

Summary
Dendritic spines are small actin-rich protrusions on the surface of dendrites whose morphological and molecular plasticity play key roles in learning and memory. Both the form and function of spines are critically dependent on the actin cytoskeleton. However, new research, using electron microscopy and live-cell super-resolution microscopy indicates that the actin cytoskeleton is more complex and dynamic than originally thought. Also, exciting recent studies from several labs indicate that microtubules, once thought to be restricted to the dendrite shaft, can make excursions into the most distal regions of dendritic spines. Moreover, microtubule invasions of spines appear to be associated with changes in synaptic activity. Thus, it is likely that dynamic interactions between microtubules and actin filaments within dendritic spines play important roles in dendritic spine plasticity.
Introduction
Organisms must be able to modify their synaptic connections during development and in adulthood. This synaptic plasticity is essential for the refinement of connections during the construction of a functional nervous system as well as for learning and memory throughout life. When processes involved in synaptic plasticity go awry, either in disease or during aging, dysfunction of the organism results. Myriad studies have provided evidence for both short- and long-term changes in synaptic efficacy. Both types of synaptic changes can be regulated at the level of either the presynaptic axon or the postsynaptic dendritic spine. Synaptic plasticity is accompanied by a change in the molecular composition of the synapse that results in either more or less efficient synaptic transmission, sometimes being regulated by changes in the morphology of dendritic spines[1].
Dendritic spines are micron-sized actin-rich protrusions on dendrites that serve as the primary source of synaptic contacts for excitatory neurons in the nervous system. Dendritic protrusions have generally been classified morphologically into several types, including filopodia, stubby spines, thin spines, mushroom-shaped spines and complex spines [2-7]. However, these shapes are not fixed. In the rodent brain most spines maintain their relative shape throughout life, although spines are capable of converting into different shapes or even appearing/disappearing [8-10]. Regardless of the ability of spines to morph into different shapes or remain stable, it is clear that the turnover of proteins, lipids and organelles in spines is an ongoing and dynamic process[11,12].
Dynamics of both actin filaments and microtubules are important for providing the structure for cellular shape changes. Moreover, changes in structure influence function. Since dendritic spines are rich in actin filaments that undergo dynamic assembly and disassembly, it follows that both the structure and function of spines is dictated by actin dynamics. But what about the function of microtubule dynamics in spine function? In this review we will present new evidence for a complex actin organization and dynamics in spines and also evaluate several recent studies that demonstrate microtubules are capable of entering dendritic spines in a dynamic, activity-dependent fashion.
Actin organization and dynamics in dendritic spines
Filamentous actin (f-actin) is the primary cytoskeletal element in dendritic spines and there are a plethora of actin-associated proteins whose functional interactions with the actin cytoskeleton are an active area of investigation [13-18]. Here we will touch upon several new studies that have focused directly on actin architecture and dynamics. The enrichment of actin in dendritic spines has been known since the early 1980’s [19-21]. One of these studies showed, with myosin-S1 decoration, that f-actin was primarily arrayed with the barbed (fast growing) end toward the plasma membrane and the actin filaments were generally short and arranged in a crosshatched and branched formation [21].
A recent study has confirmed and extended these studies using platinum replica electron microscopy to show that dendritic filopodia, which can mature into spines, have a very different actin network structure from most other filopodia, including those in non-neuronal cells and axonal growth cones [22*]. Instead of having parallel actin filaments that are oriented with barbed ends toward the tip of the filopodium, dendritic filopodia appear to form from a mixed network of both linear and branched actin filaments that extend to form a filopodium. Furthermore, in contrast to other filopodia, they appear to contain concentrations of Arp2/3, capping protein and myosin II, while lacking fascin [22*]. Fascin bundles linear actin filaments that are oriented in the same direction, while myosin II can only organize actin filaments of opposite polarity. Arp2/3 and capping protein are necessary to form branched actin filaments and maintain their short length, respectively. These data led the authors to propose a model in which dendritic filopodia and spines contain a mixed array of linear and branched actin filaments throughout the spine head, neck and base. Interestingly, they also discovered that actin filaments appear to be interdigitated among the microtubules present in the dendrite shaft, which may have implications for microtubule entry into dendritic spines.
Although electron micrographs provide extremely high resolution images of dendritic spines, they can only hint at how dynamic processes such as actin polymerization and depolymerization might be regulated in spines. Previous studies have shown that actin is in a very dynamic state in spines, with a turnover time of less than a minute [23]. Actin dynamics are also important for spine plasticity. Several previous studies have documented that actin polymerization is necessary for long-term potentiation (LTP) and is reduced following induction of long-term depression (LTD) [24,25]. Thus, determining where actin is polymerized and depolymerized within spines is critical for understanding how it regulates spine structure and function. However, these studies did not have the ability to determine how actin dynamics were regulated in sub-regions of spines.
Several recent studies have shed light on where actin is polymerized and depolymerized in different regions of the spine. An elegant study by the Kasai lab, using two photon uncaging of photoactivatable actin (PA-β-actin) in hippocampal slices, showed that there are at least three pools of f-actin in spines [26**]. The first is a dynamic pool (time constant ~40 sec.) that polymerizes near the tip of the spine and treadmills toward the center of the spine (Fig. 1a). This was the first direct demonstration that actin in spines undergoes treadmilling in a manner similar to actin in growth cones of cortical neurons [27] and newly differentiated hippocampal neurons [28]. However, the actin retrograde flow in spines (~1μm/min) is slower than that of actin in younger mammalian CNS neurons (6-8μm/min). This slower flow could be due to a different structure of actin filaments or fewer or less efficient coupling of myosin II motors [29,30] to the actin cytoskeleton. The second pool of actin was a relatively stable pool (time constant ~17 min.) that was located at the base of the spine and varied according to spine volume [26**]. The third pool of actin, termed the enlargement pool, was induced by uncaging glutamate on an individual spine and turned over relatively slowly (2-15 min.). The increase in this pool of actin was necessary but not sufficient for a long-term increase in spine size, as it was also dependent on the confinement of the pool by the spine neck.
Figure 1.
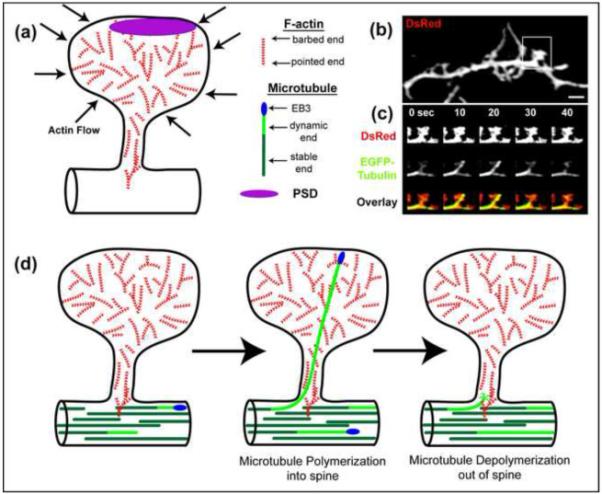
Model for dynamism of f-actin and microtubules in dendritic spines.
(a) Model of a dendritic spine showing the possible arrangement of actin filaments in the spine head, neck and dendritic shaft. Actin takes the form of short filaments with several of the filaments having their barbed (fast growing) ends near the membrane. Note that actin filaments do not have uniform polarity either in the head or shaft. The actin polymerizes at the membrane forcing a flow of actin toward the center of the spine. Myosin may play a role in this actin flow as well. (b) Image of a dendrite from a hippocampal neurons transfected with DsRed and EGFP-tubulin (DIV18). (c) Time-lapse images of the spine demarcated by the box in (b). DsRed label shows the morphology of the spine, while EGFP-tubulin labels microtubules. One dynamic microtubule extends into and retracts from the spine over a period of 40 seconds. (d) Model showing how a microtubule polymerizes into and retracts from a dendritic spine. Microtubule entry is due to polymerization of the dynamic end of the microtubule into the spine. EB3, an important microtubule +TIP protein, binds to the ends of polymerizing microtubules in both the spine and the dendrite shaft. Note that microtubules can extend to the most distal regions of the spine and have even been shown to deform the spine head membrane, forming a transient spine head protrusion.
Using a barbed-end polymerization assay and FRAP analysis another study showed that actin was capable of polymerizing at the spine tip, confirming the study by Kasai and colleagues [26**], but also demonstrating that actin can be polymerized at the base of the spine [31*]. However, actin polymerization in the base of the protrusion occurred more frequently in filopodia from less mature cultures and much less frequently in dendritic spines. Recently, another study, using single molecule tracking of fluorescent actin monomers that polymerized within otherwise non-fluorescent actin filaments, showed that actin filament treadmilling within dendritic spines is a complex and varied process that takes place throughout the spine head and shaft [32*]. However, unlike the polarized actin filament array observed in the lamellipodium of fibroblasts [32*] or newly differentiated neurons [28] these filaments were much less polarized, with only a small portion of filaments exhibiting flow (0.5-2μm/min) in any one direction. This study is both consistent with the older study that used electron microscopy [21] and the speed of actin treadmilling measured with confocal microscopy [26**].
Blanpied and colleagues have recently expanded on these findings, using single particle tracking and photoactivated localization microscopy (PALM) [33,34] of a photoswitchable actin probe (actin-mEos) to track the movements of individual actin molecules in different subregions of dendritic spines [35**]. Using this approach, the authors found that actin filaments participate in retrograde flow from the periphery towards a central region of the spine, roughly where the spine’s head meets its neck. Importantly, they found that actin filaments are particularly dynamic near the post-synaptic density (PSD) but not near endocytic zones, suggesting that stability of the PSD is conferred by an active process of continuous turnover in the actin network. Whereas, clathrin-mediated endocytosis depends on either a relatively stable f-actin network or on transient, sparsely occurring actin polymerization events.
Together these studies paint a picture of a relatively dynamic and complex actin filament array that is essential for the functional readout of the dendritic spine. Generally, it has been thought that microtubules, which are rich in the dendrite shaft, do not make excursions into dendritic spines. Consequently, microtubules were not thought to play any role in dendritic spines directly. In many other systems however, actin filaments and microtubules participate intimately in many processes [36,37]. Modern advances in high-resolution live-cell microscopy have permitted a reinvestigation of cytoskeletal dynamics in dendritic spines. Indeed, several new studies have now shown that microtubules do in fact enter dendritic spines, opening up the possibility that actin/microtubule interactions are essential for spine function as well.
History of microtubule involvement in dendritic spines
The first studies to hint at a role for microtubules in dendritic spines were by Gray and Westrum in the early 1980’s [38-40]. These studies showed with electron microscopy that microtubules could penetrate dendritic spines of neurons within intact regions of cortex, sometimes extending directly into the electron-dense post-synaptic density (PSD) [38,40]. However, these results were called into question for a number of reasons. First, the regions of cortex were dissected directly into a hypotonic solution of 20% BSA in water. This unusual procedure may have caused spurious polymerization of microtubules into spines. Furthermore, several recent studies failed to show microtubule entry into spines under physiological conditions.
In one study Harris and colleagues were able to show, using serial reconstruction electron microscopy, that microtubules could enter spines, but only the uniquely branched spines within the CA3 region in the hippocampus [41]. Another study by the same group using similar techniques showed that dendritic microtubules depolymerize after sectioning at 4°C, but after warming of the slices they appear in approximately 10-15% of hippocampal spines within the first half-hour after cutting the slices [42]. However, this study failed to find microtubules in dendritic spines of similar slices at three hours after slice equilibration or in hippocampi of animals that were directly perfused with fixative. These results led them to conclude that although microtubule entry into dendritic spines could occur, spines were devoid of microtubules under physiological conditions. Nonetheless it is possible that a population of highly dynamic, unstable microtubules were depolymerized during the fixation process, making the subcellular localization of these microtubules impossible to determine.
The absence of microtubules in spines was corroborated by two studies from Matus and colleagues, who examined mature dissociated hippocampal neurons transfected with actin and the microtubule associated protein MAP2c fused to different fluorescent proteins [43,44]. In both fixed and live cells these authors observed that MAP2c-labeled microtubules did not enter dendritic spines. However, they also found little microtubule dynamics in dendrite shafts. This may have been due to a stabilizing effect of overexpression of a structural MAP. Alternatively, MAP2c may preferentially label stable regions of microtubule that do not undergo dynamic bouts of polymerization or depolymerization. More recently, the microtubule end binding protein EB3 (fused to EGFP) was imaged in 10DIV hippocampal neurons, revealing that microtubules are indeed dynamic in younger dendrites, though this study made no mention of microtubule tip entry into dendritic protrusions [45]. In light of this study, it is likely that the methods employed in earlier electron microscopy and fluorescence imaging studies precluded detection of highly dynamic microtubules, resulting in an incomplete understanding of the activities of microtubules in neuronal dendrites and spines.
Evidence for microtubule entry into dendritic spines
Recently, four independent studies published within a one year period showed that microtubules enter spines under physiological conditions in cultured hippocampal [46**-48**] and cortical neurons [47**], as well as after tetanic stimulation of hippocampal slices [49*]. Using different methodologies, each of these studies showed that microtubule entry into spines was likely to be dependent on synaptic activity. Mitsuyama and colleagues demonstrated via electron microscopy that strong tetanic stimulation in the CA1 region of hippocampus caused dendrites to assume an undulating shape and an enlargement of postsynaptic density (PSD) [49*]. Accompanying these morphological changes to CA1 dendrites was the presence of microtubules in dendritic spines, oftentimes protruding into the PSD itself. Many studies have documented changes in spines after strong stimulation protocols but have not shown microtubule entry. What made this study different? The authors suggested that using a microtubule-stabilizing fixative was essential for preserving the otherwise labile microtubules that entered dendritic protrusions, while the use of conventional fixatives did not preserve this population of microtubules [49*]. This was similar in reasoning, although different in methodology, to the earlier studies of Gray and Westrum [38-40]. Another unanswered question from these studies was whether microtubules polymerized into spines from the dendrite shaft or were locally nucleated within the spine and then polymerized into the dendritic shaft, as suggested by others [50].
To address this issue by direct observation within living neurons, another group used time-lapse total internal reflection fluorescence microscopy (TIRFM) to directly observe microtubule polymerization from the dendritic shaft into spines [47**]. By labeling microtubules with EGFP-tubulin and imaging them at high resolution and quick succession Dent and colleagues showed that microtubule entry into dendritic protrusions (both spines and filopodia) occurred in only a small percentage of protrusions at any one time (~1% of spines) and was transient, averaging about three minutes per invasion (ranging from 20 seconds to ~30 min.) (Fig. 1b-c). By imaging microtubule dynamics, with either EGFP-tubulin or EB3-EGFP, a microtubule end binding protein, they documented that 10% of dendritic protrusions were targeted by microtubules in an hour [47**] (Fig. 1d). Extrapolating these results suggests that most dendritic protrusions would be targeted by microtubules over the course of a day. However, it is not known if some spines are preferentially targeted or if others are never targeted. Surprisingly, microtubule invasion could induce transient extensions from spine heads in a small number of cases [47**]. Microtubule invasion of spines appears to be an ongoing process because it occurred in very mature hippocampal neurons (DIV63). This is surprising because maintaining microtubules in a dynamic state is an energy intensive process governed by GTP hydrolysis, suggesting that microtubule dynamics probably serves important functions in mature dendrites. It remains unknown whether microtubule invasion of spines occurs in an intact animal throughout its lifespan, although with techniques such as two-photon imaging of cortical dendrites in living mice this hypothesis could be tested.
Microtubule entry into dendritic spines is activity-dependent
Based on the results of the Mitsuyama study [49*] it is likely that microtubule entry into dendritic spines is dependent on synaptic activity and contributes to synapse plasticity and/or formation. Three separate studies, using different methods of stimulating activity, showed that this is indeed the case [46**-48**]. Zheng and colleagues documented the presence of microtubules in dendritic spines with confocal microscopy of fixed cultures, and used BDNF to stimulate activity in these cultures [46**]. In two week old hippocampal cultures they showed that the increase in density of dendritic spines induced by BDNF treatment was augmented by pharmacological stabilization of microtubules and mitigated by pharmacological inhibition of microtubule dynamics [46**]. These results were corroborated through shRNA knockdown of the +TIP protein EB3, which also decreased spine density [46**,51]. However, in a study by Hoogenraad and colleagues, EB3 knockdown with shRNA, EB3 overexpression and pharmacological inhibition of microtubule dynamics resulted in no change to spine density. Rather, EB3 shRNA caused an increase in filopodia at the expense of spines and EB3 overexpression promoted spine development at the expense of filopodia [48**]. At present, it is unclear why these two studies, using similar techniques to disrupt EB3, obtained conflicting results with regard to spine formation and maintenance.
Another study showed that microtubule entry of spines was increased three-fold after transient KCl applications to stimulate cultured hippocampal neurons [47**]. This increase in microtubule invasions was abolished by inclusion of TTX, indicating it was action potential driven. It is likely that more physiological changes in synaptic activity, such as LTP, homeostatic plasticity and BDNF application, also cause changes in the kinetics of microtubule invasion of spines (Merriam, Hu and Dent, unpublished observations). Indeed, pharmacological inhibition of microtubule dynamics in hippocampal slice cultures could inhibit the maintenance of LTP [48**]. Nevertheless, bath application of pharmacological compounds to inhibit microtubule dynamics or knockdown of microtubule-associated proteins, such as EB3, will affect microtubule dynamics throughout the entire neuron and surrounding glia and potentially disrupt microtubule-based transport as well [46**,48**]. Thus, it is not known what specific role microtubule entry into dendritic spines plays in either BDNF induced plasticity or LTP. Further experiments that disrupt microtubule dynamics locally within individual dendritic spines, or small numbers of spines, will be necessary to determine if microtubule invasion of spines is essential for different forms of synaptic plasticity.
Conclusions and future directions
Actin is clearly essential for dendritic spine structure and function. Recently, several elegant studies have confirmed this by providing images of actin structure and dynamics at unprecedented detail with platinum replica electron microscopy [22*], two-photon confocal imaging of photactivated actin [26**], single molecule imaging of actin dynamics [32*] and single particle tracking PALM [35**]. Taken together, these studies indicate that actin in dendritic spines forms a complex, dynamic meshwork that is likely to be regulated differentially in sub-compartments of the spine. Further work will be necessary to functionally integrate all of the actin-associated proteins that are present in the spine at this level of detail, but will undoubtedly yield new surprises in dendritic spine function.
Interestingly, new studies show that microtubules enter dendritic spines in both hippocampal and cortical neurons and may be important for their morphological and functional plasticity [46**-49*]. As with other interesting studies, the findings raise more questions than they answer. Two important questions that remain are 1) how do microtubules enter spines, and 2) what function do they serve upon entry? Although we now know that MTs polymerize from the dendrite shaft into spines [47**,48**], how do they polymerize from the dendritic shaft, through a thin spine neck, into the head of the spine? Clues from the aforementioned studies indicate that actin filaments and associated proteins are likely to play an important role [13]. In fact, one study has shown that the relatively little studied protein p140Cap and the actin associated protein cortactin are likely to form a complex that regulates interactions with microtubules through EB3 [48**]. It will be interesting to determine if actin filaments in dendritic spines act as a barrier for, a scaffold for, or do not affect microtubule invasion of spines. Likely, based on what is known about the complexity of actin dynamics in spines, there will prove to be very finely tuned and complex interactions between these two cytoskeletal elements.
The function microtubules serve by transiently targeting dendritic spines is likely to involve transport of essential proteins into and out of the spine, since microtubules are the major long-distance transport machinery inside all cells. It is possible that microtubule entry into the entire extent of dendritic protrusions, either spines or filopodia, may play an important role in “tagging” synapses, or synaptic precursors, for modification. This tagging could result in the specific kinesin-mediated delivery and/or dynein-mediated removal of receptors, structural proteins, mRNA, GTPase effectors or organelles that may be required for synaptic development or plasticity. Since kinesin and dynein driven transport is an order of magnitude faster than the rate of microtubule polymerization/depolymerization it would be possible to transport substantial cargo into or out of spines on microtubules during their relatively short excursions (minutes) into spines. This hypothesis does not imply that microtubule-driven delivery directly into spines is the only route for material destined for spines. For example, it is established that myosin motor-driven processes are essential for transport of material into spines [52,53]. However, the fact that microtubules remain dynamic and capable of entering spines throughout the lifetime of neurons suggests that they serve fundamental roles in synaptic architecture and functional plasticity.
Acknowledgments
We apologize to those authors whose work we were unable to cite due to space restrictions. We thank Witchuda Saengsawang and Thomas Fothergill for critical reading of the manuscript. Work in the authors’ laboratory is supported by grants from the National Institutes of Health (R01NS064014 to E.W.D and F31NS071816 to E.B.M), the Dana Foundation (E.W.D) and the Whitehall Foundation (E.W.D).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References and recommended reading
- 1.Alvarez VA, Sabatini BL. Anatomical and physiological plasticity of dendritic spines. Annu Rev Neurosci. 2007;30:79–97. doi: 10.1146/annurev.neuro.30.051606.094222. [DOI] [PubMed] [Google Scholar]
- 2.Hayashi Y, Majewska AK. Dendritic spine geometry: functional implication and regulation. Neuron. 2005;46:529–532. doi: 10.1016/j.neuron.2005.05.006. [DOI] [PubMed] [Google Scholar]
- 3.Bonhoeffer T, Yuste R. Spine motility. Phenomenology, mechanisms, and function. Neuron. 2002;35:1019–1027. doi: 10.1016/s0896-6273(02)00906-6. [DOI] [PubMed] [Google Scholar]
- 4.Bourne J, Harris KM. Do thin spines learn to be mushroom spines that remember? Curr Opin Neurobiol. 2007;17:381–386. doi: 10.1016/j.conb.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 5.Ethell IM, Pasquale EB. Molecular mechanisms of dendritic spine development and remodeling. Prog Neurobiol. 2005;75:161–205. doi: 10.1016/j.pneurobio.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 6.Lippman J, Dunaevsky A. Dendritic spine morphogenesis and plasticity. J Neurobiol. 2005;64:47–57. doi: 10.1002/neu.20149. [DOI] [PubMed] [Google Scholar]
- 7.Tada T, Sheng M. Molecular mechanisms of dendritic spine morphogenesis. Curr Opin Neurobiol. 2006;16:95–101. doi: 10.1016/j.conb.2005.12.001. [DOI] [PubMed] [Google Scholar]
- 8.Bhatt DH, Zhang S, Gan WB. Dendritic spine dynamics. Annu Rev Physiol. 2009;71:261–282. doi: 10.1146/annurev.physiol.010908.163140. [DOI] [PubMed] [Google Scholar]
- 9.Zuo Y, Lin A, Chang P, Gan WB. Development of long-term dendritic spine stability in diverse regions of cerebral cortex. Neuron. 2005;46:181–189. doi: 10.1016/j.neuron.2005.04.001. [DOI] [PubMed] [Google Scholar]
- 10.Holtmaat AJ, Trachtenberg JT, Wilbrecht L, Shepherd GM, Zhang X, Knott GW, Svoboda K. Transient and persistent dendritic spines in the neocortex in vivo. Neuron. 2005;45:279–291. doi: 10.1016/j.neuron.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 11.Sheng M, Hoogenraad CC. The postsynaptic architecture of excitatory synapses: a more quantitative view. Annu Rev Biochem. 2007;76:823–847. doi: 10.1146/annurev.biochem.76.060805.160029. [DOI] [PubMed] [Google Scholar]
- 12.Bourne JN, Harris KM. Balancing structure and function at hippocampal dendritic spines. Annu Rev Neurosci. 2008;31:47–67. doi: 10.1146/annurev.neuro.31.060407.125646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hotulainen P, Hoogenraad CC. Actin in dendritic spines: connecting dynamics to function. J Cell Biol. 2010;189:619–629. doi: 10.1083/jcb.201003008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lin W-H, Webb DJ. Actin and Actin-Binding Proteins: Masters of Dendritic Spine Formation, Morphology, and Function. Open Neurosci J. 2009;3:54–66. doi: 10.2174/1874082000903020054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Okada H, Soderling SH. Signaling Through Actin to Regulate Spine Formation and Function. Open Neurosci J. 2009;3:97–107. [Google Scholar]
- 16.Penzes P, Jones KA. Dendritic spine dynamics--a key role for kalirin-7. Trends Neurosci. 2008;31:419–427. doi: 10.1016/j.tins.2008.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pontrello CG, Ethell IM. Accelerators, Brakes, and Gears of Actin Dynamics in Dendritic Spines. Open Neurosci J. 2009;3:67–86. doi: 10.2174/1874082000903020067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vicente-Manzanares M, Hodges J, Horwitz AR. Dendritic Spines: Similarities with Protrusions and Adhesions in Migrating Cells. Open Neurosci J. 2009;3:87–96. doi: 10.2174/1874082000903020087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Caceres A, Payne MR, Binder LI, Steward O. Immunocytochemical localization of actin and microtubule-associated protein MAP2 in dendritic spines. Proc Natl Acad Sci U S A. 1983;80:1738–1742. doi: 10.1073/pnas.80.6.1738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Matus A, Ackermann M, Pehling G, Byers HR, Fujiwara K. High actin concentrations in brain dendritic spines and postsynaptic densities. Proc Natl Acad Sci U S A. 1982;79:7590–7594. doi: 10.1073/pnas.79.23.7590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fifkova E, Delay RJ. Cytoplasmic actin in neuronal processes as a possible mediator of synaptic plasticity. J Cell Biol. 1982;95:345–350. doi: 10.1083/jcb.95.1.345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *22.Korobova F, Svitkina T. Molecular architecture of synaptic actin cytoskeleton in hippocampal neurons reveals a mechanism of dendritic spine morphogenesis. Mol Biol Cell. 2010;21:165–176. doi: 10.1091/mbc.E09-07-0596. A study using platinum replica electron microscopy for the first time on mature cultures of hippocampal neurons. The authors show that dendritic filopodia are structurally different from filopodia in other cell types. Instead of being comprised of parallel bundles of actin filaments, dendritic filopodia are formed from a network of branched and linear actin filaments. This actin network matures into a spine through expansion via an Arp2/3 dependent mechanism.
- 23.Star EN, Kwiatkowski DJ, Murthy VN. Rapid turnover of actin in dendritic spines and its regulation by activity. Nat Neurosci. 2002;5:239–246. doi: 10.1038/nn811. [DOI] [PubMed] [Google Scholar]
- 24.Matsuzaki M, Honkura N, Ellis-Davies GC, Kasai H. Structural basis of long-term potentiation in single dendritic spines. Nature. 2004;429:761–766. doi: 10.1038/nature02617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Okamoto K, Nagai T, Miyawaki A, Hayashi Y. Rapid and persistent modulation of actin dynamics regulates postsynaptic reorganization underlying bidirectional plasticity. Nat Neurosci. 2004;7:1104–1112. doi: 10.1038/nn1311. [DOI] [PubMed] [Google Scholar]
- **26.Honkura N, Matsuzaki M, Noguchi J, Ellis-Davies GC, Kasai H. The subspine organization of actin fibers regulates the structure and plasticity of dendritic spines. Neuron. 2008;57:719–729. doi: 10.1016/j.neuron.2008.01.013. An elegant study using two photon photoactivation of GFP-β-actin in hippocampal slice cultures and subsequent live cell imaging of actin dynamics within spines. This was the first study to show f-actin treadmilling or retrograde flow in dendritic spines. The authors defined three populations of actin filaments, one population in the tip of the spine that turned over rapidly and a pool with slower turnover kinetics at the spine base. A third pool, termed the “enlargment pool” of f-actin was induced by local release of caged glutamate and was necessary but not sufficient for the enlargement of spines. To give rise to an enlarged spine this pool had to be confined to the spine head by the spine neck and be coincident with activation of Ca2+/calmodulin-dependent protein kinase II.
- 27.Dent EW, Kalil K. Axon branching requires interactions between dynamic microtubules and actin filaments. J Neurosci. 2001;21:9757–9769. doi: 10.1523/JNEUROSCI.21-24-09757.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dent EW, Kwiatkowski AV, Mebane LM, Philippar U, Barzik M, Rubinson DA, Gupton S, Van Veen JE, Furman C, Zhang J, et al. Filopodia are required for cortical neurite initiation. Nat Cell Biol. 2007;9:1347–1359. doi: 10.1038/ncb1654. [DOI] [PubMed] [Google Scholar]
- 29.Ryu J, Liu L, Wong TP, Wu DC, Burette A, Weinberg R, Wang YT, Sheng M. A critical role for myosin IIb in dendritic spine morphology and synaptic function. Neuron. 2006;49:175–182. doi: 10.1016/j.neuron.2005.12.017. [DOI] [PubMed] [Google Scholar]
- 30.Zhang H, Webb DJ, Asmussen H, Niu S, Horwitz AF. A GIT1/PIX/Rac/PAK signaling module regulates spine morphogenesis and synapse formation through MLC. J Neurosci. 2005;25:3379–3388. doi: 10.1523/JNEUROSCI.3553-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- *31.Hotulainen P, Llano O, Smirnov S, Tanhuanpaa K, Faix J, Rivera C, Lappalainen P. Defining mechanisms of actin polymerization and depolymerization during dendritic spine morphogenesis. J Cell Biol. 2009;185:323–339. doi: 10.1083/jcb.200809046. A comprehensive study of the mechanisms of actin dynamics in cultured hippocampal dendritic spines. Using a actin barbed end labeling assay the authors show that actin polymerization occurs both at the tips and base of dendritic filopodia, however actin barbed ends are localized to the distal regions of spines only. The authors determine the contribution of a number of actin-associated proteins, including mDia, Rif, Arp2/3 and cofilin, in spine architecture and dynamics.
- *32.Tatavarty V, Kim EJ, Rodionov V, Yu J. Investigating sub-spine actin dynamics in rat hippocampal neurons with super-resolution optical imaging. PLoS ONE. 2009;4:e7724. doi: 10.1371/journal.pone.0007724. The first study to use single molecule imaging of actin in dendritic spines. This study confirms previous fixed cell and live cell imaging of actin filaments in spines and concludes that actin dynamics are complex within spines because there is not a coordinated treadmilling of many actin filaments as documented in fibroblast or growth cone lamellipodia. Rather, actin filaments are arranged in a heterogeneous array of short filaments.
- 33.Betzig E, Patterson GH, Sougrat R, Lindwasser OW, Olenych S, Bonifacino JS, Davidson MW, Lippincott-Schwartz J, Hess HF. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 2006;313:1642–1645. doi: 10.1126/science.1127344. [DOI] [PubMed] [Google Scholar]
- 34.Hess ST, Girirajan TP, Mason MD. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys J. 2006;91:4258–4272. doi: 10.1529/biophysj.106.091116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **35.Frost NA, Shroff H, Kong H, Betzig E, Blanpied TA. Single-molecule discrimination of discrete perisynaptic and distributed sites of actin filament assembly within dendritic spines. Neuron. 2010;67 doi: 10.1016/j.neuron.2010.05.026. In press. An elegant study using single particle tracking photoactivated localization microscopy (sptPALM) to image single actin molecules in living cells to construct comprehensive velocity maps of actin filament flow within dendritic spines. The authors find a heterogeneous flow of actin filaments, corroborating the Tatavarty study (32). Interestingly, they find a relatively rapid turnover of actin filaments near the postsynaptic density but not in more lateral regions of spines where the bulk of endocytosis takes place.
- 36.Dent EW, Gertler FB. Cytoskeletal dynamics and transport in growth cone motility and axon guidance. Neuron. 2003;40:209–227. doi: 10.1016/s0896-6273(03)00633-0. [DOI] [PubMed] [Google Scholar]
- 37.Rodriguez OC, Schaefer AW, Mandato CA, Forscher P, Bement WM, Waterman-Storer CM. Conserved microtubule-actin interactions in cell movement and morphogenesis. Nat Cell Biol. 2003;5:599–609. doi: 10.1038/ncb0703-599. [DOI] [PubMed] [Google Scholar]
- 38.Gray EG, Westrum LE, Burgoyne RD, Barron J. Synaptic organisation and neuron microtubule distribution. Cell Tissue Res. 1982;226:579–588. doi: 10.1007/BF00214786. [DOI] [PubMed] [Google Scholar]
- 39.Westrum LE, Gray EG, Burgoyne RD, Barron J. Synaptic development and microtubule organization. Cell Tissue Res. 1983;231:93–102. doi: 10.1007/BF00215777. [DOI] [PubMed] [Google Scholar]
- 40.Westrum LE, Jones DH, Gray EG, Barron J. Microtubules, dendritic spines and spine appratuses. Cell Tissue Res. 1980;208:171–181. doi: 10.1007/BF00234868. [DOI] [PubMed] [Google Scholar]
- 41.Chicurel ME, Harris KM. Three-dimensional analysis of the structure and composition of CA3 branched dendritic spines and their synaptic relationships with mossy fiber boutons in the rat hippocampus. J Comp Neurol. 1992;325:169–182. doi: 10.1002/cne.903250204. [DOI] [PubMed] [Google Scholar]
- 42.Fiala JC, Kirov SA, Feinberg MD, Petrak LJ, George P, Goddard CA, Harris KM. Timing of neuronal and glial ultrastructure disruption during brain slice preparation and recovery in vitro. J Comp Neurol. 2003;465:90–103. doi: 10.1002/cne.10825. [DOI] [PubMed] [Google Scholar]
- 43.Kaech S, Fischer M, Doll T, Matus A. Isoform specificity in the relationship of actin to dendritic spines. J Neurosci. 1997;17:9565–9572. doi: 10.1523/JNEUROSCI.17-24-09565.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kaech S, Parmar H, Roelandse M, Bornmann C, Matus A. Cytoskeletal microdifferentiation: a mechanism for organizing morphological plasticity in dendrites. Proc Natl Acad Sci U S A. 2001;98:7086–7092. doi: 10.1073/pnas.111146798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stepanova T, Slemmer J, Hoogenraad CC, Lansbergen G, Dortland B, De Zeeuw CI, Grosveld F, van Cappellen G, Akhmanova A, Galjart N. Visualization of microtubule growth in cultured neurons via the use of EB3-GFP (end-binding protein 3-green fluorescent protein) J Neurosci. 2003;23:2655–2664. doi: 10.1523/JNEUROSCI.23-07-02655.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- **46.Gu J, Firestein BL, Zheng JQ. Microtubules in dendritic spine development. J Neurosci. 2008;28:12120–12124. doi: 10.1523/JNEUROSCI.2509-08.2008. The first published study to show microtubules present in spines of cultured hippocampal neurons with immunocytochemistry. Intriguingly, only a small percentage (1-5%) of spines contain a microtubule in fixed images. The authors demonstrate that knockdown of EB3, an important microtubule end-binding protein, is sufficient to decrease the numbers of dendritic spines but does not change their length or width. Furthermore, they show that inhibiting microtubule dynamics with low concentrations of nocodazole or stabilizing microtubules with low concentrations of taxol can either inhibit or enhance, respectively, the increase in spine number induced by addition of BDNF.
- **47.Hu X, Viesselmann C, Nam S, Merriam E, Dent EW. Activity-dependent dynamic microtubule invasion of dendritic spines. J Neurosci. 2008;28:13094–13105. doi: 10.1523/JNEUROSCI.3074-08.2008. The first published study to show dynamic microtubule invasion of dendritic spines in cultured hippocampal and cortical neurons. The authors use time-lapse total internal reflection fluorescence microscopy (TIRFM) of EGFP-tubulin and EB3-EGFP to demonstrate that microtubules polymerize into a small subset of dendritic spines at any one time, but when integrated over several hours microtubules can target a substantial (10-20%) number of spines. Interestingly, microtubule entry of spines is transient, averaging only a few minutes at a time. Surprisingly, they also find that a portion of microtubules remain dynamic throughout the time in culture (>DIV60), implying that microtubule dynamics may continue throughout the life of the animal. Furthermore, they demonstrate with KCl infusion that microtubule invasion of spines is enhanced following transient elevations in neuronal activity.
- **48.Jaworski J, Kapitein LC, Gouveia SM, Dortland BR, Wulf PS, Grigoriev I, Camera P, Spangler SA, Di Stefano P, Demmers J, et al. Dynamic microtubules regulate dendritic spine morphology and synaptic plasticity. Neuron. 2009;61:85–100. doi: 10.1016/j.neuron.2008.11.013. An expansive study confirming microtubule entry into hippocampal dendritic spines using EB3-EGFP. The authors also show that EB3 knockdown decreases dendritic spines but increases dendritic filopodia, with no overall change in the overall number of dendritic protrusions. These results conflict with those of Zheng and colleagues (46**). The authors then conduct a number of detailed experiments showing that p140Cap interacts with both the microtubule end-binding protein EB3 and an actin-associated protein, cortactin, implicating a complex interaction of microtubule-associated and actin-associated proteins within dendritic spines.
- *49.Mitsuyama F, Niimi G, Kato K, Hirosawa K, Mikoshiba K, Okuya M, Karagiozov K, Kato Y, Kanno T, Sanoe H, et al. Redistribution of microtubules in dendrites of hippocampal CA1 neurons after tetanic stimulation during long-term potentiation. Ital J Anat Embryol. 2008;113:17–27. An intriguing study showing the presence of microtubules in dendritic spines of CA1 hippocampal dendrites after strong tetanic stimulation (inducing LTP) of adult hippocampal slices. The authors show with electron microscopy, after fixation in a microtubule-stabilizing fixative, that microtubules often associate with the enlarged postsynaptic densities (PSDs) within stimulated spines.
- 50.van Rossum D, Hanisch UK. Cytoskeletal dynamics in dendritic spines: direct modulation by glutamate receptors? Trends Neurosci. 1999;22:290–295. doi: 10.1016/s0166-2236(99)01404-6. [DOI] [PubMed] [Google Scholar]
- 51.Gu J, Zheng JQ. Microtubules in Dendritic Spine Development and Plasticity. Open Neurosci J. 2009;3:128–133. doi: 10.2174/1874082000903020128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang Z, Edwards JG, Riley N, Provance DW, Jr., Karcher R, Li XD, Davison IG, Ikebe M, Mercer JA, Kauer JA, et al. Myosin Vb mobilizes recycling endosomes and AMPA receptors for postsynaptic plasticity. Cell. 2008;135:535–548. doi: 10.1016/j.cell.2008.09.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Correia SS, Bassani S, Brown TC, Lise MF, Backos DS, El-Husseini A, Passafaro M, Esteban JA. Motor protein-dependent transport of AMPA receptors into spines during long-term potentiation. Nat Neurosci. 2008;11:457–466. doi: 10.1038/nn2063. [DOI] [PubMed] [Google Scholar]