

Abstract
Lysosomal and microsomal acid phosphatase activity was estimated in midgut tissue of silkworm larvae, Bombyx mori L. (Lepidoptera: Bombycidae), after induced oxidative stress by D-galactose. The larvae were simultaneously were treated with ethanolic extracts of Bacopa monniera and Lactuca sativa to study their antioxidant properties. Lipid peroxidation and fluorescence was measured to analyze extent of oxidative stress. The ethanolic extract of Lactuca sativa was found to be more effective in protecting membranes against oxidative stress than Bacopa monniera.
Key words : D-galactose, Bacopa monniera and Lactuca sativa
Introduction
Oxidative stress is caused by free radicals such as reactive oxygen species (ROS), which includes superoxide (O2.-), peroxyl, alkoxyl, hydroxyl and nitric oxide. ROS are characterized by presence of an unpaired electron in their outer orbit. In addition to these ROS radicals in living organisms, there are other ROS non-radicals such as the singlet oxygen (1O2), hydrogen peroxide and hypochlorous acid (Pietta, 2000). Small quantities of ROS are formed spontaneously under normal conditions as byproducts of redox processes such as oxidative phosphorylation in the mitochondria and βoxidation of fatty acids. However, the production of ROS is increased when the organism is subjected to irradiation, chemicals or infection (Knapowski et al. 2002). Overproduction of ROS damages cellular lipids, nucleic acids, proteins and leads to lipid peroxidation, genome instability or gene mutation; protein carbonyl formation and enzymatic inactivity resulting in degenerative processes leading to aging (Martin et al. 1996; Berlett and Standtman 1997; Finkel and Halbroook 2000). To defend against the ROS formed, animal cells use three enzymes, superoxide dismutase, catalase and glutathione peroxidase. Superoxide dismutase converts superoxide anion to oxygen and hydrogen peroxide. In biological tissues superoxide can also be converted nonenzymically into the nonradical species hydrogen peroxide and singlet oxygen (Steinbeck et al. 1993). Catalase reduces hydrogen peroxide to water and oxygen (Fridovich 1978). Glutathione peroxidase neutralizes hydrogen peroxide by taking hydrogens from two glutathione molecules resulting in two H2O and one molecule of an oxidized form of glutathione. Besides these cellular antioxidant enzymes there are non-enzyme antioxidants such as α-tocopherol, βcarotin, lycopene and ascorbic acid.
Evidence has accumulated showing that plant polyphenols are an important class of defense antioxidants. These compounds are widespread virtually in all plant foods, often at high levels, and include phenols, phenolic acids, flavonoids, tannins and lignans. Acid phosphatase (ACP) is a lysosomal enzyme that catalyzes the hydrolysis of variety of phosphate monoesters and phosphoprotein in an acidic medium. Phosphatases are capable of transphosphorylation in addition to hydrolysis. Phosphatases thus play an important role in the metabolism of carbohydrates, phospholipids and nucleotides (Hollander 1971; Van Rees 1888; Crossley 1964, 1965; Givindwar and Bhawane 1989, Bhawane and Bhanot 1989, 1990, 1994, Bhanot and Bhawane 1991). Acid phosphatase is found mainly in cytosol of midgut tissue cells of Diptera (Ferreira and Terra 1980) and Lepidoptera (Santos and Terra 1984). ACP activities in the blood and midgut tissues are strongly related to silk protein synthesis, digestion, and absorption of phosphorylating substances in silkworm larvae (Wu 1993). Malondialdehyde and 4-hydroxynonenal are the products of lipid peroxidation. Malondialdehyde and lipofuscin granules are the indicators of oxidative stress and increase with age (Nakano et al. 1995; Sitte et al. 2001).
In present investigation the silkworm Bombyx mori is used to study the effects of D-galactose-induced oxidative stress and the antioxidant properties of ethanolic extracts of Bacopa monniera and Lactuca sativa. Alcoholic extracts of L. sativa and B. monniera contain polyphenols and flavonoids.
It is well known that polyphenols and flavonoids have antioxidant properties. Previous studies suggest that antioxidant polyphenol compounds found in fruits and vegetables may reverse aging (Joseph et al. 2005). L. sativa contains quercetin a flavonol-type flavonoid ubiquitously present in fruits and vegetables (Bilyk and Supers 1985; Zhang, 2005). Quercetin has the antioxidant property of scavenging free radicals such as superoxide radicals, peroxyl and hydroxyl radicals generated in various cellular processes (Dok-Go et al. 2003; Ioku et al. 1995; Kefalas et al. 2003). B. monniera contains mixture of triterpenoid saponins and bacosides (Chatterjee et al. 1963). Previous studies on ethanolic extracts of L. sativa have shown neuroprotective effects by strengthening the antioxidant system of nervous cells in mice (Deshmukh et al. 2007). An ethanolic extract of B. monniera has shown to reduce lipid peroxidation and promotes antioxidant status (Rohini et al. 2004). In insects, the midgut is a dynamic tissue as it is the only section of the alimentary canal where the process of digestion and absorption of digested food takes place. Therefore this is the active organ in insect body organization selected in this study.
Material and Methods
Preparation of extracts
Fresh leaves of Bacopa monniera and Lactuca sativa were collected and washed with distilled water and shade dried. After complete drying, leaves were powdered and kept in alcohol for 72 hours for extraction. The alcohol was allowed to evaporate and the resulting paste was collected and stored for further use.
Silkworm rearing and administration of dose
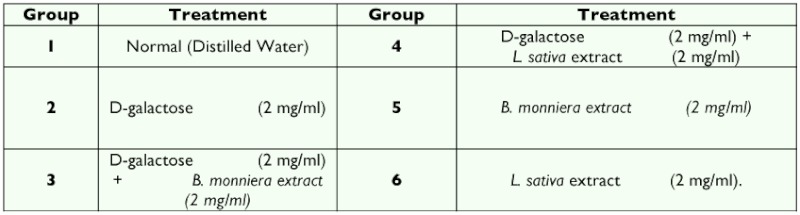
The silkworm B. mori larvae of race PM were reared according to standard methods (Krishanaswami et.al 1978). Treatment was given feeding the fifth instar larvae for the first three days by dipping and air-drying mulberry leaves of equal weight in various solutions of doses prepared by dissolving Dgalactose and ethanolic herbal extracts (Table 1). Control mulberry leaves were dipped in distilled water. The larvae were treated for first three days of fifth instar. On the fourth day, midgut tissue was dissected from larvae.
Table 1.
Groups and treatments.
Sub-cellular fractionation
The midgut tissue was homogenized in 0.25 M sucrose containing 1mM EDTA. The homogenates were subjected to sub-cellular fractionation at 4° C. The homogenate was first centrifuged at 1,000 g to remove unbroken cells and nuclei. The pellet was discarded and the supernatant was again centrifuged at 10,000 g to separate lysosomes and mitochondria. The supernatant was used as microsomal enzyme source. The pellet was resuspended in 0.25M sucrose containing 1mM EDTA that served as lysosomal enzyme source.
Measurement of lipid peroxidation and fluorescence
Lipid peroxidation was estimated using TCA-TBA-HCl reagent. The reaction mixture contained 1 ml homogenate prepared in 0.8% NaCl and 2 ml of TCA-TBA-HCl reagent. The tubes were kept in boiling water bath for 10 min. Tubes were removed and cooled and centrifuged, absorbance was measured at 532 nm using a spectrophotometer. Malondialdehyde was estimated using the extinction coefficient 1.56 × 105/M MDA/cm2.
Fluorescence was measured using the Dillard and Tappel method (1971). One ml of homogenate was extracted at room temperature with 6 ml of chloroform and methanol mixture (2:1) the extracts were centrifuged for two min. at 1500 g. After mixing briefly with 6 ml of water two layers were formed. Four ml of the upper chloroform layer was removed to a test tube and 0.4 ml methanol was added. The fluorescence was determined using a photofluorometer calibrated with quinine sulfate at 1mg/ml.
Estimation of ACP
The acid phosphatase activity was estimated using the Linhardt and Walter method. The assay mixture contained 0.2 ml of enzyme source and 0.8 ml of citrate buffer (pH 4.0) containing 5.5 × 10-3 M p-nitrophenyl phosphate as substrate. The assay tubes were incubated for 30 min. at 40° C. The reaction was stopped by adding 4 ml of 0.1 N NaOH. The absorbance was measured at 405 nm. Zero absorbance was adjusted by control devoid of enzyme source. The protein content in enzyme sources was estimated by the method of Lowry et al. (1951).
Results and Discussion
D-galactose is a reducing sugar that accelerates the process of aging by increasing oxidative stress as was observed in Drosophila melanogaster and Musca domestica (Xu et al. 2004). With age, lysosomes of postmitotic cells increasingly become full with aggregates of oxidized, glycated and crosslinked proteins that are resistant to enzymatic degradation, which is called lipofuscin. Accumulation of lipofuscin is a major manifestation of aging in insects, the concentration of fluorescent age pigment increases with age in tissues (Sohal 1981). The results show a significant increase (p < 0.001) in midgut lipid peroxidation and fluorescent products in D-galactose treated larvae (Figures 1 and 2), indicating the increased oxidative stress due to D-galactose. Lysosomal acid phosphatase activity in midgut tissue of D- galactose treated larvae was decreased significantly, but the microsomal ACP activity was increased significantly (p < 0.001) as compared to the control group (Figure 3).
Figure 1.

Lipid peroxidation in midgut tissue. Data are means ± SD (n = 6). µ*** shows highly significant change (p < 0.001), ** Shows significant change (p<0.05), ns shows non-significant change over the control group. High quality figures are available online.
Figure 2.

Estimation of fluorescent product in midgut tissue. Data are means ± SD (n = 6). *** shows highly significant change (p < 0.001), ** Shows significant change (p< 0.05), ns shows non-significant change over the control group. High quality figures are available online.
Figure 3.

Estimation of ACP activity in lysosomal and microsomal fractions in midgut tissue. Data are means ± SD (n = 6). *** shows highly significant change (p < 0.001), ** Shows significant change (p< 0.05), ns shows non-significant change over the control group. High quality figures are available online.
Cellular organelles are surrounded with lipid membranes that gets damaged due to lipid peroxidation which is result of oxidative stress. Damaged membrane looses its fluidity and integrity. The biological membranes can be disrupted by degrading phospholipids that are the primary molecular component of biological membranes (Tappel 1980; Donato and Sohal 1981). Lysosomal acid phosphatase is transported as a transmembrane protein to dense lysosomes, the pathway of lysosomal ACP to lysosomes include the passage through the plasma membrane (Martin et al. 1989).
Injured lysosomes release hydrolytic enzymes in cytoplasm leading to auto degradation of cellular proteins, damage to endoplasmic reticulum interferes with protein synthesis and intracellular transport of vital compounds (Zs-Nagy 1979 and 2002). Lipid peroxidation in membrane lipids plays an important role in cell physiology and pathology, there are number of membrane bound enzymes and their activity is altered by lipid peroxidation. Lipid peroxidation and fluorescent products were lower in midgut of larvae treated with herbal extract than the D-galactose treated group showing their antioxidant properties. There was no significant difference in fluorescent products as compared with control group. The ethanolic extract of Bacopa monniera was found to be less effective in protecting lipids against lipid peroxidation than the Lactuca sativa extract. Simultaneous treatments with extracts of B. monniera or L. sativa with D-galactose resulted in an increase in the lysosomal ACP activity (Figure 3) than D-galactose treated group, where as the microsomal ACP activity was significantly less.
The results showed that the ethanolic extract of L. sativa was more effective in protecting the lysosomal membrane integrity than B. monniera. Overall lysosomal ACP activity was found to be higher when there is a low level of oxidative stress. At higher levels of oxidative stress the lysosomal membrane was damaged that resulted in an increase in microsomal ACP activity.
Abbreviations
- ACP
acid phosphatase;
- ROS
reactive oxygen species
References
- Berlett SS, Stadtman ER. Protein oxidation in aging, disease, and oxidative stress. Journal of Biological Chemistry. 1997;272:20313–20316. doi: 10.1074/jbc.272.33.20313. [DOI] [PubMed] [Google Scholar]
- Bhanot RK, Bhawane GP. Effect of plant extractives on intestinal phosphatases of the adult, Holotrichia serrata (Coleoptera: Scarabaeidae). Journal of Current Bioscience. 1991;8:39–44. [Google Scholar]
- Bhawane GP, Bhanot RK. Alkaline and acid phosphatases of cockchafers Holotrichia fissa Brinske. Indian Journal of Entomology. 1994;56(4):342–346. [Google Scholar]
- Bhawane GP, Bhanot RK. Insecticidal action of phorate 10 G on the phosphatases in Leucopholis lepidophora BI. Larvae. Journal of Soil Biology and Ecology. 1989;9(12):126–132. [Google Scholar]
- Bhawane GP, Bhanot RK. Studies on phosphatases in different tissues of adult Leucopholis lepidophora BI. (Coleoptera: Scarabaeidae). Indian Journal of Comparative Animal Physiology. 1990;8(1):53–58. [Google Scholar]
- Bilyk A, Supers G. Distribution of Quercetin and Kaempferol in Lettuce, Kale, Chive, Garlic Chive, Leek, Horseradish, Red Radish, and Red Cabbage Tissues. Journal of Agricultural Food Chemistry. 1985;1085(33):226–228. [Google Scholar]
- Chatterjee N, Rastogi RP, Dhar ML. Chemical examination of Bacopa monniera; Wettest Part I: Isolation of chemical constituents. Indian Journal of chemistry. 1963;1:212–215. [Google Scholar]
- Crossley ACS. An experimental analysis of the origins and physiology of haemocytes in the blue blow-fly Calliphora erythrocepliala. Journal of Experimental Zoology. 1964;157:375–398. doi: 10.1002/jez.1401570309. [DOI] [PubMed] [Google Scholar]
- Crossley ACS. Transformations in the abdominal muscles of the blue blow-fly Calliphora erythrocephala (Meig.) during metamorphosis. Embryology and Experimental Morphology. 1965;14:80–110. [PubMed] [Google Scholar]
- Deshmukh AA, Gajare KA, Pillai MM. Neuroprotective effects of lettuce (Lactuca sativa Lin.) in D-galactose induced aging in female albino mice. Journal of Cell and Tissue Research. 2007;7(1):997–1002. [Google Scholar]
- Dok-Go H, Lee KH, Kim HJ, Lee EH, Lee J, Song YS, Lee YH, Jin C, Lee YS, Cho J. Neuroprotective effects of antioxidative flavonoids, quercetin, (1) dihydroquercetin and quercetin 3-methyl ether, isolated from Opuntia ficus-indica var. saboten. Brain Research. 2003;965:130–136. doi: 10.1016/s0006-8993(02)04150-1. [DOI] [PubMed] [Google Scholar]
- Donato S, Sohal RS. Relation of lipofuscin accumulation of aging. In: Florini JR, Adelman RC, Roth GS, editors. CRC handbook of Biochemistry in aging. CRC Press; 1980. pp. 221–227. [Google Scholar]
- Ferreira C, Bellinello GL, Ribeiro AF, Terra WR. Digestive enzymes associated with the glycocalyx microvillar membranes and secretory vesicles from midgut cells of Tenebrio molitor larvae. Insect Biochemistry. 1990;20:839–847. [Google Scholar]
- Ferreira C, Terra WR . Intracellular distribution of hydrolases in midgut caeca cells from an insect with emphasis on plasma membrane-bound enzymes. Comparative Biochemistry and Physiology. 1980;66B:467–473. [Google Scholar]
- Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000;408:239–247. doi: 10.1038/35041687. [DOI] [PubMed] [Google Scholar]
- Fridovich I. The biology of oxygen radicals. Science. 1978;201:875–880. doi: 10.1126/science.210504. [DOI] [PubMed] [Google Scholar]
- Gong GQ, Xu FB. Study of aging model in mice. Journal of China Pharmaceutical University. 1991;22:101–103. [Google Scholar]
- Govindwar SP, Bhawane GP. Effect of BHC on acid and alkaline-phosphatase activity in larvae of Leucopholis lepidophora B1. (Coleoptera: Scarabaeidae). Entomon. 1989;14(1 and 2):45–48. [Google Scholar]
- Hallman PC, Katan MB. Absorption, metabolism and health effects of dietary flavonoids in man. Biomedicine and Pharmacotherapy. 1997;51(8):305–310. doi: 10.1016/s0753-3322(97)88045-6. [DOI] [PubMed] [Google Scholar]
- Huesken BCP, Dejong J, Beekman B, Onderwater RCA. Flavonoids as cardioprotective agents. Cancer Chemathorapy and Pharmacolology. 1955;37:55–62. [Google Scholar]
- Ioku K, Tsushida T, Takeri Y, Nakatani N, Terajo J. Antioxidative activity of quercetin and quercetin monoglucosides in solution and phospholipids bilayers. Biochimica et Biophysica Acta. 1995;1234:99–104. doi: 10.1016/0005-2736(94)00262-n. [DOI] [PubMed] [Google Scholar]
- Kefalas P, Kallithraka S, Parejo I, Makris DP. A comparative study on the in vitro antiradical activity and hydroxyl free radical scavenging activity in aged red wines. Food Science and Technoology International. 2003;9:383–387. [Google Scholar]
- Knopowski J, Wieczorowska-Tobis K, Witowski J. Pathophysiology of aging. Journal of Physiology and Pharmacology. 2002;53(2):135–146. [PubMed] [Google Scholar]
- Krishnaswami S, Narsimhana N, Suryanarayana SK, Kumararaji S. Silkworm rearing, Manual on Sericulture Vol .2. FAO. Agricultural services Bulletin. No. 15. 1978.
- Lowry OH, Rosebrough NJ, Farr AL, Randal RJ . Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry. 1951;193:265–275. [PubMed] [Google Scholar]
- Martin B, Abdul W, Kurt von F. Lysosomal acid phosphatase is transportaed to lysosomes via the cell surface. The EMBO Journal. 1989;8(12):3633–3640. doi: 10.1002/j.1460-2075.1989.tb08537.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin GM, Austad SN, Johnson TE. Genetic analysis of aging: Role of oxidative damage and environmental stresses. Nature Genetics. 1996;13:25–34. doi: 10.1038/ng0596-25. [DOI] [PubMed] [Google Scholar]
- Nakano M, Onzil F, Mizuno T, Gotoh S. Age related changes in lipofuscin accumulation of brain and heart. Gerontology. 1995;41:69–80. doi: 10.1159/000213726. [DOI] [PubMed] [Google Scholar]
- Pietta PG. Flavonoids as antioxidants. Journal of Natural Products. 2000;63:1035–1042. doi: 10.1021/np9904509. [DOI] [PubMed] [Google Scholar]
- Rohini G, Sabitha KE, Shyamala Devi CS. Bacopa monniera Linn. extracts modulates antioxidant and marker enzyme status in fibrosarcoma bearing rats. Indian Journal of Experimental biology. 2004;42:776–780. [PubMed] [Google Scholar]
- Santos CD, Terra WR. Plasma membrane-associated amylase and trypsin: intracellular distribution of digestive enzymes in the midgut of the cassava hornworm, Erinnyis ello. Insect Biochemistry. 1984;14:587–595. [Google Scholar]
- Sitte N, Merker K, Grune T, Zglinicki TV. Lipofuscin accumulation in proliferating fibroblasts in vitro: an indicator of oxidative stress. Experimental Gerontology. 2001;36:475–486. doi: 10.1016/s0531-5565(00)00253-9. [DOI] [PubMed] [Google Scholar]
- Sohal RS. Relationship between metabolic rate, lipofuscin accumulation and lysosomal enzyme activity during aging in Musca domestica. Experimental Gerontology. 1981;16:347–355. doi: 10.1016/0531-5565(81)90055-3. [DOI] [PubMed] [Google Scholar]
- Steinbeck MJ, Khan AU, Karnovsky Mj. Extracellular production of singlet oxygen by stimulated macrophages quantified using 9, 10-diphenylanthracene and perylene in a polystyrene film. Journal of Biological Chemistry. 1993;268:15649–15654. [PubMed] [Google Scholar]
- Tappel AL. Measurement and protection from in vivo lipid peroxidation. In: Pryor WA, editor. Free Radical Biology, IV. Academic Press; 1980. pp. 1–47. [Google Scholar]
- Van Rees J. Beitrage zur Kenntniss der inneren Metamorphose von Musca vomitoria. Zoologische Jahrbücher, Abteilung für Anatomie und Ontogenie. 1888;3:1–134. [Google Scholar]
- Vigh L, Maresca B, Harwood J. Does the membrane's physical state control the expression of heat shock and other genes? Trends in Biochemical Sciences. 1998;23:369–374. doi: 10.1016/s0968-0004(98)01279-1. [DOI] [PubMed] [Google Scholar]
- Xu C, Luning W, Pingping Z, Zhitao H, Zhengyu F, Wenbin L, Jiankang L. D-galactose-caused life shortening in Drosophila melanogaster and Musca domestica is associated with oxidative stress. Biogerontology. 2004;5:317–325. doi: 10.1007/s10522-004-2570-3. [DOI] [PubMed] [Google Scholar]
- Zhang UMC. Protective effect of quercetin on aroclor 1254-induced oxidative damage in cultured chicken spermatogonial cells. Toxicological Sciences. 2005;88(2):545–550. doi: 10.1093/toxsci/kfi333. [DOI] [PubMed] [Google Scholar]
- Zhang X, Zhang BZ, Yang XD, Zhang WW, Li L. Behavior and memory changes in D-galactose-induced aging rat model. Chinese Journal of Gerontology. 1996;16:230–232. [Google Scholar]
- Zs- Nagy Imre. The role of membrane structure and function in cellular aging: A review. Mechanisms of aging and Development. 1979;9(3–4):237–246. doi: 10.1016/0047-6374(79)90102-7. [DOI] [PubMed] [Google Scholar]
- Zs- Nagy Imre. The biological waste product formation in the light of the membrane hypothesis of aging. Archives of Gerontology and Geriatrics. 2002;34(3):329–341. doi: 10.1016/s0167-4943(02)00010-9. [DOI] [PubMed] [Google Scholar]