

Abstract
Alzheimer’s disease (AD) is characterized by the self-assembly of amyloid beta (Aβ) peptides. Recent models implicate some of the earliest Aβ oligomers, such as trimers and tetramers, in disease. However, the roles of these structures remain uncertain, in part, because selective probes of their formation are not available. Towards that goal, we generated bivalent versions of the known Aβ ligand, the pentapeptide KLVFF. We found that compounds containing sufficiently long linkers (~19 to 24 Å) recognized primarily Aβ trimers and tetramers, with little binding to either monomer or higher order structures. These compounds might be useful probes for early Aβ oligomers.
Both in vitro and in vivo, the Aβ monomer will self-assemble into higher order structures, including dimers, trimers, tetramers and larger oligomers. Eventually, this peptide will form the elongated fibrils that are observed in late-stage AD patients (Figure 1a). Recent studies have suggested that the accumulation of smaller aggregates, not the large fibrils, might better correlate with neurotoxicity.1 For example, patient-derived Aβ dimers and trimers have been shown to inhibit long-term potentiation and damage synaptic plasticity.2 Similarly, synthetic Aβ trimers and tetramers are two-fold more toxic to cultured neurons than either Aβ monomers, dimers or fibrils.3 Together, these studies suggest that some of the earliest structures in the aggregation pathway might be the most diagnostic of disease.
Figure 1.

Design of KLVFF-based probes. (a) Schematic of the Aβ aggregation pathway, highlighting the earliest structures. (b) The sequence of Aβ (1–40), including the core KLVFF motif. (c) Proposed features of a bivalent, KLVFF-based probe. The Aβ is shown in grey, the KLVFF in purple, the linker in red and the biotin tag in green. (d) Snapshot from a molecular dynamics simulation of a KLVFF-based probe bound to a model Aβ trimer. Based on these simulations, an approximate distance between KLVFF sites was estimated.
The Aβ peptide is a 40 or 42 amino acid fragment of the amyloid precursor protein (APP) (Figure 1b). Portions of this peptide are thought to form β-sheets upon release from APP and subsequent stacking of these region appears to nucleate Aβ self-association.4 Within these sites, residues 16 through 20, KLVFF, are especially important.5 Specifically, this motif is thought to interact with itself in adjacent β-strands, with the phenylalanine residues forming key, repetitive inter-molecular contacts.6 The interactions of KLVFF with itself has been studied extensively and these studies have suggested that many Aβ structures have an exposed KLVFF motif at each “end”.7 Consistent with this model, KLVFF-based peptides will inhibit Aβ aggregation at high concentrations, presumably by blocking these sites.6e However, KLVFF binds only weakly (Kd > 1 mM)8 and multivalent displays have been found to be required for potent inhibition.9 Multivalent binding is known to significantly enhance avidity and selectivity in many systems, by elevating local ligand concentration, favoring multi-site binding and other mechanisms. Based on these observations, we envisioned that bridging two KLVFF peptides with a linker of the appropriate length might provide a probe for the earliest Aβ oligomers (Figure 1c). This strategy was designed to address a central challenge in building probes that are specific for a subset of Aβ structures. Namely, these oligomers are assembled from identical monomer units and; therefore, they contain many degenerate molecular features, such as high beta-sheet content. By exploiting one of their few distinguishing properties (e.g. end-to-end distance between KLVFF motifs), we hoped to circumvent these issues.
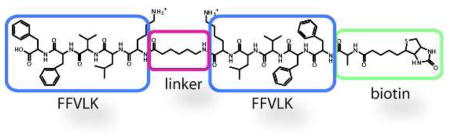
To estimate the minimal distance needed to span the ends of an early Aβ aggregate, we assembled a representative KLVFF-based probe in silico and then employed molecular dynamics (MD) simulations to examine its binding to a model Aβ repeating unit.7f These studies roughly estimated the distance between KLVFF sites as 13–15 Å in a dimer, 19–20 Å in a trimer, and approximately 24–25 Å in a tetramer (Figure 1d and Supplemental Figure 1). Using microwave-assisted, solid-phase peptide coupling, we then constructed a control compound in which the KLVFF peptide was linked to biotin (Figure 2a). Similarly, we generated four molecules (d7, d13, d19, and d24), each containing two KLVFF motifs separated by a variable number of aminohexanoic acid (Ahx) units and a biotin at the N-terminus (Figure 2b). These compounds were named according to the approximate length of the extended linker (e.g. d7 has an estimated linker length of ~ 7 Å).11 We found that these probes were soluble and non-aggregating in aqueous solution at low concentrations (below 10 μM).
Figure 2.

KLVFF-based probes selectively bind Aβ trimers and tetramers. (a) Chemical structure of the monovalent, KLVFF-biotin probe. (b) Chemical structures and binding properties of bivalent probes. Aβ (1–40) was cross-linked as described, separated using electrophoresis, and transferred to nitrocellulose. Compounds (2 μM) were incubated with the membranes, which were then washed and imaged with streptavidin-HRP. (c) The Aβ band intensities are plotted against the maximum linker length. Results are representative of four independent experiments.
To test binding of these probes, we employed an established, UV cross-linking approach to produce Aβ samples containing a mixture of small oligomers.12 Briefly, 25 μM Aβ (1–40) was cross-linked with a Ru(II) catalyst and excess catalyst was removed.13 The resulting “ladder” was separated by native-gel electrophoresis. By silver staining, we observed approximately equal levels of monomer, dimer, and trimer, along with lesser quantities of tetramer and pentamer (Figure 2b). We then transferred these samples to nitrocellulose, incubated with the KLVFF probes (2 μM) and washed extensively. The bound material was localized using streptavidin coupled to horseradish peroxidase (HRP).
Under these conditions, we observed no binding by the KLVFF-biotin control (Figure 2b), a result consistent with its weak affinity.8 Similarly, the KLVFF-based probes with relatively short linkers, d7 and d13, also had weak binding, with a faint band at the molecular weight of an Aβ trimer (Figure 2b and 2c). However, the compounds with longer linkers, d19 and d24, interacted strongly with the trimer and tetramer regions, with some binding to the dimers and pentamers (Figure 2b and 2c). The relatively poor binding to the Aβ dimers might suggest that it is not as ordered as the other structures, a concept that is consistent with recent MD and NMR studies.6g,7d Importantly, Aβ monomer was not recognized by any of the ligands at these concentrations, supporting an important role for multivalent interactions.
Based upon the results using crosslinked Aβ samples, we wanted to further test binding in more dynamic mixtures. Towards that goal, we used ligand d24 to probe aged samples of non-crosslinked Aβ (1–42). Samples prepared by this method are known to contain a mixture of monomer (4.5 kDa), poorly resolved trimers and tetramers (~ 12 to 18 kDa) and higher order oligomers (~40 to 200 kDa).14 Consistent with those patterns, we observed Aβ structures of these sizes by either silver staining or Western blots with the anti-Aβ antibody 6E10 (Figure 3a). The KLVFF-biotin control did not recognize any of the bands under these conditions. However, d24 remained bound to the region corresponding to trimer and tetramer, suggesting that selectivity is maintained (Figure 3a). Minimal binding to monomer was seen and, importantly, we did not observe any interactions with the higher molecular weight oligomers, further emphasizing the selectivity of these probes for early structures in the Aβ aggregation pathway.
Figure 3.

Ligand d24 selectively recognize trimers and tetramers in mixed Aβ samples and human cerebrospinal fluid (CSF). (a) Aβ (1–42) was separated and probed as described above. Silver staining or probing with the anti-Aβ antibody 6E10 showed a mixture of monomers, trimers, tetramers, and high-molecular weight oligomers, but d24 bound predominantly to trimers and tetramers. (b) Cross-linked Aβ (1–40) was added to human CSF and probed with d24. Results are representative of at least two independent replicates.
Finally, we wanted to evaluate binding in human cerebrospinal fluid (CSF), which provides a more challenging environment than aqueous buffers in which to retain binding. To determine if d24 could still bind Aβ in this milieu, we added 1 μg of cross-linked Aβ (1–40) to CSF samples from non-AD patients (6 μg of total protein) and characterized the resulting mixture by silver stain, 6E10 antibody and d24. From these studies, we concluded that d24 recognized several unrelated bands within the CSF sample; however, this off-target reactivity was largely restricted to proteins > 50 kDa. Thus, we were still able to visualize binding to Aβ trimers and tetramers (Figure 3b). Similar findings were obtained using Aβ (1–42) (Supplemental Figure 2). Together, these results suggest that the multivalent probes can exploit unique inter-KLVFF distances to distinguish between otherwise closely related Aβ structures. Based on these findings, we anticipate that derivatives of d24, with further improvements in affinity and selectivity, may be promising probes for detecting the appearance of the earliest Aβ aggregates.
Supplementary Material
Acknowledgments
A.A.R. was supported by a fellowship from the Biogerontology NIA Training Grant (AG000114). This work was also supported by a grant from the Alzheimer’s Association (NIRG-08-89471) to J.E.G. The authors thank C. Evans and P. Marinec for assisting with ligand characterization, M. Wallace for helpful discussions, and D. Giacherio of the UM Hospital for CSF samples.
Footnotes
Supporting Information Available: Detailed protocols for ligand synthesis and characterization, Aβ preparation and crosslinking, Western blots and MD simulations. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.(a) Caughey B, Lansbury PT. Annu Rev Neurosci. 2003;26:267–98. doi: 10.1146/annurev.neuro.26.010302.081142. [DOI] [PubMed] [Google Scholar]; (b) Hung LW, Ciccotosto GD, Giannakis E, Tew DJ, Perez K, Masters CL, Cappai R, Wade JD, Barnham KJ. J Neurosci. 2008;28:11950–8. doi: 10.1523/JNEUROSCI.3916-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.(a) Shankar GM, Bloodgood BL, Townsend M, Walsh DM, Selkoe DJ, Sabatini BL. J Neurosci. 2007;27:2866–75. doi: 10.1523/JNEUROSCI.4970-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, Smith I, Brett FM, Farrell MA, Rowan MJ, Lemere CA, Regan CM, Walsh DM, Sabatini BL, Selkoe DJ. Nat Med. 2008;14:837–42. doi: 10.1038/nm1782. [DOI] [PMC free article] [PubMed] [Google Scholar]; (c) Townsend M, Shankar GM, Mehta T, Walsh DM, Selkoe DJ. J Physiol. 2006;572:477–92. doi: 10.1113/jphysiol.2005.103754. [DOI] [PMC free article] [PubMed] [Google Scholar]; (d) O’Nuallain B, Freir DB, Nicoll AJ, Risse E, Ferguson N, Herron CE, Collinge J, Walsh DM. J Neurosci. 2010;30:14411–19. doi: 10.1523/JNEUROSCI.3537-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ono K, Condron MM, Teplow DB. Proc Natl Acad Sci U S A. 2009;106:14745–50. doi: 10.1073/pnas.0905127106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.(a) Aguzzi A, O’Connor T. Nat Rev Drug Discov. 2010;9:237–48. doi: 10.1038/nrd3050. [DOI] [PubMed] [Google Scholar]; (b) Roychaudhuri R, Yang M, Hoshi MM, Teplow DB. J Biol Chem. 2009;284:4749–53. doi: 10.1074/jbc.R800036200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Esler WP, Stimson ER, Ghilardi JR, Lu YA, Felix AM, Vinters HV, Mantyh PW, Lee JP, Maggio JE. Biochemistry. 1996;35:13914–21. doi: 10.1021/bi961302+. [DOI] [PubMed] [Google Scholar]
- 6.(a) Ahmed M, Davis J, Aucoin D, Sato T, Ahuja S, Aimoto S, Elliott JI, Van Nostrand WE, Smith SO. Nat Struct Mol Biol. 2010;17:561–7. doi: 10.1038/nsmb.1799. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Bieschke J, Siegel SJ, Fu Y, Kelly JW. Biochemistry. 2008;47:50–9. doi: 10.1021/bi701757v. [DOI] [PubMed] [Google Scholar]; (b) Bowerman CJ, Ryan DM, Nissan DA, Nilsson BL. Mol Biosyst. 2009;5:1058–69. doi: 10.1039/b904439f. [DOI] [PubMed] [Google Scholar]; (c) Chini MG, Scrima M, D’Ursi AM, Bifulco G. J Pept Sci. 2009;15:229–34. doi: 10.1002/psc.1095. [DOI] [PubMed] [Google Scholar]; (d) Hilbich C, Kisters-Woike B, Reed J, Masters CL, Beyreuther K. J Mol Biol. 1992;228:460–73. doi: 10.1016/0022-2836(92)90835-8. [DOI] [PubMed] [Google Scholar]; (e) Tjernberg LO, Naslund J, Lindqvist F, Johansson J, Karlstrom AR, Thyberg J, Terenius L, Nordstedt C. J Biol Chem. 1996;271:8545–8. doi: 10.1074/jbc.271.15.8545. [DOI] [PubMed] [Google Scholar]; (f) Wood SJ, Wetzel R, Martin JD, Hurle MR. Biochemistry. 1995;34:724–30. doi: 10.1021/bi00003a003. [DOI] [PubMed] [Google Scholar]; (g) Yu L, Edalji R, Harlan JE, Holzman TF, Lopez AP, Labkovsky B, Hillen H, Barghorn S, Ebert U, Richardson PL, Miesbauer L, Solomon L, Bartley D, Walter K, Johnson RW, Hajduk PJ, Olejniczak ET. Biochemistry. 2009;48:1870–7. doi: 10.1021/bi802046n. [DOI] [PubMed] [Google Scholar]
- 7.(a) Antzutkin ON, Balbach JJ, Leapman RD, Rizzo NW, Reed J, Tycko R. Proc Natl Acad Sci U S A. 2000;97:13045–50. doi: 10.1073/pnas.230315097. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Costa PR, Kocisko DA, Sun BQ, Lansbury PT, Griffin RG. J Am Chem Soc. 1997;119:10487–10493. [Google Scholar]; (c) Hetenyi C, Kortvelyesi T, Penke B. Bioorg Med Chem. 2002;10:1587–93. doi: 10.1016/s0968-0896(01)00424-2. [DOI] [PubMed] [Google Scholar]; (d) Horn AH, Sticht H. J Phys Chem B. 2010;114:2219–26. doi: 10.1021/jp100023q. [DOI] [PubMed] [Google Scholar]; (e) Hwang W, Zhang S, Kamm RD, Karplus M. Proc Natl Acad Sci U S A. 2004;101:12916–21. doi: 10.1073/pnas.0402634101. [DOI] [PMC free article] [PubMed] [Google Scholar]; (f) Luhrs T, Ritter C, Adrian M, Riek-Loher D, Bohrmann B, Dobeli H, Schubert D, Riek R. Proc Natl Acad Sci U S A. 2005;102:17342–7. doi: 10.1073/pnas.0506723102. [DOI] [PMC free article] [PubMed] [Google Scholar]; (g) Wallace JA, Shen JK. Biochemistry. 2010;49:5290–8. doi: 10.1021/bi100137y. [DOI] [PubMed] [Google Scholar]; (h) Hu Y, Su B, Kim C-S, Hernandez M, Rostagno A, Ghiso J, Kim JR. Chembiochem. 2010 doi: 10.1002/cbic.201000435. in press. [DOI] [PubMed] [Google Scholar]
- 8.Cairo CW, Strzelec A, Murphy RM, Kiessling LL. Biochemistry. 2002;41:8620–9. doi: 10.1021/bi0156254. [DOI] [PubMed] [Google Scholar]
- 9.(a) Chafekar SM, Malda H, Merkx M, Meijer EW, Viertl D, Lashuel HA, Baas F, Scheper W. Chembiochem. 2007;8:1857–64. doi: 10.1002/cbic.200700338. [DOI] [PubMed] [Google Scholar]; (b) Lowe TL, Strzelec A, Kiessling LL, Murphy RM. Biochemistry. 2001;40:7882–9. doi: 10.1021/bi002734u. [DOI] [PubMed] [Google Scholar]; (c) Watanabe K, Nakamura K, Akikusa S, Okada T, Kodaka M, Konakahara T, Okuno H. Biochem Biophys Res Commun. 2002;290:121–4. doi: 10.1006/bbrc.2001.6191. [DOI] [PubMed] [Google Scholar]; (d) Zhang G, Leibowitz MJ, Sinko PJ, Stein S. Bioconjug Chem. 2003;14:86–92. doi: 10.1021/bc025526i. [DOI] [PubMed] [Google Scholar]
- 10.Kiessling LL, Gestwicki JE, Strong LE. Angew Chem Int Ed Engl. 2006;45:2348–68. doi: 10.1002/anie.200502794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Thomas LL, Christakis TJ, Jorgensen WL. J Phys Chem B. 2006;110:21198–204. doi: 10.1021/jp064811m. [DOI] [PubMed] [Google Scholar]
- 12.Bitan G, Teplow DB. Acc Chem Res. 2004;37:357–64. doi: 10.1021/ar000214l. [DOI] [PubMed] [Google Scholar]
- 13.Marinec PS, Evans CG, Gibbons GS, Tarnowski MA, Overbeek DL, Gestwicki JE. Bioorg Med Chem. 2009;17:5763–8. doi: 10.1016/j.bmc.2009.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cerf E, Sarroukh R, Tamamizu-Kato S, Breydo L, Derclaye S, Dufrene YF, Narayanaswami V, Goormaghtigh E, Ruysschaert JM, Raussens V. Biochem J. 2009;421:415–23. doi: 10.1042/BJ20090379. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.