

Abstract
Cocaine exhibits preferential (∼15-fold) affinity for σ1 over σ2 sigma receptors, and previous research has shown an interaction of σ1 receptor-selective ligands and cocaine's behavioral effects. The present study investigated the effect of the putative sigma receptor agonist SA 4503 (1-(3,4-dimethoxyphenethyl)-4-(3-phenylpropyl)piperazine dihydrochloride) on cocaine's locomotor stimulatory and discriminative stimulus properties. At doses without intrinsic activity, SA 4503 dose-dependently attenuated cocaine-induced hyperactivity in mice. This inhibition was overcome by increasing the cocaine dose. In rats trained to use cocaine as a discriminative stimulus in a drug discrimination task, doses of SA 4503 that did not substitute for the cocaine stimulus did not alter the cocaine substitution curve. However, SA 4503 potentiated the effect of methamphetamine to substitute for the cocaine stimulus. These data support a role for sigma receptors in the locomotor-activating properties of cocaine and, importantly, indicate a role for these receptors in the discriminative stimulus effects of methamphetamine. The data also suggest sigma receptors mediate the activity of different dopamine pathways responsible for the behavioral effects of psychostimulants.
Keywords: Sigma receptors, cocaine, locomotor activity, drug discrimination
1. Introduction
SA 4503 (1-(3.4-dimethoxyphenethyl)-4-(3-phenylpropyl)piperazine dihydrochloride) binds to both σ1 and σ2 sigma receptors, but has greater affinity (∼15-fold) for the σ1 (Ki = 0.004 μM) relative to the σ2 (Ki = 0.06 μM) subtype (Lever et al., 2006). SA 4503 showed no affinity (Ki > 10 μM) for 36 other nervous system targets, including dopamine receptors, excitatory amino acid receptors, second messenger systems (e.g., protein kinase C and inositol triphosphate) and voltage-gated calcium channels (Matsuno et al., 1996), suggesting that SA 4503 exhibits selectivity for sigma receptors over other targets. A more recent study reported that SA 4503 displays affinity (Ki = 0.05 μM) for the vesicular acetylcholine transporter (VAChT) (Kawamura et al., 2006). Regarding its effect on receptor function, SA 4503 exhibits neuropharmacological properties that are similar to known agonists (e.g., pentazocine), suggesting that it is a sigma receptor agonist (Matsuno et al., 1996). However, the lack of a clearly-defined functional assay limits this evaluation.
Cocaine has affinity for sigma receptors, with preferential (∼15-fold) affinity for the σ1 compared to the σ2 sigma receptor subtype (Ki = 2 and 31 μM, respectively) (Matsumoto et al., 2002). The σ1 sigma receptor affinity of cocaine has led to several lines of research to understand the interaction between sigma ligands and the behavioral effects of cocaine. Oral administration of SA 4503 (20 – 40 mg/kg) prolonged (∼64 – 140%) cocaine-induced convulsions in mice, but SA 4503 (1 – 10 mg/kg) failed to alter cocaine-induced hyperactivity in rats (Skuza, 1999). The sigma receptor agonist DTG (1,3-di(2-tolyl)guanidine), which has nearly equal affinity for σ1 and σ2 sigma receptors (Ki = 74 and 61 μM, respectively) (Bowen et al., 1993), potentiated cocaine-induced hyperactivity in rats (Skuza, 1999). DTG did not substitute for cocaine's discriminative stimulus (SD) properties in rats, but shifted the cocaine substitution curve to the left (Ukai et al., 1997). This limited work suggests that sigma receptor agonism enhances the behavioral effects of cocaine; however, the selective role of the σ1 versus the σ2 sigma receptor subtype remains to be determined.
While other studies have shown an interaction of sigma receptor agonists on the locomotor stimulatory and SD properties of cocaine (Skuza, 1999; Ukai et al., 1997), few have investigated the potential role of σ1 sigma receptor agonists on amphetamine's behavioral effects. Repeated SA 4503 administration augmented d-amphetamine-induced hyperactivity in a forced swimming test in rats (Skuza & Rogoz, 2002). Furthermore, σ1 sigma receptor antagonist BMY 14802 attenuated the development of methamphetamine-induced sensitization in rats (Ujike et al., 1992).
The purpose of the present study was to investigate the effect of putative sigma receptor agonist SA 4503 on cocaine's locomotor stimulatory and discriminative stimulus properties, which will inform on the potential role of σ1 sigma receptors on cocaine's effects. The effect of SA 4503 on cocaine-induced hyperactivity in mice was determined to understand the potential role of sigma receptors in this behavior. To understand the role of sigma receptors on cocaine's interoceptive properties, a drug discrimination procedure was used where rats were trained to use cocaine as a SD. SA 4503 substitution tests and tests in which rats were pretreated with SA 4503 followed by cocaine were performed. The effects of SA 4503 on methamphetamine and d-amphetamine substitution also were determined presently.
2. Methods
2.1. Drugs and Chemicals
SA 4503 di-hydrochloride was synthesized by our laboratory as the di-HCl salt and was characterized by reverse phase HPLC (> 97% pure), 1H NMR and 13C NMR. All data were consistent with those obtained with previously synthesized materials (Lever et al., 2006). Cocaine hydrochloride, d-amphetamine sulfate, (±)-methamphetamine hydrochloride and procaine hydrochloride were purchased from Sigma Chemical Co. (St. Louis, MO). All drugs were prepared in saline (0.9% w/v) vehicle and throughout the manuscript, all drug doses refer to the free base weight.
2.2. Locomotor Activity
Male CD-1 mice (Charles River, 20-22 g at arrival) were used for the locomotor activity experiment. Mice were housed 3 or 4 mice per cage with ad libitum access to standard chow and water. The colony was maintained under a 12-hr/12-hr light/dark cycle and the experiments were conducted during the light phase of the cycle. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Missouri.
Preliminary experiments were conducted to determine cocaine's dose-response profile to increase locomotor activity. Mice were acclimated to standard automated activity monitors (Med Associates; Georgia VT) described previously (Polston et al., 2006) for two consecutive days. On the third consecutive day, mice were placed in the monitor for 60 min, injected (i.p.) with cocaine (33, 66 or 99 μmol/kg, [10, 20 or 30 mg/kg]) or saline vehicle, and returned to the activity monitor for 60 min. During the second 60 min period, a dose-dependent increase (saline group mean = 7331 cm, S.E.M. = ±737 cm; 33 μmol/kg cocaine group mean = 17,795 cm, S.E.M. = ±2391 cm; 66 μmol/kg cocaine group mean = 21,632 cm, S.E.M. = ±2762 cm; and 99 μmol/kg cocaine group mean = 39,416 cm, S.E.M. = 5390 cm) in locomotor activity was observed, with peak activity approximately 15 min after cocaine injection. Based on these data, a 66 μmol/kg cocaine dose was used for subsequent experiments.
To determine if SA 4503 alters cocaine-induced hyperactivity, mice were acclimated to the monitors for two consecutive days. On the third consecutive day, mice were placed in monitors for 45 min, injected (i.p) with SA 4503 (2.7, 8.1, or 27 μmol/kg) or saline vehicle, returned to the monitor for 15 min, injected (i.p.) with cocaine (66 μmol/kg) or saline vehicle, and returned to the monitor for 60 min. The experiment's time course was based on previous SA 4503 experiments indicating a relatively-short (∼30 min) period of time before the agonist's behavioral effect was observed (Horan et al., 2001). In a follow-up experiment, mice were injected (i.p) with 8.1 μmol/kg SA 4503 followed by a higher (99 μmol/kg) cocaine dose. Each group contained between 8 and 12 mice.
Distance travelled was collected in 5-min increments over the 120 min behavioral session. A 3-way repeated measures analysis of variance (RM-ANOVA) was performed with SA 4503 Dose (2.7, 8.1, or 27 μmol/kg) and Cocaine Dose (0 and 66 μmol/kg) as between-group factors and Time as a within-subjects factor. A second 3-way ANOVA was performed with SA 4503 Dose (0 and 8.1 μmol/kg), Cocaine Dose (0, 66 and 99 μmol/kg), and Time as factors. Distance travelled during the last 60 min of the session was calculated, and 2-way ANOVA was performed with SA 4503 Dose (0, 2.7, 8.1, or 27 μmol/kg) and Cocaine Dose (0, 66 and/or 99 μmol/kg) as between-group factors. Where appropriate (p < 0.05), Tukey post hoc analyses were performed.
2.3. Drug Discrimination
For the drug discrimination study, male Sprague-Dawley rats (Harlan, Indianapolis, IN; ∼200 g upon arrival to the laboratory) were weighed and handled daily prior to the start of the experiment. Rats had ad libitum access to tap water and given access to a limited amount of standard rat chow (∼25 g) after the completion of the behavioral session. Body weights were maintained at approximately 300 - 350 g.
Standard operant chambers (ENV-001; Med Associates) were used. The chambers had a house light and reinforcers (20 mg food pellets; Bio-Serv, Frenchtown, NJ) were delivered into a recessed receptacle located opposite the house light and between two response levers. Responses made on the active lever were reinforced and responses made on the inactive lever were recorded but not reinforced. All stimulus and response events were controlled and recorded by Med Associates' Med PC-IV Software. Chambers were washed with a soap solution after each rat's session.
Initially, rats (n = 10) were trained to respond on both levers in the operant chambers. Rats were placed in the operant chambers with the house light turned off. The house light was turned on and responses under a fixed-ratio (FR)-1 schedule resulted in reinforcement (delivery of a food pellet). During reinforcement the house light was off for 3 sec and responses were recorded but were not counted toward subsequent reinforcement delivery. For each FR-1 shaping session, rats were reinforced with a maximum of 30 food pellets. FR-1 shaping sessions continued until each rat successfully completed the sessions. Subsequently, the ratio requirement for each shaping session was systematically increased to FR-10. Following the successful completion of the FR-10 shaping sessions, discrimination training commenced.
During the training sessions, rats were injected with cocaine (16 μmol/kg, [5 mg/kg]) or saline, put back into their home cage for 10 min, and then placed in the operant chamber. After administration of cocaine, one lever was active (the opposite lever was inactive) and following saline administration, the other lever was active (the opposite lever was inactive). The cocaine- and saline-paired levers were counterbalanced. The session was terminated after 30 min or 300 responses, whichever occurred first. Only one discrimination training session was administered daily and the presentation of cocaine or saline across sessions was maintained by the following pattern – saline (S), drug (D), D, S, S, D, S, D, S, S, D, D.
Within a discrimination training session, performance was assessed on the first completed ratio. A correct lever selection was recorded when the rat made 10 responses on the active lever with no more than 5 responses on the inactive lever. The criterion for acquisition of the discrimination (stimulus control) for each rat was correct lever selection on 8 out of 10 successive daily sessions.
After all rats had reached stimulus control, substitution test sessions began using a discrete-trials procedure. In each test session only one drug dose and post-injection interval was assessed. In addition, only one test session was completed in a day and at least two training sessions followed each test session. In the test program, both levers were active and 10 responses on either lever delivered a food pellet. The test session was terminated after delivery of food reinforcement or 15 min, whichever occurred first.
In the substitution tests, rats were administered a single dose of cocaine (i.p., 0.01 – 33 μmol/kg), (±)-methamphetamine (s.c., 0.00003 μmol/kg – 3.3 μmol/kg), d-amphetamine (s.c., 0.002 – 2.2 μmol/kg), SA 4503 (i.p., 0.81 – 81 μmol/kg) or procaine (i.p., 4.23 – 127.1 μmol/kg). Rats were returned to the home cage for a 10 min delay, and then placed in the operant chamber when a test session commenced. Additional SA 4503 substitution tests were performed with a 20 min delay between injection time and test session commencement. The 10 min delay for cocaine, methamphetamine, d-amphetamine, and procaine were based on previous studies (Cunningham et al., 2006). The delay periods for SA 4503 were investigated in the current study and the data are presented in Figure 2.
Figure 2.
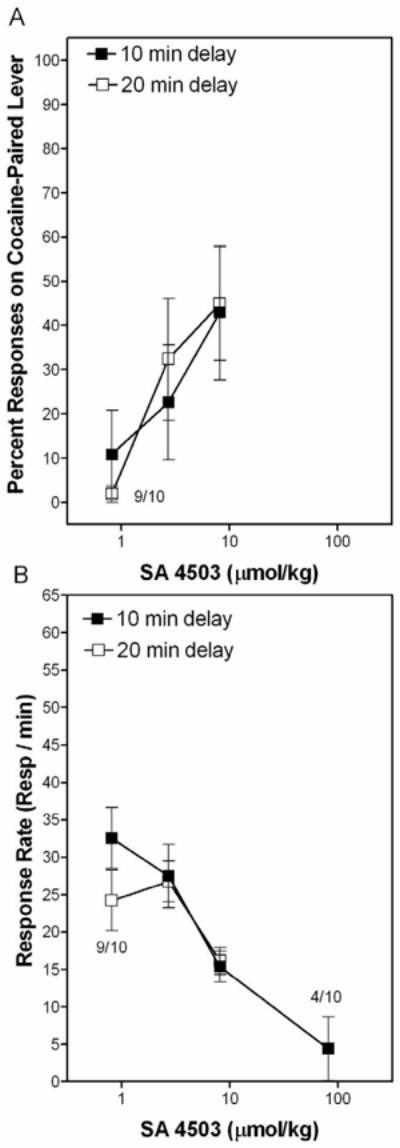
SA 4503 partially substitutes for the cocaine SD. Rats were administered SA 4503, returned to the home cage for a 10 or 20 min delay, and then placed in the chamber where testing commenced. Closed symbols represent data with a 10 min delay and open symbols represent data with a 20 min delay. Data represent mean (±SEM) percent of responses on the cocaine-paired lever (Panel A) and the mean (±SEM) response rates (Panel B). (n = 10 rats/group, except where indicated for a specific drug dose and time point).
In the pretreatment tests, rats were administered a single dose of SA 4503 (0.81 – 2.7 μmol/kg), returned to the home cage for a 10 min delay, injected with either cocaine (0.01 – 16 μmol/kg), methamphetamine (0.00003 μmol/kg – 3.3 μmol/kg), or d-amphetamine (0.002 – 2.2 μmol/kg), returned to the home cage for a 10 min delay, and then placed in the operant chamber when a test session commenced.
For the test sessions, the percent of responses on the cocaine-paired lever and response rates were calculated. “Partial” substitution was defined as 40 - 80% and “full” substitution was defined as > 80% of responses made on the cocaine-paired lever. A nonlinear regression analysis was performed on the mean dose response curve when “full” substitution was observed and an ED50 value was calculated. Dose-response curves were considered to be significantly different when the 95% confidence intervals of their ED50 values did not overlap. Response rates were determined by dividing the total number of responses on both (cocaine- and saline-paired) levers by the session duration. For substitution tests, a one-way RM-ANOVA was performed on response rate data with Drug Dose as the within-subjects factor. For pretreatment tests, a two-way RM-ANOVA was performed on response rate data with Pretreatment Condition and Drug Dose as within-subjects factors. Post hoc paired comparisons were performed when necessary. Animals that exhibited poor inter-test training performance (i.e. did not respond on the appropriate-paired lever during two training sessions consecutively before a test session) were removed from analyses.
3. Results
3.1. Locomotor Activity
Locomotor activity data for mice administered SA 4503 followed by cocaine (66 μmol/kg) are presented in Figure 1. Data analyses revealed a significant 3-way interaction of SA 4503 Dose, Cocaine Dose, and Time [F(69, 622) = 3.22, p<0.05]. Post hoc analyses determined that, as expected, cocaine (66 μmol/kg) produced a significant increase in locomotor activity, relative to mice treated with only saline (compare open symbols in all panels of Figure 1).
Figure 1.
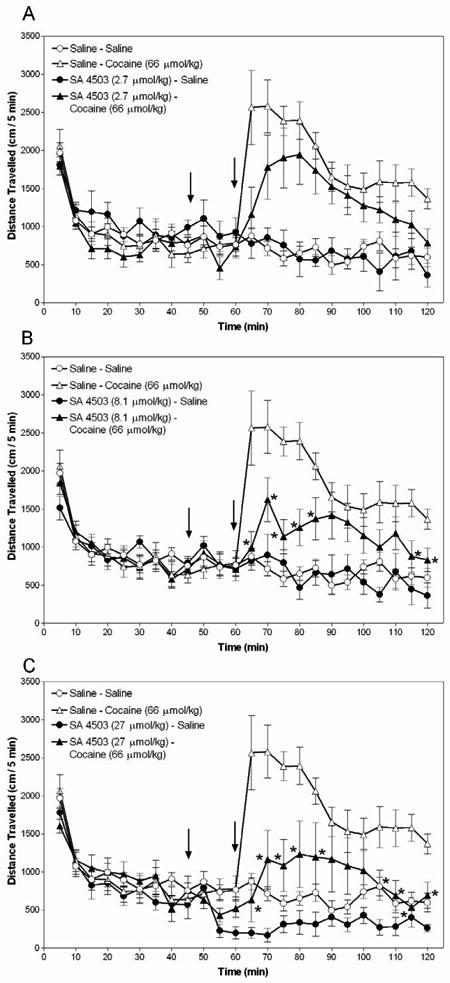
SA 4503 attenuates cocaine-induced hyperactivity. Mice were placed in an automated activity monitor for 45 min, administered SA 4503 (2.7, 8.1 or 27 μmol/kg) or saline, returned to the monitor for 15 min, administered cocaine (66 μmol/kg) or saline, and returned to the monitor for 60 min. Data represent mean (±SEM) distance travelled for the entire 120 min session. Across panels the left arrow designates the first injection (SA 4503 or saline) and the right arrow designates the second injection (cocaine or saline). Open symbols represent data from mice that received saline for the first injection and closed symbols represent data from mice that received 2.7, 8.1 or 27 μmol/kg SA 4503 (Panels A, B and C, respectively) for the first injection. Asterisks designate a significant (p < 0.05) difference from the mice administered saline followed by cocaine.
To determine the effect of SA 4503 on basal activity, groups of mice were treated with SA 4503 or saline vehicle, but did not receive cocaine. For these groups, there were no differences among mice treated with 2.7 or 8.1 μmol/kg SA 4503 and those treated with only saline (compare open and closed circles in Panels A and B of Figure 1). However, mice treated with 27 μmol/kg SA 4503 were less active than saline-treated mice at the 55-75 min time points (compare open and closed circles in Panel C of Figure 1).
Mice were treated with SA 4503 followed by cocaine to determine the interaction between these drugs. There were no significant differences between mice administered 2.7 μmol/kg SA 4503 followed by cocaine (66 μmol/kg), and those administered saline and cocaine at any time point (compare open and closed triangles in Panel A of Figure 1). However, mice treated with 8.1 μmol/kg SA 4503 followed by cocaine were less active than mice that received saline pretreatment at the 65-85, 115 and 120 min time points (compare open and closed triangles in Panel B of Figure 1). Furthermore, differences between mice pretreated with 27 μmol/kg SA 4503 followed by cocaine and those pretreated with saline followed by cocaine were observed at the 65-85 and 105-120 min time points (compare open and closed triangles in Panel C of Figure 1).
SA 4503-induced inhibition of cocaine's effects also was observed with analysis of the cumulative 60 min period after cocaine (66 μmol/kg) injection [F(3, 54) = 4.01, p<0.05]. Mice pretreated with 8.1 μmol/kg SA 4503 (mean = 14,185 cm, S.E.M. = ±2418 cm) and 27 μmol/kg SA 4503 (mean = 11,319 cm, S.E.M. = ±3451 cm) were less active than mice administered saline followed by cocaine (mean = 22,787 cm, S.E.M. = 1881 cm). No differences were observed between mice administered 2.7 μmol/kg SA 4503 (mean = 16,877 cm, S.E.M. = ±2818 cm) followed by cocaine and mice treated with saline and cocaine.
To determine if the SA 4503 (8.1 μmol/kg)-induced inhibition of cocaine's (66 μmol/kg) effects was surmountable, a follow-up experiment was performed in which mice were treated with 8.1 μmol/kg SA 4503 followed by a higher (99 μmol/kg) cocaine dose (data not shown graphically). No significant differences were observed between mice that received 8.1 μmol/kg SA 4503 and 99 μmol/kg cocaine (mean = 36, 318 cm, S.E.M. = ±3187 cm) and those that received saline and this higher cocaine dose (mean = 36,149 cm, S.E.M. = ±5769 cm).
3.2. Drug Discrimination
3.2.1. Training
All rats reached criteria for stimulus control (median = 41 sessions, range = 16-53 sessions).
3.2.2. SA 4503 Tests
Substitution tests were conducted in which rats were administered SA 4503 and placed in the operant chamber 10 or 20 min later. The percent of responses on the cocaine-paired lever and response rates are presented in Panels A and B of Figure 2.
With a 10 min post-injection interval, 8.1 μmol/kg SA 4503 decreased response rates significantly [F(2,17) = 6.42, p<0.05] relative to the saline (0 μmol/kg SA 4503) test session. The 81 μmol/kg SA 4503 dose was omitted from this analysis because the test session was terminated after the first four rats were unresponsive. Partial substitution was evident at the 8.1 μmol/kg SA 4503 dose.
With a 20-min post-injection interval, there were no significant differences in response rates among the SA 4503 doses. Data for one of the ten rats was excluded for the 0.81 μmol/kg SA 4503 dose due to poor inter-test training performance. Partial substitution was evident at the 8.1 μmol/kg SA 4503 dose.
3.2.3. Cocaine Tests
For the drug discrimination tests with cocaine, response rates are presented in Panel D of Figure 3 and the percent responding on the cocaine-paired lever is presented Panel A of Figure 3.
Figure 3.
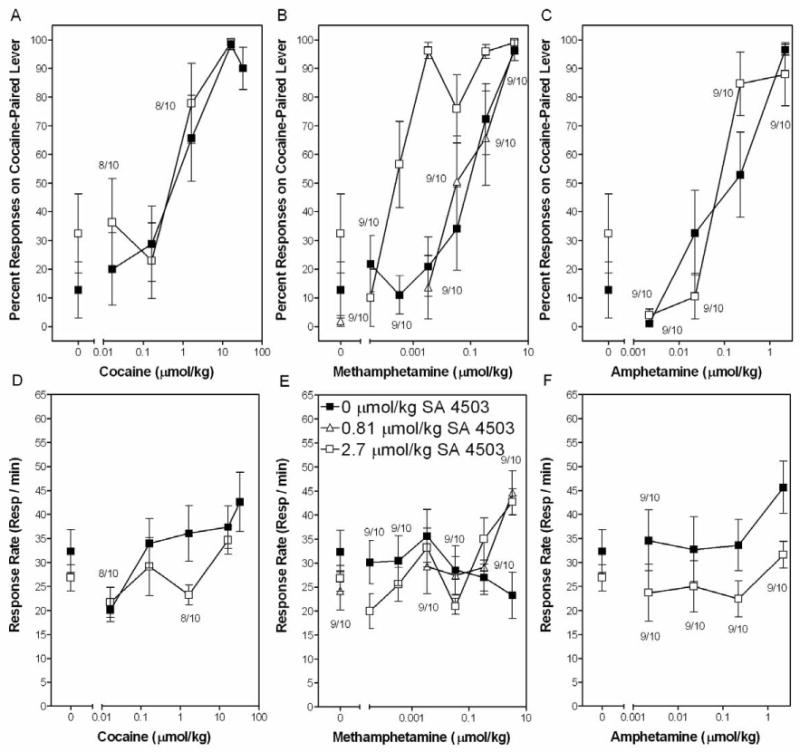
SA 4503 augments the effect of methamphetamine to substitute for the cocaine SD in a dose-dependent manner. Rats were administered SA 4503, returned to the home cage for 10 min, injected with cocaine, methamphetamine, or d-amphetamine and placed in the chamber 10 min later. Closed symbols represent substitution tests where rats were administered cocaine, methamphetamine, or d-amphetamine in the absence of SA 4503. Open symbols represent pretreatment with 0.81 μmol/kg (open triangles) or 2.7 μmol/kg (open squares) SA 4503. Data represent mean (±SEM) percent of responses on the cocaine-paired lever (Panels A-C) and the mean (±SEM) response rates (Panels D-F). (n = 10 rats/group, except where indicated for a specific drug dose and time point).
One the cocaine substitution test, there were no significant differences in response rates among cocaine doses (see closed symbols in Panel D of Figure 3). Regarding the percent of responses on the cocaine-paired lever (see closed symbols in Panel A of Figure 3), full substitution was evident at the 16 and 33 μmol/kg cocaine doses. Partial substitution was evident at the 1.6 μmol/kg cocaine dose. The ED50 value was 0.46 μmol/kg (95% C.I. = 0.16-1.15 μmol/kg).
For the SA 4503 pretreatment tests with cocaine, rats were pretreated with 2.7 μmol/kg SA 4503 (a dose that did not substitute for the SD) prior to cocaine administration. Pretreatment with SA 4503 did not significantly alter response rates, relative to the absence of SA 4503 (compare open and closed symbols in Panel D of Figure 3). Data for two of the ten rats was excluded due to poor inter-test training performance for the 0.016 and 1.6 μmol/kg cocaine doses in the 2.7 μmol/kg SA 4503 pretreatment condition. Regarding the percent of responses on the cocaine-paired lever (see open symbols in Panel A of Figure 3), full substitution was evident at the 16 μmol/kg cocaine dose and partial substitution was evident at the 1.6 μmol/kg cocaine dose in the presence of SA 4503. Analysis of the regression line determined an ED50 value of 0.26 μmol/kg (95% C.I. = 0.06-0.9 μmol/kg). This ED50 value did not differ from the value in the absence of SA 4503 pretreatment (0.46 μmol/kg).
3.2.4. Procaine Tests
None of the procaine doses (4.23, 42.3, and 127.1 μmol/kg) significantly altered response rates (34.46, 33.27, and 28.48 responses/min, respectively) and only partial substitution (26, 39, and 70%, respectively) was achieved (data not shown), indicating that the effect of 16 μmol/kg cocaine to serve as the SD is mediated primarily by the central nervous system (Cunningham et al., 2006).
3.2.5. Methamphetamine Tests
For the drug discrimination tests with methamphetamine, response rates are presented in Panel E of Figure 3 and the percent responding on the cocaine-paired lever is presented Panel B of Figure 3.
On the methamphetamine substitution test, none of the methamphetamine doses significantly altered response rates (see closed symbols in Panel E of Figure 3). Data from one of the ten rats was excluded from the 0.00003 and 0.0003 μmol/kg methamphetamine doses due to poor inter-test training performance. Regarding the percent of responses of the cocaine-paired lever (see closed symbols in Panel B of Figure 3), partial substitution was evident at 0.3 μmol/kg methamphetamine and full substitution was evident at 3.3 μmol/kg methamphetamine. Regression analysis determined an ED50 value of 0.06 μmol/kg (95% C.I. = 0.02-0.13 μmol/kg).
In order to investigate the effect of SA 4503 on the dose-response curve for methamphetamine, rats were pretreated with 0.81 and 2.7 μmol/kg SA 4503 (doses that did not substitute for the SD) prior to methamphetamine administration. Regarding response rates, a significant Methamphetamine Dose × SA 4503 Dose interaction was found [F(3,31) = 3.89, p<0.05]. Post hoc analyses revealed that pretreatment with 0.81 and 2.7 μmol/kg SA 4503 increased response rates at the 3.3 μmol/kg methamphetamine dose, relative to the absence of SA 4503 pretreatment (compare open and closed symbols in Panel E of Figure 3).
Regarding the percent of responses on the cocaine-paired lever (see Panel B of Figure 3), for the 0.81 μmol/kg SA 4503 pretreatment condition, full substitution was evident at the 3.3 μmol/kg methamphetamine dose and partial substitution was evident at the 0.3 and 0.03 μmol/kg methamphetamine doses. Regression analysis determined an ED50 value of 0.05 μmol/kg (95% C.I. = 0.01-0.13 μmol/kg), which was not different from the ED50 value determined in the absence of SA 4503 (0.06 μmol/kg). For the 2.7 μmol/kg SA 4503 pretreatment condition, full substitution was evident at the 0.003, 0.3, and 3.3 μmol/kg methamphetamine doses and partial substitution was evident at the 0.03 and 0.0003 μmol/kg methamphetamine doses. Regression analysis determined an ED50 value of 0.0003 μmol/kg (95% C.I. = 0.00006-0.001 μmol/kg), which was different from the absence of SA 4503, indicating that SA 4503 (2.7 μmol/kg) pretreatment shifted (∼200 fold) the methamphetamine substitution dose-response curve to the left.
3.2.6. d-Amphetamine Tests
For the drug discrimination tests with d-amphetamine, response rates are presented in Panel F of Figure 3 and the percent responding on the cocaine-paired lever is presented Panel C of Figure 3.
On the d-amphetamine substitution test, none of the d-amphetamine doses significantly altered response rates (see closed symbols in Panel F of Figure 3). Only data from nine of the ten rats was included for the 0.002 μmol/kg d-amphetamine dose due to poor inter-test training performance. Regarding the percent of responses on the cocaine-paired lever (see closed symbols in Panel C of Figure 3), full substitution was evident at 2.2 μmol/kg d-amphetamine and partial substitution was evident at 0.2 μmol/kg d-amphetamine. Regression analysis determined an ED50 value of 0.09 μmol/kg (95% C.I. = 0.04-0.2 μmol/kg).
For the SA 4503 pretreatment tests with d-amphetamine, rats were pretreated with 2.7 μmol/kg SA 4503 (a dose that did not substitute for the SD) prior to d-amphetamine. Pretreatment with SA 4503 did not significantly alter response rates, relative to the absence of SA 4503 (compare open and closed symbols in Panel F of Figure 3). For the 2.7 μmol/kg SA 4503 pretreatment condition, data from one of the ten rats was excluded for all d-amphetamine doses due to poor inter-test training performance. Regarding the percent of responses on the cocaine-paired lever (see Panel C of Figure 3), full substitution was evident at the 0.2 and 2.2 μmol/kg d-amphetamine doses in the presence of SA 4503. Analysis of the regression line determined an ED50 value of 0.07 μmol/kg (95% C.I. = 0.03-0.14 μmol/kg), which did not differ from the absence of SA 4503 (0.09 μmol/kg).
4. Discussion
The present study investigated the effect of SA 4503, a putative sigma receptor agonist with preferential (∼15-fold) affinity for σ1 over σ2 sigma receptors, on the behavioral effects of cocaine. SA 4503 attenuated cocaine's locomotor stimulatory properties, but did not alter its interoceptive properties in drug discrimination. Interestingly, SA 4503 dose-dependently shifted the stimulus-substitution curve for methamphetamine to the left by approximately 200-fold. But, SA 4503 did not alter the SD properties of d-amphetamine. This study suggests a complex interaction between these receptors and the locomotor and interoceptive properties of these stimulants.
The present study was not the first to investigate the behavioral effects of SA 4503. As mentioned previously, SA 4503 did not alter cocaine-induced hyperactivity in rats (Skuza, 1999). The discrepancy from the present results may be due to methodological differences between the studies (specifically, oral administration in the previous study and parenteral treatment presently). Recently, our laboratory found that SA 4503 dose-dependently attenuated methamphetamine-induced hyperactivity in rats (Rodvelt, Manuscript under review). Our locomotor activity results support the proposed mechanism by Su and colleagues (2009) in which sigma receptor agonists can facilitate different behavioral consequences depending on the ligand concentration. In other previous reports, nicotine, but not SA 4503, produced significant conditioned place preference (Horan et al., 2001), and SA 4503 pretreatment blocked the acquisition of nicotine-induced conditioned place preference (Horan et al., 2001), suggesting SA 4503 is sufficient to attenuate the conditioned-reinforcing properties of nicotine. Together, the present results and previous research indicate sigma receptors are involved in locomotor activity and drug reinforcement.
Regarding the drug discrimination studies, the cocaine substitution curve was cocaine dose-dependent, and methamphetamine and d-amphetamine fully substituted for the cocaine SD as reported many times in the literature. Interestingly, SA 4503 (0.81 and 2.7 mol/kg) did not substitute for the cocaine SD following either a 10- or 20-min delay. A higher (8.1 mol/kg) SA 4503 dose partially substituted for the cocaine SD, but this dose significantly attenuated response rates, suggesting a nonspecific drug effect. This is in line with a previous study in which the sigma receptor agonist DTG failed to substitute for cocaine in rats trained to discriminate cocaine (33 μmol/kg) from saline (Ukai et al., 1997). In other assays that have probed the psychological properties of sigma receptor agonists, igmesine and PRE-084 (2-(4-morpholinoethyl-1-phenylcyclohexane-1-carboxylate) failed to produce a conditioned place preference (Romieu et al., 2002), supporting the lack of conditioned-reinforcing properties for these sigma receptor agonists. Collectively, these drug discrimination and place preference studies indicate that sigma receptor agonists produce a subjective state different from cocaine. This suggests that binding to sigma receptors (Lever et al., 2006; Matsumoto et al., 2002) is not sufficient for SA 4503 or cocaine to produce an interoceptive state similar to cocaine.
In the SA 4503 pretreatment tests, SA 4503 dose-dependently potentiated the effect of methamphetamine to substitute for the cocaine SD. Pretreatment with 2.7 μmol/kg SA 4503 increased methamphetamine's ED50 value ∼200-fold, compared to the value determined in the absence of SA 4503. These findings indicate that SA 4503 increased the potency of methamphetamine to substitute for the cocaine SD. Although no other studies have shown this enhancement of methamphetamine's effects with sigma receptor agonists, previous studies have shown that sigma receptor agonists can augment the effects of cocaine in operant behavior tasks. As mentioned, DTG shifted (∼2-fold) the cocaine substitution curve to the left (Ukai et al., 1997). Recently, DTG and PRE-084 similarly shifted the cocaine self-administration dose-response curve to the left in a dose-dependent manner (Hiranita et al., 2010).
While SA 4503 augmented methamphetamine's effects, it had no influence on d-amphetamine's substitution for the cocaine SD. One possible explanation is d-amphetamine's lack of affinity (Ki > 10 μM) for σ1 sigma receptors (Walker et al., 1990). The doses of d-amphetamine tested presently were likely well-below d-amphetamine's affinity for sigma receptors. In contrast, methamphetamine has affinity for σ1 sigma receptors (Ki = 2.2 μM) (Nguyen et al., 2005), which may be responsible for the augmentation of methamphetamine's effects on the cocaine SD. Nguyen and colleagues further reported that methamphetamine has much lower (∼21 fold) affinity (Ki = 46 μM) for σ2 sigma than σ1 sigma receptors, suggesting an important role for the σ1 sigma receptor in methamphetamine's SD properties.
Despite the previous studies in which non-subtype selective sigma receptor agonists DTG and PRE-084 augmented the effect of cocaine (Hiranita et al., 2010; Ukai et al., 1997), SA 4503 pretreatment was insufficient to alter the cocaine SD. This could be related to SA 4503's greater (∼15-fold) affinity for the σ1 over the σ2 sigma receptor. Neither DTG nor PRE-084 have the selectivity for σ1over σ2 sigma receptors that is demonstrated by SA 4503 (Bowen et al., 1993; Cobos et al., 2005). While more research is needed, these findings speculate that the σ2 sigma receptor plays a more important role in enhancing cocaine's effects than the σ1 sigma receptor. Future cocaine research with selective σ2 sigma receptor agonists will be important for this research line.
An emerging explanation for the differential effects of SA 4503 on locomotor activity and drug discrimination lies in the intracellular activity of sigma receptor ligands, rather than in a direct agonist/antagonist interaction with plasmalemmel receptors. Intracellular sigma receptors mediate downstream dopamine systems that control locomotor and drug discrimination behaviors. Low and high doses of sigma receptor ligands can produce amplification and inhibition, respectively (Su et al., 2009). High concentrations of sigma receptor ligands may inhibit plasmalemmal ion channels and block downstream dopamine systems such that attenuation may be produced. Low concentrations of sigma receptor ligands may increase intracellular calcium levels and produce amplification of downstream dopamine systems, such that a potentiation may occur. Acute SA 4503 administration increased the number of spontaneously active dopamine neurons in the ventral tegmental area (Minabe et al., 1999), the origination of the mesolimbic dopamine pathway. The mesolimbic dopamine system is likely mediating the SD properties of cocaine (Callahan et al., 1997). In contrast, acute SA 4503 administration significantly decreased the number of spontaneously active dopamine neurons in the substantia nigra (Minabe et al., 1999) which is aligned with earlier work showing pentazocine and DTG inhibit the firing rat of substantia nigra dopamine neurons (Steinfels et al., 1989). These data suggest that sigma receptors play an inhibitory role in the motor effects mediated by dopamine neurons in the nigrostriatal pathway; however, these effects may be reflective of presynaptic neuronal alterations by sigma receptor agonists. The previous and present reports of sigma receptor agonists both potentiating and attenuating cocaine-induced locomotor activity may be the result of postsynaptic sigma receptors in the nigrostriatal dopamine pathway. Cocaine-induced extracellular dopamine release binds postsynaptically to dopamine receptors and facilitates intracellular signaling cascades. Activation of postsynaptic intracellular sigma receptors may be differentially affecting the postsynaptic intracellular signaling cascades produced by dopamine in a concentration-dependent manner. Together, these support a hypothesis that SA 4503 can produce both intracellular amplification and inhibition.
An alternative explanation for the effects of SA 4503 on cocaine's locomotor stimulatory and interoceptive properties is the potential involvement of VAChT. SA 4503 displays high affinity (Ki = 0.05 μM) for VAChT (Kawamura et al., 2006); however, no studies have investigated the role of VAChT on cocaine-associated behaviors. Thus, future research on VAChT-selective ligands and cocaine is needed in order to understand how SA 4503 might modulate cocaine's effects via this mechanism.
In summary, SA 4503 dose-dependently attenuated cocaine-induced hyperactivity and potentiated the effect of methamphetamine to substitute for the cocaine SD. These data suggest a role for sigma receptors in the locomotor activating and SD properties of cocaine, results supported by other work with sigma receptor antagonists. Additionally, the present results indicate a role for these receptors in the discriminative stimulus effects of methamphetamine, where activation of sigma receptors enhances methamphetamine's ability to substitute for a cocaine SD. The data also support electrophysiological studies in which sigma receptors differentially mediate the activity of mesolimbic and nigrostriatal pathways, responsible for drug discrimination and locomotor behaviors, respectively.
Cocaine exhibits affinity for sigma receptors, and previous research has shown an interaction of receptor-selective ligands and cocaine's behavioral effects
The sigma receptor agonist SA 4503 dose-dependently attenuated cocaine-induced hyperactivity in mice.
SA 4503 potentiated the effect of methamphetamine to substitute for the cocaine stimulus in a standard drug discrimination task in rats.
These data support a role for sigma receptors in the locomotor-activating properties of cocaine and, importantly, indicate a role for the σ1 sigma receptor in the behavioral effects of methamphetamine
Acknowledgments
This research was partially supported by a grant from the National Institute on Drug Abuse (DA028477) and by the University of Missouri Department of Psychological Sciences. The authors appreciate the assistance of Clark Oelrichs, Lauren Hediger and David Lefevers in the conduct of this research.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bowen WD, De Costa B, Hellewell SB, Walker JM, Rice KC. [3H]-(+)-Pentazocine: a potent and highly selective benzomorphans-based probe for sigma-1 receptors. Mol Neuropharmacol. 1993;3:117–26. [Google Scholar]
- Callahan PM, De La Garza R, 2nd, Cunningham KA. Mediation of the discriminative stimulus properties of cocaine by mesocorticolimbic dopamine systems. Pharmacol Biochem Behav. 1997;57(3):601–7. doi: 10.1016/s0091-3057(96)00434-0. [DOI] [PubMed] [Google Scholar]
- Cobos EJ, Baeyens JM, Del Pozo E. Phenytoin differentially modulates the affinity of agonist and antagonist ligands for σ1 receptors of guinea pig brain. Synapse. 2005;55(3):192–5. doi: 10.1002/syn.20103. [DOI] [PubMed] [Google Scholar]
- Cunningham CS, Polston JE, Jany JR, Segert IL, Miller DK. Interaction of lobeline and nicotinic receptor ligands with the discriminative stimulus properties of cocaine and amphetamine. Drug Alcohol Depend. 2006;84(3):211–22. doi: 10.1016/j.drugalcdep.2006.01.012. [DOI] [PubMed] [Google Scholar]
- Hiranita T, Soto PL, Tanda G, Katz JL. Reinforcing effects of σ-receptor agonists in rats trained to self-administer cocaine. J Pharmacol Exp Ther. 2010;332(2):515–24. doi: 10.1124/jpet.109.159236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horan B, Gardner EL, Dewey SL, Brodie JD, Ashby CR., Jr The selective σ1 receptor agonist, 1-(3,4-dimethoxyphenethyl)-4-(phenylpropyl)piperazine (SA4503), blocks the acquisition of the conditioned place preference response to (-)-nicotine in rats. Eur J Pharmacol. 2001;426(1-2):R1–2. doi: 10.1016/s0014-2999(01)01229-8. [DOI] [PubMed] [Google Scholar]
- Kawamura K, Shiba K, Tsukada H, Nishiyama S, Mori H, Ishiwata K. Synthesis and evaluation of vesamicol analog (-)-O-[11C]methylvesamicol as a PET ligand for vesicular acetylcholine transporter. Ann Nucl Med. 2006;20(6):417–24. doi: 10.1007/BF03027377. [DOI] [PubMed] [Google Scholar]
- Lever JR, Gustafson JL, Xu R, Allmon RL, Lever SZ. σ1 and σ2 receptor binding affinity and selectivity of SA4503 and fluoroethyl SA4503. Synapse. 2006;59(6):350–8. doi: 10.1002/syn.20253. [DOI] [PubMed] [Google Scholar]
- Matsumoto RR, McCracken KA, Pouw B, Zhang Y, Bowen WD. Involvement of sigma receptors in the behavioral effects of cocaine: evidence from novel ligands and antisense oligodeoxynucleotides. Neuropharmacology. 2002;42(8):1043–55. doi: 10.1016/s0028-3908(02)00056-4. [DOI] [PubMed] [Google Scholar]
- Matsuno K, Nakazawa M, Okamoto K, Kawashima Y, Mita S. Binding properties of SA4503, a novel and selective σ1 receptor agonist. Eur J Pharmacol. 1996;306(1-3):271–9. doi: 10.1016/0014-2999(96)00201-4. [DOI] [PubMed] [Google Scholar]
- Minabe Y, Matsuno K, Ashby CR., Jr Acute and chronic administration of the selective σ1 receptor agonist SA4503 significantly alters the activity of midbrain dopamine neurons in rats: An in vivo electrophysiological study. Synapse. 1999;33(2):129–40. doi: 10.1002/(SICI)1098-2396(199908)33:2<129::AID-SYN3>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Nguyen EC, McCracken KA, Liu Y, Pouw B, Matsumoto RR. Involvement of sigma (σ) receptors in the acute actions of methamphetamine: receptor binding and behavioral studies. Neuropharmacology. 2005;49(5):638–45. doi: 10.1016/j.neuropharm.2005.04.016. [DOI] [PubMed] [Google Scholar]
- Polston JE, Cunningham CS, Rodvelt KR, Miller DK. Lobeline augments and inhibits cocaine-induced hyperactivity in rats. Life Sci. 2006;79(10):981–90. doi: 10.1016/j.lfs.2006.05.006. [DOI] [PubMed] [Google Scholar]
- Rodvelt KR, Oelrichs CE, Blount LR, Fan KH, Lever SZ, Lever JR, Miller DK. The sigma receptor agonist both attenuates and enhances the effects of methamphetamine. doi: 10.1016/j.drugalcdep.2010.12.018. Manuscript under review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romieu P, Phan VL, Martin-Fardon R, Maurice T. Involvement of the sigma1 receptor in cocaine-induced conditioned place preference: possible dependence on dopamine uptake blockade. Neuropsychopharmacology. 2002;26(4):444–55. doi: 10.1016/S0893-133X(01)00391-8. [DOI] [PubMed] [Google Scholar]
- Skuza G. Effect of sigma ligands on the cocaine-induced convulsions in mice. Pol J Pharmacol. 1999;51(6):477–83. [PubMed] [Google Scholar]
- Skuza G, Rogoz Z. A potential antidepressant activity of SA4503, a selective σ1 receptor agonist. Behav Pharmacol. 2002;13(7):537–43. doi: 10.1097/00008877-200211000-00003. [DOI] [PubMed] [Google Scholar]
- Steinfels GF, Tam SW, Cook L. Electrophysiological effects of selective sigma-receptor agonists, antagonists, and the selective phencyclidine receptor agonist MK-801 on midbrain dopamine neurons. Neuropsychopharmacology. 1989;2(3):201–8. doi: 10.1016/0893-133x(89)90023-7. [DOI] [PubMed] [Google Scholar]
- Su TP, Hayashi T, Vaupel DB. When the endogenous hallucinogenic trace amine N,N-dimethyltryptamine meets the σ1 receptor. Sci Signal. 2009;2(61):pe12. doi: 10.1126/scisignal.261pe12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ujike H, Kanzaki A, Okumura K, Akiyama K, Otsuki S. Sigma (σ) antagonist BMY 14802 prevents methamphetamine-induced sensitization. Life Sci. 1992;50(16):PL129–34. doi: 10.1016/0024-3205(92)90466-3. [DOI] [PubMed] [Google Scholar]
- Ukai M, Mori E, Kameyama T. Modulatory effects of morphine, U-50488H and 1,3-di-(2-tolyl)guanidine on cocaine-like discriminative stimulus in the rat using two-choice discrete-trial avoidance paradigm. Methods Find Exp Clin Pharmacol. 1997;19(8):541–6. [PubMed] [Google Scholar]
- Walker JM, Bowen WD, Walker FO, Matsumoto RR, De Costa B, Rice KC. Sigma receptors: biology and function. Pharmacol Rev. 1990;42(4):355–402. [PubMed] [Google Scholar]