

Abstract
Purpose
To determine the immunogenicity of diphtheria toxoid (DT) formulated in two types of vesicles following transcutaneous immunization (TCI) of mice onto microneedle array-treated skin.
Methods
DT-containing cationic liposomes or anionic surfactant-based vesicles were prepared by extrusion and sonication. The physicochemical properties were characterized in terms of size, ζ-potential, vesicle elasticity and antigen association. TCI was performed by applying formulations onto intact or microneedle array-pretreated mice skin, using cholera toxin as an adjuvant. Subcutaneous and intradermal immunizations were as control. Immune responses were evaluated by IgG and neutralizing antibody titers, and the immune-stimulatory properties were assessed using cultured dendritic cells.
Results
Stable DT-containing cationic liposomes (∼150 nm) and anionic vesicles (∼100 nm) were obtained. Incorporation of Span 80 increased liposome elasticity. About 90% and 77% DT was associated with liposomes and vesicles, respectively. TCI of all formulations resulted in substantial antibody titers only if microneedle pretreatment was applied. Co-administration of cholera toxin further augmented the immune responses of TCI. However, vesicle formulations didn’t enhance the immunogenicity on either intact or microneedle-treated skin and showed low stimulatory activity on dendritic cells.
Conclusions
Microneedle pretreatment and cholera toxin, but not antigen association to vesicles, enhances the immunogenicity of topically applied DT.
KEY WORDS: cholera toxin, diphtheria toxoid, microneedle array, transcutaneous immunization, vesicles
INTRODUCTION
Vaccination is a cost-effective way to fight infectious diseases. Currently, most vaccines are administered by subcutaneous (s.c.) or intramuscular injection. The problems with administration via these routes are that they are painful, cause stress (especially in children), require trained personnel and may lead to side effects. This results in incomplete vaccination coverage and, hence, unnecessary deaths from vaccine-preventable diseases. Consequently, research is focused on alternative vaccine delivery through oral (1), dermal (2), nasal (3) and pulmonary (4) routes. Vaccination via the dermal route is of particular interest since the skin is the biggest organ and is easily accessible. Antigen-presenting cells (APCs) in the skin, in particular the Langerhans cells in the epidermis and the dermal dendritic cells (DCs) in the dermis, are the targets for transcutaneous immunization (TCI), i.e. topical vaccine application onto intact or pretreated skin. Epidermal Langerhans cells usually represent about 1% of the total epidermal cell population while covering 20% of the skin surface (5). Upon contact with certain antigens together with danger signals, e.g. skin barrier disruption or inflammation, Langerhans cells and dermal DCs become activated. They capture antigens/pathogens, migrate to the peripheral draining lymph nodes, process and present the antigen to the naïve T cells and initiate immune responses (6). However, a major hurdle is to overcome the stratum corneum: the upper-most layer of the skin, which serves as a barrier. This layer consists of corneocytes embedded in a highly organized crystalline lamellar structure of the intercellular lipid matrix. The unique arrangement of this layer (15–20 μm thick in humans and thinner in mice) makes it impermeable for macromolecules, including antigens (7).
In the 1970s, Gerstel and Place proposed the use of microneedle arrays as a way to overcome this barrier in a minimally invasive manner (8). However, it was not employed in (trans)dermal drug delivery studies until about 10 years ago, when the technology for fabrication in micron dimensions became available and cost-effective (9). After the skin is treated with a microneedle array, tiny conduits are formed transiently with little pain sensation. Antigens are able to diffuse through the conduits to the APCs in the epidermis and dermis (10). It has been shown previously that microneedle array pretreatment leads to major improvement in the immunogenicity of topically applied diphtheria toxoid (DT) in mice (11). Other studies have also demonstrated the promise of microneedle array-aided TCI: Matriano et al. studied the dosing of a model antigen, ovalbumin, and the kinetics and the magnitudes of antibody titers induced in microneedle-mediated TCI in hairless guinea pigs (12); Widera et al. investigated the influences of important fabrication parameters, e.g. the needle length, needle density, and covered skin area of the ovalbumin-coated microneedle arrays, on the effectiveness of TCI (13).
Another approach to overcome the skin barrier in TCI is to formulate antigens with vesicles. Elastic vesicles, consisting of highly deformable bilayers, have been reported to be able to improve transdermal drug delivery. These vesicle formulations were applied non-occlusively as the transdermal osmotic gradient has been reported to be the driving force of the transport across the stratum corneum (14). For example, Transfersomes® have been used successfully for a range of small molecule drugs, including steroids, non-steroidal anti-inflammatory drugs and local anesthetics (15–17), as well as peptides and proteins, e.g. cyclosporin A and insulin (18,19). Moreover, TCI with hepatitis B surface antigen-loaded elastic liposomes and tetanus toxoid-loaded Transfersomes® has been shown to elicit immune responses equivalent to those produced by intramuscular injection of alum-adsorbed antigens in mouse and rat, respectively (20,21). In our group, elastic vesicles composed of surfactants have previously been developed. After non-occlusive application on the skin, vesicle structures were observed in the deeper layers of the stratum corneum close to the stratum corneum-viable epidermis junction. Enhanced delivery of lidocaine, rotigotine and ketorolac was observed when the drug was associated with vesicles and the formulation applied on the skin in a non-occlusive manner (22–24). Therefore, there is a potential for antigen-loaded vesicles as an effective formulation for TCI.
In this study, for the first time, these two approaches—microneedle pretreatment and antigen-loaded vesicle formulations—were combined to assess their potential in TCI, using DT (MW: 58 KDa; isoelectric point: 4.7) as an antigen. Two types of vesicles have been included. Cationic liposomes were made of soybean phosphatidylcholine (SPC) and 1,2-dioleoyl-3-trimethylammonium-propane chloride salt (DOTAP). Span 80 was incorporated to increase elasticity of the vesicle bilayer (25). These liposomes in the absence or presence of Span 80 are referred to as DT-Lip and DT-ELip (DT Elastic Liposome), respectively. Characterization of their physicochemical properties is reported in this study. Surfactant-based vesicles were composed of sucrose-laurate ester (L-595) and sodium bistridecyl sulfosuccinate (TR-70). Octaoxyethylene-laurate ester (PEG-8-L) was used to increase the vesicle elasticity. This DT-containing vesicle formulation has been developed and characterized previously and referred to as DT-Ves in this paper (26). Selected formulations were applied in TCI onto intact or microneedle-treated skin of mice and compared with intradermal (i.d.) and s.c. immunization. Their immunogenicity was evaluated by measuring serum IgG (subtype) and neutralizing antibody titers. Their immune-stimulatory properties were assessed in vitro using human peripheral blood mononuclear cell-derived immature DCs.
MATERIALS AND METHODS
Materials
SPC and DOTAP were kindly supplied by Lipoid GmbH (Ludwigshafen, Germany). Diphtheria toxin (batch 79/1), DT (batch 98/40, protein content 12.6 mg/ml by BCA assay, 1 μg equals to approximately 0.3 Lf), horse anti-DT and horseradish peroxidase (HRP) conjugated anti-DT were provided by the Netherlands Vaccine Institute (NVI, Bilthoven, the Netherlands). HRP-conjugated goat anti-mouse (HRP-GAM) IgG (γ-chain specific), IgG1 (γ1-chain specific) and IgG2a (γ2a-chain specific) were purchased from Southern Biotech (Birmingham, US). Adju-Phos® (alum) was obtained from Brenntag Biosector (Copenhagen, Denmark). Chromogen 3, 3′, 5, 5′-tetramethylbenzidine (TMB) and the substrate buffer were purchased from Biosource B.V. (Nivelles, Belgium). Tween 20, lyophilized bovine serum albumin, Folin Ciocalteu’s phenol reagent, cholera toxin and Span 80 were ordered from Sigma-Aldrich (Zwijndrecht, the Netherlands). Tween 80 was purchased from Merck (Darmstadt, Germany). Ficoll and Percoll were ordered from GE Healthcare (Eindhoven, the Netherlands). Nimatek® (100 mg/ml ketamine), Rompun® (20 mg/ml xylasine) and the injection fluid (0.9% NaCl) were obtained from a local pharmacy. All other chemicals used were of analytical grade, and all solutions were prepared with distilled water.
Methods
DT Vesicle Formulation Preparation
The compositions of the DT vesicle formulations are listed in Table I. The DT-Lip and DT-ELip were prepared using the film rehydration and extrusion method. SPC, Span 80 and DOTAP, dissolved in chloroform, were mixed in an appropriate ratio and formed a thin film at the bottom of the flask using a rotary evaporator. Residual organic solvent in the film was removed by 30 min nitrogen flow. The film was rehydrated by 10 mM phosphate buffer (PB, pH 7.4, 7.7 mM Na2HPO4 and 2.3 mM NaH2PO4) or 10 mM citrate buffer (CB, pH 5.0, 4.0 mM H3C6H5O7 and 6.0 mM Na3C6H5O7) with or without saline (153 mM NaCl, PBS or CBS) containing 1.5 mg/ml DT. The concentration of lipids in the buffer was 5% w/w. The resulting dispersions were shaken at 200 rpm in the presence of glass beads for 2 h at room temperature, followed by 2 × 15 s tip sonication (20% energy output, Branson Ultrasonics, Danbury, UK). The formulations were extruded (LIPEX™ Extruder, Northern Lipids Inc., Canada) 3 times through a pair of polycarbonate filters with pore size of 200 nm and 3 times through another pair with pore size of 100 nm (Millipore, Amsterdam, the Netherlands). DT-Ves were prepared and characterized in a previous study (26). Briefly, the surfactants were dissolved in ethanol, while TR-70 was dissolved in an ethanol/isopropanol mixture. After mixing in an appropriate ratio, the organic solvents were evaporated overnight in a vacuum centrifuge and rehydrated with 10 mM CB (pH 5.0) containing 1.5 mg/ml DT. Vesicles were formed after 3 × 5 s tip sonication (20% energy output, Branson Ultrasonics). Subsequently, the pH of the buffer was lowered to 4.5 by addition of 0.1 M HCl in order to associate DT with the vesicles. The final concentration of the surfactants in buffer was 10% w/w.
Table I.
DT-Loaded Vesicle Formulations
| Buffer (pH) | Components and molar ratio | Abbreviated name |
|---|---|---|
| L-595:PEG-8-L:TR-70 | ||
| CB (4.5) | 5:5:1 | DT-Ves |
| SPC:Span 80:DOTAP | ||
| PBS (7.4) | 9:0:1 | DT-Lip-PBS |
| PBS (7.4) | 9:3:1 | DT-ELip-PBS |
| PB (7.4) | 9:0:1 | DT-Lip |
| PB (7.4) | 9:3:1 | DT-ELip |
| CBS (5.0) | 9:0:1 | DT-Lip-CBS |
| CBS (5.0) | 9:3:1 | DT-ELip-CBS |
| CB (5.0) | 9:0:1 | DT-Lip-5 |
| CB (5.0) | 9:3:1 | DT-ELip-5 |
Size and ζ-Potential Measurements
The size and polydispersity index (PDI) of all formulations were measured by dynamic light scattering using a Zetasizer (Nano ZS, Malvern Ltd., United Kingdom). All size measurements were performed at 25 °C at an angle of 173° between the laser and the detector. Before measuring, all formulations were diluted in their corresponding buffer. Formulations were measured at regular time points during a period of 2 weeks after preparation to evaluate the colloidal stability. The ζ-potential was measured by laser Doppler electrophoresis using the same device. The formulations were stored at 4 °C in between these measurements.
DT-Vesicle Association
To determine the DT-vesicle association ratios in the formulations, size exclusion chromatography was performed using a Sepharose® CL-4B (Amersham, Uppsala, Sweden) column as previously described (26). Plain vesicles (cationic liposomes or anionic surfactant vesicles) and free DT in buffer solution served as controls to determine the time window for fraction collection. Each formulation was run with its corresponding buffer as mobile phase at a flow speed of 0.40 ml/min. The recovery rate of DT after SEC was determined by a modified Lowry-Peterson protein assay, which separates the antigen from the lipids/surfactants through a precipitation step and measures total protein content in each fraction (27). The antigenicity of DT in each fraction was measured by ELISA, using horse anti-DT serum and a HRP-conjugated horse anti-DT, which measures the antigenicity of accessible DT in the presence of the vesicle components. Data were analyzed using the principle of parallel-line analysis of the OD-log concentration plots as described by Tierney et al. (28).
To examine the status of DT-liposome association when encountering physiological condition, the formulations were eluted in SEC using PBS. The resulting liposome-associated-DT fractions were further treated by 1% Triton X-100 to disrupt the liposome structure and release the encapsulated DT. The antigenicity of DT in these fractions was determined by ELISA and compared to the corresponding fractions eluted by the original buffers.
Microneedle Array and Applicator
The microneedle array used in this study was manufactured from commercially available 30G hypodermic needles (Becton Dickinson, Alphen a/d Rijn, the Netherlands) as described previously (29). The needles were assembled as a 300 µm-long, 4 × 4 array on a polymer back plate with a surface area of about 0.5 cm2. The microneedles were inserted into the skin at a speed of 3 m/s using an electric impact applicator (11).
Evaluation of Vesicle Elasticity
The elasticity of the DT-loaded vesicles was evaluated by extrusion through a modified Avanti Mini-Extruder® (Avanti Polar Lipids Inc., Delfzyl, the Netherlands). This method evaluates vesicle elasticity by comparing the extruded volume of dispersions under constant pressure as a function of time. In detail, the Mini-Extruder® was mounted on a scaffold with both syringes standing perpendicularly, donor above receptor. A polycarbonate filter with pore size of 30 nm was inserted in between the donor and acceptor syringes, and 1 ml of vesicle dispersion was loaded in the donor syringe. A 1-kg weight was mounted to generate a constant pressure of about 10 bar on the piston of the donor syringe. Upon release of the weight, the extruded volume of the dispersion in the receptor syringe was recorded every minute over a 10-minute period. This procedure was repeated two more times using freshly prepared dispersions. The elasticity of vesicle membrane was calculated using the following formula as reported by Van den Bergh et al. (30):
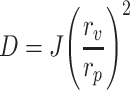 |
where, D, elasticity index of vesicle membrane; J, volume of dispersion extruded over 5 min; r v, size of vesicles; and r p, pore size of the filter.
Immunization Protocol
Female BALB/c mice (H2d), 8 weeks old at the start of the experiments, were purchased from Charles River (Maastricht, the Netherlands) and maintained under standardized conditions in the animal facility of the Leiden/Amsterdam Center for Drug Research, Leiden University. The study was conducted in conformity with the Public Health Service Policy on use of laboratory animals and had been approved by the Research Ethical Committee of Leiden University.
Three immunization studies were carried out. Negatively charged DT-Ves and positively charged DT liposome formulations were applied on intact or microneedle-treated skin in the first and second study, respectively. Formulations were applied occlusively or non-occlusively, with the presence or absence of CT as an adjuvant. Immunization of DT-Lip via i.d. injection was carried out in the third study.
One day before immunization, the abdominal skin of mice of TCI groups was shaved and rested for 24 h. During immunization, mice were anesthetized by intraperitoneal injection of 150 mg/kg ketamine and 10 mg/kg xylazine. The shaved skin area was wiped with 70% ethanol. For microneedle treatment, a skin fold was supported by styrofoam and pierced using the microneedle array right before the application of DT formulations. Seventy-μl DT formulations containing 100 µg DT (and 100 µg CT if applicable) per mouse were applied onto intact or treated skin. The formulations were carefully spread over the entire skin area of application, about 2 cm2 restricted by a metal ring. After 1 h occlusive or non-occlusive incubation, the skin area was extensively washed with lukewarm tap water and patted dry twice. In the third study, 30 µl DT-Lip or DT solution containing 5 µg DT were given intradermally. Five µg of DT adsorbed onto alum (DT-alum) in a 100 μl saline suspension was administered by s.c. injection for all three studies as a control. The DT-alum formulation was prepared as previously described, and the adsorption of DT to alum was between 70% and 80% (31). All DT formulations were freshly prepared before each vaccination. Mice were immunized at day 1, 21 and 42 (at approximately the same skin region for the TCI groups) and sacrificed at day 56. Blood was sampled from the tail vein one day before each immunization, and whole blood was collected from the femoral artery during sacrifice. Cell-free sera were obtained using MiniCollect® tubes (Greiner Bio-one, Alphen a/d Rijn, the Netherlands) by centrifugation after clot formation and stored at −80 °C until use.
Serum Antibody Assays
Serum IgG, IgG1 and IgG2a titers were determined by ELISA. Briefly, ELISA plates (Microlon®, Greiner Bio-one) were coated with DT at 4°C overnight. Two-fold serial dilutions of serum samples were applied in the plates, and the containing DT-specific antibodies were detected by HRP-GAM IgG, IgG1 or IgG2a using TMB as substrate. Antibody titers are expressed as the reciprocal of the calculated sample dilution corresponding to half of the maximum absorbance at 450 nm of a complete s-shaped absorbance-log dilution curve. Mice with serum sample unable to reach the half-saturated absorbance value at the lowest (ten-fold) dilution were considered as non-responders.
Protective immunity against diphtheria depends on the presence of circulating neutralizing antibodies, evaluated using Vero cell test: the WHO standard method to assess the success of diphtheria vaccination, which relies on the inhibition of a cytotoxic dose of diphtheria toxin (32). In brief, after complement inactivation, two-fold serial dilutions of serum samples were prepared with complete medium 199 (Gibco, Breda, the Netherlands) and applied to microtiter plates (CELLSTAR®, Greiner Bio-one). Subsequently, 2.5 × 10−5 Lf diphtheria toxin was added to each well. After 2 h incubation at 37°C for neutralization, Vero cell suspension was added to each well. Covered with a plate sealer, Vero cells were incubated at 37°C in 5% CO2 for 6 days. The end point was taken as the highest dilution protecting the Vero cells.
DC Maturation Study
Immature DCs were prepared from human peripheral blood mononuclear cells, which were isolated by Ficoll-Percoll density-gradient centrifugation on day 0. Monocytes were separated from platelets due to their adherence to the plastic surface (24 well plate for cell culture, Costar®, Corning, New York, US) after incubation at 37°C for 60 min. The adherent cells were cultured for 6 days in RPMI 1640, supplemented with 10% v/v fetal calf serum (FCS, Biosource-Invitrogen, Breda, the Netherlands), 1% glutamine, 100 U/ml penicillin and 0.1 mg/ml of streptomycin, 250 U/ml granulocyte-macrophage colony-stimulating factor (GM-CSF, Biosource-Invitrogen) and 100 U/ml interleukin-4 (IL-4, Biosource-Invitrogen) at 37 °C with 5% CO2 to differentiate into immature DCs. Medium was refreshed at day 3. At day 6, the medium was replaced by new medium containing GM-CSF and 2 μg/ml DT, either free, mixed with CT or associated in liposomes or vesicles, using lipopolysaccharide (LPS, from Escherichia coli, Sigma) as a positive control. Then DCs were incubated for 48 h at 37°C. Subsequently, DCs were washed 3 times with PBS containing 1% w/v BSA and 2% v/v FCS and incubated for 30 min with a mixture of 20× diluted anti-HLADR-FITC, anti-CD83-PE and anti-CD86-APC (Becton Dickinson) on ice. Cells were washed again, and the expression of MHC II, CD83 and CD86 was quantified using flow cytometry (FACS Canto II, Becton Dickinson). The up-regulation of these three surface markers by 50 ng/ml LPS was set as 100%. Live cells were gated based on forward and side scatter. A minimum of 10,000 DC events were analyzed in each experiment. The study was repeated using DCs from at least three different donors.
Statistical Analysis
IgG (subtype) antibody titers were analyzed with two-way ANOVA with Bonferroni post-test, and the neutralizing antibody titers were analyzed using one-way ANOVA with the same post-test. Other analyses were performed where suitable as indicated. Statistical analysis was carried out using Prism Graphpad, and a p value less than 0.05 was considered to be significant.
RESULTS
Colloidal Properties of DT Vesicle Formulations
Particle size and ζ-potential of the DT vesicle formulations are provided in Table II. Particle size and ζ-potential measured at day 7 and day 14 were very similar to the original values (data not shown), indicating good colloidal stability for all formulations listed. Particle sizes of the liposome formulations without Span 80 were smaller than those in the presence of Span 80. Formulations prepared by extrusion method show lower PDI than those prepared by sonication method.
Table II.
Characterization of DT Vesicle Formulations. Data Shown are Mean ± SD of Three Different Batches.
| Abbreviated name | Size (nm) | PDI | ζ-potential (mV) | Association %a (Lowry-Peterson) | Encapsulation %b (ELISA) in PBS |
|---|---|---|---|---|---|
| DT-Ves | 101 ± 6 | 0.28 ± 0.03 | −77.3 ± 3.0 | 76.8 ± 2.9 | – |
| DT-Lip | 164 ± 2 | 0.23 ± 0.02 | 24.5 ± 1.4 | 86.7 ± 3.9 | 36.0 ± 4.5c |
| DT-ELip | 175 ± 3 | 0.21 ± 0.03 | 24.4 ± 0.6 | 86.7 ± 5.1 | 46.9 ± 4.3d |
| DT-Lip-5 | 124 ± 9 | 0.12 ± 0.02 | 24.6 ± 2.0 | 29.9 ± 4.0 | 14.4 ± 4.0 |
| DT-ELip-5 | 149 ± 3 | 0.11 ± 0.01 | 24.8 ± 1.1 | 40.0 ± 4.3 | 13.0 ± 3.3 |
aAssociated DT: includes the surface-attached DT, bilayer-intercalated DT and the liposome-encapsulated DT
bEncapsulated DT: DT released upon Triton x-100 treatment, detected by ELISA
cSignificantly higher than those of DT-Lip-5 (p < 0.001, t-test)
dSignificantly higher than those of DT-ELip-5 (p < 0.001)
Association of DT with the Vesicles
The DT-Ves was characterized previously (Table II). The majority of DT was associated with vesicles when the pH of the buffer was lowered to 4.5 and dissociated when pH was increased to 7.4. The association and dissociation were not influenced by the ion strength of the buffer.
For the DT-containing cationic liposomes, when prepared in PB at pH 7.4 and CB at pH 5.0, high DT-liposome association was observed. After SEC of DT-Lip and DT-ELip, nearly all protein content was recovered from the liposome-associated DT fractions, whereas for DT-Lip-5 and DT-ELip-5, the recovery from the liposome-associated DT fraction was about 30% (Fig. 1a). However, when prepared in PBS and CBS instead, the liposome-associated DT in all four formulations appeared to be lower than 15% (data not shown). This indicates that ionic strength and pH of the buffer play a prominent role in the antigen-liposome association. Formulations prepared in PB and CB were selected for further studies due to higher DT-liposome association.
Fig. 1.

DT-liposome association in formulations prepared in CB or PB. The liposome-associated DT and free DT were separated through SEC fractionation of formulations by their corresponding buffers and determined with Lowry-Peterson protein assay (a) and ELISA (b); free DT in PB and CB served as control. DT liposome formulations prepared in CB and PB were eluted in SEC with PBS (referred to as “eluted by PBS,” c and d). The resulted liposome-associated DT fractions were further treated with 1% Triton X-100 to release the encapsulated DT, and both were measured with ELISA (referred to as “treated by Triton X-100”). Data are shown as mean + SD of three different batches.
When detected with ELISA, the percentages of DT antigenicity in the free-DT fractions were similar to those of protein content, while the antigenicity from the liposome-associated DT fractions was much less (Fig. 1b). This significant loss of DT antigenicity is probably due to the shielding effect of the liposome on the encapsulated DT. Selected formulations were eluted with PBS (pH 7.4) in SEC to examine the status of DT-liposome association in physiological condition. DT antigenicity of the resulting fractions were measured and compared with those eluted by CB/PB (Fig. 1c and d). These figures show that part of the liposome-associated DT, probably attached to the liposome surface, was dissociated from the liposomes. When the liposome-associated DT fractions were further treated with Triton X-100, increased DT antigenicity was detected, and the overall recovery rate of DT antigenicity was increased to about 80%, comparable to the control group. This increased DT antigenicity was previously shielded by liposome and represents the encapsulated DT under physiological condition (Table II). The DT-encapsulation ratio in PB at pH 7.4 was significantly higher than that in CB at pH 5.0 (p < 0.001, t-test), most likely due to the higher negative charge of DT at pH 7.4, which facilitates the electrostatic interaction between DT and the liposome components.
Elasticity of the DT-Loaded Vesicles
The extruded volumes of DT-loaded vesicle formulations and the calculated elasticity indices are shown in Fig. 2a and b, respectively. The unloaded surfactant vesicles showed relatively high elasticity. However, it was dramatically reduced by the loading of DT. When 0.2 mg/ml or more DT was incorporated, the dispersions could hardly be extruded, similar to the rigid DT-Lip. On the other hand, DT-ELip retained its elasticity and can be extruded through filter with 30 nm pore size. The particle size of DT-ELip (∼175 nm) was much larger than that of empty vesicles (∼100 nm). For this reason, even though its extrusion speed was slightly slower than the empty surfactant vesicles, it shows the highest elasticity index.
Fig. 2.
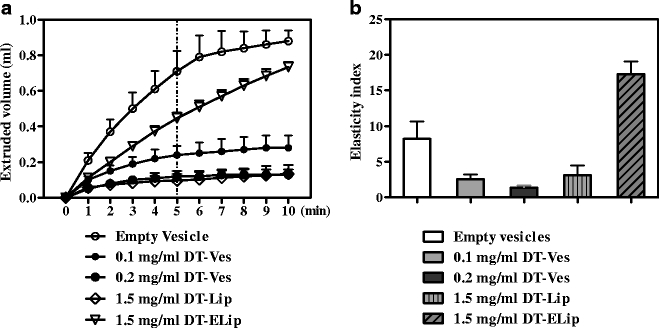
The vesicle elasticity measured with the extrusion method. DT vesicle formulations were extruded through polycarbonate filters with pore size of 30 nm under a constant pressure of 10 bar. The extruded volumes of these dispersions were recorded once per minute during 10 min (a). Particle sizes of the dispersions were measured by dynamic light scattering, and the elasticity indices were calculated using the extruded volumes after 5 min (b). Data shown are mean + SD of three batches.
Analysis of Antibody Titers Induced in TCI
DT-Loaded Anionic Surfactant Vesicles
First, TCI was performed by applying free DT and DT-Ves onto microneedle-treated or intact skin using CT as an adjuvant. The serum IgG titers after prime, the first boost and the second boost and the neutralizing antibody titers after the second boost through occlusive application are provided in Fig. 3. Non-occlusive application of the same formulations was also studied, and no significant difference was found compared to the corresponding occlusive groups. For clarity, only data from non-occlusive application of DT-Ves onto intact skin are shown in Fig. 3b. Pretreatment of the skin using the microneedle array improved the antibody responses drastically for both free DT and DT-Ves (Fig. 3a vs. b). The presence of CT further potentiated the IgG titers for DT-Ves (p > 0.05, p < 0.001 and p < 0.001 after prime, the first boost and second boost, respectively) and for free DT (p < 0.05, p < 0.001 and p < 0.05 after prime, the first boost and second boost, respectively). For DT-Ves, IgG titers were very close to those induced by s.c. DT-alum after the second boost (p > 0.05), whereas for free DT, comparable IgG titers were obtained from the first boost on (p > 0.05). Surprisingly, DT-Ves formulations did not induce higher IgG titers than free DT through either occlusive or non-occlusive application (p > 0.05 for all three time points). The IgG1 followed the IgG and showed slightly lower titers with similar trend, while the IgG2a titers were much less developed with many non-responders (data not shown).
Fig. 3.
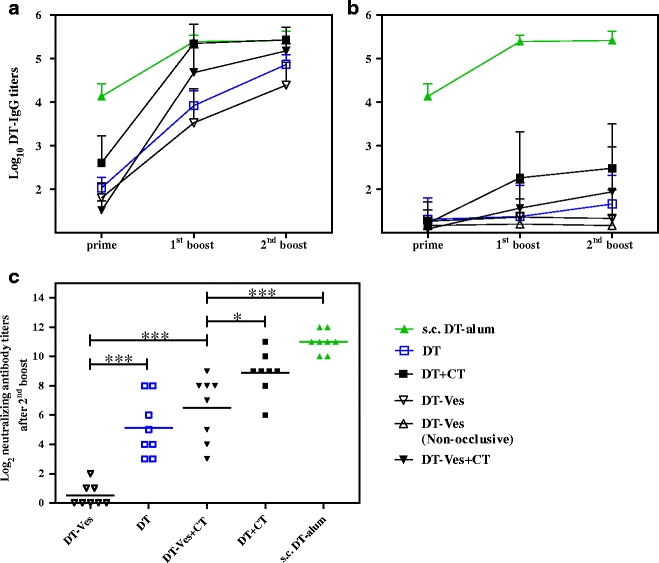
DT-specific IgG titers and neutralizing antibody titers after TCI of DT-Ves and free DT using CT as an adjuvant. Formulations were applied occlusively (unless specified in b) on microneedle-treated (a) or intact skin (b) at day 0, 21 and 42; s.c. injection of DT-alum served as control. Sera were collected after prime, the first boost and the second boost (day 20, 41 and 55), and IgG titers were determined with ELISA. Data shown as mean + SD (n = 8). Non-responders were given an arbitrary log-value of 1. Neutralizing antibody titers of TCI on microneedle-pretreated skin after the second boost were evaluated with Vero cell test. An arbitrary log-value of 0 was given when titers were below detection limit (c). Neutralizing antibody titers from intact skin were all below detection limit and therefore not shown (*: p < 0.05, **: p < 0.01, ***: p < 0.001).
The neutralizing antibody titers showed the same trend as the IgG titers after the second boost (Fig. 3c). The presence of CT enhanced the titers of both free DT and DT-Ves (p < 0.001 for both). The differences between free DT and DT-Ves are more pronounced. DT-Ves induced lower titers than DT (p < 0.001 and p < 0.05 for with and without CT, respectively). The titers induced by DT-Ves with CT were lower than those of s.c. DT-alum (p < 0.001), whereas for IgG titers, the difference was not significant. In the presence of CT, free DT induced comparable neutralizing antibody titers to those of s.c. DT-alum, the same as the IgG titers.
In TCI on intact skin, free DT and DT-Ves resulted in neither substantial antibody responses nor detectable neutralizing antibody titers. DT-Ves did not improve the immune responses of free DT, either.
As DT is released quickly from DT-Ves at pH 7.4, and the negative surface potential may reduce the interactions between vesicles and the cell membranes of APCs, positively charged vesicles were included in our studies. Initially, we focused on developing positively charged, DT-loaded, surfactant vesicles to obtain high DT-vesicle association at pH 7.4. However, by replacing TR-70 with DOTAP or dimethyl dioctadecyl ammonium as the charge inducer, stable formulations could not be obtained. The cationic liposome formulations were developed instead, in which considerable amount of DT has stable association with liposomes when diluted in PBS.
DT-Loaded Cationic Liposomes
On microneedle-treated skin, TCI was performed using DT-Lip, DT-Lip-5 and DT-ELip using CT as an adjuvant. Only occlusive application was performed, as no differences between occlusion and non-occlusion were observed in the study of DT-Ves. Serum IgG and neutralizing antibody titers are provided in Fig. 4. The presence of CT potentiated the IgG titers and neutralizing antibody titers of DT-Lip significantly (p < 0.001 for IgG titers after the first and second boost, and the neutralizing antibody titers, Fig. 4b and c). However, the DT liposome formulations did not induce higher immune responses than free DT, either. DT-ELip resulted in lower IgG titers after prime and the first boost, and lower neutralizing antibody titers after the second boost as compared to free DT (p < 0.01, p < 0.001 and p < 0.01, respectively). Only after the second boost, it induced a comparable level of IgG titers as free DT (p > 0.05), higher than those of DT-Lip-5 and DT-Lip (p < 0.05 and p < 0.001, respectively, Fig. 4a). DT-Lip and DT-Lip-5 induced lower IgG titers than free DT at all three time points (p < 0.05, p < 0.001 and p < 0.001, respectively, Fig. 4a) and lower neutralizing antibody titers (p < 0.001, Fig. 4c). Similar titers induced by DT-Lip and DT-Lip-5 indicate that the immunogenicity is not sensitive to the different DT-liposome association ratios (Fig. 4a). Same as the study of DT-Ves, the IgG1 titers of DT liposome formulations followed the IgG titers and showed a similar trend. The IgG2a titers were much less developed (data not shown).
Fig. 4.
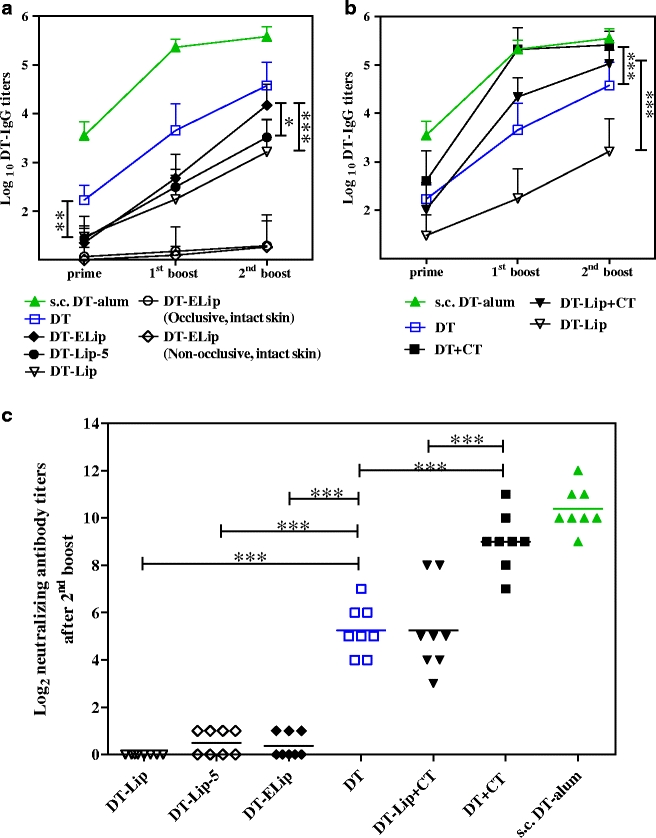
DT-specific IgG titers and neutralizing antibody titers induced by DT liposome formulations in TCI. Formulations were applied occlusively on microneedle-treated skin (unless specified in a) at day 0, 21 and 42; s.c. injection of DT-alum as control. Sera were collected after prime, the first boost and the second boost (day 20, 41 and 55), and IgG titers were determined with ELISA (a and b). Data shown as mean + SD (n = 8). Non-responders were given an arbitrary log-value of 1. Neutralizing antibody titers after the second boost were evaluated with Vero cell test (c). An arbitrary log-value of 0 was given when titers were below detection limit (*: p < 0.05, **: p < 0.01, ***: p < 0.001).
As for DT-Ves, the loss of vesicle elasticity by the loading of DT was also suspected to account for its low immunogenicity on intact skin, TCI of DT-ELip on intact skin was included because of its superior elasticity. However, the IgG levels induced were low and comparable to those of free DT and DT-Ves (Fig. 4a vs. Fig. 3), and occlusive and non-occlusive applications showed no difference in serum IgG titers (Fig. 4a).
Analysis of Antibody Titers Induced via i.d. Immunization
All the data suggest that vesicle formulations do not improve the immune response in TCI. To study whether differences in delivery efficiency across the skin barrier play a role, DT-Lip and free DT were administered via i.d. injection. The resulting serum IgG, IgG1, IgG2a and neutralizing antibody titers are shown in Fig. 5. The IgG, IgG1 and neutralizing antibody titers from free DT and DT-Lip were not significantly different. Unlike TCI, i.d. immunization of free DT and DT-Lip induced substantial IgG2a titers. Interestingly, DT-Lip induced a faster increase in IgG2a titers than free DT, and significantly higher IgG2a titers were found after the first and the second boost (Fig. 5b, p < 0.001, two-way ANOVA), suggesting immune modulation to a more Th1-biased response.
Fig. 5.
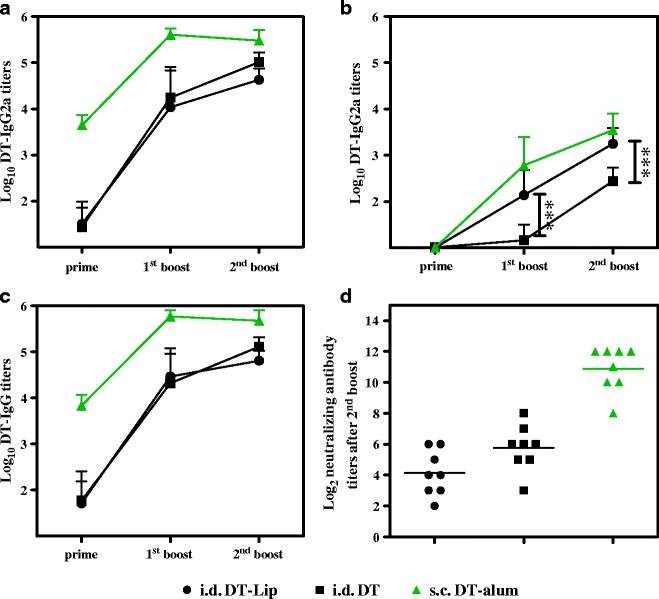
DT-specific IgG subtype antibody titers and neutralizing antibody titers after i.d. injection of DT-Lip and free DT. Vaccination was performed at day 0, 21 and 42; s.c. injection of DT-alum was as control. Sera were collected after prime, the first boost and the second boost (day 20, 41 and 55), and IgG, IgG1 and IgG2a titers were determined with ELISA (a, b and c). Data shown as mean + SD (n = 8). Non-responders were given an arbitrary log-value of 1. Neutralizing antibody titers after the second boost were evaluated with Vero cell test (d). An arbitrary log-value of 0 was given when titers were below detection limit (***: p < 0.001).
DC Maturation upon Contact with DT-Loaded Vesicles
To circumvent the complicated factors in delivery efficiency across the skin barrier, the direct stimulatory effects of DT formulations on the maturation of DCs were investigated in vitro. The up-regulation of the surface markers, MHC II, CD83 and CD86, was determined after culturing immature DCs in the presence of the DT formulations (Fig. 6). DT alone had no effect on the surface markers tested. CT showed the most potent stimulation to the immature DCs. It significantly up-regulated MHC II expression to levels higher than LPS and at the same time moderately enhanced those of CD86 as compared to free DT (p < 0.001 and p < 0.01, respectively, one way ANOVA). DT-Ves, DT-Lip and DT-ELip did not show significant up-regulation to all three surface markers as compared to free DT.
Fig. 6.

Stimulatory effects of DT formulations on immature DCs as indicated by the up-regulation of the surface markers MHC II, CD83 and CD86. The level of these three surface markers induced by 50 ng/ml LPS was set as 100%. The culture medium has a basal level of stimulation to the DCs as it contains GM-CSF. The final concentrations of each formulation component in culture medium were DT (2 μg/ml), DT+CT (2 μg/ml DT with 2 μg/ml CT), DT-Lip and DT-ELip (2 μg/ml DT with 70 μg/ml total lipids), DT-Ves (2 μg/ml DT with 140 μg/ml total surfactants). Data shown are mean + SD (n ≥ 3).
DISCUSSION
In the current study, stable DT-loaded liposome formulations were prepared and characterized in terms of their particle size, ζ-potential, degree of DT-vesicle association and vesicle elasticity. Two types of DT vesicle formulations, cationic liposomes and anionic surfactant vesicles, were studied in TCI, and the skin was either pretreated with microneedle arrays or left intact. Pretreatment with the microneedle array and co-administration with CT improved the immune responses of all of the formulations dramatically. However, the immunogenicity was not improved further by using vesicle formulations.
Initially, surfactant vesicles were chosen to formulate DT for TCI as a vaccine delivery system, as they have been shown to enhance the transport of small molecule drugs across human skin. Evidence for an exceptional interaction between these vesicles and human skin in vivo was obtained by freeze fracture electron microscopy and Fourier transform infrared studies (24,33); vesicular structures were observed in the deep layers of the stratum corneum after 1 h non-occlusive application. However, there was no evidence of vesicle material in the viable epidermis. The same vesicle system was employed for DT delivery in the current study. However, on intact skin, no substantial immune response was induced by DT-Ves and free DT, following either occlusive or non-occlusive application. The comparable immunogenicity of DT-Ves and free DT on intact skin indicates that vesicles do not efficiently transport the antigen across the stratum corneum. As this could be due to fast DT release from vesicles at pH 7.4 and the loss of vesicle elasticity when DT is incorporated, it was decided to prepare DT-ELip, which showed both association of DT with the vesicles/liposomes at pH 7.4 and an increased elasticity when DT is associated with the liposomes. However, in the subsequent in vivo immunization study, DT-ELip did not enhance the immunogenicity of topically applied DT on intact and microneedle-pretreated skin. As the composition of the Span liposomes is very similar to that of Transfersomes® used in TCI for hepatitis B surface antigen and gap junction protein, the outcome of our studies is in contrast to those reporting that ultradeformable vesicles induce potent immune responses on intact skin after non-occlusive application (20,34). In our hands, elastic vesicles enhance the transport of small molecules across the skin (22–24), but do not improve the immunogenicity of topically applied antigens, e.g. DT. In addition, there is no difference between occlusive and non-occlusive application for the elastic liposomes. Therefore, the osmotic gradient does not seem to play a central role as a driving force for antigen diffusion using these formulations.
To facilitate the transport of antigens into the viable epidermis, inclusion of skin barrier disruption methods, e.g. microneedle array pretreatment, tape striping or electroporation etc., appears to be crucial (35). Microneedle pretreatment has been shown to enhance the transcutaneous transport significantly. Henry et al. demonstrated four orders of magnitude increase in permeability for calcein and BSA through human epidermis in vitro after pretreatment with a microneedle array of 150 μm needle length (9), while Verbaan et al. showed penetration of 72 kDa dextran across microneedle-pretreated human skin in vitro using 300 μm long microneedle array (29). In the current study, the improved immunogenicity provided by the microneedle pretreatment may be attributed not only to enhanced antigen transport across the skin barrier, but also to the danger signals (IL-1α, IL-1β, GM-CSF and TNF-α released from the surrounding keratinocytes) caused by the skin barrier disruption, which sensitize and mobilize the skin APCs (36).
The induced immune response depends on both the permeation along the conduits and the interaction of the formulation with the skin APCs. I.d. vaccination was performed to exclude the influence of transport along the conduits on the immune responses. In addition, the DC study provided equal contact of DT to the APCs in vitro. I.d. vaccination of free DT and DT-Lip induced similar IgG1 and neutralizing antibody titers. This indicates that liposomes do not stimulate maturation of DCs. When comparing i.d. vaccination with TCI on microneedle-pretreated skin, where the same dose of DT in DT-Lip induced significantly lower immune response than free DT, it is clear that the transport of DT-loaded liposomes along the conduits is one of the limiting factors. This might be due to non-specific bindings of the positively charged liposomes to other cell membranes and negatively charged intercellular proteins, as reported by Yan et al. (37). As the negatively charged vesicles also reduced the immune response, it seems that a lower diffusion rate along the conduits may also play a role. An application period longer than 1 h may enhance the diffusion and improve the immune responses in TCI, as the conduits formed by the microneedles remain open for at least a few hours as reported by Bal et al. (38) and Banga et al. (39).
In TCI on microneedle-pretreated skin, it is notable that free DT induced faster IgG development than DT liposome formulations (Fig. 4). Most probably the soluble antigen diffuses faster along the conduits into the skin and arrives earlier in the viable layers and the peripheral draining lymph nodes. The role of direct lymphatic drainage of free antigen and the subsequent capture by blood-derived, lymph node-resident DCs for presentation to T cells has been highlighted in recent studies (40–43). So far, a DT/CT mixture, soluble antigen with soluble adjuvant, is the most immunogenic formulation in TCI on microneedle-pretreated skin. CpG ODN also demonstrated potent immune potentiation and modulation properties in our previous TCI study of DT (44). Moreover, the alum-adsorbed DT, big particles of several microns, reduced IgG titers dramatically and diminished the neutralizing antibody titers to undetectable level when applied on microneedle-treated skin (data not shown). Therefore, including other potent and soluble adjuvants is one of the first directions for further optimization of TCI formulations. Antigen-adjuvant conjugates and antigen-adjuvant fusion constructs may also be potential candidates. Furthermore, it has been reported that following i.d. injection in mice, 20 nm particles are taken up into the lymphatics more readily than 45 or 100 nm particles and are retained in the lymph nodes for a longer time (up to 120 h) (45). These ultra-small particles as well as some specific types of vesicles, e.g. ISCOMS, virus-like particles and surface-modified liposomes (46,47), hold promise in TCI when combined with microneedle array pretreatment. Last but not least, instead of relying on passive diffusion following microneedle treatment, the development of a microneedle array through which the vaccine formulation could be injected would provide more precise dose control and improve the vaccination efficiency.
CONCLUSION
We have demonstrated in the current study that the microneedle array pretreatment of the skin and co-administration of CT significantly enhances the immunogenicity of topically applied DT. However, formulating the antigen with vesicles, rigid or elastic, anionic or cationic, applied occlusively or non-occlusively, does not further improve the immune responses.
ACKNOWLEDGEMENTS
Tsambika Hahn is acknowledged for her efforts in developing stable, DT-loaded, surfactant-based, cationic vesicle formulations. We’d also like to thank Dr. Robert Poole and Dr. Myrra Carstens for their critical reading of this manuscript. This work was financially supported, in part, by the Chinese Scholarship Council.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
ABBREVIATIONS
- Alum
Adju-Phos®, colloidal aluminum phosphate
- APCs
antigen-presenting cells
- CB(S)
citrate buffer(ed saline)
- CT
cholera toxin
- DC
dendritic cell
- DOTAP
1,2-dioleoyl-3-trimethylammonium-propane chloride salt
- DT
diphtheria toxoid
- GM-CSF
granulocyte-macrophage colony-stimulating factor
- HRP-GAM
horseradish peroxidase conjugated goat anti-mouse
- i.d.
intradermal
- IL-4
interleukin-4
- L-595
sucrose-laurate ester
- LPS
lipopolysaccharide
- PB(S)
phosphate buffer(ed saline)
- PDI
polydispersity index
- PEG-8-L
octaoxyethylene laurate ester
- s.c.
subcutaneous
- SEC
size exclusion chromatography
- SPC
soybean phosphatidyl choline
- TCI
transcutaneous immunization
- TMB
chromogen 3, 3′, 5, 5′-tetramethylbenzidine
- TR-70
sodium bistridecyl sulfo succinate
REFERENCES
- 1.Simerska P, Moyle PM, Olive C, Toth I. Oral vaccine delivery—new strategies and technologies. Curr Drug Deliv. 2009;6:347–358. doi: 10.2174/156720109789000537. [DOI] [PubMed] [Google Scholar]
- 2.Mikszta JA, Laurent PE. Cutaneous delivery of prophylactic and therapeutic vaccines: historical perspective and future outlook. Expert Rev Vaccines. 2008;7:1329–1339. doi: 10.1586/14760584.7.9.1329. [DOI] [PubMed] [Google Scholar]
- 3.Slutter B, Hagenaars N, Jiskoot W. Rational design of nasal vaccines. J Drug Target. 2008;16:1–17. doi: 10.1080/10611860701637966. [DOI] [PubMed] [Google Scholar]
- 4.Lu D, Hickey AJ. Pulmonary vaccine delivery. Expert Rev Vaccines. 2007;6:213–226. doi: 10.1586/14760584.6.2.213. [DOI] [PubMed] [Google Scholar]
- 5.Valladeau J, Saeland S. Cutaneous dendritic cells. Semin Immunol. 2005;17:273–283. doi: 10.1016/j.smim.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 6.Glenn GM, Kenney RT, Ellingsworth LR, Frech SA, Hammond SA, Zoeteweij JP. Transcutaneous immunization and immunostimulant strategies: capitalizing on the immunocompetence of the skin. Expert Rev Vaccines. 2003;2:253–267. doi: 10.1586/14760584.2.2.253. [DOI] [PubMed] [Google Scholar]
- 7.Bos JD, Meinardi MM. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp Dermatol. 2000;9:165–169. doi: 10.1034/j.1600-0625.2000.009003165.x. [DOI] [PubMed] [Google Scholar]
- 8.Gerstel MS, Place VA. Drug delivery device, US, 1976.
- 9.Henry S, McAllister DV, Allen MG, Prausnitz MR. Microfabricated microneedles: a novel approach to transdermal drug delivery. J Pharm Sci. 1998;87:922–925. doi: 10.1021/js980042+. [DOI] [PubMed] [Google Scholar]
- 10.Prausnitz MR. Microneedles for transdermal drug delivery. Adv Drug Deliv Rev. 2004;56:581–587. doi: 10.1016/j.addr.2003.10.023. [DOI] [PubMed] [Google Scholar]
- 11.Ding Z, Verbaan FJ, Bivas-Benita M, Bungener L, Huckriede A, van den Berg DJ, et al. Microneedle arrays for the transcutaneous immunization of diphtheria and influenza in BALB/c mice. J Control Release. 2009;136:71–78. doi: 10.1016/j.jconrel.2009.01.025. [DOI] [PubMed] [Google Scholar]
- 12.Matriano JA, Cormier M, Johnson J, Young WA, Buttery M, Nyam K, et al. Macroflux microprojection array patch technology: a new and efficient approach for intracutaneous immunization. Pharm Res. 2002;19:63–70. doi: 10.1023/A:1013607400040. [DOI] [PubMed] [Google Scholar]
- 13.Widera G, Johnson J, Kim L, Libiran L, Nyam K, Daddona PE, et al. Effect of delivery parameters on immunization to ovalbumin following intracutaneous administration by a coated microneedle array patch system. Vaccine. 2006;24:1653–1664. doi: 10.1016/j.vaccine.2005.09.049. [DOI] [PubMed] [Google Scholar]
- 14.Cevc G, Gebauer D. Hydration-driven transport of deformable lipid vesicles through fine pores and the skin barrier. Biophys J. 2003;84:1010–1024. doi: 10.1016/S0006-3495(03)74917-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cevc G, Blume G. Hydrocortisone and dexamethasone in very deformable drug carriers have increased biological potency, prolonged effect, and reduced therapeutic dosage. Biochim Biophys Acta. 2004;1663:61–73. doi: 10.1016/j.bbamem.2004.01.006. [DOI] [PubMed] [Google Scholar]
- 16.Cevc G, Blume G. Biological activity and characteristics of triamcinolone-acetonide formulated with the self-regulating drug carriers, Transfersomes. Biochim Biophys Acta. 2003;1614:156–164. doi: 10.1016/S0005-2736(03)00172-X. [DOI] [PubMed] [Google Scholar]
- 17.Benson HA. Transfersomes for transdermal drug delivery. Expert Opin Drug Deliv. 2006;3:727–737. doi: 10.1517/17425247.3.6.727. [DOI] [PubMed] [Google Scholar]
- 18.Cevc G. Transdermal drug delivery of insulin with ultradeformable carriers. Clin Pharmacokinet. 2003;42:461–474. doi: 10.2165/00003088-200342050-00004. [DOI] [PubMed] [Google Scholar]
- 19.Guo J, Ping Q, Sun G, Jiao C. Lecithin vesicular carriers for transdermal delivery of cyclosporin A. Int J Pharm. 2000;194:201–207. doi: 10.1016/S0378-5173(99)00361-0. [DOI] [PubMed] [Google Scholar]
- 20.Mishra D, Dubey V, Asthana A, Saraf DK, Jain NK. Elastic liposomes mediated transcutaneous immunization against Hepatitis B. Vaccine. 2006;24:4847–4855. doi: 10.1016/j.vaccine.2006.03.011. [DOI] [PubMed] [Google Scholar]
- 21.Gupta PN, Mishra V, Singh P, Rawat A, Dubey P, Mahor S, et al. Tetanus toxoid-loaded transfersomes for topical immunization. J Pharm Pharmacol. 2005;57:295–301. doi: 10.1211/0022357055515. [DOI] [PubMed] [Google Scholar]
- 22.Honeywell-Nguyen PL, Arenja S, Bouwstra JA. Skin penetration and mechanisms of action in the delivery of the D2-agonist rotigotine from surfactant-based elastic vesicle formulations. Pharm Res. 2003;20:1619–1625. doi: 10.1023/A:1026191402557. [DOI] [PubMed] [Google Scholar]
- 23.Honeywell-Nguyen PL, Van den Bussche MH, Junginger HE, Bouwstra JA. The effect of surfactant-based elastic and rigid vesicles on the penetration of lidocaine across human skin. STP Pharma. 2002;12:257–262. [Google Scholar]
- 24.Honeywell-Nguyen PL, Gooris GS, Bouwstra JA. Quantitative assessment of the transport of elastic and rigid vesicle components and a model drug from these vesicle formulations into human skin in vivo. J Invest Dermatol. 2004;123:902–910. doi: 10.1111/j.0022-202X.2004.23441.x. [DOI] [PubMed] [Google Scholar]
- 25.El Maghraby GM, Williams AC, Barry BW. Oestradiol skin delivery from ultradeformable liposomes: refinement of surfactant concentration. Int J Pharm. 2000;196:63–74. doi: 10.1016/S0378-5173(99)00441-X. [DOI] [PubMed] [Google Scholar]
- 26.Ding Z, Bivas-Benita M, Hirschberg H, Kersten GFA, Jiskoot W, Bouwstra JA. Preparation and characterization of diphtheria toxoid-loaded elastic vesicles for transcutaneous immunization. J Drug Target. 2008;16:555–563. doi: 10.1080/10611860802228673. [DOI] [PubMed] [Google Scholar]
- 27.Peterson GL. Determination of total protein. Methods Enzymol. 1983;91:95–119. doi: 10.1016/S0076-6879(83)91014-5. [DOI] [PubMed] [Google Scholar]
- 28.Tierney R, Beignon AS, Rappuoli R, Muller S, Sesardic D, Partidos CD. Transcutaneous immunization with tetanus toxoid and mutants of Escherichia coli heat-labile enterotoxin as adjuvants elicits strong protective antibody responses. J Infect Dis. 2003;188:753–758. doi: 10.1086/377287. [DOI] [PubMed] [Google Scholar]
- 29.Verbaan FJ, Bal SM, van den Berg DJ, Groenink WH, Verpoorten H, Luttge R, et al. Assembled microneedle arrays enhance the transport of compounds varying over a large range of molecular weight across human dermatomed skin. J Control Release. 2007;117:238–245. doi: 10.1016/j.jconrel.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 30.van den Bergh BA, Wertz PW, Junginger HE, Bouwstra JA. Elasticity of vesicles assessed by electron spin resonance, electron microscopy and extrusion measurements. Int J Pharm. 2001;217:13–24. doi: 10.1016/S0378-5173(01)00576-2. [DOI] [PubMed] [Google Scholar]
- 31.Metz B, Jiskoot W, Hennink WE, Crommelin DJ, Kersten GF. Physicochemical and immunochemical techniques predict the quality of diphtheria toxoid vaccines. Vaccine. 2003;22:156–167. doi: 10.1016/j.vaccine.2003.08.003. [DOI] [PubMed] [Google Scholar]
- 32.Miyamura K, Tajiri E, Ito A, Murata R, Kono R. Micro cell culture method for determination of diphtheria toxin and antitoxin titres using VERO cells. II. Comparison with the rabbit skin method and practical application for seroepidemiological studies. J Biol Stand. 1974;2:203–209. doi: 10.1016/0092-1157(74)90016-X. [DOI] [PubMed] [Google Scholar]
- 33.Honeywell-Nguyen PL, de Graaff AM, Groenink HW, Bouwstra JA. The in vivo and in vitro interactions of elastic and rigid vesicles with human skin. Biochim Biophys Acta. 2002;1573:130–140. doi: 10.1016/s0304-4165(02)00415-4. [DOI] [PubMed] [Google Scholar]
- 34.Paul A, Cevc G, Bachhawat BK. Transdermal immunisation with an integral membrane component, gap junction protein, by means of ultradeformable drug carriers, transfersomes. Vaccine. 1998;16:188–195. doi: 10.1016/S0264-410X(97)00185-0. [DOI] [PubMed] [Google Scholar]
- 35.Mahe B, Vogt A, Liard C, Duffy D, Abadie V, Bonduelle O, et al. Nanoparticle-based targeting of vaccine compounds to skin antigen-presenting cells by hair follicles and their transport in mice. J Invest Dermatol. 2009;129:1156–1164. doi: 10.1038/jid.2008.356. [DOI] [PubMed] [Google Scholar]
- 36.Partidos CD, Muller S. Decision-making at the surface of the intact or barrier disrupted skin: potential applications for vaccination or therapy. Cell Mol Life Sci. 2005;62:1418–1424. doi: 10.1007/s00018-005-4529-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yan W, Chen W, Huang L. Mechanism of adjuvant activity of cationic liposome: phosphorylation of a MAP kinase, ERK and induction of chemokines. Mol Immunol. 2007;44:3672–3681. doi: 10.1016/j.molimm.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 38.Bal SM, Caussin J, Pavel S, Bouwstra JA. In vivo assessment of safety of microneedle arrays in human skin. Eur J Pharm Sci. 2008;35:193–202. doi: 10.1016/j.ejps.2008.06.016. [DOI] [PubMed] [Google Scholar]
- 39.Banga AK. Microporation applications for enhancing drug delivery. Expert Opin Drug Deliv. 2009;6:343–354. doi: 10.1517/17425240902841935. [DOI] [PubMed] [Google Scholar]
- 40.Kissenpfennig A, Henri S, Dubois B, Laplace-Builhe C, Perrin P, Romani N, et al. Dynamics and function of Langerhans cells in vivo: dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells. Immunity. 2005;22:643–654. doi: 10.1016/j.immuni.2005.04.004. [DOI] [PubMed] [Google Scholar]
- 41.Reddy ST, van der Vlies AJ, Simeoni E, Angeli V, Randolph GJ, O’Neil CP, et al. Exploiting lymphatic transport and complement activation in nanoparticle vaccines. Nat Biotechnol. 2007;25:1159–1164. doi: 10.1038/nbt1332. [DOI] [PubMed] [Google Scholar]
- 42.Iezzi G, Frohlich A, Ernst B, Ampenberger F, Saeland S, Glaichenhaus N, et al. Lymph node resident rather than skin-derived dendritic cells initiate specific T cell responses after Leishmania major infection. J Immunol. 2006;177:1250–1256. doi: 10.4049/jimmunol.177.2.1250. [DOI] [PubMed] [Google Scholar]
- 43.Sixt M, Kanazawa N, Selg M, Samson T, Roos G, Reinhardt DP, et al. The conduit system transports soluble antigens from the afferent lymph to resident dendritic cells in the T cell area of the lymph node. Immunity. 2005;22:19–29. doi: 10.1016/j.immuni.2004.11.013. [DOI] [PubMed] [Google Scholar]
- 44.Ding Z, Van Riet E, Romeijn S, Kersten GF, Jiskoot W, Bouwstra JA. Immune modulation by adjuvants combined with diphtheria toxoid administered topically in BALB/c mice after microneedle array pretreatment. Pharm Res. 2009;26:1635–1643. doi: 10.1007/s11095-009-9874-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Reddy ST, Rehor A, Schmoekel HG, Hubbell JA, Swartz MA. In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide) nanoparticles. J Control Release. 2006;112:26–34. doi: 10.1016/j.jconrel.2006.01.006. [DOI] [PubMed] [Google Scholar]
- 46.Saupe A, McBurney W, Rades T, Hook S. Immunostimulatory colloidal delivery systems for cancer vaccines. Expert Opin Drug Deliv. 2006;3:345–354. doi: 10.1517/17425247.3.3.345. [DOI] [PubMed] [Google Scholar]
- 47.Altin JG, van Broekhoven CL, Parish CR. Targeting dendritic cells with antigen-containing liposomes: antitumour immunity. Expert Opin Biol Ther. 2004;4:1735–1747. doi: 10.1517/14712598.4.11.1735. [DOI] [PubMed] [Google Scholar]