

Abstract
Pregnancy-associatedPlasmodium falciparum malaria (PAM) is a major cause of morbidity and mortality in African women and their offspring. PAM is characterized by accumulation of infected erythrocytes (IEs) that adhere to chondroitin sulphate A (CSA) in the placental intervillous space. We show here that human monoclonal IgG antibodies with specificity for variant surface antigens (VSA) specifically expressed by CSA-adhering IEs (VSAPAM) can be used in vitro to select parasites from nonpregnant donors to express VSAPAM and that this selection for VSAPAM expression results in preferential transcription of var2csa. The results corroborate current efforts to develop PAM-specific vaccines based on VAR2CSA.
Keywords: human monoclonal IgG, PfEMP1, Plasmodium falciparum, pregnancy, VAR2CSA
INTRODUCTION
Children living in areas of stable Plasmodium falciparum transmission acquire substantial protective immunity against malaria during the first decade of life. Protection is mediated to a large extent by variant surface antigen (VSA)-specific IgG. Nevertheless, women in such areas remain highly susceptible to P. falciparum infection if they become pregnant, as the parasites can switch to expression of particular VSA (called VSAPAM), which allow infected erythrocyte (IE) sequestration in the placenta, but which are not compatible with parasite survival in a nonpregnant host. Acquired immunity to PAM is mediated by VSAPAM-specific IgG that either opsonizes IEs for phagocytosis or interferes with chondroitin sulphate A (CSA)-specific adhesion of IEs (1). VAR2CSA, which is a member of the P. falciparum erythrocyte membrane protein 1 (PfEMP1) family of VSA, appears to be the dominant or only pregnancy-associated Plasmodium falciparum malaria (PAM) type VSA in the P. falciparum genome, and is therefore the main target of current efforts to develop a vaccine against PAM (2). To assist this work, we have developed a panel of VSAPAM-specific human IgG1 monoclonal antibodies (3), which can opsonize VSAPAM-expressing IEs for phagocytosis and interfere with IE adhesion to CSA (Barfod et al. unpublished data). These antibodies can be used to enrich for VSAPAM expression in parasites not expressing VSAPAM but obtained from women with PAM (3). In the present study we used two of the above-mentioned monoclonal antibodies to select for VSAPAM expression in parasites from nonpregnant donors. One antibody (PAM1·4) was chosen because it appears to react with most or all parasites expressing VSAPAM. The other antibody (PAM8·1) was chosen because it reacts with a well-defined, but inter-clonally variant, epitope in the DBL3X domain of VAR2CSA.
MATERIALS AND METHODS
Monoclonal antibodies
We used eight VSAPAM-specific monoclonal IgG1 antibodies generated as described elsewhere (3; 4). VSAPAM is defined here as IE surface-expressed VSA, which are significantly better recognized by plasma IgG from P. falciparum-exposed multigravidae than from sympatric men, and where the recognition by plasma IgG from these men is not significantly different from recognition by plasma IgG from nonexposed controls. In contrast, typical non-PAM VSAs are better recognized by plasma IgG from P. falciparum-exposed adults than from nonexposed donors, without marked sex-dependent differences. Seven of the monoclonal antibodies used here are specific for inter-clonally variant epitopes in either the DBL3-X or the DBL5-ɛ domain of VAR2CSA (3). The exact specificity of the last antibody (PAM1·4) remains undefined, but it appears to recognize a conformational, and possibly discontinuous, epitope in VAR2CSA that is difficult to reproduce in recombinant constructs (3). PAM1·4 recognizes VSAPAM expressed by most or all P. falciparum genotypes, whereas the VAR2CSA DBL3-X epitope recognized by PAM8·1 is present in some, but not all P. falciparum clones (3). Both PAM1·4 and PAM8·1 can opsonize VSAPAM-expressing IEs for phagocytosis and interfere with their adhesion to CSA (Barfod et al. in preparation).
Malaria parasites
We used the two long-term in vitro-adapted parasites 3D7 and HB3. The 3D7 clone was originally derived from NF54 parasites isolated from a Dutch girl near Amsterdam airport (5). It was chosen here because it can be selected for expression of VSAPAM that react with PAM1·4 but not PAM8·1 (3). HB3 was cloned from the Honduras I/CDC parasite strain (6), and was chosen because it can be selected for expression of VSAPAM reactive with PAM1·4 and PAM8·1. All parasites were grown in 0+ erythrocytes as described (3).
Antibody selection for VSAPAM expression
Selection for VSAPAM expression was done essentially as described elsewhere (7). In brief, monoclonal PAM1·4 or PAM8·1 antibodies were immobilized on Protein A-coated magnetic beads (Dynal) and mixed with 3D7- or HB3-IEs. IEs adhering to the antibody-coated beads were isolated in a strong magnetic field and subsequently returned to in vitro culture. Selection protocol was repeated when multiplication of the antibody-selected parasites allowed it. VSA expression was assessed by flow cytometry analysis as described elsewhere (8). We used plasma from 10 P. falciparum-exposed multigravidae, 10 sympatric men, and 10 nonexposed controls to assess the sex-specificity of VSA recognition. According to the original criteria (9), sex-specific recognition requires that (i) levels of IE-surface reactive IgG are significantly higher in P. falciparum-exposed multigravidae than in sympatric men, and that (ii) the difference in IgG levels in P. falciparum-exposed men and nonexposed controls is not statistically significant. We used term plasma from 30 P. falciparum-exposed women (10 pregnant for the first time, 10 for the third time, and 10 for the fifth time) to assess parity-dependency, which requires a statistically significant relationship between IgG levels and number of pregnancies in P. falciparum-exposed women (9). Parasite isolates were only considered to express VSAPAM if plasma IgG recognition of IEs was both sex-specific and parity-dependent.
Analysis of var gene transcription
Late-stage IEs were isolated by magnetic separation as described (8) and returned to culture overnight to obtain ring-stage IEs. Genomic DNA was isolated with a QIAamp blood kit (Qiagen), and total RNA extracted (TRIzol, Invitrogen) and treated with DNase I (Invitrogen) for 30 min (10). The absence of DNA in RNA samples was confirmed as described (11). Reverse transcription was performed using Superscript II (Invitrogen) and random hexamer primers, followed by real-time PCR to quantify var transcript abundances as described (11). We used primer pairs specific for the 59 vargenes in the 3D7 genome (12) and for 45 var genes in the HB3 genome (Table S1 in Supporting Information). The latter were tested on genomic DNA dilutions to ascertain appropriate fragment size, melting temperature, and amplification efficiency compared to the internal control seryl-tRNA synthetase. All primer pairs varied less than two Ct values from that of the internal control and amplified single fragments of the expected sizes.
Statistical analysis
Differences in antibody levels among plasma from P. falciparum-exposed multigravidae, P. falciparum-exposed men, and nonexposed control donors were analysed by one-way anova, or by one-way analysis of ranks for non-normal distributed data. If statistically significant (P < 0·05) overall differences were detected, significant pair-wise differences were isolated by Holm-Sidak's or Dunn's test for normal- and non-normal-distributed data, respectively. Parity-dependency was analysed by Pearson product moment correlation.
RESULTS
Human monoclonal IgG can select Plasmodium falciparum-IEs for expression of VSAPAM
The VSA expressed on the surface of unselected 3D7- and HB3-IEs were significantly better recognized by plasma IgG from P. falciparum-exposed donors of both sexes than by IgG from nonexposed donors, whereas levels in exposed men and multigravidae were not significantly different from each other. Furthermore, IE surface-reactive IgG levels did not depend on parity (Table 1 and Figure 1a). Thus, unselected 3D7 and HB3 both showed typical non-PAM-type plasma antibody recognition patterns of IE surface-expressed VSA. This was confirmed by the nonreactivity of 3D7- and HB3-IEs with all eight VSAPAM-specific monoclonal antibodies (Figure 1d). When VSA expression was re-assessed after three rounds of selection (about 6 weeks after the first round of selection), the recognition of 3D7- and HB3-IEs selected on either PAM1·4 or PAM8·1 all showed an indeterminate VSA phenotype, where one but not both criteria for sex-specific antibody recognition were met (Table 1 and Figure 1b). PAM8·1-selected 3D7 remained nonreactive with the four monoclonal antibodies used for testing at this time (PAM1·4, PAM3·10, PAM4·7, and PAM8·1), whereas the other three parasite lines showed reactivity with at least one of them (Figure 1e). These results suggested that further rounds of selection might lead to definite VSAPAM expression, at least for PAM1·4-selected 3D7, PAM1·4- and PAM8·1-selected HB3. Indeed, PAM1·4-selected 3D7, as well as PAM1·4- and PAM8·1-selected HB3 had all acquired a typical sex-specific and parity dependent VSAPAM expression pattern after four additional rounds of selection (Table 1 and Figure 1c), and reacted with all the monoclonal antibodies except the VAR2CSA DBL5-ɛ-specific PAM4·7 (Figure 1f). In contrast, 3D7 selected seven times for reactivity with PAM8·1 retained an indeterminate sex-specificity pattern also seen after three rounds of selection, did not acquire the parity-dependent pattern typical of VSAPAM-expressing lines (Table 1), and did not react with any of the eight VSAPAM-specific monoclonal antibodies (Figure 1f). Thus, PAM8·1 could not be used to select 3D7 for VSAPAM reactivity, consistent with the absence of the predicted epitope for this antibody in the VAR2CSA DBL3-X domain of this parasite (3).
Table 1.
Changes in plasma antibody recognition pattern of infected erythrocytes following selection with VSAPAM-reactive human monoclonal antibodies
| Rounds of selection for antibody recognition |
|||||||
|---|---|---|---|---|---|---|---|
| Parasite | Antibodya | 0 | 3 | 7 | |||
| Sex-specificityb | Parity-dependency | Sex-specificity | Parity-dependency | Sex-specificity | Parity-dependency | ||
| 3D7 | PAM1·4 | No | No: P(r) = 0·87 | Indeterminate | n.d. | Yes | Yes: P(r) = 0·002 |
| PAM8·1 | Indeterminate | n.d. | Indeterminate | No: P(r) = 0·41 | |||
| HB3 | PAM1·4 | No | No: P(r) = 0·55 | Indeterminate | n.d. | Yes | Yes: P(r) = 0·006 |
| PAM8·1 | Indeterminate | n.d. | Yes | Yes: P(r) = 0·037 | |||
Human monoclonal VSAPAM-specific antibody used for selection.
Determination of sex-specificity involves a two-step procedure: first establishment that IE surface-reactive IgG levels in at least one of the three donor categories (exposed multigravidae, exposed men, and unexposed controls) differs from at least one other category. Only if this is the case (as it was here: P < 0·001 in all cases), can sex-specificity be confidently assessed by post hoc testing for significant (P < 0·05) pair-wise differences. The result of this post hoc testing is indicated here as No: none of the two post hoc criteria for sex-specificity were met, Indeterminate: only one of the criteria was met. Yes: both criteria were met. See Materials and Methods for further details.
Figure 1.
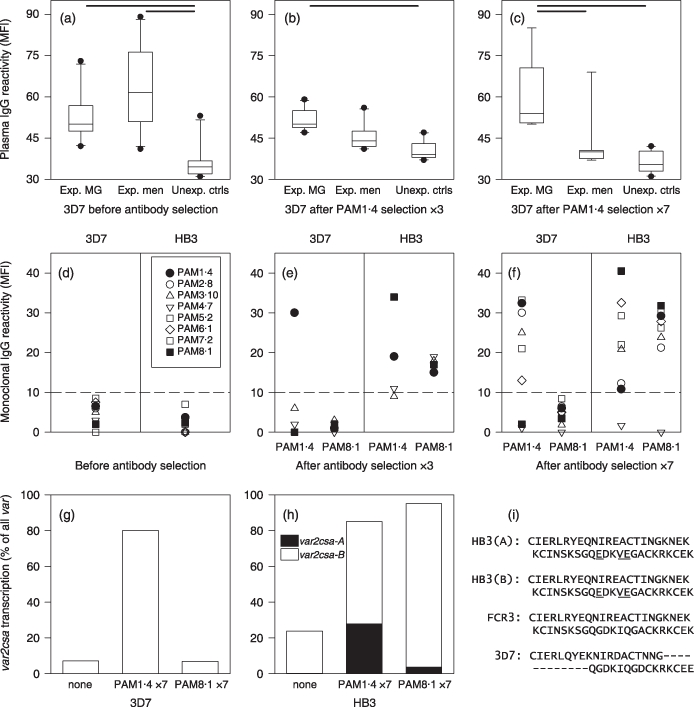
Changes in VSA expression andvar gene transcription following selection for expression of VSAPAM reactive with human monoclonal antibodies.Recognition of IEs by plasma IgG from 10 P. falciparum-exposed multigravidae (Exp. MG), 10 sympatric men (Ex. men) and 10 nonexposed controls (Unexp. ctrls) before selection (a), and after three (b) or seven (c) rounds of selection by human monoclonal IgG antibody PAM1·4. Results in panels A-C are presented as medians (horizontal line), central 50% of data points (boxes), central 80% of data points (whiskers) and outliers (•). In addition, statistically significant (P < 0·05) pair-wise differences are indicated by heavy horizontal bars along the top of the panels. Recognition of IEs by human monoclonal VSAPAM-specific IgG antibodies before selection (d), and after three (e) or seven (f) rounds of selection. Results in panels D–F are presented as individual data points. Negative cut-off, defined as the upper level of recognition of unselected parasites, is indicated as a dashed horizontal line. The proportion of var2csa transcripts among all var transcripts in 3D7 (g) and HB3 (h) before selection and after seven rounds of selection. Panel (i) shows the amino acid sequence of the PAM8·1-specific region of VAR2CSA DBL3-X in the two VAR2CSA paralogs in HB3, FCR3, and 3D7 parasites. Amino acid differences between the two HB3 sequences and the PAM8·1-reactive FCR3 sequence are underlined.
Acquisition of VSAPAM expression following antibody-selection is associated with selective transcription of var2csa
Transcripts of var2csa (PFL0030c) constituted 7% of total measured var gene transcripts in unselected 3D7 parasites, increasing to 80% after seven rounds of PAM1·4 selection. In contrast, PAM8·1 selection did not affect the proportion of var2csa transcripts in 3D7 (Figure 1g). The HB3 genome contains two var2csa paralogs, var2csa-A and var2csa-B (13). In unselected HB3 parasites, var2csa-A transcripts constituted 1% of the measured var transcripts, increasing to 28% and 4% after seven rounds of selection by PAM1·4 and PAM8·1, respectively (Figure 1h). Transcript levels of the var2csa-B gene increased from 23% to 57% and 92% after seven rounds of selection by PAM1·4 and PAM8·1, respectively (Figure 1h). No var transcript other than var2csa showed marked changes, and var2csa was therefore the dominant transcript in all the parasites following antibody selection. The dominance of var2csa-B transcripts relative to var2csa-A in HB3 after PAM8·1 selection, suggested that the DBL3-X domain in the protein encoded by var2csa-B might be of the FCR3-type recognized by PAM8·1, whereas the corresponding domain encoded by var2csa-A might be of the 3D7-type (PFL0030 c) not recognized by PAM8·1 (2). However, var2csa-A and var2csa-B encode an identical amino acid sequence in the region spanning the PAM8·1 epitope, and this sequence was of the FCR3-type (Figure 1i). The var2csa-A : var2csa-B transcripts may therefore reflect a founder effect.
DISCUSSION
The VSA subset VSAPAM is expressed by P. falciparum involved in pregnancy-associated malaria (PAM). VSAPAM-specific IgG mediates acquired immunological protection against PAM, and the PfEMP1 variant VAR2CSA appears to be the main or only target of these antibodies. VAR2CSA is therefore the leading candidate for development of vaccines against PAM.
We used monoclonal human IgG antibodies to select erythrocytes infected by two genotypically distinct laboratory P. falciparum clones derived from nonpregnant donors for expression of VSAPAM. Parasites acquiring expression of VSAPAM following selection showed increased levels of transcripts encoding the PfEMP1 variant VAR2CSA, which appears to be the only PAM-type VSA in the P. falciparum genome. The results obtained in the study are important for several reasons.
First they support the hypothesis that all P. falciparum parasites have the capacity to express VSAPAM. This hypothesis is supported by previously published data that all P. falciparum genomes appear to contain at least one paralog of the gene encoding the only known VSAPAM-type antigen, VAR2CSA (11,14). Furthermore, this gene is selectively transcribed by placental parasites (and following selection for adhesion to CSA in vitro) and VAR2CSA is expressed on the IE surface (15–17).
Second, they indicate that P. falciparum parasites regularly and spontaneously switch to expression of VSAPAM in the absence of an external signal, for example pregnancy-associated hormonal changes. It has been speculated that switching to VSAPAM expression requires signals from the pregnant host, for example hormones, and that selection therefore might not be possible unless the parasite is derived from such a host. It has also been argued that selection of parasites by panning on CSA in vitro might result in expression of antigens of dubious relevance to the antigens expressed as a result of in vivo selection occurring in the pregnant woman. Our data show that switching to expression of genuine VSAPAM can occur in vitro in the absence of external signals, in line with other recent evidence (18). By extension, these findings suggest that switching to VSAPAM in vivo also occurs spontaneously regardless of the pregnancy status of the host. In a pregnant host, such parasites will often be at a selective advantage (because of the frequent absence of VSAPAM-specific immunity in women of low parity) (19), whereas they appear to be unable to survive in a nonpregnant host (20).
Third, our results support the hypothesis that although interclonal variation in VAR2CSA is the result of antibody-driven positive selection, the diversity of functionally important antibody epitopes in the molecule is constrained (21). Thus, some parasites can express VAR2CSA without being vulnerable to recognition by certain VAR2CSA-reactive antibodies, because of variation in defined parts of VAR2CSA. The persistent nonrecognition by PAM8·1 of VAR2CSA-expressing 3D7 is a case in point. At the same time, the PAM8·1 antibody, originally identified by its reactivity with VSAPAM-expressing FCR3-IEs (3), was effectively selected for VSAPAM expression in the genetically unrelated HB3 clone. The significance of these findings is underscored by the fact that PAM8·1 efficiently opsonizes IEs for phagocytosis as well as interferes with IE adhesion to CSA (Barfod et al. in preparation). Our study also corroborates existing evidence that PAM1·4 recognizes most if not all VSAPAM-expressing parasites by recognizing a functionally highly constrained and conformation-dependent discontinuous epitope in VAR2CSA (3). Like PAM8·1, PAM1·4 is an opsonin and capable of interfering with IE adhesion to CSA (Barfod et al. in preparation). Therefore, the PAM1·4 epitope is a highly attractive candidate for development of a PAM-specific vaccine, and its characterization is a matter of the highest priority in our current research. Although the theoretical possibility of another, non-PfEMP1 target of PAM1·4 remains, available evidence point to VAR2CSA.
In conclusion, our evidence support current efforts to develop PAM-specific vaccines based on VAR2CSA and highlight the versatility of human monoclonal antibodies generated from clinically immune donors in these investigations.
Acknowledgments
The study received financial support from the Commission of the European Communities (grant LHSP-CT-2006-036838 HUMALMAB), Seattle Biomedical Research Institute, Seattle, WA (sub-contracts MADP/PMI-3035 and UC-3036), and Copenhagen University Hospital Rigshospitalet (410-005). The Ph.D. study program of JS is partially financed by grants from University of Copenhagen and the Danish Technical University.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Table S1 Real-time PCR primers for HB3 var genes
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
REFERENCES
- 1.Hviid L. The immuno-epidemiology of pregnancy-associated malaria: a variant surface antigen-specific perspective. Parasite Immunol. 2004;26:477–486. doi: 10.1111/j.0141-9838.2004.00733.x. [DOI] [PubMed] [Google Scholar]
- 2.Hviid L, Salanti A. VAR2CSA and protective immunity against pregnancy-associated Plasmodium falciparum malaria. Parasitology. 2007;134:1871–1876. doi: 10.1017/S0031182007000121. [DOI] [PubMed] [Google Scholar]
- 3.Barfod L, Bernasconi N, Dahlbäck M, et al. Human pregnancy-associated malaria-specific B cells target polymorphic, conformational epitopes in VAR2CSA. Mol Microbiol. 2007;63:335–347. doi: 10.1111/j.1365-2958.2006.05503.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Traggiai E, Becker S, Subbarao K, et al. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nat Med. 2004;10:871–875. doi: 10.1038/nm1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Delemarre BJ, Van der Kaay HJ. Malaria tropica op natuurlijke wijze verkregen in Nederland. Ned Tijdschr Geneeskd. 1979;123:1981–1982. [PubMed] [Google Scholar]
- 6.Bhasin VK, Trager W. Gametocyte-forming and non-gametocyte-forming clones of Plasmodium falciparum. Am J Trop Med Hyg. 1984;33:534–537. doi: 10.4269/ajtmh.1984.33.534. [DOI] [PubMed] [Google Scholar]
- 7.Staalsoe T, Nielsen MA, Vestergaard LS, et al. In vitro selection of Plasmodium falciparum 3D7 for expression of variant surface antigens associated with severe malaria in African children. Parasite Immunol. 2003;25:421–427. doi: 10.1111/j.1365-3024.2003.00652.x. [DOI] [PubMed] [Google Scholar]
- 8.Staalsoe T, Giha HA, Dodoo D, et al. Detection of antibodies to variant antigens on Plasmodium falciparum infected erythrocytes by flow cytometry. Cytometry. 1999;35:329–336. doi: 10.1002/(sici)1097-0320(19990401)35:4<329::aid-cyto5>3.3.co;2-p. [DOI] [PubMed] [Google Scholar]
- 9.Ricke CH, Staalsoe T, Koram K, et al. Plasma antibodies from malaria-exposed pregnant women recognize variant surface antigens on Plasmodium falciparum-infected erythrocytes in a parity-dependent manner and block parasite adhesion to chondroitin sulphate A. J Immunol. 2000;165:3309–3316. doi: 10.4049/jimmunol.165.6.3309. [DOI] [PubMed] [Google Scholar]
- 10.Rottmann M, Lavstsen T, Mugasa JP, et al. Differential expression of var gene groups is associated with morbidity caused by Plasmodium falciparum infection in Tanzanian children. Infect Immun. 2006;74:3904–3911. doi: 10.1128/IAI.02073-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Salanti A, Staalsoe T, Lavstsen T, et al. Selective upregulation of a single distinctly structured var gene in CSA-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- 12.Dahlbäck M, Lavstsen T, Salanti A, et al. Changes in vargene mRNA levels during erythrocytic development in two phenotypically distinct Plasmodium falciparum parasites. Malaria J. 2007;6:78. doi: 10.1186/1475-2875-6-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kraemer SM, Kyes SA, Aggarwal G, et al. Patterns of gene recombination shape vargene repertoires in Plasmodium falciparum: comparisons of geographically diverse isolates. BMC Genomics. 2007;8:45. doi: 10.1186/1471-2164-8-45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bockhorst J, Lu F, Janes JH, et al. Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol. 2007;155:103–112. doi: 10.1016/j.molbiopara.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 15.Magistrado P, Salanti A, Tuikue Ndam NG, et al. VAR2CSA expression on the surface of placenta-derived Plasmodium falciparum-infected erythrocytes. J Infect Dis. 2008;198:1071–1074. doi: 10.1086/591502. [DOI] [PubMed] [Google Scholar]
- 16.Salanti A, Dahlbäck M, Turner L, et al. Evidence for the involvement of VAR2CSA in pregnancy-associated malaria. J Exp Med. 2004;200:1197–1203. doi: 10.1084/jem.20041579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tuikue Ndam NG, Salanti A, Bertin G, et al. High level of var2csatranscription by Plasmodium falciparum isolated from the placenta. J Infect Dis. 2005;192:331–335. doi: 10.1086/430933. [DOI] [PubMed] [Google Scholar]
- 18.Nunes MC, Sterkers Y, Gamain B, et al. Investigation of host factors possibly enhancing the emergence of the chondroitin sulfate A-binding phenotype in Plasmodium falciparum. Microbes Infect. 2008;10:828–832. doi: 10.1016/j.micinf.2008.05.006. [DOI] [PubMed] [Google Scholar]
- 19.Fried M, Nosten F, Brockman A, et al. Maternal antibodies block malaria. Nature. 1998;395:851–852. doi: 10.1038/27570. [DOI] [PubMed] [Google Scholar]
- 20.Nguyen-Dinh P, Steketee RW, Greenberg AE, et al. Rapid spontaneous postpartum clearance of Plasmodium falciparum parasitaemia in African women. Lancet. 1988;2:751–752. doi: 10.1016/s0140-6736(88)90229-2. [DOI] [PubMed] [Google Scholar]
- 21.Dahlbäck M, Rask TS, Andersen PH, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in Plasmodium falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.