

Summary
We recently reported that differentiation of CD8+ T cells from the naïve to the effector state involves the upregulation of glucose-dependent metabolism. Glucose deprivation or inhibition of glycolysis by 2-deoxy-D-glucose (2-DG) selectively inhibited production of IFN-γ but not of IL-2. To determine a more global role of glucose metabolism on effector T cell function, we performed gene array analysis on CD8+ effector T cells stimulated in the presence or absence of 2-DG. We observed that expression of only 10% of genes induced by TCR/CD28 signaling was inhibited by 2-DG. Among these were genes for key cytokines, cell cycle molecules, and cytotoxic granule proteins. Consistent with these results, production of IFN-γ and GM-CSF, cell cycle progression, upregulation of cyclin D2 protein, cytolytic activity, and upregulation of granzyme B protein but also conjugate formation were exquisitely glucose-dependent. In contrast to glucose, oxygen was little utilized by CD8+ effector T cells, and relative oxygen deprivation did not inhibit these CTL functional properties. Our results indicate a particularly critical role for glucose in regulating specific effector functions of CD8+ T cells, and have implications for the maintenance of the effector phase of cellular immune responses in target tissue microenvironments such as a solid tumor.
Keywords: T Cells, Cytotoxicity, Gene Regulation, Cellular Activation
Introduction
Mature CD8+ effector T cells recognize foreign peptides on MHC class I molecules displayed on virally-infected or cancerous cells. During the effector phase of the immune response, lytic granules are released in a directional manner, causing rapid death of target cells. In addition, effector cytokines and chemokines are secreted, regulating and recruiting other immune cells to the site. Finally, CD8+ effector T cells can undergo additional rounds of clonal expansion to amplify the immune response and effectively eradicate antigen-expressing cells. Execution of these effector functions requires increases in metabolic substrates, cytoskeletal changes, and cell division which come at an energy cost for the cell.
Compared to naïve cells, differentiated CD8+ effector T cells display increased glucose-dependent metabolism, including augmented expression of glycolytic proteins and greater glycolytic rate [1]. Given that effector cells need to be poised for rapid activation, we reasoned that the maintenance and execution of effector functions may require a certain amount of ATP. Furthermore, the effectiveness of an anti-viral or anti-tumor immune response may depend on the amount of energy substrates available in the surrounding microenvironment. Previously, we showed that maintenance of IFN-γ gene expression depended on glucose availability [1]. Earlier studies have also suggested a glucose requirement for cytolytic activity [2], arguing that glucose specifically might be required for critical effector functions.
ATP can be synthesized via two distinct processes of respiration, anaerobic (O2-independent) or aerobic (O2-dependent). Glycolysis is the anaerobic mechanism by which one molecule of glucose is converted into two molecules of pyruvate. Two molecules of ATP are generated per molecule of glucose. In the absence of O2, two additional molecules of ATP can be generated by the conversion of pyruvate into lactate by lactate dehydrogenase. By contrast, aerobic respiration produces 36 molecules of ATP per molecule of glucose. Although more energy efficient, the oxidative phosphorylation step of aerobic respiration produces negative byproducts such as reactive oxygen species, and it is thought that for this reason, among others, that some cell systems preferentially utilize the less energy efficient anaerobic glucose metabolic pathway to provide cellular ATP. The role of cell metabolism in T cell responsiveness to TCR/CD28 ligation has recently been reviewed [3]. This work suggests that regulation of nutrient utilization, including glucose handling, is a critical component of T cell activation at multiple levels.
Based on these considerations, we examined in more detail the specific requirements for glucose versus oxygen in specific CD8+ gene expression events and effector functions. We observed that 10% of inducible transcripts were glucose-dependent. Based on the nature of these genes, we went on to show that effector cytokine production, cytolysis, and proliferation were T cell functions inhibited by 2-deoxy-D-glucose (2-DG). In contrast, oxygen deprivation had minimal affects on IFN-γ production and cytolysis, and did not cause a decrease in steady-state ATP levels in CD8+ effector T cells. Taken together, our data suggest that glucose may be the most critical source of energy for CD8+ effector T cells to carry out critical immune functions in target tissue microenvironments.
Results
Expression of only a subset of induced genes is inhibited by 2-DG
Previously, we showed that production of the effector cytokine IFN-γ, but not of IL-2, was exquisitely sensitive to glucose-deprivation or addition of 2-deoxy-D-glucose (2-DG) [1]. This observation suggested that a subset of inducible genes in CD8+ effector cells might be preferentially glucose-dependent. To test this notion directly, we utilized Affymetrix arrays containing approximately 35,000 oligonucleotides to examine the expression profiles of 2C CD8+ effector T cells stimulated with or without 2-DG for 12 hours. We found that 1194 transcripts were inducibly upregulated upon anti-CD3 and anti-CD28 stimulation. Of these, expression of only 10% (124 of 1194) was inhibited by 2-DG. This gene set seemed to be enriched for transcripts associated with T cell effector functions (Supplemental Table 1). As shown in Figures 1A and 1B, mRNA levels of several effector cytokine genes were diminished in cells stimulated with 2-DG. IFN-γ expression was inhibited as we had shown previously. But in addition, GM-CSF and several other cytokines also were suppressed by 2-DG. However, we found that transcripts for the chemokine MIP-1β and the cytokine IL-2 remained similar after stimulation despite the presence of 2-DG (Figure 1C). We also observed the downregulation of genes involved in cytolysis (Perforin and Granzyme C, Figure 1D) and proliferation (Cyclin D2, Figure 1E). Of note, the expression of several genes, such as TCRβ and γ-tubulin, remained unchanged in all three states (Figure 1F). These results argue that glucose is required for expression of a selected subset of genes in CD8+ T cells. The categories of genes affected suggest that several key T cell effector functions might be particularly sensitive to glucose deprivation.
Figure 1. Gene expression analyzed using Affymetrix chips representing 35,000 oligonucleotides.
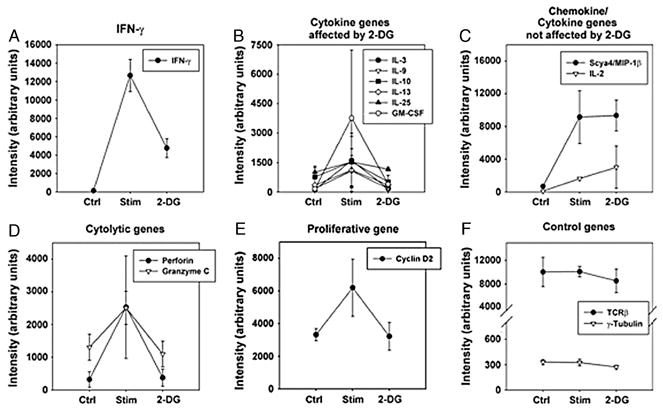
2-DG affects only a subset of genes in stimulated CD8+ effector T cells. CD8+ effector T cells were stimulated for 12 hours with anti-CD3 and anti-CD28 mAb-coated beads in the presence or absence of 50 mM 2-DG. Control (Ctrl), unstimulated; stimulated (Stim); stimulated in the presence of 50 mM 2-DG (2-DG). Genes affected by 2-DG treatment include IFN-γ (A), several cytokines including GM-CSF, IL-3, IL-9, IL-10 (B) but not MIP-1β and IL-2 (C). Cytolytic and proliferative genes like perforin, granzyme B (D) and cyclin D2 (E) were also highly affected by 2-DG treatment as compared to unmodulated control genes like γ-tubulin and TCR-β (F). Individual hybridizations were normalized using dChip software, then averaged. Data represent the average + SD of two independent experiments.
Effector cytokine production is glucose-dependent
As confirmatory assays of our gene array data, real-time RT-PCR analysis was performed for a subset of cytokine genes. Consistent with the gene expression profiling, induction of transcripts encoding IFN-γ and GM-CSF was blocked by 2-DG as detected by real-time RT-PCR (Figures 2B and 2D). Similarly, secretion of IFN-γ and GM-CSF protein was nearly completely blocked upon inhibition of glucose metabolism (Figures 2A and 2C). In contrast, as we reported previously [1], IL-2 production was relatively resistant to inhibition by 2-DG and TNF-α secretion also was blocked by 2-DG (data not shown). These data indicate that key effector cytokines are exquisitely dependent on glucose for their production.
Figure 2. Effector cytokine production, proliferation, and cytolytic activity are glucose-dependent.
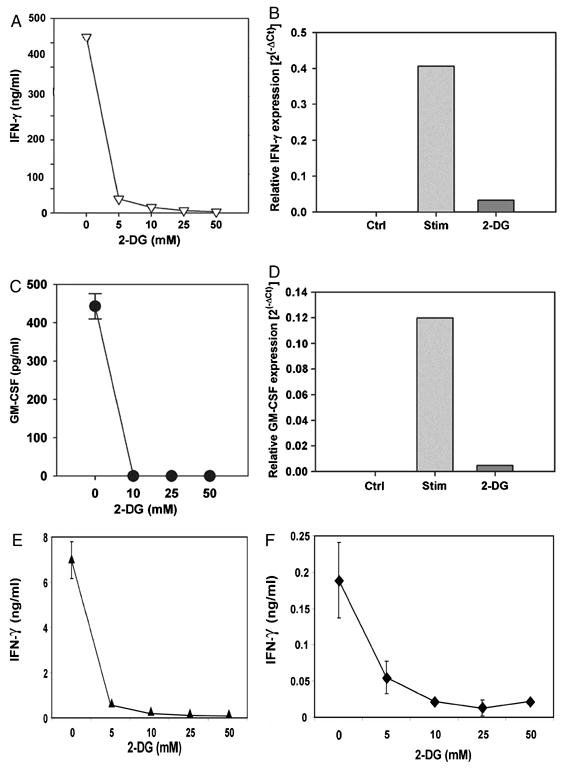
A. IFN-γ production by CD8+ effector T cells following anti-CD3 and anti-CD28 stimulation were measured by ELISA. Supernatant was collected after 24 hours of stimulation. Data represent the average + SD of three independent experiments and the 2-DG effect was statistically significant (p<0.01). B. Quantitative RT-PCR of IFN-γ mRNA in cells that were treated with 50 mM 2-DG for 12 hours. C. GM-CSF production by CD8+ effector T cells following stimulation with anti-CD3 and anti-CD28 mAb-coated beads was measured by ELISA. Supernatant was collected after 24 hours of stimulation. Data represent the average + SD of three independent experiments and the inhibition by 2-DG was statistically significant at all concentrations (p<0.001). D. Quantitative RT-PCR of GM-CSF mRNA in cells that were treated with 50 mM 2-DG for 12 hours. Data are representative of at least two independent experiments. E. IFN-γ production by CD8+ T cells primed in vivo with OVA peptide was measured by ELISA. Supernatant was collected after 20 hours of in vitro re-stimulation with OVA peptide. The 2-DG effect on IFN-γ was statistically significant (p<0.001) for all the concentrations tested. F. IFN-γ production by CD8+ T cells primed in vivo with OVA peptide was measured by ELISA. Supernatant was collected after 20 hours of stimulation with anti-CD3 and anti-CD28 mAb-coated beads. The 2-DG effect on IFN-γ was statistically significant (p<0.001) for all the concentrations tested. Data for E and F represent the average + SD of two independent experiments.
In order to confirm the importance of glucose metabolism for effector functions of T cells primed in vivo, mice were immunized with ovalbumin protein in complete Freund's adjuvant. T cells from lymph nodes were recovered and activated overnight, stimulated with ovalbumin in presence or absence of 2-DG, and tested for IFN-γ secretion. Like with in vitro primed 2C cells, 2-DG strongly inhibited IFN-γ secretion by in vivo primed polyclonal T cells (Figure 2E). To be certain whether the same effect would be observed when the only cells in the culture that could be affected by 2-DG were the T cells, stimulation was also performed without APCs but instead with anti-CD3/anti-CD28 mAb-coated beads. IFN-γ production was similarly inhibited by 2-DG under these conditions (Figure 2F).
Cytolytic activity of CD8+ effector T cells is glucose-dependent
Our gene array data indicated reduced expression of perforin and granzyme C mRNA in effector cells stimulated in the presence of 2-DG, suggesting that cytolysis might also be glucose-dependent. The reduction in these transcripts with 2-DG was confirmed by real-time RT-PCR (Figure 3A). To examine the glucose-dependence of cytolysis, primed 2C/RAG2-/- CD8+ T cells were used as effectors, which recognize Ld expressed by P815 targets. The addition of 50 mM 2-DG nearly completely abolished the ability of CD8+ effector T cells to lyse targets in a 4-hour 51Cr-release assay (Figure 3B). We showed previously that T cell death by apoptosis is not increased by 2-DG for up to 20 hours [1], arguing that this reduced cytolytic activity is not explained by a reduction in T cell viability.
Figure 3. 2-DG blocks cytolytic activity in CD8+ effector T cells.
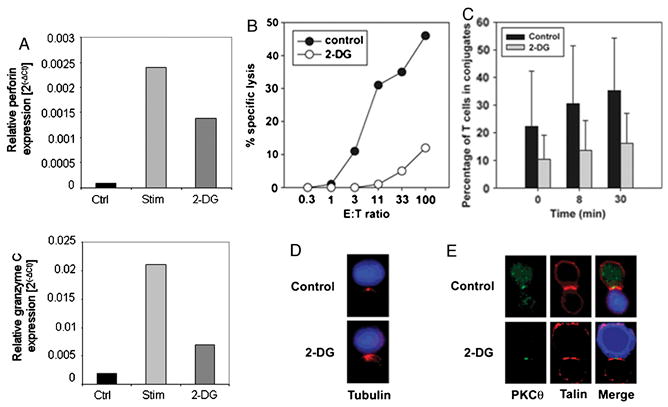
A. Effect of 50mM 2-DG on induction of perforin and granzyme C mRNA levels, as assessed by quantitative RT-PCR. Data represent average of two independent assays. B. Cytolytic activity of CD8+ effector T cells was determined in a 4-hour 51Cr-release assay, in the presence or absence of 50 mM 2-DG. Data represent the average of five independent experiments. Inhibition by 2-DG was statistically significant at each E:T ratio (P<0.01). C. Conjugate formation was assessed between 2C and P815-B7.1 targets, in the presence or absence of 50mM 2-DG. Calcein AM-labeled T cells (green) were combined with PKH-labeled P815-B7.1 cells (red) for indicated periods of time. Flow cytometry was used to determine the percentage of conjugates out of the total number of cells. Data represent the average + SD of three pooled experiments. D. MTOC reorientation in T cell/target cell conjugates in the presence or absence of 50mM 2-DG, as measured by tubulin staining (red). Data shown is a representative image of at least 20 conjugates analyzed. E. Formation of the cSMAC in T cell/target cell conjugates. Antibodies against PKCθ (green) and talin (red) were used. Target cells were stained with CMAC (blue). Images are representative of at least 20 conjugates analyzed.
Although we observed a reduction in gene expression of perforin and multiple granzyme isoforms in cells stimulated in 2-DG for 12 hours, we reasoned that reduced gene expression could not completely account for the dramatic reduction in a 4-hour cytolytic assay. Differentiated CD8+ effector T cells already express mature cytotoxic granules in lysosome-like compartments [4]. Upon engagement of target effector cells, these lytic granules rapidly polarize toward the target cell and are then secreted into the intermembrane space between the cells. Thus, we examined other features of the lytic process to examine their glucose dependence. The establishment of conjugate formation between CD8+ effector T cells and targets is a complex process of adhesion that could require vigorous ATP generation. We therefore examined conjugate formation using a flow cytometry-based assay in the presence or absence of 2-DG. As shown in Figure 3C, effector cells formed readily detectable conjugates detected at the earliest assayable time point. Indeed, the presence of 2-DG reduced the efficiency of conjugation by approximately 50%, arguing that CTL adhesion to target cells is glucose-dependent.
An additional step in the lytic process is the polarization of the cytolytic machinery toward the target cell. This cellular polarization is reflected by movement of the microtubule organizing center (MTOC). Using confocal microscopy, we tested whether CD8+ effectors that do successfully form conjugates in the presence of 2-DG could reorganize the MTOC. As shown in figure 3D, 2-DG did not inhibit MTOC reorientation, which was still observed in the vast majority of conjugates. In keeping with the process of proper polarization of the cell, we also examined establishment of features of the immunologic synapse in the presence or absence of 2-DG. The immunological synapse can be characterized by accumulation of talin along the T cell/target cell interface. In addition, the mature synapse in many systems includes central clearing of talin and partitioning of signaling molecules such as PKC-θ in the central supramolecular activation cluster (cSMAC) [5]. We therefore used fluorescence microscopy to determine if a mature immunological synapse could be formed in T cells that successfully formed conjugates with target cells, in the presence of 2-DG. In fact, recruitment of talin to the interface was readily detected, as was focusing of PKCθ to the cSMAC (Figure 3E). These results are consistent with the fact that glucose deprivation does not inhibit early TCR-mediated signaling events [1]. Collectively, these data suggest that 2-DG inhibits cytolytic activity both by blocking conjugate formation and by inhibiting induction of new granule proteins.
T cell proliferation is glucose-dependent
Results from our gene array experiments also indicated that glucose utilization was required for induction of cyclin D2 mRNA, suggesting that cell cycle progression might be glucose-dependent. Real-time RT-PCR confirmed that less cyclin D2 was induced in effector cells stimulated with 50 mM 2-DG (Figure 4A). We therefore analyzed effector cells for [3H]thymidine incorporation after stimulation for 48 hours with mitomycin C-treated P815-B7.1 cells in the presence of increasing concentrations of 2-DG. We observed a profound inhibitory effect of 2-DG on [3H]thymidine incorporation (Figure 4B). To examine directly blockade of thymidine incorporation was reflected at the level of cell division, CFSE dilution was examined. Primed 2C/RAG2-/- CD8+ effector T cells were labeled with CFSE and stimulated with P815-B7.1 cells for 48 hours. As shown in Figure 4C, the normally brisk cell division was completely abrogated in the presence of 2-DG. To determine which phase of the cell cycle is blocked by 2-DG, we examined the DNA content of stimulated effector cells by PI staining (Nicoletti assay) [6]. As shown in Figure 4D, cells stimulated under control conditions showed 2n and 4n peaks of DNA. By contrast, we observed a prominent single 2n peak in cells stimulated with 10 mM 2-DG, indicating that the cells arrested at the G1 to S phase of the cell cycle. These data correlate with the reduced mRNA expression of cyclin D2, which specifically regulates the G1 to S progression.
Figure 4. 2-DG inhibits cell cycle progression of stimulated CD8+ effector T cells.
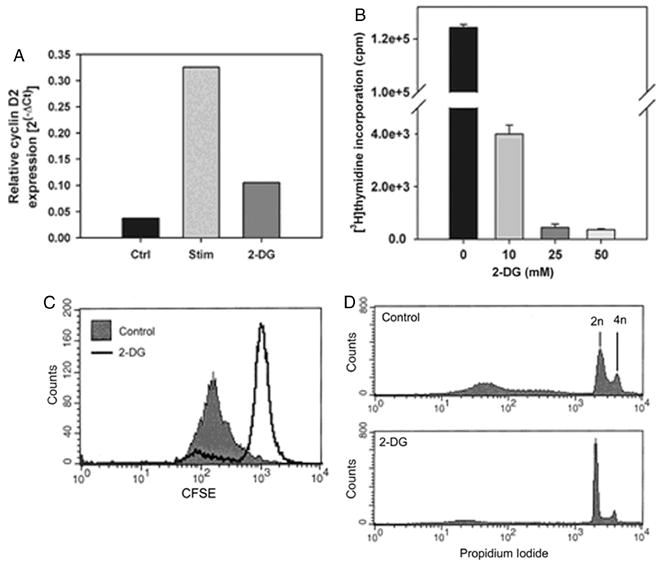
A. Quantitative RT-PCR of cyclin D2 mRNA levels in cells treated anti-CD3 and anti-CD28 mAb-coated beads in the presence or absence of 2-DG. Data represent the average of two independent experiments. B. [3H]thymidine incorporation was assessed in cells stimulated with anti-CD3/anti-CD28 mAb-coated beads, in the presence or absence of 2-DG. Data represent the average + SD of three independent experiments and the inhibition by 2-DG was statistically significant (p<0.001). C. CFSE-labeled 2C CD8+ effector T cells were stimulated with P815-B7.1 cells in the presence or absence of 10 mM 2-DG. After 48 hours, dilution of CFSE was analyzed by flow cytometry. D. DNA content was assessed of effector T cells stimulated for 48 hours with or without 10 mM 2-DG.
Western blot confirmation of inhibition of key gene products
The level of inhibition by 2-DG of several key transcripts appeared to be less profound than the degree of inhibition in the functional assays. We reasoned that one potential explanation may be a greater inhibition revealed at the protein level, as this reflects the total integration of transcription and translation over time. To this end, western blot analysis was performed for selected proteins. As shown in Figure 5, induction of IFN-γ protein detected in whole cell lysates was nearly completely blocked by 2-DG. Similarly, upregulated expression of cyclin D2 and Granzyme B proteins was nearly completely inhibited. Total ERK was used as an internal control, as we had found previously that this signaling pathway was unaffected by glucose deprivation [1]. Therefore, 2-DG has a profound inhibitory effect on the production of selected key proteins involved in T cell function.
Figure 5. Western blot analysis of selected proteins.
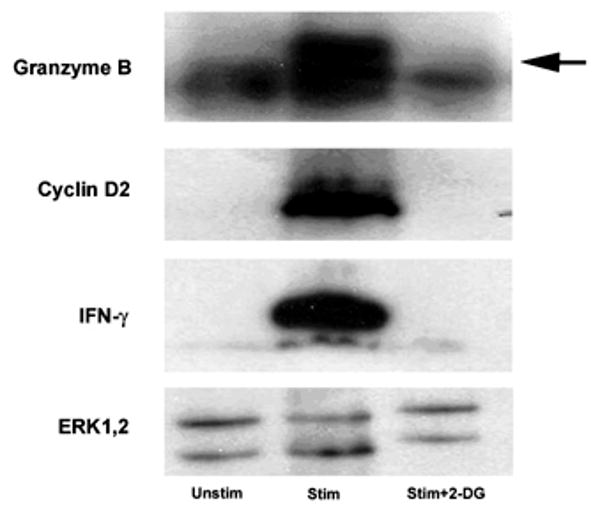
2C/RAG2-/- effector CD8+ T cells were stimulated with anti-CD3/anti-CD28 mAb-coated beads for 16 hours, in the presence or absence of 2-DG. Cells were collected, lysed, separated by SDS-PAGE, and blotted for the indicated proteins. The arrow indicates the Granzyme B band above a background band. Similar results were seen in 2 independent experiments.
Selected CD8+ effector functions are minimally affected by oxygen deprivation
In addition to glucose, oxygen can be an important resource for generating ATP. We therefore assessed the dependence of cytokine production and cytolytic activity of CD8+ effector T cells on oxygen. To this end, CD8+ effector cells were stimulated under 1% O2 (hypoxic) conditions. Primed 2C/RAG2-/- T cells stimulated in the setting of hypoxia were still able to produce comparable levels of IL-2 compared to cells stimulated under 21% O2 (control) conditions (Figure 6A). In contrast to the complete inhibition by 2-DG (Figure 2A), IFN-γ production also was still detectable under hypoxic conditions (Figure 6B). Furthermore, cytolytic activity was equal if not better under hypoxia compared to control conditions (Figure 6C).
Figure 6. Select effector functions are minimally affected by oxygen deprivation.
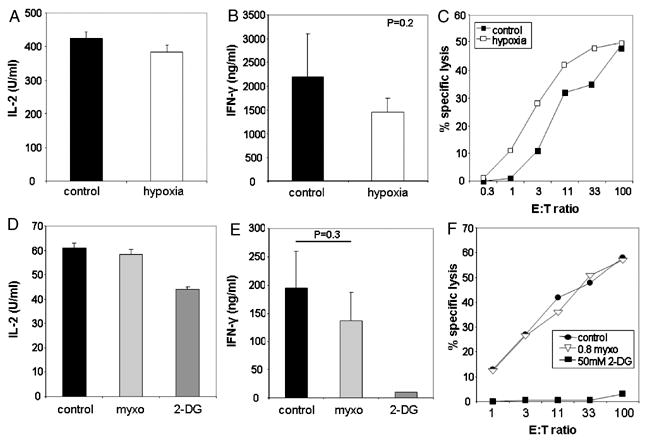
A. and B. Production of IL-2 (A) or IFN-γ (B) under hypoxia as assessed by ELISA. Supernatant was collected after 24 hours of stimulation with anti-CD3 and anti-CD28 mAb-coated beads. Data represent the average + SD of at least three independent experiments. C. Cytolytic activity of CD8+ effector T cells was measured in hypoxic or normoxic conditions. P815 cells were used as specific targets. Control, 21% O2; hypoxia, 1% O2. Data represent the average of two independent experiments. D. and E. Production of IL-2 (D) and IFN-γ (E) stimulated with anti-CD3 and anti-CD28 mAb-coated beads in the presence of 0.8 μM myxothiazol (myxo), a cytochrome b-c1 inhibitor, or 50 mM 2-DG, as assessed by ELISA. Supernatant was collected after 24 hours of stimulation. Data represent the average + SD of at least five independent experiments. F. Cytolytic activity of CD8+ effector T cells was performed in the presence or absence of 2-DG or myxothiazol. Data are represent the average of two independent experiments. As in the other studies, inhibition of IFN-γ production and cytolysis by 2-DG was statistically significant (p<0.05), whereas differences seen with myxothiazol or hypoxia were not.
The 1% oxygen tension used in the experiments was as low as could be achieved by this method. Nevertheless, since 1% oxygen is sufficient to allow some oxidative phosphorylation, we perfomed an alternative approach to block oxygen-dependent metabolism by using myxothiazol, an inhibitor of the mitochondrial cytochrome b-c1 complex, to block aerobic respiration and therefore mimic anoxia. We observed that a concentration of 0.8 mM myxothiazol was sufficient to block oxygen uptake (data not shown). Interestingly, as we observed with hypoxia, myxothiazol had very little effect on IL-2 production (Figure 6D), IFN-γ production (Figure 6E), or cytolysis (Figure 6F). These data support the notion that CD8+ effector functions can be maintained under very low oxygen tension arguing that oxygen is minimally required as an energy source for CD8+ effector T cells when compared to glucose.
Oxygen is not a main energy source for CD8+ effector T cells
Our experiments using hypoxia and myxothiazol suggested that CD8+ effector cells might utilize very little oxygen. To determine this directly, we measured the rate of O2 consumption of CD8+ effector cells. Over the first few minutes of the assay, a very slow rate of oxygen utilization was detected (Figure 7A). In order to determine the maximal and minimal potential rates of oxygen consumption by these cells, additional manipulations were performed. Mitochondrial respiration can be blocked by rotenone, an inhibitor of NADH dehydrogenase. Addition of rotenone (5 μM) blocked this low level of O2 consumption, suggesting that effector cells were indeed metabolizing oxygen via mitochondrial respiration, albeit at a very low level. The potential intrinsic rate of oxygen consumption can be revealed by the supplementation of N, N, N′, N′-tetramethyl-p-phenylenediamine (TMPD) and ascorbate, which are artificial substrates for cytochrome c oxidase complex. TMPD and ascorbate bypass the inhibition of mitochondrial respiration by rotenone. We observed that TMPD and ascorbate induced an extremely brisk rate of oxygen consumption, suggesting that CD8+ effector cells are normally consuming much less oxygen than their full capacity would support. Finally, to determine whether glucose or oxygen is more important for maintaining ATP levels, we stimulated CD8+ effector cells with anti-CD3 and anti-CD28 antibody-coated beads for 3 hours in the presence of 2-DG or under hypoxic conditions. We observed that the concentration of ATP remained in the range of 15-20 μM (per ∼3333 cells) in cells stimulated in the setting of hypoxia but that ATP levels were reduced by greater than 50% with 2-DG (Figure 7B). These data suggest that glucose is the main substrate for ATP generation in these cells, with oxidative phosphorylation making a lesser contribution.
Figure 7. Oxygen is not a main energy source for CD8+ effector T cells.
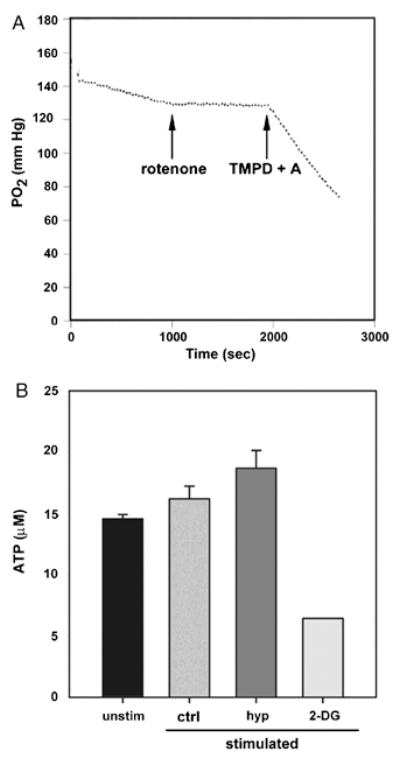
A. The rate of O2 consumption of 3 × 107 unstimulated CD8+ effector T cells. A reduced rate of consumption by the addition of rotenone (5 μM) shows that O2 consumption is mediated via mitochondria. The addition of N, N, N′, N′-tetramethyl-p-phenylenediamine (TMPD; 200 μM) and ascorbate (A; 400 μM) reflects the maximum potential for O2 consumption by these cells. Data are representative of two independent experiments. B. Steady-state levels of ATP in CD8+ effector T cells stimulated with anti-CD3 and anti-CD28 coated beads for 3 hours under the indicated conditions. Control (ctrl), 21% O2, 5% CO2; hypoxia (hyp), 1% O2, 5% CO2; 2-DG (2-DG), 50 mM 2-DG, 21% O2, 5% CO2. Inhibition by 2-DG was statistically significant (p<0.001). Data represent the average + SD of three independent experiments.
Discussion
Based on our previous evidence that glucose-dependent metabolism was required for IFN-γ production by CD8+ effector T cells [1], we examined in the current report the breadth of transcripts that required glucose for inducible expression following CD3/CD28 ligation. Surprisingly, 2-DG blocked expression of only 10% subset of these genes, arguing for a selective effect of glucose deprivation. Also noteworthy was the observation that most genes associated with effector functions were among this subset. This included genes encoding several cytokines, granule proteins, and cell cycle regulators. Thus, glucose metabolism appears to be critically important for executing T cell effector functions and presumably for maintaining immune responses.
The inhibitory effect of 2-DG or glucose deprivation on T cell cytokine production is likely due to inhibition at the level of transcription. The mechanism of this block is not clear, as we have previously shown that defined TCR/CD28-induced signal transduction pathways and nuclear translocation of defined transcription factors were not inhibited by 2-DG [1]. It is important to note that general protein synthesis does not seem to be impaired under our experimental conditions, as numerous proteins were found to be expressed equivalently under conditions of glucose deprivation. No global diminution of transcription occurs either, as 90% of inducible transcripts remained intact. In addition, we had observed no effect on IFN-γ mRNA stability following glucose deprivation. In contrast to regulation of cytokines, the inhibitory effect of 2-DG on cytolytic activity is more complex. In addition to inhibition of transcription of granule-associated proteins, stable conjugate formation between T cells and target cells was diminished by 2-DG. These results are consistent with those of MacDonald et al, who observed that 2-DG inhibited conjugate formation between T cells and allogeneic tumor cells [7]. The mechanism by which conjugate formation is interrupted is not clear. 2-DG does not likely work by preventing N-linked glycosylation of adhesion molecules because treatment with tunicamycin has been reported not to inhibit cytolytic activity [7]. It seems likely that 2-DG is inhibiting the mechanics of adhesion through integrins, perhaps by interfering with “inside-out” signaling through TCR-mediated activation of LFA-1. 2-DG did not affect surface expression of LFA-1 as determined by flow cytometry (data not shown). 5-thio-d-glucose (5-SHG) also has been reported not to inhibit cytolysis [2]. 5-SHG and 2-DG are both competitive inhibitors of glucose for the glycolytic proteins hexokinase, phosphoglucoisomerase, and phosphoglucomutase [8, 9]. While 2-DG may also have specific effects on T cells beyond its inhibition on glycolysis, in our model system, removal of glucose from the medium and addition of 2-DG had similar functional and biochemical effects [1]. We were curious as to whether the effects of 2-DG treatment might have been the result of an induction of ER stress in CD8+ T cells. However, a gel shift assays for the induction of YY1, a transcription factor known to be upregulated after ER stress, did not show any difference between control cells and cells treated by different concentration of 2-DG (data not shown). Still, this pathway may be of interest to investigate further in future studies.
Our observed inhibition of cytolysis by 2-DG are somewhat at odds with the published results of MacDonald and colleagues who showed that 2-DG in the presence of oxygen did not inhibit cytolytic activity [2]. The discrepancy may perhaps be explained by the fact that our study used a pure population of CD8+ T cells in contrast to the mixed population of activated polyclonal T cells used by MacDonald et al. CD4+ and CD8+ T cells may have different metabolic requirements for granule-mediated cytolytic activity. In addition, NK cells may have distinct signal transduction requirements for granule exocytosis compared to CD8+ T cells [10] and also could be differentially susceptible to 2-DG. Since we observed a drop in steady-state levels of ATP in the presence of 2-DG, energy depletion by 2-DG is likely a major factor in our system.
We also observed that 2-DG inhibited cell cycle progression of CD8+ T cells. This inhibition was associated with diminished induction of cyclin D2 and arrest in G1. However, additional modes of action in cell cycle regulation are also possible. Arrest at the G1 to S transition may be promoted by the activation of adenosine-monophosphate-activated protein kinase (AMPK). Tamas, et al. recently showed that 2-DG treatment resulted in the phosphorylation and activation of AMPK [11]. Phosphorylated AMPK has been shown to induce a p53-dependent cell cycle arrest at the G1 to S phase in primary mouse embryonic fibroblasts [12].
We were surprised to find that hypoxic conditions, and even addition of myxothiazol, did not suppress cytokine production and cytolysis in primed CD8+ effector T cells. These results were supported by minimal detected oxygen utilization by these cells, as well as maintained total cellular ATP levels under conditions of hypoxia. Although MacDonald and Koch also reported that hypoxia did not affect cytolytic activity, several subsequent publications contradicted these data [13]. In our hands, even complete inhibition of oxygen utilization by myxothiazol had no effect on cytolysis. Recently, Yi, et al. showed that 5 μM rotenone (which blocks the electron transport complex I) was sufficient to block naïve CD8+ T cell proliferation and other aspects of activation [14]. However, concentrations greater than 100 μM rotenone were required to suppress production of TNF-α and IFN-γ by effector cells, consistent with our results.
A low basal level of oxygen consumption was observed in resting effector CD8+ T cells, suggesting that these cells are not completely anaerobic. However, these measurements were taken at atmospheric (21% O2) conditions; therefore this observation may be restricted to these experimental conditions. We also had difficulty measuring oxygen consumption following T cell activation due to technical limitations. Oxygen concentrations available in vivo would likely be considerably less than the ambient oxygen concentration present in tissue culture conditions, suggesting the possibility that a significant rate of oxygen consumption might not be observed in vivo. The minimal need for high oxygen levels for T cell functions is supported by other published reports. Cell viability has been reported to be completely unaffected by prolonged exposure to hypoxic (2% O2) conditions [15]. Hypoxia actually slightly increased IL-2, IFN-γ, and IL-4 production from human PBMCs stimulated with PHA for 16 hours. Interestingly, a hypoxic environment (2.5% O2) during T cell differentiation was found to generate greater CD8+ cytolytic T cell activity [16]. These data indicate that 2-2.5% oxygen is an environment that can support T cell function. In fact, these conditions are similar to the conditions found in vivo, as oxygen tension measured in the spleen ranged from 0.5-4% O2 [16].
Hypoxic conditions are known to induce upregulation of HIF-1α, which promotes expression of genes encoding glycolytic enzymes but also has other physiologic targets [17]. Moreover, HIF-1α expression itself does not require hypoxia to be expressed. Conditional deletion of HIF-1α in the T cell compartment has been shown to improve T cell function in models of bacterial infection [18], arguing that some HIF-1α gene targets are inhibitory for T cell activation. Based on our current results regarding the importance of glyocolysis for T cell effector functions, the critical HIF-1α targets in that scenario seems unlikely to be the glycolytic machinery. Consistent with other potential roles for HIF-1α beyond glucose metabolism, stabilized expression of HIF-1α in thymocytes has been shown to inhibit calcium signaling through upregulated expression of the calcium pump SERCA2 [19]. Thus, the regulatory effects of HIF-1α in T cells are multiple and complex.
Why T cells would preferentially utilize glucose as an energy source over oxygen, despite less efficient ATP generation, is not clear. Our results are not likely just a reflection of in vitro experimental conditions, as Ahmed and colleagues also reported upregulated expression of glycolytic enzyme genes in effector and memory T cells analyzed ex vivo [20]. Utilization of glucose even in the presence of excess oxygen is known as aerobic glycolysis. This process seen in CD8+ effector T cells is reminiscent of what has been observed in rapidly proliferating cancer cells. In 1930, Otto H. Warburg was the first to describe that tumor cells switch to glycolysis for energy [21]. In our hands, CD8+ effector T cells consumed little oxygen, suggesting that aerobic glycolysis was largely sustaining the energy demands of these cells. It is conceivable that amino acids and lipid precursors, normally prime sources of ATP during catabolic metabolism, are enabled to be redirected into the production of macromolecules that maintain effector function. In addition, effector CD8+ T cells may prefer to utilize glycolysis over aerobic respiration to prevent the generation of reactive oxygen species (ROS). It is well-established that ROS induce DNA damage and can lead to aberrant activation of AP-1 and NF-κB [22]. In T cells, ROS also can mediate activation-induced cell death [23]. In addition, pyruvate, an end product of glycolysis, can serve as a scavenger for ROS [24], adding another benefit from pure glycolysis.
Our results have implications for the tissue requirements to support the effector phase of a CD8+ T cell response. In particular, the tumor microenvironment containing proliferating cancer cells that also preferentially utilize glucose as a nutrient source might be depleted of adequate nutrients for supporting T cell function. We recently observed that tumor cells displayed a greater rate of glucose uptake compared to effector CD8+ T cells in vitro [1]. This could be a reflection of what is happening in a solid tumor, where infiltrating T cells may be starved of glucose, resulting in impaired effector functions. In fact, the dysfunctional state reported for tumor-infiltrating lymphocytes includes defective proliferation, cytokine production, cytolysis [25]. In addition, defects in cell adhesion and conjugate formation have recently been reported [26], making the list of defects quite similar to those seen in glucose-deprived T cells. Dysfunctional tumor-infiltrating lymphocytes can usually recover effector functions after culture ex vivo [27], which is always performed in standard culture medium rich in glucose. It will be of interest to determine directly in future studies whether tumors indeed deprive T cells of glucose. Developing strategies to improve T cell resistance to glucose deprivation may be a novel approach for maintaining T cell effector function in vivo.
Materials and Methods
Mice
2C/RAG2-/- mice were bred in a specific pathogen-free animal facility at The University of Chicago. C57BL/6J and DBA/2J mice were obtained from The Jackson Laboratory (Bar Harbor, ME). Experimental procedures involving mice were approved by the Institutional Animal Care and Use Committee at The University of Chicago.
Cells
The generation of effector 2C TCR transgenic CD8+ T cells has been described [28]. Briefly, naïve 2C cells were purified from the spleens of 2C/RAG2-/- mice using the StemSep Enrichment Cocktail for Murine CD8+ T cells (Stem Cell Technologies, Vancouver, BC, Canada). To generate effector cells, naïve cells were plated with mitomycin C-treated P815-B7.1 cells for 4 days. On the fourth day of stimulation, the cells were collected, subjected to Ficoll-Hypaque centrifugation, and re-primed for another 4 days to generate day 8 effector cells. Day 8 cells routinely were demonstrated to secrete high levels of IFN-γ and to lyse antigen-expressing target cells (data not shown).
In vivo primed T cells were generated by immunization of C57BL/6J mice in the hind footpads with the chicken ovalbumin (1 mg/ml) emulsified in complete Freund's adjuvant. T cells were recovered and purified from lymph nodes, incubated overnight with ovalbumin or with anti-CD3/anti-CD28 mAb-coated beads, and tested for IFN-γ secretion.
Gene array analysis
Total RNA was isolated by using Trizol reagent (Invitrogen, Carlsbad, CA) followed by treatment with DNase I (Invitrogen) and RNeasy Mini column purification (Qiagen, Chatsworth, CA) according to manufacturer's instructions. Integrity of RNA was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA). The purity/concentration was determined using a GeneSpec III (Miraibio). All RNA samples used for hybridization had an OD260/280 and OD260/230 ratio >1.8 and total RNA concentration > 1μg/ml.
All gene array hybridizations were performed at the Functional Genomics Facility, University of Chicago. Targets were prepared according to the Affymetrix GeneChip Expression Analysis Manual (Affymetrix, Santa Clara, CA). Briefly, 10 μg of total RNA was used to synthesize double-stranded cDNA using the Superscript Choice System (Invitrogen). First strand cDNA synthesis was primed with a T7-(dT24) oligonucleotide. From the phase-log gel-purified cDNA, biotin-labeled antisense cRNA was synthesized using BioArray High Yield RNA Transcript Labeling Kit (Enzo Diagnostics, Farmingdale, NY). After precipitation with 4 M LiCl, 20 μg of cRNA was fragmented in fragmentation buffer (40 mM Tris-Acetate, pH 8.1, 100 mM KOAc, 30 mM MgOAc) for 35 minutes at 94° C. Fragmented cRNA (12 μg) was hybridized to arrays for 16 hours at 45° C and 60 rpm in an Affymetrix Hybridization Oven 640. Arrays were washed and stained with streptavidin-PE in an Affymetrix Fluidics Station 450 using the Affymetrix GeneChip protocol and scanned using the Affymetrix GeneChip Scanner 3000.
The acquisition and initial quantification of array images were performed using the GCOS (Affymetrix). Subsequent data analysis involved: i) the normalization of array values by dCHIP software [29], ii) the averaging of two independent samples for each experiment, iii) determining relative expression using normalized values in a Microsoft Excel spreadsheet.
51Cr-release assay
To assess cytolytic activity, 2 × 103 51Cr-labeled targets were plated with 2 × 105 naive or effector CD8+ T cells in a 96-well V-bottom plate (ICN Biomedicals, Costa Mesa, CA). After 4 h of incubation at 37 °C, 50 μl of supernatant was transferred to a LumaPlate-96 (PerkinElmer Life Sciences) and allowed to dry overnight. Plates were then counted using TopCount-NXT (PerkinElmer Life Sciences).
Assays for proliferation
For the [3H]thymidine incorporation assay, 5 × 104 purified CD8+ T cells were stimulated with 5 × 104 mitomycin C-treated P815.B7–1 cells in a 96-well microtiter plate. At various times after plating, 1 μCi of [methyl-3H]thymidine (Amersham Biosciences) was added to the cells. After a 6-hour pulse, the plates were frozen until harvested. The wells were harvested for determination of [3H]thymidine incorporation using a Packard Filtermate Harvester and TopCount-NXT (PerkinElmer Life Sciences).
For CFSE labeling, CD8+ effector T cells were labeled with 5 nM CFSE on ice for 2 minutes. Cells were then quenched with 15 ml FBS, washed twice more with complete 10% FBS/DMEM, and then stimulated with P815-B7.1 cells as APCs.
For the Nicoletti assay, 5 × 105 cells were stimulated in 96-well plates under the indicated conditions and cultured overnight (∼20 hours). Cells were then transferred to fresh 1.5 ml tubes, pelleted for 5 minutes at 2000 rpm, and resuspended in 100 μl Nicoletti buffer (0.1% sodium citrate/0.1% Triton X-100/50 μg/ml PI) for at least 4 hours. DNA content was assessed by flow cytometry.
Cytokine ELISAs
To evaluate cytokine production, 105 purified day 8 effector 2C CD8+ T cells were stimulated with 5 × 105 polystyrene beads (Dynal Biotech Inc., Lake Success, NY) coated with anti-CD3 (2C11; 1 μg/ml) and anti-CD28 (PV-1; 1 μg/ml) antibodies in 96-well flat-bottom plates for approximately 20 hours at 37°C. Supernatants were collected after 20 hours of stimulation and analyzed for cytokine production by ELISA (IL-2 or IFN-γ, antibody pairs from BD Biosciences Pharmingen; GM-CSF, OptEIA for GM-CSF, BD Biosciences Pharmingen).
Western blot analysis
T cells were stimulated as indicated and cell lysates were prepared as described [30]. Proteins were resolved on 10% SDS-PAGE gels, transferred to PVDF membranes, and blotted using published techniques [30]. The antibodies utilized were: anti-Granzyme B (eBiosciences, San Diego, CA), anti-Cyclin D2 (MBL International, Woburn, MA), anti-IFN-γ (BD Biosciences, San Jose, CA), and anti-ERK (Zymed Laboratories, Carlsbad, CA). For the Granzyme B and IFN-γ assays, GolgiPlug (BD Biosciences) was added 2 hours before lysing the cells, to diminish secretion.
Quantitative RT-PCR
Reactions were performed in 50 μl volumes, and carried out in 96-well optical plates. ABI 7700 thermal cycler (Applied Biosystems, Foster City, CA) was used for amplification. Each gene was evaluated using a specific primer/probe set purchased from Applied Biosystems, and labeled with FAM dye. ΔCT of a particular gene was normalized against the ΔCT of β-actin. Data was analyzed using SDS Software (Applied Biosystems).
Hypoxia experiments
Complete DMEM containing 10% FBS was equilibrated under hypoxic (1% O2/5% CO2; 37°C) conditions overnight prior to use. Cells and reagents were prepared under ambient (21% O2/5% CO2; 37°C) conditions. At the final step, the experiment was set up inside the hypoxia chamber. Experiments conducted under hypoxic (1% O2/5% CO2; 37°C) conditions were performed, as described, at 37°C in a sealed, humidified hypoxia chamber (Coy Laboratory Products, Grass Lake, MI). Gas pressures of O2 and CO2 were maintained by mass flow controllers. Parallel controls were conducted under ambient incubator conditions.
Oxygen consumption
The rate of oxygen consumption from 30 × 106 naïve and effector 2C CD8+ T cells was measured in ambient air at room temperature using a Warburg chamber and a polarographic O2 electrode. Data was obtained and analyzed using Pegasus software (Lakeshore Technologies, Chicago, IL). Rotenone, N, N, N′, N′-tetramethyl-p-phenylenediamine (TMPD), and ascorbic acid (all from Sigma, St. Louis, MO) was added directly into Warburg chamber during the experiment.
Measurement of ATP levels
Steady-state ATP levels were determined using the ATP Bioluminescence Kit HS II (Roche), according to manufacturer's instructions. 106 effector CD8+ T cells were cultured with either 5 × 106 uncoated or 5 × 106 anti-CD3 and anti-CD28 antibody-coated beads (0.5 ml total volume) under the indicated conditions in a 1.5 ml microfuge tube with a poked hole to allow for gas exchange. Following a 3-hour stimulation, cells were washed once with ice-cold PBS and resuspended in 0.2 ml cell lysis buffer (included in kit). Lysate was further diluted with dilution buffer (included in kit) in two serial 1:10 dilutions for a final volume of 0.2 ml. An equal volume of cell lysis buffer was added to the lysate. Triplicate samples of 0.1 ml lysate and 0.1 ml luciferase substrate were then read in triplicate using a Monolight luminometer (BD Pharmingen) and averaged. Therefore, the ATP concentration of 3333 cells is represented.
Conjugate formation
This assay was performed similarly as described previously [31]. Briefly, T cells were labeled with calcein AM (Molecular Probes, Eugene, OR), and either EL4 cells or P815 cells were labeled with PKH (Sigma, Saint Louis, MO). T cells (2.5×105) and targets (5×105) were mixed, centrifuged, lightly vortexed, and incubated at 37°C for the indicated times. Cells were then vortexed vigorously for 30 seconds and immediately fixed. 2-color flow cytometric analysis was performed, and percent conjugates were determined by calculating the ratio of double positive conjugates to the total number of T cells.
Immunofluorescence microscopy
To distinguish APCs from T cells, APCs were loaded with the vital dye CMAC Cell-Tracker Blue (Molecular Probes) as previously described [31]. APCs (1.5×105) were mixed with an equal number of Ficoll-Hypaque purified T cells in DMEM with 10% FBS, centrifuged at 5000 rpm for 30 seconds, and incubated for the indicated time (minus 2 minutes) at 37°C. Supernatant was aspirated, and conjugates were then gently resuspended in serum free DMEM using a 1000-μl pipette and plated onto poly-L-lysine (m.w. 30,000–70,000, Sigma)-coated slides for 2 min before fixation. Slides were fixed in 3% (w/v) paraformaldehyde in PBS for 15 minutes. Samples were permeabilized in 0.3% (v/v) Triton X-100 (Sigma) in PBS for 10 minutes, and then rinsed in PBS and blocked in DMEM containing 10% FBS for 5 minutes. All subsequent antibody incubations were performed in calcium/magnesium-free DPBS containing 2% FBS. Primary and secondary Abs were applied sequentially for 60 minutes at room temperature and washed five times after each incubation with DPBS. After fluorochrome labeling, specimens were mounted in Mowiol 4-88 (Hoechst Celanese, Charlotte, NC), with 10% 1,4-diazobicyclo[2.2.2]octane (Sigma) added as an anti-fade. Samples were analyzed using a Zeiss Axiovert100 microscope. Image capture and deconvolution analysis (where appropriate) was performed using SlideBook software (Intelligent Imaging Innovations, Denver, CO).
Statistics
Differences between groups were analyzed using a student's t-test with Excel software. A p-value less than 0.05 was used for significance.
Acknowledgments
We gratefully acknowledge Janel Washington for expert mouse assistance, Helena Harlin for help with gene array data analysis, Matthew Mack, Robert Guzy, and Paul Schumacker for valuable assistance with hypoxia/oxygen experiments and advice, and Maria-Luisa Alegre, Kenneth Frauwirth, and Jeffrey Rathmell for helpful discussions. We also thank Xinmin Li at the Functional Genomics Facility of The University of Chicago for gene array analysis.
This work was supported by RO1 AI47919 from the NIH.
Abbreviations
- 2-DG
2-deoxy-d-glucose
- ROS
reactive oxygen species
- O2
oxygen
- cSMAC
central supramolecular activating cluster
- pSMAC
peripheral supramolecular activating cluster
- mTOC
microtubule organizing center
- IS
immunological synapse
- PI
propidium iodide
- 5-SHG
5-thio-d-glucose
- TMPD
tetramethyl-p-phenylenediamine
- AMPK
adenosine-monophosphate-activated protein kinase
Footnotes
Conflict of interest: The authors declare no financial or commercial conflict of interest.
References
- 1.Cham CM, Gajewski TF. Glucose availability regulates IFN-gamma production and p70S6 kinase activation in CD8+ effector T cells. J Immunol. 2005;174:4670–4677. doi: 10.4049/jimmunol.174.8.4670. [DOI] [PubMed] [Google Scholar]
- 2.MacDonald HR. Energy metabolism and T-cell-mediated cytolysis. II. Selective inhibition of cytolysis by 2-deoxy-D-glucose. J Exp Med. 1977;146:710–719. doi: 10.1084/jem.146.3.710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fox CJ, Hammerman PS, Thompson CB. Fuel feeds function: energy metabolism and the T-cell response. Nat Rev Immunol. 2005;5:844–852. doi: 10.1038/nri1710. [DOI] [PubMed] [Google Scholar]
- 4.Stinchcombe JC, Bossi G, Booth S, Griffiths GM. The immunological synapse of CTL contains a secretory domain and membrane bridges. Immunity. 2001;15:751–761. doi: 10.1016/s1074-7613(01)00234-5. [DOI] [PubMed] [Google Scholar]
- 5.Monks CR, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 1998;395:82–86. doi: 10.1038/25764. [DOI] [PubMed] [Google Scholar]
- 6.Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J Immunol Methods. 1991;139:271–279. doi: 10.1016/0022-1759(91)90198-o. [DOI] [PubMed] [Google Scholar]
- 7.MacDonald HR, Cerottini JC. Inhibition of T cell-mediated cytolysis by 2-deoxy-D-glucose:dissociation of the inhibitory effect from glycoprotein synthesis. Eur J Immunol. 1979;9:466–470. doi: 10.1002/eji.1830090610. [DOI] [PubMed] [Google Scholar]
- 8.Chen M, Whistler RL. Action of 5-thio-D-glucose and its 1-phosphate with hexokinase and phosphoglucomutase. Arch Biochem Biophys. 1975;169:392–396. doi: 10.1016/0003-9861(75)90180-0. [DOI] [PubMed] [Google Scholar]
- 9.Wick AN, Drury DR, Nakada HI, Wolfe JB. Localization of the primary metabolic block produced by 2-deoxyglucose. J Biol Chem. 1957;224:963–969. [PubMed] [Google Scholar]
- 10.Ting AT, Schoon RA, Abraham RT, Leibson PJ. Interaction between protein kinase C-dependent and G protein-dependent pathways in the regulation of natural killer cell granule exocytosis. J Biol Chem. 1992;267:23957–23962. [PubMed] [Google Scholar]
- 11.Tamas P, Hawley SA, Clarke RG, Mustard KJ, Green K, Hardie DG, Cantrell DA. Regulation of the energy sensor AMP-activated protein kinase by antigen receptor and Ca2+ in T lymphocytes. J Exp Med. 2006;203:1665–1670. doi: 10.1084/jem.20052469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jones RG, Plas DR, Kubek S, Buzzai M, Mu J, Xu Y, Birnbaum MJ, Thompson CB. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol Cell. 2005;18:283–293. doi: 10.1016/j.molcel.2005.03.027. [DOI] [PubMed] [Google Scholar]
- 13.Nathan CF, Mercer-Smith JA, Desantis NM, Palladino MA. Role of oxygen in T cell-mediated cytolysis. J Immunol. 1982;129:2164–2171. [PubMed] [Google Scholar]
- 14.Yi JS, Holbrook BC, Michalek RD, Laniewski NG, Grayson JM. Electron transport complex I is required for CD8+ T cell function. J Immunol. 2006;(177):852–862. doi: 10.4049/jimmunol.177.2.852. [DOI] [PubMed] [Google Scholar]
- 15.Naldini A, Carraro F, Silvestri S, Bocci V. Hypoxia affects cytokine production and proliferative responses by human peripheral mononuclear cells. J Cell Physiol. 1997;173:335–342. doi: 10.1002/(SICI)1097-4652(199712)173:3<335::AID-JCP5>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- 16.Caldwell CC, Kojima H, Lukashev D, Armstrong J, Farber M, Apasov SG, Sitkovsky MV. Differential effects of physiologically relevant hypoxic conditions on T lymphocyte development and effector functions. J Immunol. 2001;167:6140–6149. doi: 10.4049/jimmunol.167.11.6140. [DOI] [PubMed] [Google Scholar]
- 17.Sitkovsky M, Lukashev D. Regulation of immune cells by local-tissue oxygen tension: HIF1 alpha and adenosine receptors. Nat Rev Immunol. 2005;5:712–721. doi: 10.1038/nri1685. [DOI] [PubMed] [Google Scholar]
- 18.Thiel M, Caldwell CC, Kreth S, Kuboki S, Chen P, Smith P, Ohta A, et al. Targeted deletion of HIF-1alpha gene in T cells prevents their inhibition in hypoxic inflamed tissues and improves septic mice survival. PLoS ONE. 2007;2:e853. doi: 10.1371/journal.pone.0000853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Neumann AK, Yang J, Biju MP, Joseph SK, Johnson RS, Haase VH, Freedman BD, Turka LA. Hypoxia inducible factor 1 alpha regulates T cell receptor signal transduction. Proc Natl Acad Sci U S A. 2005;102:17071–17076. doi: 10.1073/pnas.0506070102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kaech SM, Hemby S, Kersh E, Ahmed R. Molecular and functional profiling of memory CD8 T cell differentiation. Cell. 2002;111:837–851. doi: 10.1016/s0092-8674(02)01139-x. [DOI] [PubMed] [Google Scholar]
- 21.Warburg OH. The metabolism of tumours. 1930 [Google Scholar]
- 22.Klaunig JE, Kamendulis LM. The role of oxidative stress in carcinogenesis. Annu Rev Pharmacol Toxicol. 2004;44:239–267. doi: 10.1146/annurev.pharmtox.44.101802.121851. [DOI] [PubMed] [Google Scholar]
- 23.Hildeman DA, Mitchell T, Teague TK, Henson P, Day BJ, Kappler J, Marrack PC. Reactive oxygen species regulate activation-induced T cell apoptosis. Immunity. 1999;10:735–744. doi: 10.1016/s1074-7613(00)80072-2. [DOI] [PubMed] [Google Scholar]
- 24.O'Donnell-Tormey J, Nathan CF, Lanks K, DeBoer CJ, de la Harpe J. Secretion of pyruvate. An antioxidant defense of mammalian cells. J Exp Med. 1987;(165):500–514. doi: 10.1084/jem.165.2.500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Radoja S, Frey AB. Cancer-induced defective cytotoxic T lymphocyte effector function: another mechanism how antigenic tumors escape immune-mediated killing. Mol Med. 2000;6:465–479. [PMC free article] [PubMed] [Google Scholar]
- 26.Koneru M, Monu N, Schaer D, Barletta J, Frey AB. Defective adhesion in tumor infiltrating CD8+ T cells. J Immunol. 2006;176:6103–6111. doi: 10.4049/jimmunol.176.10.6103. [DOI] [PubMed] [Google Scholar]
- 27.Radoja S, Rao TD, Hillman D, Frey AB. Mice bearing late-stage tumors have normal functional systemic T cell responses in vitro and in vivo. J Immunol. 2000;(164):2619–2628. doi: 10.4049/jimmunol.164.5.2619. [DOI] [PubMed] [Google Scholar]
- 28.Cham CM, Xu H, O'Keefe JP, Rivas FV, Zagouras P, Gajewski TF. Gene array and protein expression profiles suggest post-transcriptional regulation during CD8+ T cell differentiation. J Biol Chem. 2003;278:17044–17052. doi: 10.1074/jbc.M212741200. [DOI] [PubMed] [Google Scholar]
- 29.Li C, Wong WH. Model-based analysis of oligonucleotide arrays: expression index computation and outlier detection. Proc Natl Acad Sci U S A. 2001;98:31–36. doi: 10.1073/pnas.011404098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rivas FV, O'Herrin S, Gajewski TF. CD28 is not required for c-Jun N-terminal kinase activation in T cells. J Immunol. 2001;167:3123–3128. doi: 10.4049/jimmunol.167.6.3123. [DOI] [PubMed] [Google Scholar]
- 31.Morgan MM, Labno CM, Van Seventer GA, Denny MF, Straus DB, Burkhardt JK. Superantigen-induced T cell:B cell conjugation is mediated by LFA-1 and requires signaling through Lck, but not ZAP-70. J Immunol. 2001;167:5708–5718. doi: 10.4049/jimmunol.167.10.5708. [DOI] [PubMed] [Google Scholar]