

Abstract
Background
Persistence of myofibroblasts is believed to contribute to the development of fibrosis in idiopathic pulmonary fibrosis (IPF). Transforming growth factor-β1 (TGF-β1) irreversibly converts fibroblasts into pathological myofibroblasts, which express smooth muscle α-actin (α-SMA) and produce extracellular matrix proteins, such as procollagen I (α1). Reactive oxygen species (ROS) produced by NADPH oxidases (NOX) have been shown to regulate cell differentiation. We hypothesized that NOX could be expressed in parenchymal pulmonary fibroblasts and could mediate TGF-β1–stimulated conversion of fibroblasts to myofibroblasts.
Methods
Fibroblasts were cultured from the lung of 9 Control and 8 IPF patients. We quantified NOX4, α-SMA and procollagen I (α1) mRNA and protein expression, ROS production and Smad2/3 phosphorylation, in the absence and in the presence of incubation with TGF-β1. PDGF-induced fibroblasts migration was also assessed.
Results
We found that (1) NOX4 mRNA and protein expression was upregulated in pulmonary fibroblasts from patients with IPF and correlated with mRNA expression of α-SMA and procollagen I (α1) mRNA; (2) TGF-β1 upregulated NOX4, α-SMA and procollagen I (α1) expression in control and IPF fibroblasts; (3) the change in α-SMA and procollagen I (α1) expression in response to TGF-β1 was inhibited by antioxidants and by a NOX4 siRNA; (4) NOX4 modulated α-SMA and procollagen I (α1) expression by controlling activation of Smad 2/3, and (5) NOX4 modulated PDGF-induced fibroblasts migration.
Conclusion
NOX4 is critical for modulation of pulmonary myofibroblast phenotype in IPF, probably by modulating the response to TGF-β1 and PDGF.
Keywords: Adult; Aged; Cell Differentiation; drug effects; physiology; Cells, Cultured; Female; Fibroblasts; drug effects; enzymology; pathology; Gene Expression Regulation, Enzymologic; Humans; Idiopathic Pulmonary Fibrosis; enzymology; pathology; Lung; enzymology; pathology; Male; Middle Aged; NADPH Oxidase; biosynthesis; genetics; physiology; Platelet-Derived Growth Factor; pharmacology; RNA, Messenger; genetics; Reactive Oxygen Species; metabolism; Reverse Transcriptase Polymerase Chain Reaction; methods; Smad2 Protein; metabolism; Smad3 Protein; metabolism; Transforming Growth Factor beta1; pharmacology; Up-Regulation
INTRODUCTION
Idiopathic pulmonary fibrosis (IPF) is the most common idiopathic interstitial pneumonia. IPF is a disease with poor prognosis and an aggressive nature, and poses major challenges to clinicians (1) (2). Currently, no effective therapies exist to stop ongoing fibrosis in IPF. Accumulation and persistence of myofibroblasts is believed to contribute to the development of fibrosis (3). Alpha-smooth muscle actin (α-SMA) expression, increased proliferative capacity, and increased generation and secretion of the extracellular matrix (ECM) proteins such as collagen and fibronectin are key hallmarks of myofibroblast differentiation in fibrotic disorders (4). It has been established that myofibroblast differentiation occurs through combinatorial signals involving TGF-β1 and integrin signalling (5).
Several studies suggest that oxidant–antioxidant imbalances in the lower respiratory tract play a significant role in the pathogenesis of IPF. Reactive oxygen species (ROS) can promote a profibrogenic environment in the lung through the modulation of the protease-antiprotease balance (6). Also, ROS activate and/or mediate the effects of growth-regulatory cytokines, including TGF-β1 (7) (8). However, the endogenous sources of oxidants in IPF are poorly known.
The NADPH oxidases (NOX) proteins generate ROS, namely superoxide, by electron transfer to oxygen. Superoxide can be further converted to hydrogen peroxide by cellular superoxide dismutases. NADPH oxidases consist of membrane-associated and cytosolic subunits (for review, see reference (9)). The most thoroughly investigated NADPH oxidase is the phagocytic gp91phox (NOX2)-containing subunit that interacts with a second membrane bound subunit, p22phox, and several cytosolic and regulatory subunits including p47phox, p67phox, and Rac. Both NOX1 and NOX4 are homologs of NOX2, and are expressed in different nonphagocytic cells (9). It has been recently shown that NOX1 and NOX4 are expressed in pulmonary arterial adventitial fibroblasts and that NOX4 contributes to the increase in ROS generation under hypoxic conditions, stimulating proliferation and inhibiting apoptosis of these cells (10). Furthermore, NOX4 is involved in TGF-β1-induced human cardiac fibroblast differentiation into myofibroblasts (11). Therefore, we hypothetized that NOX1 and NOX4 could be also expressed in parenchymal pulmonary fibroblasts and could mediate the effects of TGF-β1.
The aim of this study was to investigate NOX1 and NOX4 expression in pulmonary fibroblasts from control subjects and IPF patients and to analyse their respective role in mediating TGF-β1-induced fibroblast differentiation into myofibroblast, a key phenomenon in IPF pathogenesis (12). The myofibroblast phenotype was analyzed by examining α-SMA and procollagen I (α1) expression.
MATERIAL AND METHODS
See online supplement for further details.
Lung fibrosis patients
Fibroblasts were derived from lung tissue sampled from 8 IPF patients. Lung samples were obtained by open lung biopsy (n=4) or at the time of lung transplantation (n=4). IPF was diagnosed according to the ATS-ERS consensus criteria (17). Patients (6 males; 2 females) had a median age of 50 years [range, 44 to 69 yr]. 6 were ex-smokers, 1 never smoked and 1 was an active smoker. At the time of lung sampling, 3 patients were treated with low dose oral corticosteroids, associated with azathioprin in one patient.
Control patients
Fibroblasts were derived from lung samples from 9 patients (6 males, 3 females) undergoing lung surgery for removal of a primary lung tumor. Normal lung was obtained from a noninvolved segment, remote from the solitary lesion. Median age was 54 years [28 to 68]. Two patients had never smoked, 4 were ex-smokers and 3 were active smokers.
Culture of fibroblasts
Human lung fibroblasts were cultured from lung explants until passage 5 as previously described (13). In some experiments, cells were incubated in the presence of TGF-β1 (10 ng/ml, R&D Systems, Lille, France) or vehicle.
Cellular Toxicity
Cellular toxicity and viability were assessed by trypan blue exclusion and lactate dehydrogenase (LDH) release in the medium, as described previously (14).
Intracellular ROS production
Intracellular ROS production was quantified by measuring H2-DCFH-DA oxidation, as described previously (14) (15). Results are expressed in fluorescent arbitrary units.
Real-time RT-PCR
NOX1, 2, 3, 4 and 5, α-SMA and procollagen I (α1) mRNA expression were quantified by quantitative real-time RT-PCR by use of the PCR ABI 7700 apparatus (Applied Biosystems, Carlsbad, CA). Expression of mRNA was normalized to that of ubiquitin. In previous experiments, we verified that ubiquitin mRNA expression did not change during the different experimental conditions.
Cell transfection with siRNA
To suppress endogenous NOX4 expression, experiments involved a specific siRNA (Xeragon-Qiagen, Valencia, CA,) against NOX4 designed on the target region from the NOX4 gene (5′-AAACCGGCAGGAGUUUACCCAG-3′). Scrambled siRNA (nonhomologous to the human genome, 5′-AACCAGCAAGGUGUAUCGCCAC-3′) was used as a control.
Western blot analysis
Western blot was performed as described previously (16) with use of the following antibodies: polyclonal anti-human NOX4 (1/1000, Novus Biologicals, Inc., Littleton, CO), monoclonal anti α-actin (1/3000, Sigma-Aldrich, Saint Quentin Fallavier, France), and monoclonal anti-human anti Smad 2 and 3 (1/500, R&D Systems, Lille, France). The expression of NOX4 was normalized in respect to the expression of β-actin. The expression of non-phosphorylated Smad 2 (monoclonal antibody, 1/400, Cell Signalling, Saint Quentin Yvelines, France) was used to normalize the expression of phospho Smad 2/3.
Cell migration assay
Cell motility was examined as previously described (17) using a modified Boyden chamber apparatus (Transwell Costar) in response to 24 h stimulation with rhPDGF-BB 10ng/ml (R&D systems) or media alone. Chemotactic index was determined as the ratio of cells moving in response to a chemoattractant relative to cells moving in response to media control.
Immunohistochemical analysis
Immunohistochemistry was performed as previously described (16). 4–6 μm thick cryostat sections were fixed in acetone and reacted with the primary antibody rabbit polyclonal anti-NOX4 (1/500 dilution, kindly provided by Pr Victor Thannickal, University of Alabama at Birmingham, Birmingham, Alabama).
Statistical analysis
See online supplement for further details.
All the data are displayed as medians, interquartile range and minimum and maximum values. All of the analysis were performed by non-parametric tests. p<0.05 was considered as statistically significant.
RESULTS
Expression of NOX mRNA and oxidants production in pulmonary fibroblasts
Lung fibroblasts from control patients expressed NOX1, 2, 4 and 5 mRNA at similar degrees, whereas expression of NOX3 was not detected in these cells (fig 1A to D). When comparing IPF and C fibroblasts, NOX4 was the only NOX whose mRNA expression was increased in the former as compared to the latter condition (p=0.043). The increased NOX4 expression in IPF fibroblasts was confirmed at the protein level by western blot (p=0.025 C vs IPF, fig 1E). NOX4 mRNA and protein induction in IPF fibroblasts were accompanied by an increased intracellular ROS production in these cells as compared to control fibroblasts (p=0.028, fig 1F). Immunohistochemistry performed on control and IPF lung samples demonstrated that lung fibroblasts expressed immunoreactive NOX-4 in situ in the fibrotic lung (fig 2). Bronchial and alveolar epithelial cells, and pulmonary endothelial cells also expressed immunoreactive NOX-4.
Figure 1.
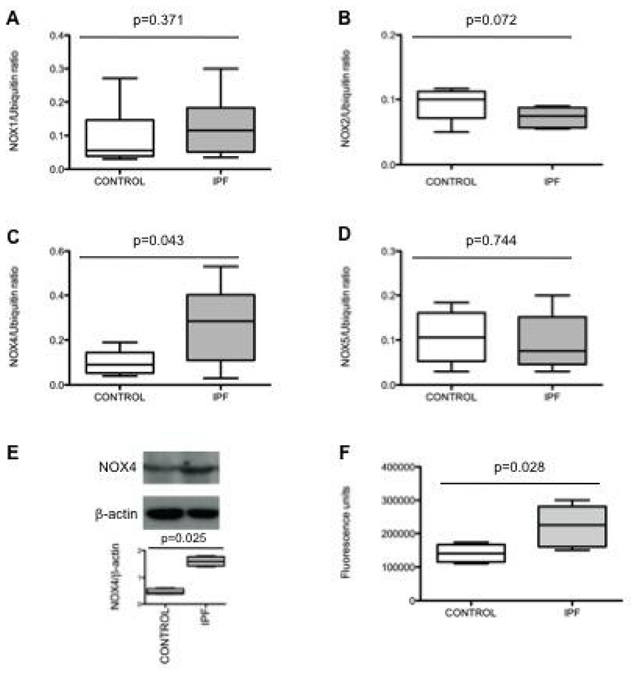
A, B, C and D: NOX1, 2, 4 and 5 mRNA expression in lung fibroblasts from patients with idiopathic pulmonary fibrosis (IPF) compared to controls expressed as the ratio to ubiquitin mRNA levels. Data are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values. E: Typical western-blot and quantification of NOX4 protein expression (67 kD) in lung fibroblasts from patients with IPF and controls. NOX4 is expressed as the ratio to β-actin expression. Data are presented as box-and- whiskers plot with median, interquartile range and minimum and maximum values. F: reactive oxygen species production by IPF and control fibroblasts assessed by measurement of H2-DCFH-DA oxidation. The values, expressed as arbitrary fluorescence units, are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values.
Figure 2.
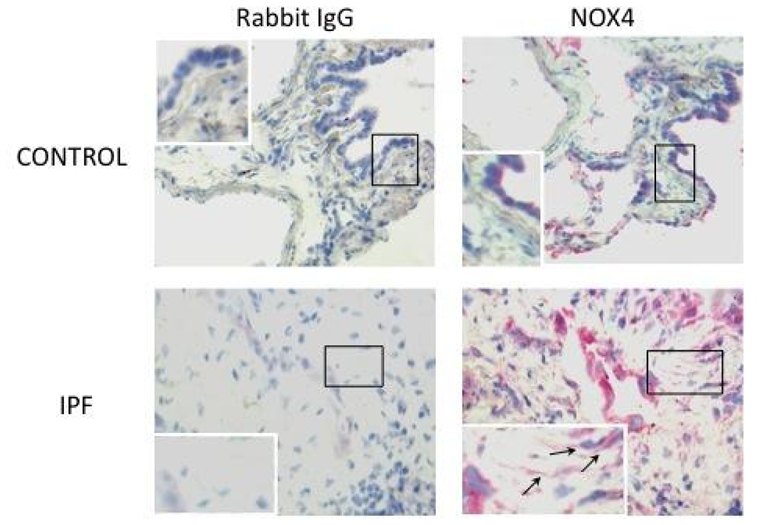
Detection of immunoreactive NOX-4 in the lung. Immunohistochemistry shows that bronchial and alveolar epithelial cells express NOX-4 in the normal lung. In IPF lung samples, hyperplastic alveolar cells and fibroblasts (arrows) are strongly labelled. The control antibody gave always a negative signal.
Expression of markers of myofibroblast diferentiation
The content of α-SMA mRNA was increased in IPF fibroblasts as compared to controls (p=0.045, fig E1 in on line supplement) whereas procollagen I (α1) mRNA content was similar in both groups (p=0.310, fig E1 in on line supplement). Analysis of fibroblasts from IPF patients showed a significant correlation between α-SMA or procollagen I (α1) mRNA and NOX4 mRNA expression (Spearman ρ = 0.994, p<0.0001 in both cases, fig E1 in on line supplement), suggesting a role of this NADPH oxidase homolog in myofibroblast differentiation.
Effect of TGF-β1
Since NOX4 was overexpressed in IPF fibroblasts and correlated with markers of fibroblast differentiation into myofibroblast, and since TGF-β1 is an essential inducer of myofibroblast differentiation (5), we analyzed if TGF-β1 modulated NOX4 expression by lung fibroblasts. Incubation of control fibroblasts with TGF-β1 for 18h induced a 3-fold increase of NOX4 mRNA content (fig 3A, p=0.014). In IPF fibroblasts, TGF-β1 induced a 8-fold increase of NOX4 mRNA content (fig 3B, p=0.008). As expected, TGF-β1 increased α-SMA and procollagen I (α1) mRNA expression in control and IPF fibroblasts (fig 3A and B, p=0.009, p=0.008, p=0.034, and p=0.029 for α-SMA in control and IPF fibroblasts and for procollagen I (α1) in control and IPF fibroblasts respectively). The increase in α-SMA was also observed at the protein level (fig 3C). Incubation of control fibroblasts with TGF-β1 induced a significant increase in ROS production, measured by oxidation of DCFH (fig 3C). This increase was more important in IPF than in control cells at 1, 3, and 24 h post-TGF-β1 (p=0.033, p=0.039 and p=0.028 at each time respectively, fig 3D).
Figure 3.

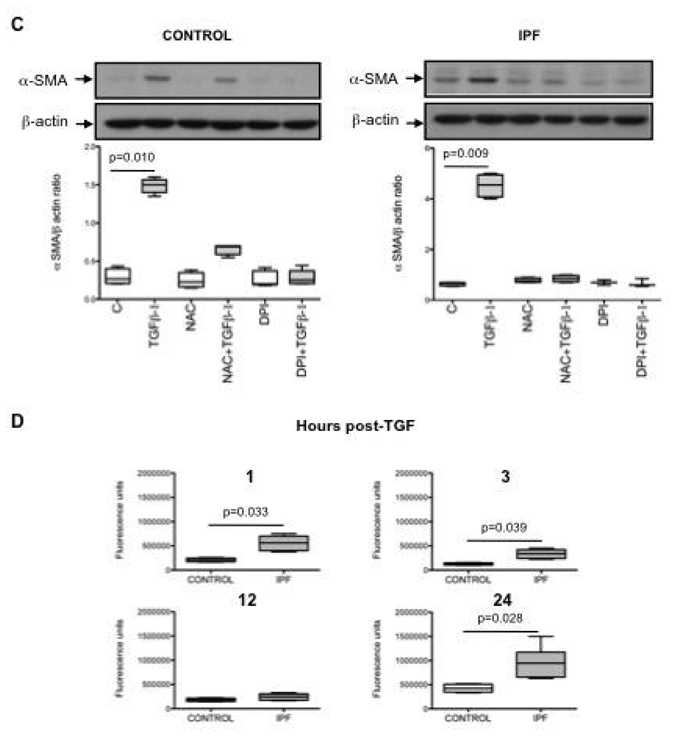
A and B: Effect of TGF-β1 in the presence or in the absence of N-acetylcysteine (NAC, 1 mM) or diphenylene iodonium (DPI, 10 μm) on NOX4, α smooth muscle actin (α-SMA) and procollagen I (α1) mRNA expression in lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF). Data are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values. C: Typical western blot and quantification of α-SMA protein expression in lung fibroblasts after TGF-β1 incubation in the presence or in the absence of NAC (1 mM) or DPI (10 μm). α-SMA is expressed as the ratio to β-actin expression. Data are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values. D: TGF-β1 stimulates ROS production in human lung fibroblasts. Control and IPF fibroblasts were stimulated with TGF-β1 for 1 to 24 h and ROS production was assessed by measurement of H2-DCFH-DA oxidation. The 4 figures show production by the 2 groups of cells after 1, 3 12 and 24 h post-TGF-β1 respectively. Values are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values.
Role of NOX4 on TGF-β1 induced myofibroblast differentiation
We first examined if ROS, the product of NOX4 activity, were involved on TGF-β1 induced myofibroblast differentiation. First, we examined the effect of the antioxidant N-acetylcystein. Pretreatment of control and IPF fibroblasts for 1 h with N-acetylcysteine (1 mM) totally prevented the increase of α-SMA and procollagen I (α1) mRNA expression induced by TGF-β1 (fig 3A, B and C). This result strongly suggested that the oxidant-antioxidant imbalance is involved in the effect of TGF-β1.
In order to examine the involvement of NOX4 on TGF-β1 effects, we first incubated the fibroblasts for 1 h with diphenylene iodonium (DPI, 10 μM), an inhibitor of flavoprotein-dependent oxidases including NOX (18). DPI prevented TGF-β1 induced α-SMA and procollagen I (α1) mRNA expression in both control and IPF fibroblasts (fig 3A, B and C).
Second, we examined the effects of cell transfection with a NOX4 siRNA. NOX4 siRNA significantly reduced NOX4 mRNA levels at 24 h in both control and IPF fibroblasts whereas no such effect was observed with lipofectamine or scrambled siRNA (fig 4A and B).
Figure 4.
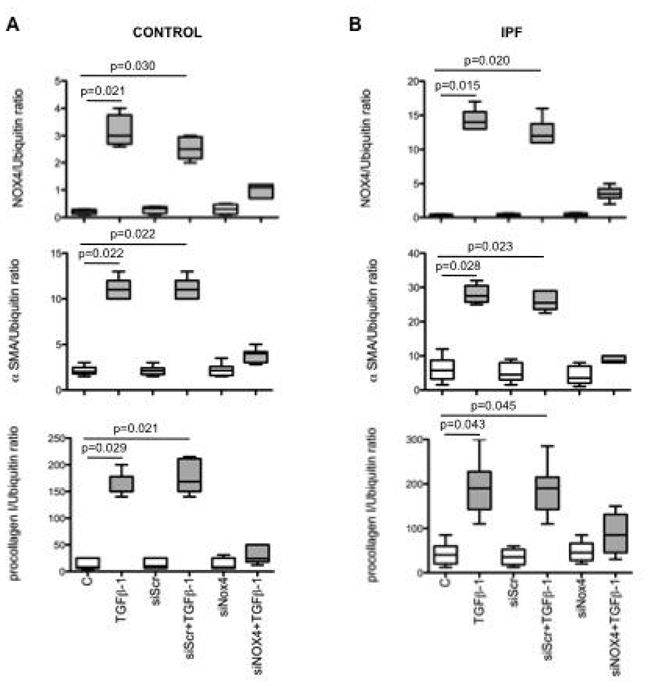
A and B: Effect of cell transfection with a siRNA anti-NOX4 on NOX4, α-SMA and procollagen I (α1) mRNA expression induced by TGF-β1 in lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF). siScr: scrambled siRNA. NOX4, α-SMA and procollagen I (α1) mRNA levels are expressed as the ratio to ubiquitin mRNA levels. Values are presented as box-and-whiskers plot with median, interquartile range and minimum and maximum values.
Transfection with NOX4 siRNA significantly inhibited the effect of TGF-β1 on myofibroblats differentiation in both control and IPF cells (fig 4A and B). Indeed, expression of α-SMA and procollagen I (α1) were reduced by 67% and 68% and by 64% and 54% in control and IPF fibroblasts respectively. These results demonstrate that NOX4 mediates the effect of TGF-β1 on myofibroblast differentiation.
It has to be noted that both N-acetylcysteine and DPI reversed NOX4 mRNA induction by TGF-β1, showing probably an autocrine role of NOX4-derived ROS on NOX4 induction by TGF-β1.
None of the different pharmacological treatments and transfection procedures altered cell viability (data not shown).
Role of NOX4 on TGF-β1 induced SMAD 2 and 3 phosphorylation
It has been shown that Smads mediate TGF-β1 induction of α-SMA in human lung fibroblasts (19) (20). Since phosphorylation of Smad2/3 can be modulated by ROS (11), we analyzed whether NOX4 was involved on Smad2/3 phosphorylation induced by TGF-β1. The results of these experiments show that 1 h cell pretreatment with both N-acetylcysteine and DPI (1mM and 10 μM respectively) prevented Smad2/3 phosphorylation induced by 18 h incubation with TGF-β1 in control and IPF fibroblasts (fig 5A and B). Cell transfection with the NOX4 siRNA gave similar results (fig 5A and B). These results demonstrate that NOX4-produced ROS are involved in Smad2/3 phosphorylation by TGF-β1.
Figure 5.
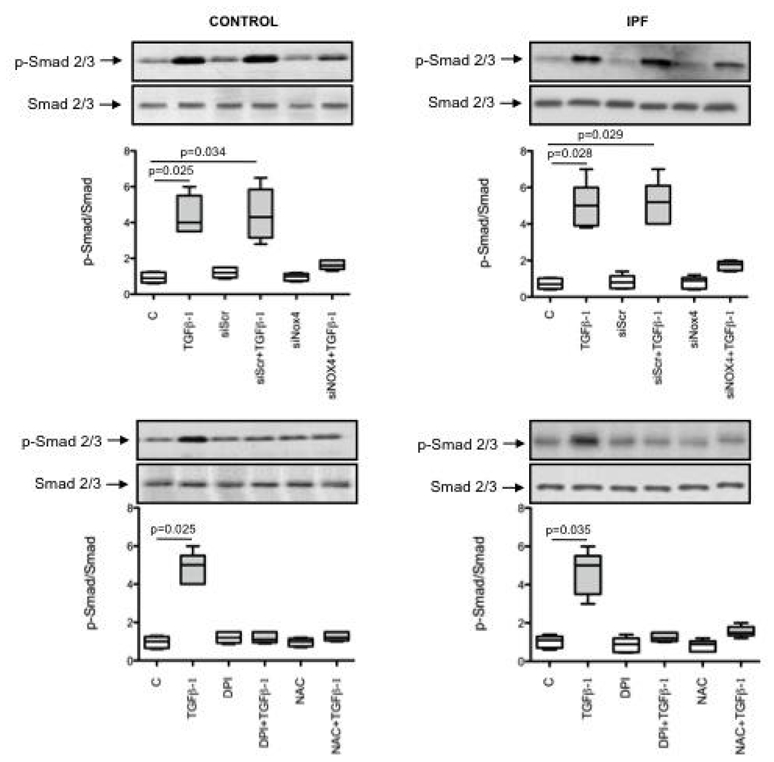
Typical western bolt showing the expression of phosphorylated (p-Smad 2/3) and non-phosphorylated (Smad 2/3) in human lung fibroblasts from controls and patients with idiopathic pulmonary fibrosis (IPF) after incubation with TGF-β1 in the presence or in the absence of a scrambled siRNA (siScr), a siRNA anti-NOX4 (siNOX4), diphenylene iodonium (DPI, 10 μm) or N-acetylcysteine (NAC, 1 mM). Box-and-whiskers plot with median, interquartile range and minimum and maximum values show quantification of p-Smad 2/3 expressed as the ratio of Smad 2/3 expression.
Role of NOX4 on PDGF induced fibroblast migration
Finally, we analyzed if NOX4 was involved in fibroblasts migration induced by PDGF-BB, a strong migration stimulus. First, we examined if PDGF modulated NOX4 expression by lung fibroblasts. Incubation of control fibroblasts and IPF fibroblasts with PDGF for 24 h (10 ng/ml) induced a 1.5 and 2.5-fold increase of NOX4 mRNA content respectively (fig E2 in online supplement, p=0.028 and p=0.002 respectively).
Having demonstrated that NOX4 was upregulated by PDGF, we analyzed its role on cell migration by incubating cells with NAC or DPI. Both NAC and DPI suppressed PDGF-induced fibroblasts migration in control and IPF fibroblasts, showing a role of NOX4 in this process (fig E2 in online supplement and data not shown).
DISCUSSION
The main results of the present study are that (1) NOX4 mRNA expression was upregulated in pulmonary fibroblasts from patients with IPF and correlated with mRNA expression of α-SMA and procollagen I (α1) mRNA; (2) TGF-β1 upregulates NOX4, α-SMA and procollagen I (α1) mRNA and α-SMA protein expression in control and IPF human pulmonary fibroblasts; (3) the change in α-SMA and procollagen I (α1) expression in response to TGF-β1 requires NOX4-derived ROS; (4) NOX4 modulates α-SMA and procollagen I (α1) expression by controlling activation of Smad 2/3, and (5) NOX4 modulates fibroblasts migration induced by PDGF-BB. These new findings support the notion that NOX4 is critical for modulation of pulmonary myofibroblast phenotype in IPF, probably by modulating the response to TGF-β1 and PDGF. Similar results concerning the response to TGF-β1 were very recently reported by Hecker and colleagues (21) in an article published after the present study was submitted for publication.
Expression of NOX4 in control fibroblasts
Expression of NOX4 protein was detected in lungs from both control subjects and patients with idiopathic pulmonary artery hypertension, the expression was predominantly in the medial layer of pulmonary arteries and was higher in patients than in controls (10). Accordingly, pulmonary artery adventitial fibroblasts in culture also expressed both NOX1 and NOX4 (22). In agreement with these findings, in the present study we found that parenchymal pulmonary fibroblasts from control and IPF patients expressed both NOX1 and NOX4. This result is different from data reported in cardiac fibroblasts in which NOX1 expression was barely detectable (11), stressing the cell specificity of NOX expression. To the best of our knowledge this is the first demonstration that non-vascular pulmonary fibroblasts express NOX. It is noteworthy that our experiments were performed on cells in the fifth passage, since progressive down-regulation of NOX4 expression has been correlated with increasing passage number (23). However, our immunohistochemical results demonstrate that fibroblasts do express immunoreactive NOX-4 in situ in the fibrotic lung.
NOX4 up-regulation in IPF fibroblasts
The mechanism(s) involved in NOX4 up-regulation in IPF fibroblasts are difficult to analyze since very few data concerning the regulation of NOX4 activity and expression are available. Recent data from recombinant NOX4 expression suggest that NOX4 enzymatic activity depends on the membrane-associated p22phox subunit, whereas cytosolic subunits, phosphorylation, or interaction with Rac are apparently not required for its activation (24) (25). Concerning the regulation of gene expression, and the relevant transcription factors involved, very few data are available since promoter studies of NOX4 are lacking. Our data clearly show that TGF-β1 increases NOX4 expression both in control and in IPF fibroblasts, in accordance with previous data in cardiac fibroblasts and airway smooth muscle (11) (26) (27). Interestingly, the increase was stronger in IPF fibroblasts than in controls. One can speculate that TGF-β1 can be responsible of NOX4 up-regulation in IPF fibroblasts ex vivo via an autocrine loop since expression of TGF-β1 has been shown to be increased in these cells (28). Furthermore, high levels of TGF-β1 produced by epithelial cells could contribute to up-regulation in fibroblasts in situ in the lung of IPF patients (29).
NOX4 induction by TGF-β1 was inhibited by N-acetylcysteine and DPI suggesting that NOX-produced ROS are implicated in NOX4 gene expression regulation. We obtained similar results in human alveolar bronchiolar and epithelial cell lines (14), but the molecular mechanisms involved are unknown. Clearly, more studies are needed to understand NOX4 gene expression regulation in IPF fibroblasts.
Role of NOX4 on myofibroblasts differentiation in IPF
Whatever the mechanism(s) involved in gene expression regulation, NOX4 was involved in myofibroblast differentiation in IPF. This conclusion is supported by 1) the correlation between basal NOX4 and α-SMA and procollagen I (α1) mRNA expression, and 2) the prevention of TGF-β1 differentiating effects by the NOX4 siRNA. This last phenomenon was also observed in cardiac fibroblasts (11). It has to be noted that we can not ensure that NOX4-produced ROS were involved in myofibroblats differentiation in the present study since we did not measure fibroblast ROS production in NOX4 siRNA treated cells. However, several experimental evidences strongly suggest that the effects we observed were mediated by ROS produced by NOX4. First, NOX4 was the only NOX homolog up regulated in IPF fibroblasts; second, ROS production was higher in these cells as compared to control cells, and third similar results in terms of α-SMA and procollagen I (α1) expression were obtained by incubating these cell with the antioxidant NAC and by transfecting them with a NOX4 siRNA. Furthermore, a recent study with controlled recombinant expression of NOX4 demonstrated a strong correlation between NOX4 mRNA level and ROS generation (30). Thus, in this experimental setting, the activity of NOX4 was demonstrated to depend directly on its gene expression levels. Moreover, we and others have previously shown that siRNA against NOX4 reduced basal ROS level in rat smooth muscle cells and in pulmonary epithelial cell lines significantly (14) (31). Collectively, these data strongly support a role of NOX4-produced ROS on myofibroblast differentiation.
Interestingly, immunohistochemistry showed a strong expression of NOX4 by hyperplastic alveolar epithelium in the fibrotic lung (fig 2). In the kidney, NOX4 has been involved in the regulation of epithelial-mesenchymal transition (32). Although not specifically studied, this phenomenon could also occur in the lung and implicate NOX4 in the fibrogenic process through multiple ways.
Role of NOX4 on TGF-β1 induced myofibroblasts differentiation and PDGF-induced migration
TGF-β1 stimulates differentiation of fibroblasts into myofibroblasts using Smad 2/3 signalling (20). After initial phosphorylation, Smad 2/3 activation in response to TGF-β1 is prolonged (hours to days depending on the cell type). Interestingly, we showed that Smad 2/3 phosphorylation was up-regulated by TGF-β1 after 18 h of incubation. The antioxidant N-acetylcysteine, the NOX inhibitor DPI, and the siRNA anti NOX4 lead to a marked inhibition of Smad2/3 phosphorylation, as demonstrated previously in human cardiac fibroblasts (11). These data imply that oxidants produced by NOX4 mediate α-SMA and procollagen I (α1) upregulation in pulmonary fibroblasts in response to TGF-β1 probably by stimulating phosphorylation and activation of Smads 2/3. In addition, NOX4 mediates PDGF-induced fibroblasts migration.
In summary, we have shown NOX4 upregulation in pulmonary fibroblasts from IPF patients, and identified a novel role for NOX4 as essential mediator of Smad2/3 transcription factor activation in response to TGF-β1 in pulmonary fibroblasts. Because of the critical role of TGF-β1–induced myofibroblast activation in IPF, these data provide insight into novel mechanisms with potential therapeutic implications for IPF.
Supplementary Material
Acknowledgments
We acknowledge Françoise Zerah (Assistance Publique – Hôpitaux de Paris, Service d’Explorations Fonctionnelles, Hôpital Henri Mondor, Créteil, France) for statistical advice.
FUNDING
Nadia Amara was supported by Chancellerie des Universités de Paris (legs Poix), and Jorge Boczkowski by INSERM and Assistance Publique-Hôpitaux de Paris (Contrat d’Interface). This work was supported by the European Commission (FP 7, European IPF Network) and by the Agence Nationale de la Recherche (ANR Physio 2006, FIBROPNEUMO).
Footnotes
Publisher's Disclaimer: The Corresponding Author has the right to grant on behalf of all authors and does grant on behalf of all authors, an exclusive licence (or non exclusive for government employees) on a worldwide basis to the BMJ Publishing Group Ltd and its Licensees to permit this article (if accepted) to be published in [THORAX] editions and any other BMJPG Ltd products to exploit all subsidiary rights, as set out in our licence
ETHICS APPROVAL
This study was approved by the local ethics committee of Saint Germain en Laye hospital (20 rue Amargis, 78100 Saint Germain en Laye, France) and stored biopsies were reported to our institutional board (Délégation à la Recherche Clinique, Assistance Publique-Hôpitaux de Paris, Carré Historique de l’Hôpital Saint- Louis, 1 avenue Claude Vellefaux 75010 Paris, France).
Competing Interest: None to declare.
References
- 1.Dempsey OJ. Clinical review: Idiopathic pulmonary fibrosis--past, present and future. Respir Med. 2006;100:1871–1885. doi: 10.1016/j.rmed.2006.08.017. [DOI] [PubMed] [Google Scholar]
- 2.Maher TM, Wells AU, Laurent GJ. Idiopathic pulmonary fibrosis: Multiple causes and multiple mechanisms? Eur Respir J. 2007;30:835–839. doi: 10.1183/09031936.00069307. [DOI] [PubMed] [Google Scholar]
- 3.Kuhn C, McDonald JA. The roles of the myofibroblast in idiopathic pulmonary fibrosis. Ultrastructural and immunohistochemical features of sites of active extracellular matrix synthesis. Am J Pathol. 1991;138:1257–1265. [PMC free article] [PubMed] [Google Scholar]
- 4.Zhang K, Gharaee-Kermani M, McGarry B, et al. In situ hybridization analysis of rat lung alpha 1(i) and alpha 2(i) collagen gene expression in pulmonary fibrosis induced by endotracheal bleomycin injection. Lab Invest. 1994;70:192–202. [PubMed] [Google Scholar]
- 5.Serini G, Bochaton-Piallat ML, Ropraz P, et al. The fibronectin domain ed-a is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1. J Cell Biol. 1998;142:873–881. doi: 10.1083/jcb.142.3.873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kinnula VL, Fattman CL, Tan RJ, et al. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am J Respir Crit Care Med. 2005;172:417–422. doi: 10.1164/rccm.200501-017PP. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yin L, Morita A, Tsuji T. Tobacco smoke extract induces age-related changes due to modulation of tgf-beta. Exp Dermatol. 2003;12 (Suppl 2):51–56. doi: 10.1034/j.1600-0625.12.s2.8.x. [DOI] [PubMed] [Google Scholar]
- 8.Amarnath S, Dong L, Li J, et al. Endogenous TGF-beta activation by reactive oxygen species is key to foxp3 induction in tcr-stimulated and HIV-1-infected human cd4+cd25- t cells. Retrovirology. 2007;4:57. doi: 10.1186/1742-4690-4-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol. 2004;4:181–189. doi: 10.1038/nri1312. [DOI] [PubMed] [Google Scholar]
- 10.Mittal M, Roth M, Konig P, et al. Hypoxia-dependent regulation of nonphagocytic Nadph oxidase subunit NOX4 in the pulmonary vasculature. Circ Res. 2007;101:258–267. doi: 10.1161/CIRCRESAHA.107.148015. [DOI] [PubMed] [Google Scholar]
- 11.Cucoranu I, Clempus R, Dikalova A, et al. Nad(p)h oxidase 4 mediates transforming growth factor-beta1-induced differentiation of cardiac fibroblasts into myofibroblasts. Circ Res. 2005;97:900–907. doi: 10.1161/01.RES.0000187457.24338.3D. [DOI] [PubMed] [Google Scholar]
- 12.Hinz B, Phan SH, Thannickal VJ, et al. The myofibroblast: One function, multiple origins. Am J Pathol. 2007;170:1807–1816. doi: 10.2353/ajpath.2007.070112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Marchand-Adam S, Marchal J, Cohen M, et al. Defect of hepatocyte growth factor secretion by fibroblasts in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2003;168:1156–1161. doi: 10.1164/rccm.200212-1514OC. [DOI] [PubMed] [Google Scholar]
- 14.Amara N, Bachoual R, Desmard M, et al. Diesel exhaust particles induce matrix metalloprotease-1 in human lung epithelial cells via a Nadp(h) oxidase/NOX4 redox-dependent mechanism. Am J Physiol Lung Cell Mol Physiol. 2007;293:L170–181. doi: 10.1152/ajplung.00445.2006. [DOI] [PubMed] [Google Scholar]
- 15.Thabut G, El-Benna J, Samb A, et al. Tumor necrosis factor-alpha increases airway smooth muscle oxidants production through a Nadph oxidase-like system to enhance myosin light chain phosphorylation and contractility. J Biol Chem. 2002;277:22814–22821. doi: 10.1074/jbc.M200315200. [DOI] [PubMed] [Google Scholar]
- 16.Goven D, Boutten A, Lecon-Malas V, et al. Altered Nrf2/keap1-bach1 equilibrium in pulmonary emphysema. Thorax. 2008;63:916–924. doi: 10.1136/thx.2007.091181. [DOI] [PubMed] [Google Scholar]
- 17.Quesnel C, Nardelli L, Piednoir P, et al. Alveolar fibroblasts in acute lung injury: Biological behaviour and clinical relevance. Eur Respir J. 2009 doi: 10.1183/09031936.00074709. [DOI] [PubMed] [Google Scholar]
- 18.Hancock JT, White JI, Jones OT, et al. The use of diphenylene iodonium and its analogues to investigate the role of the nadph oxidase in the tumoricidal activity of macrophages in vitro. Free Radic Biol Med. 1991;11:25–29. doi: 10.1016/0891-5849(91)90184-5. [DOI] [PubMed] [Google Scholar]
- 19.Evans RA, Tian YC, Steadman R, et al. TGF-beta1-mediated fibroblast-myofibroblast terminal differentiation-the role of smad proteins. Exp Cell Res. 2003;282:90–100. doi: 10.1016/s0014-4827(02)00015-0. [DOI] [PubMed] [Google Scholar]
- 20.Massague J. How cells read tgf-beta signals. Nat Rev Mol Cell Biol. 2000;1:169–178. doi: 10.1038/35043051. [DOI] [PubMed] [Google Scholar]
- 21.Hecker L, Vittal R, Jones T, et al. Nadph oxidase-4 mediates myofibroblast activation and fibrogenic responses to lung injury. Nat Med. 2009;15:1077–1081. doi: 10.1038/nm.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li S, Tabar SS, Malec V, et al. NOX4 regulates ros levels under normoxic and hypoxic conditions, triggers proliferation, and inhibits apoptosis in pulmonary artery adventitial fibroblasts. Antioxid Redox Signal. 2008 doi: 10.1089/ars.2008.2035. [DOI] [PubMed] [Google Scholar]
- 23.Sturrock A, Cahill B, Norman K, et al. Transforming growth factor-beta1 induces nox4 nad(p)h oxidase and reactive oxygen species-dependent proliferation in human pulmonary artery smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2006;290:L661–L673. doi: 10.1152/ajplung.00269.2005. [DOI] [PubMed] [Google Scholar]
- 24.Ambasta RK, Kumar P, Griendling KK, et al. Direct interaction of the novel NOX proteins with p22phox is required for the formation of a functionally active nadph oxidase. J Biol Chem. 2004;279:45935–45941. doi: 10.1074/jbc.M406486200. [DOI] [PubMed] [Google Scholar]
- 25.Kawahara T, Ritsick D, Cheng G, et al. Point mutations in the proline-rich region of p22phox are dominant inhibitors of nox1- and NOX2-dependent reactive oxygen generation. J Biol Chem. 2005;280:31859–31869. doi: 10.1074/jbc.M501882200. [DOI] [PubMed] [Google Scholar]
- 26.Hu T, Ramachandrarao SP, Siva S, et al. Reactive oxygen species production via Nadph oxidase mediates tgf-beta-induced cytoskeletal alterations in endothelial cells. Am J Physiol Renal Physiol. 2005;289:F816–825. doi: 10.1152/ajprenal.00024.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sturrock A, Huecksteadt TP, Norman K, et al. NOX4 mediates tgf-beta1-induced retinoblastoma protein phosphorylation, proliferation, and hypertrophy in human airway smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2007;292:L1543–1555. doi: 10.1152/ajplung.00430.2006. [DOI] [PubMed] [Google Scholar]
- 28.Ramos C, Montano M, Garcia-Alvarez J, et al. Fibroblasts from idiopathic pulmonary fibrosis and normal lungs differ in growth rate, apoptosis, and tissue inhibitor of metalloproteinases expression. Am J Respir Cell Mol Biol. 2001;24:591–598. doi: 10.1165/ajrcmb.24.5.4333. [DOI] [PubMed] [Google Scholar]
- 29.Khalil N, O’Connor RN, Flanders KC, et al. TGF-beta 1, but not TGF-beta 2 or TGF-beta 3, is differentially present in epithelial cells of advanced pulmonary fibrosis: An immunohistochemical study. Am J Respir Cell Mol Biol. 1996;14:131–138. doi: 10.1165/ajrcmb.14.2.8630262. [DOI] [PubMed] [Google Scholar]
- 30.Serrander L, Cartier L, Bedard K, et al. NOX4 activity is determined by mrna levels and reveals a unique pattern of ROS generation. Biochem J. 2007;406:105–114. doi: 10.1042/BJ20061903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schroder K, Helmcke I, Palfi K, et al. Nox1 mediates basic fibroblast growth factor-induced migration of vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2007;27:1736–1743. doi: 10.1161/ATVBAHA.107.142117. [DOI] [PubMed] [Google Scholar]
- 32.Rhyu DY, Yang Y, Ha H, et al. Role of reactive oxygen species in TGF-beta1-induced mitogen-activated protein kinase activation and epithelial-mesenchymal transition in renal tubular epithelial cells. J Am Soc Nephrol. 2005;16:667–675. doi: 10.1681/ASN.2004050425. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.