

Abstract
Yolk sac and placenta are required to sustain embryonic development in mammals, yet our understanding of the genes and processes that control morphogenesis of these extraembryonic tissues is still limited. The chato mutation disrupts ZFP568, a Krüppel-Associated-Box (KRAB) domain Zinc finger protein, and causes a unique set of extraembryonic malformations, including ruffling of the yolk sac membrane, defective extraembryonic mesoderm morphogenesis and vasculogenesis, failure to close the ectoplacental cavity and incomplete placental development. Phenotypic analysis of chato embryos indicated that ZFP568 does not control proliferation or differentiation of extraembryonic lineages, but rather regulates the morphogenetic events that shape extraembryonic tissues. Analysis of chimeric embryos showed that Zfp568 function is required in embryonic-derived lineages, including the extraembryonic mesoderm. Depleting Zfp568 affects the ability of extraembryonic mesoderm cells to migrate. However, explanted Zfp568 mutant cells could migrate properly when plated on appropriate extracellular matrix conditions. We show that expression of Fibronectin and Indian Hedgehog are reduced in chato mutant yolk sacs. This data suggests that ZFP568 controls the production of secreted factors required to promote morphogenesis of extraembryonic tissues. Our results support previously undescribed roles of the extraembryonic mesoderm in yolk sac morphogenesis and in the closure of the ectoplacental cavity and identify a novel role of ZFP568 in the development of extraembryonic tissues.
Keywords: placenta, yolk sac, chorion, ectoplacental cavity, extraembryonic mesoderm, KRAB-domain protein, mouse
INTRODUCTION
Yolk sac and placenta are extraembryonic tissues that play critical roles in the early development of mammalian species. Although extraembryonic tissues do not contribute to the adult organism, they are critical to sustain embryonic life inside the uterus by providing nourishment and secreting factors that maintain pregnancy (Watson and Cross, 2005). At early embryonic stages, the yolk sac forms a diffusion barrier that mediates the absorption of nutrients directly from the uterine compartment (Jollie, 1990; Rossant and Tam, 2002). Additionally, the yolk sac functions as the first hematopoietic organ, playing a critical role nourishing embryos once they depend on a circulatory system to deliver nutrients (Ferkowicz and Yoder, 2005; Fraser and Baron, 2009). Once embryonic blood circulation is established, the chorio-allantoic placenta (referred onwards as placenta) becomes essential for embryonic survival, as it mediates the exchange of gases, metabolites and waste products between the maternal and fetal circulatory systems. Placental cell types also secrete hormones that maintain pregnancy and impede immunological rejection of the fetus by the maternal immune system (Watson and Cross, 2005). The importance of extraembryonic tissues is underscored by the embryonic growth restriction and early lethality observed in mouse mutants that disrupt yolk sac and placental development (Argraves and Drake, 2005; Baron, 2003; Watson and Cross, 2005).
There are three critical stages in the morphogenesis of extraembryonic tissues: the early specification of trophectoderm (TE) and visceral endoderm (VE) as extraembryonic lineages separate from the embryonic inner cell mass; the contribution of extraembryonic mesoderm during gastrulation; and the subsequent differentiation and morphogenesis of these cell types to form the mature yolk sac and placenta (Rossant and Tam, 2009). Especially relevant is the emergence of the extraembryonic mesoderm, an embryonic-derived cell type that delaminates from the primitive streak and triggers a profound rearrangement in the other extraembryonic lineages (Inman and Downs, 2007). Starting at embryonic day (E) 6.5, mesoderm cells migrate into the extraembryonic region, displacing TE lineages proximally into the conceptus and originating the exocoelomic and ectoplacental cavities. As the extraembryonic mesoderm thins out to line the exocoelom, it apposes the VE and forms the yolk sac. Additionally, the extraembryonic mesoderm contacts TE lineages and forms the chorion, an extraembryonic membrane essential for placental formation (Rossant and Cross, 2001; Watson and Cross, 2005). Completion of placental morphogenesis also requires the contribution of the allantois, another mesoderm-derived structure that extends and fuses with the chorion, becoming the umbilical cord (Inman and Downs, 2007). Simultaneously, the chorion rises and the ectoplacental cavity collapses, causing the apposition of the chorion with the ectoplacental cone, a pre-requisite for the formation of the labyrinth, the vascularized portion of the placenta where maternal and fetal circulation contact (Inman and Downs, 2007; Watson and Cross, 2005).
The study of mouse mutants has greatly contributed to our understanding of the genes and processes that underlie yolk sac and placenta morphogenesis. More than 90 genes have been described to affect the development of these extraembryonic tissues (for a comprehensive review of mouse mutants with placental phenotypes see Inman and Downs, 2007; Watson and Cross, 2005). Some mutations affect differentiation of extraembryonic cell types: for instance, transcription factors Achaete-Scute complex homolog-2 (ASCL2) and Heart and neural crest derivatives expressed transcript-1 (HAND1) regulate the differentiation of trophoblast giant cells from TE stem cell precursors (El-Hashash et al., 2010; Hu and Cross, 2010). Other mutants affect extraembryonic mesoderm cells or their derivatives, as in the case of mutations in Brachyury (T) and Bone morphogenetic protein (Bmp) signaling, which affect extraembryonic mesoderm morphogenesis and the formation of the allantois (Downs et al., 2004; Inman and Downs, 2006; Rashbass et al., 1991; Tremblay et al., 2001; Winnier et al., 1995). Mouse studies have also highlighted the mutual dependence of TE, VE and extraembryonic mesoderm for morphogenesis of extraembryonic tissues, and have identified signaling molecules that mediate these interactions, including Vascular endothelial growth factor (VEGF), Indian Hedgehog (IHH), Fibroblast growth factor (bFGF), Fibronectin (FN) and Transforming growth factor β (TGFβ) (Bohnsack et al., 2004; Byrd et al., 2002; Damert et al., 2002; Flamme and Risau, 1992; Goumans et al., 1999). Also, TE derived lineages secrete polypeptide hormones that impact proliferation and differentiation of the placenta (Rossant and Cross, 2001; Watson and Cross, 2005). Studies on mouse mutants have also identified many other genes that control placental morphogenetic processes such as chorioallantoic attachment, labyrinth branching morphogenesis and labyrinth vascularization (reviewed in Inman and Downs, 2007; Watson and Cross, 2005). These include growth factors, adhesion proteins, transmembrane receptors, protein kinases, transcription factors and epigenetic regulators (Bartholin et al., 2008; Inman and Downs, 2007; Papadaki et al., 2007; Rossant and Cross, 2001; Tian et al., 2009; Watson and Cross, 2005).
KRAB Zinc finger proteins represent one of the largest families of transcriptional regulators in mammals, with more than 300 genes (Urrutia, 2003). However, the biological processes controlled by these proteins have been elusive, mostly due to the lack of mouse mutants in individual KRAB-domain proteins. Here we report the characterization of the extraembryonic defects of chato mouse mutants. chato disrupts ZFP568, a KRAB Zinc finger protein previously described to disrupt mammalian convergent extension (Garcia-Garcia et al., 2008). chato mutants display strong extraembryonic defects, including a ruffling of the yolk sac, defective vasculogenesis and incomplete placental morphogenesis. Analysis with markers of different extraembryonic cell types showed proper specification of extraembryonic lineages in chato embryos, but morphogenetic cellular and tissue rearrangements were disrupted. Zfp568 is expressed ubiquitously throughout mid-gestation, but at high levels in TE-derived tissues, including the ectoplacental cone and the extraembryonic ectoderm (Garcia-Garcia et al., 2008). Analysis of chimeric Zfp568 embryos obtained through tetraploid complementation and through the use of a Cre-responsive Zfp568 reversible allele indicated that Zfp568 is required in embryonic-derived tissues for the morphogenesis of both yolk sac and placenta. chato mutant yolk sacs contained clumped extraembryonic mesoderm cells, indicating that yolk sac and trophoblast defects in chato mutants could be caused by an inability of extraembryonic mesoderm to migrate. We show that chato yolk sac explants migrate and behave similar to wild type when plated in appropriate tissue culture conditions, but their migratory ability is diminished in the absence of Fibronectin. Expression of Fn1 and other factors involved in morphogenesis of extraembryonic tissues is decreased in chato mutant embryos. Taken together, these results indicate that Zfp568 is required in embryonic-derived tissues to provide an environment that supports proper morphogenesis of extraembryonic tissues. Our data attests to the important role of the extraembryonic mesoderm in the morphogenesis of the yolk sac and placenta and defines a requirement of ZFP568 for proper development of these tissues.
MATERIALS AND METHODS
Mouse strains
Extraembryonic phenotypes of Zfp568chato and Zfp568null alleles (Garcia-Garcia et al., 2008) were characterized on C3H/FeJ, CAST/Ei and 129Sv/ImJ. Meox2Cre (Tallquist and Soriano, 2000), Sox2-Cre (Hayashi et al., 2002) and ROSA 26 (Friedrich and Soriano, 1991) mice were obtained from Jackson Laboratory. myr-VENUS mice were obtained from Dr. Kat Hadjantonakis (Rhee et al., 2006). The Zfp568rGT-mutant reversible allele was generated from the German Gene Trap Consortium (GGTC) clone P103E09 (Schnutgen et al., 2005). Complete disruption of Zfp568 splicing in Zfp568rGT-mutant was confirmed by RT-PCR using primers in Zfp568 first and second coding exons. For creation of chimeras, Meox2Cre+/−; Zfp568+/null or Sox2Cre+/−; Zfp568+/null males were crossed to Zfp568rGT-mutant/+ females.
Embryo analysis
Embryos were dissected in 0.4% BSA PBS. Scanning electron microscopy samples were prepared as described (Garcia-Garcia et al., 2008) and imaged with a Hitachi 4500 microscope. In situ hybridizations using DIG-labeled RNA probes were conducted as previously described (Garcia-Garcia and Anderson, 2003; Welsh and O’Brien, 2000). Embryos were imaged in methanol, then processed for 16 μm cryosections. Immunohistochemistry was performed as described (Nagy, 2003) on 8 μm cryosections or whole embryos. Antibodies were anti-phospho-histone H3 (1:200; Upstate); anti-VCAM1 (1:250; BD Biosciences), anti-α4 integrin (1:250; Millipore), anti-Laminin (1:50; Sigma-Aldrich), anti-Fibronectin (1:1000 Sigma-Aldrich), anti-E-Cadherin (1:500 Sigma-Aldrich) and anti-rabbit/anti-rat Alexa Fluor 488 (1:200; Molecular Probes). Alexa Fluor 488 Phalloidin (1:20; Molecular Probes) was used for counterstaining. TUNEL was conducted using the ApopTag Kit (Chemicon) as previously described (Garcia-Garcia et al., 2008). Western blot analysis was performed on yolk sacs from E8.5 embryos using standard protocols and anti-Laminin (1:1000; Sigma-Aldrich), anti-Fibronectin (1:1000; Sigma-Aldrich), anti-GAPDH HRP (1:8000; Abcam) and anti-rabbit HRP (1:10000; Jackson ImmunoResearch) antibodies. Intensity of Western blot bands was quantified with ImageJ. Images of mutant embryos and their respective wild type controls are shown at the same magnification. Expression of molecular markers in chato mutants of different categories was assayed, but has not been reported unless a modification in the expression pattern was detected. Similarly, analysis of proliferation and apoptosis includes results from chato embryos of different classes unless stated. Statistical analysis used two-tailed t-tests and the Prism software (GraphPad).
Tetraploid complementation assays
Generation of Rosa26 lacZ+ Zfp568chato and Zfp568chato/+ ES cell lines, tetraploid complementation experiments and staining for β-galactosidase activity were conducted as described (Nagy, 2003). Similar results were obtained using either of two Zfp568chato ES cell lines.
Cell migration assays
Yolk sacs from GFP+ (myr-VENUS; Rhee et al., 2006) wild type embryos and from chato mutants were dissected at E8.5 and treated with 0.25% trypsin. Trypsinized cells were mixed, then plated onto 5μg/cm2 Fibronectin-coated chambers. 24 hours after plating, confluent cells were scratched and cultured in media containing 5% FBS. Cell migration was assessed after 16 hours. Individually grown yolk sac explants were performed as described (Nagy, 2003) using Lab-Tek II Chamber Slides (Nunc) coated with 5μg/cm2 Fibronectin, 5μg/cm2 Laminin, 0.1% gelatin or without coating. Quantification of migration areas was done at 16 hours using ImageJ. Explants lacking migrating cells at 16 hours (30–60%) were not used to quantify the migratory ability of explants. Differences in explant attachment efficiency between wild type and chato mutant embryos, or between different substrate conditions, were not significant.
qRT-PCR
For quantitative reverse transcription PCR analysis of yolk sac samples, RNA was purified from two independent pools of E8.5 wild type and mutant yolk sacs (RNA STAT-60, Tel-Test). SYBR Green real-time PCR was conducted under standard conditions with cDNA prepared using Superscript III First-Strand Synthesis System (Invitrogen). Calculations were conducted using the 2−ΔΔCT method (Livak and Schmittgen, 2001) with Gapdh as standard. For primer sequences, see Figure S10.
RESULTS
ZFP568 is required for yolk sac morphogenesis
chato is a recessive chemically-induced null mutation in the KRAB domain Zinc Finger protein ZFP568 that disrupts embryo elongation and causes lethality at E9 (Garcia-Garcia et al., 2008). In addition to previously characterized embryonic defects, chato mutants display malformations in extraembryonic tissues. In most chato embryos (96% n=73), the yolk sac is noticeably ruffled by E8.5 (Figure 1A–B; Garcia-Garcia et al., 2008). These yolk sac bubble-like protrusions were observed in embryos as early as E7.5, although at this stage the yolk sac ruffles were not as large and profuse (Figure 1C–D; Garcia-Garcia et al., 2008). We investigated whether chato yolk sac defects originate from abnormalities in the VE or extraembryonic mesoderm, the two cell types that constitute the yolk sac. Analysis of VE markers showed that expression of α-fetoprotein (Afp) (Dziadek and Andrews, 1983), Transthyretin (Ttr) (Barron et al., 1998) and HNF1 Homeobox B (HNF1b) (Coffinier et al., 1999) was unaffected in chato mutants (Figure 1E–H, Figure S1A–D). Likewise, molecular markers of the extraembryonic mesoderm, including Hand1 (Firulli et al., 1998) and the Forkhead box F1a gene (Foxf1) (Mahlapuu et al., 2001), were correctly expressed (Figure 1I–L and not shown). We evaluated whether an overgrowth of the VE layer may underlie the yolk sac ruffling in chato mutants. To this end, we quantified the number of mitotic cells in sagittal sections using phospho-histone H3 antibodies. The average number of VE mitotic cells in chato mutants at E8.5 (6.48±2.38; Figure S1L–M, blue) was slightly smaller than in wild type littermates (7.97±3.18; Figure S1K & S1M, orange), although the difference was not statistically significant (p=0.0655). To evaluate whether the ruffled yolk sac could originate from an excess of VE cells due to increased cell survival, we quantified apoptosis using TUNEL. Consistent with previous reports, we found that wild type yolk sac (Goh et al., 1997) and extraembryonic tissues (Gladdy et al., 2006) have very low levels of apoptosis, and that these levels were similarly low in chato mutants (Figure S2). Therefore, our data does not support overproliferation or increased cell survival of the VE as the causes of yolk sac ruffling.
Figure 1. Yolk sac defects in chato embryos.

(A–D) Scanning electron micrographs of E8.5 (A,B) and E7.5 embryos (C,D). (E–L) In situ hybridizations in whole mount embryos (E,F, I,J) and yolk sac sections (G,H, K, L) with Afp (E–H) and Hand1 (I–L) probes in E8.5 wild type (E,G, I, K) and chato (F, H, J, L) embryos. The chato embryos shown in F, H, J and L correspond to the class II category as specified in Figure 3. In this and other figures, the expression of markers in their respective cell types did not change between embryos of different categories unless otherwise noted. Arrowheads in B and D indicate yolk sac ruffles. Arrowhead in H marks compression of VE cells. Red arrowheads in L highlight detachment of VE from extraembryonic mesoderm. Black arrowheads in K, L point to extraembryonic mesoderm cells. Asterisks (I,J) indicate placental expression of Hand1. VE, visceral endoderm. Scale bars represent 100μm in A, B, C, D, E and I; and 50μm in G and K. In this and following figures, pictures of wild type and mutant embryos are taken at the same magnification.
Analysis of chato embryo sections revealed abnormalities in the organization of both VE and extraembryonic mesoderm. In chato embryos, VE cells were compressed at some areas (Figure 1H, black arrowhead) and detached from the underlying extraembryonic mesoderm at other locations (Figure 1L, red arrowheads; Figure S1B, S1D). These defects were not accompanied by abnormal VE epithelial integrity or polarity, as assessed by analysis of Cadherin 1 (CDH1) and actin localization in chato mutants (Figure S1G–J and S1K–L). Integrity and composition of extraembryonic basement membranes were not noticeably changed in chato embryos, as judged from immunohistochemistry analysis of Fibronectin and Laminin (Figure S9). Also, expression of receptors for these extracellular components such as α4-Integrin and β1-Integrin was not affected in chato mutants (Figure S3 and not shown). Similar to wild type embryos, chato mutants contained mesoderm cells lining interiorly the exocoelomic cavity (Figure 1K–L, small arrowheads). However, in chato mutants extraembryonic mesoderm cells clumped at the embryonic-extraembryonic junction (Figure 1H, S1, 2D, 2F). Analysis of Tbx4, a marker of allantoic mesoderm (Chapman et al., 1996), revealed that posteriorly accumulated cells in chato embryos correspond to the allantois, which failed to extend as compared to wild type littermates (Figure 2A–F). In the most severe chato embryos, Tbx4 failed to be expressed (Figure 2E–F). Analysis of markers of vascular and hematopoietic precursors revealed abnormalities in these extraembryonic mesoderm lineages. Immunohistochemistry using Platelet/endothelial cell adhesion molecule-1 (PECAM1) antibodies (Baldwin et al., 1994) showed that in chato mutants the vascular endothelial cells are specified, but the extraembryonic vascular plexus failed to extend throughout the yolk sac and remained restricted to the blood island region (Figure 2G–H). Hematopoietic cells were also specified in chato mutants (Figure 2I–J, Figure S1E–F) as highlighted by expression of the hematopoietic marker βH1globin (McGrath et al., 2003). However, consistent with the failure of the primitive plexus vasculature to extend in chato embryos, they remained confined to the blood island region in chato mutants (Figure 2I–J, arrowhead). Altogether, these results indicate that the yolk sac ruffling in chato embryos could originate from a loss of adhesiveness between VE and extraembryonic mesoderm, and/or by defective extraembryonic mesoderm morphogenesis.
Figure 2. Abnormal extraembryonic mesoderm morphogenesis in chato mutants.

(A–F) Whole mount in situ hybridizations of E8.5 wild type (A,B) and chato (C–F) embryos using Tbx4 probe. (A,C,E) posterior views. (B,D,F) sagittal sections of the corresponding embryos. According to the categories specified in Figure 3, panels D and F are a class I and a class III chato mutant, respectively. (G,H) Immunohistochemistry with anti-PECAM antibodies in E8.5 wild type (G) and chato (H) embryos. Endothelial cells in chato mutants are small and formed a collapsed vascular plexus. (I,J) βH1-globin whole mount in situ hybridizations in E8.5 wild type (I) and chato (J) embryos. Open arrowheads in B, D, F point to allantoic mesoderm. Arrowhead in J indicates accumulation of blood cells at blood islands. Small arrowheads in B, D and F mark the chorion (ch). Images in G and H were acquired with confocal microscopy using a 40× objective.
chato disrupts placental morphogenesis
Morphogenesis of the labyrinth layer of the placenta requires that the allantois grows and makes contact with the chorion (Inman and Downs, 2007; Watson and Cross, 2005). Consistent with the inability of the allantois to extend in chato mutants (Figure 2A–F), we found that chorioallantoic attachment was disrupted in all chato embryos (n>76), preventing labyrinth formation (Figures 3A–D, S3 and S4). In chato mutants containing allantoic buds, adhesion molecules required for chorioallantoic attachment, including Vascular cell adhesion molecule-1 (VCAM1) and its ligand α4-integrin (Gurtner et al., 1995; Kwee et al., 1995; Yang et al., 1995), were localized in allantois and chorionic mesoderm at levels similar to those of wild type littermates (Figure S3). Also, allantoic cell proliferation and cell death were not significantly altered in chato mutants (Figure S3M–P). Altogether, these results indicate that chato impedes chorioallantoic attachment by disrupting either sustained growth of the allantois or the ability of allantoic cells to migrate.
Figure 3. chato mutants display chorionic defects.

(A–D) Sagittal sections of E8.5 wild type (A) and chato (B–D) embryos. Mutants were categorized into class I, II and III based on the severity of chorionic ectoderm expansion. Original pictures of these embryos without labeling signs are shown in Figure S4. (E–F) Immunohistochemistry with anti-phospho-histone H3 antibodies (PH3, red) in E8.5 sagittal sections of the chorion in wild type (E) and chato (F) embryos counterstained with Phalloidin (green). (G) Quantification of the number of PH3 positive cells per section in the chorionic ectoderm of wild type (orange; 4.61±0.33) and chato class II (blue; 5.67±0/64) embryos (p=0.1178). Broken black lines mark the chorion (ch). Broken red lines mark the amnion (amn). Exocoelomic cavity (exc) is colored in blue, ectoplacental cavity (ec) is colored in red. Arrowhead in C highlights the expansion of the chorion. Arrow in D points to the enlarged ectoplacental cavity. Bracket in B indicate the relatively smooth yolk sac of class I chato mutants. Brackets in C and D indicate yolk sac bubbles concentrated at the embryonic-extraembryonic boundary. Brackets in E and F mark the developing labyrinth in a wild type embryo and the flat chorion of a chato mutant, respectively. Error bars indicate s.d.; n=embryos and s=sections quantified. Scale bars represent 100μm in A and 25μm in E.
Analysis of embryonic sections revealed additional malformations in developing placental tissues in chato mutants. The majority of chato embryos contained an enlarged chorion (92% n=73 Figure 3C–D, Figure S4). Depending on the expansion and arrangement of the chorion we established three chato phenotypic groups: In class II chato embryos (63%) the chorion expanded laterally in direct contact with the VE (Figure 3C, Figure S4C, S4F), in some cases reaching the embryonic-extraembryonic boundary (Figure 3C, arrowhead). In class III chato mutants (29%), the chorion failed to rise and the enlarged chorionic ectoderm formed an expanded ectoplacental cavity (Figure 3D, Figure S4D, S4G). Only 8% of chato mutants (class I) contained a normally sized chorion and an ectoplacental cavity that collapsed similar to wild type littermates (Figure 3B).
Expansion of the chorion and failure of the ectoplacental cavity to collapse have not been associated with defects in allantoic growth, as supported by the lack of these defects in mouse mutants that disrupt allantoic extension such as Smad1 (Lechleider et al., 2001) and Cdx2 (Chawengsaksophak et al., 2004). To further investigate the role of Zfp568 in proper morphogenesis of the chorion, we explored whether the expanded chorion in class II and class III chato mutants could originate from overproliferation. We assessed the number of phospho-histone H3 positive cells in sagittal section of the chorionic ectoderm at E8.5, when chorioallantoic attachment had just occurred in wild type embryos (Rossant and Cross, 2001) and the chorionic ectoderm cells could still be morphologically discerned from other placental cell types (bracket in Figure 3E). At this stage, we did not observe an overall dramatic difference in the number of phospho-histone H3 positive cells in the chorion of chato mutants as compared with wild type littermates (Figure 3E–F). To quantify possible differences, we counted the number of phospho-histone H3 positive cells in the flat chorionic ectoderm of class II chato mutants and compared it with the number of TE phospho-histone H3 positive cells closer to the exocoelomic cavity of wild type littermates. Using this method, we found that the chorionic plate of class II chato embryos contained only an average of 1.06 more phospho-histone H3 positive cells than those in wild type littermates (wt=4.61 chato=5.67; Figure 3G). The fact that this difference is not statistically significant, together with the consideration that our quantification method might slightly underestimate the number of proliferating chorionic ectoderm cells in wild type embryos, suggests that chorion proliferation is not substantially increased in chato mutants. As previously indicated, TUNEL analysis in extraembryonic tissues did not reveal different rates of cell apoptosis between wild type and chato littermates (Figure S2), indicating that differences in cell survival do not account for the excess of chorionic ectoderm cells in chato embryos.
We next explored whether the expanded chorionic ectoderm in class II and III chato mutants could originate from an imbalance amongst different TE lineages. The chorion contains trophoblast stem cells that perdure until the last stages of placental morphogenesis and differentiate to produce ectoplacental cone and trophoblast giant cells (Cross, 2000; Hu and Cross, 2010). Conditions that disrupt the differentiation of trophoblast stem cells lead to an expansion of certain cell types at the expense of others (Guillemot et al., 1994; Luo et al., 1997; Riley et al., 1998). However, analysis of molecular markers characteristic of different placental populations by in situ hybridization in whole mount embryos and in sections showed that expression of markers of different placental cell types in chato embryos, regardless of their phenotypic class, was similar to that of wild type littermates (Figure S5). The trophoblast giant cell marker Placental lactogen 1 (Pl1); (Linzer and Fisher, 1999) was correctly expressed in chato mutants (Figure S5A–D, S5O–P). Likewise, markers expressed in the ectoplacental cone and in precursors of the spongiotrophoblast and labyrinth, including Ascl2 (Guillemot et al., 1994) and Trophoblast specific protein alpha (Tpbpa) (Lescisin et al., 1988), were expressed in chato embryos in the appropriate location and in a similar number of cells as in wild type littermates (Figure S5E–L). Also, Glial cells missing homolog 1 (Gcm1), a transcription factor expressed in clusters of chorion cells that mark the sites of future labyrinth branching (Basyuk et al., 1999) was expressed in clusters of cells in the chorion of chato embryos (Figure S5M–N). These results indicate that chato does not influence the differentiation of trophoblast cell types.
Altogether, results from these experiments indicate that ZFP568 does not regulate cell proliferation, cell survival or differentiation of trophoblast lineages. Hypotheses about how the expansion of the chorion might originate in chato mutants are provided in the Discussion section.
The severity of trophoblast defects in chato embryos correlates with that of yolk sac ruffling and extraembryonic mesoderm defects
We observed that the severity of defects in chato trophoblast tissues correlated with that of the yolk sac ruffling and malformations in extraembryonic mesoderm derivatives. In particular, the amount and location of yolk sac ruffles correlated with the extent of chorion expansion. Thus, in chato mutants where the chorion expanded to the embryonic-extraembryonic boundary (class II and III), yolk sac protrusions were substantial and concentrated to a small band at this junction (brackets in Figure 3C–D, Figure S5F), while in embryos with a small chorion expansion (some class II) or no chorion expansion (class I), the yolk sac was smoother and the ruffles were distributed throughout a wider band (brackets in Figure 3B, Figure S5J).
Extraembryonic mesoderm defects in chato embryos also correlated with the yolk sac and chorion phenotypes. In class I chato mutants, clumps of extraembryonic mesoderm were not observed and, even though the vascular plexus was defective, an underdeveloped allantois was present (Figure 2D). However, class II and III chato mutants had more severe extraembryonic mesoderm defects such as presence of clumps of extraembryonic mesoderm (Figure 3C) and complete lack of an extended allantoic bud (Figure 2F, Figure 3D).
In contrast to the correlations between the severity of extraembryonic defects in chato mutants, we found that neither yolk sac ruffling, chorion expansion, nor extraembryonic mesoderm malformations matched up with the severity of chato embryonic phenotypes (not shown; Garcia-Garcia et al., 2008). Taken together, these observations suggest that defects in yolk sac and trophoblast tissues in chato mutants have a common developmental origin, while embryonic malformations likely originate from an independent requirement of ZFP568 in embryonic tissues. Analysis of Zfp568 expression pattern did not resolve what tissues primarily require ZFP568 function, since Zfp568 is expressed in all embryonic and extraembryonic tissues at early developmental stages, albeit with higher expression levels in TE cells (Garcia-Garcia et al., 2008; Figure S6).
ZFP568 is required in embryonic-derived tissues to control morphogenesis of embryonic and extraembryonic tissues
To investigate ZFP568 requirements in embryonic and extraembryonic lineages, we analyzed the phenotype of chimeric embryos obtained through tetraploid complementation assays. By aggregating wild type tetraploid embryos with chato mutant ES cells, we obtained tetraploidwt ↔ ES cellchato chimeras containing wild type VE and TE (Figure 4A, orange) and chato mutant embryonic-derived tissues, including the extraembryonic mesoderm (Figure 4A, blue). We found that tetraploidwt ↔ ES cellchato chimeric embryos (n=12) recapitulated all extraembryonic malformations observed in class II and III chato mutant embryos, including yolk sac ruffling (Figure 4B–C, arrowheads; Figure S4F–G), defective extraembryonic and allantoic mesoderm morphogenesis (Figure 4C, Figure S4F–G), expansion of the chorionic ectoderm (Figure 4C, Figure S4F–G) and failure of the chorion to rise and form the placenta (Figure 4C, arrow, Figure S4G). Additionally, tetraploidwt ↔ ES cellchato chimeric embryos displayed embryonic defects similar to chato mutants (Garcia-Garcia et al., 2008), including failure to undergo axial rotation and convergent extension defects in lateral plate mesoderm and definitive endoderm (Figure 4D–E). In comparison, control experiments using wild type tetraploid embryos and wild type ES cells, produced embryos without developmental defects (n=13, Figure 4F–I, Figure S4E). Therefore, these experiments demonstrate that wild type ZFP568 function in VE and TE tissues is not sufficient to rescue extraembryonic and embryonic malformations in chato mutants and that ZFP568 is required in embryonic-derived tissues to promote both extraembryonic and embryonic morphogenesis.
Figure 4. Tetraploid complementation assays.

(A) Schematic representation of wild type (orange) and chato mutant (blue) cell types in tetraploidwt ↔ ES cellchato chimeric embryos shown in B–E. Tetraploidwt ↔ ES cellchato (B–E) and control tetraploidwt ↔ ES cellwt (F–I) chimeric embryos were processed for Xgal staining (B–C, F–G) or Foxf1 in situ hybridization (D–E, H–I). B, F, D, H; lateral views. E, I; anterior views. C, G; sagittal sections. C′, G′; higher magnifications of boxed areas in C, G. Arrowheads indicate yolk sac ruffles. Arrow in C points to chorion (ch). Brackets in D, H highlight the lateral plate mesoderm. Discontinuous lines in E, I mark the width of the definitive endoderm.
Restoring Zfp568 function in embryonic-derived tissues rescues embryonic and extraembryonic defects of chato mutants
To further confirm ZFP568 requirement in embryonic-derived tissues, we analyzed the phenotype of chimeric embryos reciprocal to those obtained through tetraploid complementation and thus containing Zfp568 mutant VE and TE and wild type embryonic-derived tissues (VE+TEmutant ↔ Em+Exmwt chimeras, Figure 5C). To obtain these embryos, we used a Zfp568 reversible allele in combination with the embryonic-specific Cre lines Meox2-Cre (Tallquist and Soriano, 2000) and Sox2-Cre (Hayashi et al., 2002). The Zfp568 reversible allele (Zfp568rGT-mutant) contains a genetrap insertion that completely disrupts the activity of Zfp568 (Figure 5B), but, upon Cre recombinase expression, the orientation of the genetrap cassette can be inverted, restoring partial function to the Zfp568 locus (Zfp568rGT-restored allele; Figure S7).
Figure 5. Analysis of chimeras with restored Zfp568 expression in embryonic-derived tissues.

Photographs of E8.5 wild type (A) and Zfp568rGT-mutant/null (B) embryos. (C) Schematic representation of the distribution of Zfp568 mutant cells (blue) and cells with restored Zfp568 function (orange) in Meox2Cre; Zfp568 rGT-mutant/null and Sox2Cre; Zfp568 rGT-mutant/null chimeric embryos. (D–F) Photographs of Meox2Cre; Zfp568 rGT-mutant/null embryos. Blue staining in D and F corresponds to Zfp568 and T expression, respectively. Other embryos were not assayed for expression of these markers. (G,H) Quantification of embryos with rescued embryonic (G) and yolk sac phenotypes (H) in Meox2Cre; Zfp568 rGT-mutant/null and Sox2Cre; Zfp568 rGT-mutant/null chimeras. Trophoblast defects could not be assessed in all chimeras, but severe chorionic defects such as those observed in class II and III chato mutants were detected in only 17% (2/12) Sox2Cre; Zfp568 rGT-mutant/null embryos, suggesting that trophoblast defects were restored in some chimeras. Orange bars indicate the % embryos in which phenotypes were rescued. Blue bars represent % embryos with chato-like phenotypes. Note that yolk sacs were smooth in a small percentage of Zfp568rGT-mutant/null embryos without Cre (panel H; orange area in second column from the left; these embryos had a phenotype similar to those shown in panel E). Control embryos include Sox2Cre/+ and Meox2Cre/+ embryos. n = embryos quantified. Arrowheads highlight yolk sac ruffles.
To resolve whether restoring Zfp568 expression in embryonic-derived tissues is sufficient to rescue the extraembryonic and embryonic defects of chato mutants, we analyzed the phenotypes of Meox2-Cre; Zfp568rGT-mutant/null and Sox2-Cre; Zfp568rGT-mutant/null embryos. Both allelic combinations generate VE+TEZfp568-mutant ↔ Em+ExmZfp568-partially rescued chimeras (Figure 5C). The phenotypes of Meox2-Cre; Zfp568rGT-mutant/null and Sox2-Cre; Zfp568rGT-mutant/null chimeric embryos fell into one of the three following categories: some embryos showed all the phenotypic hallmarks of chato mutants (Figure 5F); in other embryos, yolk sac and chorion defects were rescued, but embryos still had chato-like defects in embryonic tissues (Figure 5E); finally, some chimeric embryos had a wild-type appearance in both embryonic and extraembryonic tissues (Figure 5D). The lack of rescue in some chimeric embryos (Figure 5E–F) could be due either to incomplete efficiency of the Cre recombinase in all embryonic cells, or to incomplete rescue of ZFP568 function due to the hypomorphic condition of the Zfp568rGT-restored allele (Figure S7). Quantification of embryos showing rescue of yolk sac and embryonic phenotypes showed that a larger percentage of embryos were rescued in experiments using Sox2-Cre (Figure 5G–H). This result is consistent with the previously reported higher efficiency of Sox2-Cre vs. Meox2-Cre (Hayashi et al., 2002) and confirms that the extent of phenotypic rescue in the chimeras depends, at least partially, on the number of embryonic-derived cells in which ZFP568 function was restored. Regardless of the incomplete Cre efficiency and incomplete restoration of ZFP568 function, the fact that some VE+TEZfp568-mutant ↔ Em+ExmZfp568-partially rescued chimeras were completely wild type in appearance (Figure 5D, G, H) indicates that relatively small levels of Zfp568 expression in embryonic-derived tissues can rescue both embryonic and extraembryonic defects. Additionally, the presence of embryonic defects in some VE+TEZfp568-mutant ↔ Em+ExmZfp568-partially rescued chimeras with rescued yolk sac and placental malformations (Figure 5E) indicates that embryonic defects are not a secondary consequence of the yolk sac and placental malformations in chato mutants.
Together with results from the analysis of tetraploid chimeras, these experiments demonstrate that Zfp568 is not required in the VE or TE for early morphogenesis of the yolk sac, placenta and embryonic tissues. Furthermore, the rescue of extraembryonic, but not embryonic malformations in some VE+TEZfp568-mutant ↔ Em+ExmZfp568-partially rescued chimeras (Figure 5E) suggests that the requirements for Zfp568 in extraembryonic mesoderm might be lower, and separate from the requirements of Zfp568 in embryonic tissues.
chato mutant cells migrate properly in explant assays
Since analysis of chimeric embryos showed that Zfp568 is required in embryonic-derived tissues (including extraembryonic mesoderm), we sought to test whether extraembryonic defects in chato embryos are due to an inability of extraembryonic mesoderm cells to migrate properly. To this end, we compared the migratory ability of wild type and chato mutant cells side-by-side in yolk sac explant assays. Yolk sac tissues from GFP-labelled wild type embryos and chato mutants were individually trypsinized, then mixed. The resultant cell mixture was then plated onto Fibronectin-coated plates and the migratory behavior of wild type and chato mutant cells was evaluated through their ability to populate a scratch as previously described (materials and methods; Figure 6A). If the migratory ability of chato mutant cells were hindered, we anticipated that the wound would be preferentially populated by wild type (GFP+) cells. In our experiments, we observed that, at 16 hours after wounding, the scratch had completely healed and contained a similar number of wild type (GFP+) and chato mutant (GFP−) cells with mesenchymal morphology that were distributed evenly throughout the wounded area (Figure 6B–C, n=7 assays). We did not detect any abnormality in the morphology of chato mutant cells in our tests.
Figure 6. Yolk sac cell migration assays.

(A) Bright field image of a scratch made 24 hours after plating a mix of GFP-positive wild type (green in B and C) and chato mutant cells from yolk sac explants onto Fibronectin-coated slides. (B–C) Phalloidin (red) and DAPI (purple/blue) staining on the same field of cells 16 hours later. C corresponds to the boxed region in B. (D–K) Yolk sacs from E7.5 wild type (D, F, H, J) and Zfp568 mutant (E, G, I, K) embryos were cut into 2–4 fragments, plated (extraembryonic mesoderm face down) onto chambers coated with Fibronectin (D,E) not coated (F,G), coated with Gelatin (H,I) or Laminin (J,K) and imaged 16 hours afterwards. Images shown are representative examples of each category. For plots of all explants analyzed see Figure S8. (L) Quantification of explant migration shown as the area populated by migrating cells from the explant (traced by broken lines in D–K) relative to the size of the explant (dark area in D–K). Broken lines in A indicate edges of scratch. Broken lines in D–K indicate outer edge of migrating cells. Error bars in L represent s.d. between explants. Scale bars represent 100μm in A, D. Orange bars represent wt embryos, blue bars represent Zfp568null embryos.
The lack of migratory defects in chato mutant explants contrasts with the extraembryonic and embryonic phenotypes we observed in chato mutants. As described previously, chato embryos contained clumps of extraembryonic mesoderm accumulated in the embryonic-extraembryonic boundary (Figure 2C–F) and extraembryonic mesoderm derivatives fail to properly rearrange to form the vascular plexus (Figure 2G–H). Additionally, chato mutants fail to undergo convergent extension (Garcia-Garcia et al., 2008). Since these phenotypes indicate that chato regulates cellular rearrangements, we reasoned that Zfp568 might be required to influence the behavior and/or migration of mesenchymal cells in the context of the embryo. Cell migration depends on cues laid down in the extracellular environment, such as growth factors, adhesion molecules and chemotactic cues (Ridley et al., 2003). Hence, it is possible that in our explant assays, chato mutant cells migrated normally because these cues were provided by either the simultaneous presence of wild type cells or the cell culture conditions. To address this possibility, we tested the migratory ability of wild type and chato mutant yolk sac explants when grown separately and under different growth conditions.
We found that different cell culture substrates supported migration from wild type yolk sac explants, although migration was favored when slides were coated with FN (Figure 6D, F, H, J; orange bars in Figure 6L). Previous reports have described similar effects of different extracellular matrix components on cell culture behavior, with particular cell types showing distinct responses for a given protein substrate (Burdsal et al., 1993; Hashimoto et al., 1987; Tzu and Marinkovich, 2008). When plated on FN-coated slides, chato yolk sac explants migrated similar to wild type controls (p=0.41; Figure 6D–E, 6L, Figure S8). However, migration from chato yolk sacs was significantly reduced when plated on non-coated slides or onto slides coated with inactivated Collagen (gelatin) or Laminin (LAM) (Figure 6F–L, Figure S8). These results indicate that in the absence of FN, chato cells do not have the conditions appropriate for cell migration. These results support our previous observation that lack of ZFP568 does not impede cells from migrating in an appropriate environment and suggest that ZFP568 controls the production of specific secreted factors required for cell migration in the context of the embryo.
chato disrupts signaling between VE and extraembryonic mesoderm
Within extraembryonic tissues, a number of molecules secreted to the extracellular environment are required for proper yolk sac morphogenesis. Amongst these, FN1 is known to be produced by extraembryonic mesoderm cells and promote both extraembryonic mesoderm development and VE survival and homeostasis (Bohnsack et al., 2004; Dickson et al., 1995; Goumans et al., 1999; Morikawa and Cserjesi, 2004). Since our explant assays indicated that FN-coating could restore migration from chato mutant explants (Figure 6L–M), we investigated whether FN1 was correctly expressed in chato embryos. We found that, in chato mutants, both FN1 and Laminin A1 (LAMA1)were localized at the basement membranes of extraembryonic and embryonic tissues similar to wild type littermates (Figure S9). However, the levels of FN1 in chato yolk sac extracts were 32.2% those of wild type controls as assayed by Western blotting (Figure 7A). Interestingly, the levels of LAMA1 were similar between chato and wild type samples (Figure 7B), indicating that chato specifically disrupts the production of some, but not all basement membrane components.
Figure 7. Yolk sac signaling is affected in chato mutants.
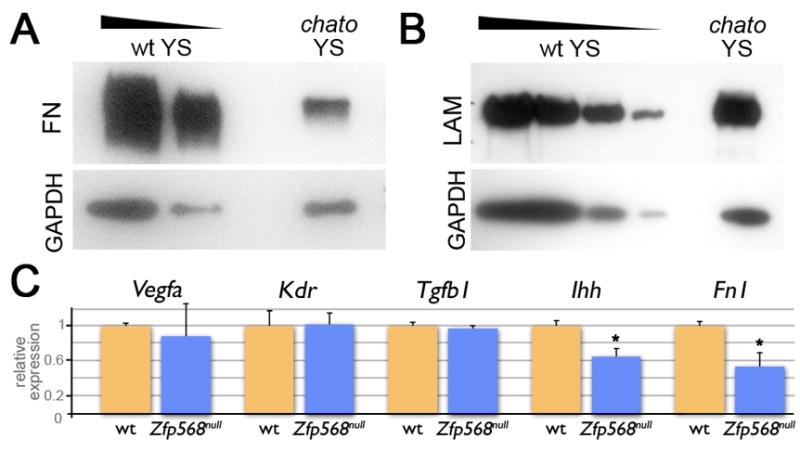
(A–B) Western blots of yolk sac extracts from chato and littermate wild type embryos using anti-Fibronectin (A) and anti-Laminin (B) antibodies. GAPDH was used as a loading control. Fibronectin levels were reduced 32.2% (as quantified from 3 different experiments), while Laminin levels were comparable between wild type and chato mutant extracts (2 experiments) (C) Expression of Vegfa, Kdr, Tgfb1, Ihh and Fn1 in chato (blue) yolk sacs relative to wt (orange) was assessed by qRT-PCR. Error bars represent s.d. between two independent pools of yolk sacs. * indicates p<0.05.
In addition to FN1, other secreted factors have been identified to play roles in yolk sac development. VEGF and IHH, are secreted by the VE and promote differentiation and morphogenesis of the vascular system in the adjacent extraembryonic mesoderm layer, where they are bound by receptors exclusively expressed in these cells (Bohnsack et al., 2004; Breier et al., 1992; Farrington et al., 1997; Yamaguchi et al., 1993). Other proteins promoting yolk sac morphogenesis signal within the extraembryonic mesoderm, as is the case of TGFβ1 (Dickson et al., 1995; Larsson et al., 2001; Oshima et al., 1996). To test whether expression of any of these factors or their receptors is regulated by ZFP568, we quantified their levels of expression in yolk sac tissues of wild type and Zfp568 mutant embryos using qRT-PCR. We found that expression of Vegfa, its receptor Kdr, and Tgfβ1 were not significantly different between wild type and Zfp568 mutant samples (Figure 7C). However, Zfp568 yolk sac tissues had on average reduced levels of Ihh (64%) and Fn1 transcripts (54%) as compared to wild type littermate controls (Figure 7C). The fact that only some of the factors we tested showed abnormal expression in chato mutants excludes the possibility that the chato mutation has a general effect on transcription and, hence, attests of the specificity of these results. Together with the experiments above, these results provide strong evidence that ZFP568 controls the production of specific secreted factors required to promote morphogenesis of extraembryonic tissues.
DISCUSSION
The characterization of chato extraembryonic defects demonstrates a role of the KRAB domain Zinc finger protein ZFP568 in the morphogenesis of both yolk sac and placental tissues. In the yolk sac, ZFP568 is required for extraembryonic mesoderm morphogenesis, vasculogenesis, allantois extension and proper organization of the VE layer. ZFP568 loss of function also affects the development of the placenta. In chato mutants the allantois fails to extend, disrupting choriallantoic attachment. Furthermore, loss of Zfp568 causes expansion of chorionic ectoderm and failure of the ectoplacental cavity to collapse, a process required for proper labyrinth formation. Analysis of molecular markers failed to reveal abnormalities in the specification of extraembryonic lineages in chato mutants (Figures 1–3). Therefore, our data supports that extraembryonic defects in chato embryos arise from abnormalities in the morphogenetic processes that shape these tissues, rather than an imbalance in different extraembryonic cell types.
ZFP568 is expressed ubiquitously throughout embryonic and extraembryonic tissues (Garcia-Garcia et al., 2008). However, the analysis of chato chimeric embryos established that ZFP568 is required exclusively in embryonic-derived tissues for early embryo morphogenesis (Figures 4–5). Because extraembryonic mesoderm originates from embryonic lineages during gastrulation (Lu et al., 2001), it is likely that malformations in yolk sac and trophoblast tissues in chato mutants arise because of defects in the extraembryonic mesoderm. ZFP568 is also required in embryonic-derived tissues to control morphogenetic processes in the embryo, including convergent extension and axial rotation (Figure 4). Analysis of Cre-induced chimeras using a reversible Zfp568 allele showed that restoring Zfp568 function in the embryo and extraembryonic mesoderm, in some cases only rescues defects in yolk sac and trophoblast tissues, but not embryonic morphogenesis (Figure 5). These results support distinct requirements for Zfp568 in extraembryonic mesoderm and embryonic cells to control morphogenesis of extraembryonic and embryonic tissues, respectively. Together with observations that the severity of embryonic defects do not correlate with that of the extraembryonic malformations, these results support independent roles of ZFP568 in the morphogenesis of embryonic and extraembryonic tissues.
chato affects the secretion of extracellular cues required for proper morphogenesis of extraembryonic tissues
chato mutants have strong defects in extraembryonic mesoderm-derived structures such as the primitive vascular plexus and the allantois (Figure 2). Results from the analysis of chimeric embryos support that vasculogenesis and allantoic malformations in chato mutants primarily arise from defects in the extraembryonic mesoderm. Explant assays designed to test the migratory ability of extraembryonic mesoderm cells in chato mutants indicate that loss of Zfp568 does not disrupt the cell machinery required for cell migration (e.g. the cytoskeleton). However, we found that the expression of key molecules required for extraembryonic mesoderm and yolk sac morphogenesis is reduced in chato embryos (Figure 7). Fibronectins are well known for their roles in cell adhesion and migration (Glukhova and Thiery, 1993). Specifically, absence of Fn1 in mouse embryos leads to severe vasculogenesis defects and a separation of the VE from the extraembryonic mesoderm (George et al., 1997). Therefore, the reduced levels of FN1 in chato embryos (Figure 7A & C) might be partially responsible for the abnormal vascular plexus, extraembryonic mesoderm migration and VE adhesion defects in chato mutants. Noteworthy, chato mutant cells migrate properly when explanted onto FN-coated slides, but their migration is reduced when FN is not provided exogenously (Figure 6, Figure S8).
qRT-PCR experiments also revealed downregulation of Ihh levels in chato mutants (Figure 7C). Ihh is required to sustain yolk sac vasculogenesis (Byrd et al., 2002; Dyer et al., 2001). Hence, lower levels of Ihh expression in chato embryos may also contribute to the vascular plexus abnormalities in chato mutants. Interestingly, Ihh is secreted by VE cells (Becker et al., 1997). Since chato is required in embryonic-derived tissues, Ihh downregulation in chato embryos suggests that loss of ZFP568 in the extraembryonic mesoderm indirectly causes malfunctions in the adjacent VE layer. Signaling between the VE and extraembryonic mesoderm is known to be required for proper yolk sac morphogenesis and a number of molecules, including FN1, have been described to mediate these reciprocal interactions (Bohnsack et al., 2004). Therefore, we propose that ZFP568 functions in embryonic-derived tissues to produce factors that signal to adjacent tissues and generate an environment appropriate for extraembryonic mesoderm migration and yolk sac morphogenesis. Comparision of the chato phenotype with that of mutants in Fn1 (George et al., 1997) and Ihh (Byrd et al., 2002; Dyer et al., 2001) suggests that the complexity of extraembryonic defects in chato embryos is unlikely due to the downregulation of just these factors. We presume that expression of additional molecules required for extraembryonic mesoderm migration and yolk sac morphogenesis might be compromised in chato mutants.
chato disrupts morphogenesis of the yolk sac and placenta
chato embryos show severe malformations in non-embryonic-derived tissues, including the yolk sac VE and the chorionic ectoderm. Since chato tetraploid chimeras recapitulate the VE ruffling and chorion abnormalities of chato mutants, it is likely that these phenotypes are caused by extraembryonic mesoderm defects. It is possible that downregulation of Fn1, Ihh and/or additional factors in chato mutants affects morphogenesis of the VE and chorion. Another possibility is that abnormal morphogenesis of extraembryonic mesoderm in chato mutants physically restricts the development of the adjacent VE and TE lineages. Defective genesis (from the primitive streak), migration and/or proliferation of extraembryonic mesoderm might affect the distribution and/or number of cells lining the exocoelomic cavity, which could impair the ability of the VE to extend and in turn cause loss of adhesiveness between these two adjacent layers, originating yolk sac ruffling such as that observed in chato embryos.
chato defects in extraembryonic mesoderm also prevent placental morphogenesis. Mouse mutants in which chorioallantoic fusion is blocked retain a flat chorion and display disrupted labyrinth formation (Gurtner et al., 1995; Hunter et al., 1999; Kwee et al., 1995; Yang et al., 1995). Therefore, failure of the allantois to extend and reach the chorion likely prevents labyrinth formation in chato mutants (Figure 3F). Additionally, defects in extraembryonic mesoderm migration in chato embryos prevent the rising of the chorion and closure of the ectoplacental cavity, processes required for labyrinth formation. Rising of the chorion and closure of the ectoplacental cavity have been described to influence proliferation within the chorionic ectoderm (Uy et al., 2002). Therefore, we hypothesize that delayed or impaired rising of the chorion in chato mutants could lead to a subtle increase in chorionic ectoderm proliferation undetectable by phospho-histone H3 analysis, but sustained over time during development. This sustained proliferation could originate the chorionic phenotypes we observed in class II and class III chato embryos.
While these observations support that extraembryonic mesoderm defects in chato embryos impose a physical constrain in other extraembryonic tissues, mutants that affect extraembryonic mesoderm morphogenesis, such as T or Bmp4 have not been described to cause such severe defects in extraembryonic tissues as chato embryos (Rashbass et al., 1991; Winnier et al., 1995). Therefore, we favor the hypothesis that extraembryonic mesoderm in chato mutants both fails to secrete factors that promote morphogenesis and exerts a physical constrain on adjacent tissues.
ZFP568 has overlapping and divergent roles with factors previously described to regulate morphogenesis of extraembryonic tissues
A number of mutants have been identified that affect morphogenesis of extraembryonic tissues. However, chato mutants present a unique combination of extraembryonic malformations. As previously indicated, mouse mutants that affect extraembryonic mesoderm morphogenesis (Rashbass et al., 1991; Winnier et al., 1995), allantoic extension (Chawengsaksophak et al., 2004; Lechleider et al., 2001) or chorioallantoic attachment (Mahlapuu et al., 2001; Solloway and Robertson, 1999; Xu et al., 1998), have not been described to cause chorion expansion, failure to close the ectoplacental cavity or as severe yolk sac ruffling as we have observed in chato embryos. Amongst other mutants known to disrupt placental morphogenesis (Inman and Downs, 2007; Rossant and Cross, 2001; Watson and Cross, 2005), only mutants in the transcription factors Hand1 (Firulli et al., 1998; Riley et al., 1998; Scott et al., 1999) and Erf (Papadaki et al., 2007) resembled some of the characteristic features of chato mutants.
Similar to chato embryos, Hand1 mutants have yolk sac ruffling and vasculogenesis defects, but Hand1 embryos have an imbalance in the differentiation of TE-derived lineages that causes a reduction in Pl1-positive trophoblast giant cells with a concomitant expansion of the spongiotrophoblast population expressing Acsl2 (Riley et al., 1998), a phenotype that we have not observed in chato embryos (Figure S5). Mutants in the ets transcriptional repressor Erf also show defects remarkably similar to chato, including abnormal vasculogenesis, defective chorioallantoic attachment, failure to close the ectoplacental cavity and an expanded chorion layer (Papadaki et al., 2007). However, Erf mutants do not show yolk sac ruffles and, unlike chato embryos, loss of Erf affects the differentiation of trophoblast stem cells causing a reduction in the number of cells expressing Tpbpa and lack of Gcm1 expression in the chorion at E8.5 (Papadaki et al., 2007). The phenotypic similarities and differences of chato embryos with Hand1 and Erf mutants suggest overlapping, as well as divergent roles for these factors on the morphogenesis of extraembryonic tissues. Future studies will help clarify how ZFP568, HAND1 and ERF coordinately and separately control development of the yolk sac and placenta.
CONCLUSIONS
The biological processes controlled by KRAB Zinc finger proteins have been elusive, mostly due to the lack of mouse models disrupting individual KRAB-domain family members. The study of chato mutants described here identifies a role of Zfp568 in the morphogenesis of extraembryonic tissues and defines a primary requirement for Zfp568 in the extraembryonic mesoderm. Our data supports a requirement for ZFP568 in the production of factors important for cell migration and tissue morphogenesis. The differences between the phenotype of chato embryos with that of mouse mutants previously described to affect yolk sac and placenta development suggest that Zfp568 controls extraembryonic development through distinct molecular mechanisms. Future systems biology approaches will be able to identify the precise targets of ZFP568 ultimately responsible for the morphogenesis of extraembryonic tissues.
RESEARCH HIGHLIGHTS.
the yolk sac in chato mutants has bubble like protrusions
chato disrupts allantoic extension, preventing chorioallantoic attachment
chato causes an expansion of the chorion and failure of the ectoplacental cavity to close
Zfp568 is required in embryonic-derived tissues, including extraembryonic mesoderm
Zfp568 controls the expression of factors required for morphogenesis of extraembryonic tissues
Supplementary Material
(A–F) In situ hybridizations in E8.5 wild type (A, C, E) and chato mutants (B, D, F) with Ttr (A,B) Hnf1b (C,D) and ®H1 globin (E,F) probes. High magnifications of sagittal sections through the yolk sac region are shown. (B) class III and (D, F) class II chato mutants (G–J) Wild type (G) and chato mutant sagittal sections were stained with anti-CDH1 antibody (green) and counterstained with DAPI (blue). (I,J) higher magnifications of boxed areas in G, H respectively. (H, J) show a class II chato mutant. (K–L) Immunohistochemistry with anti-phospho-histone H3 antibodies (PH3, red) counterstained with Phalloidin (green) as shown on a class III chato mutant (M) Quantification of the number of PH3 positive cells per section in the VE of wild type (orange; 7.97±3.18) and chato (blue; 6.48±2.38) embryos (p=0.0655). Error bars indicate s.d.; n=embryos and s=sections quantified. VE, visceral endoderm; amn, amnion; TE, trophectoderm; Exm, extraembryonic mesoderm; ch, chorion. Scale bars represent 25μm.
(A–D) TUNEL (green) and DAPI (blue) staining on sagittal sections of wild type (A) and a class II chato (B) embryo. (C,D) Higher magnifications of boxed regions in A, B. Arrowheads point to TUNEL positive nuclei. (E,F) Quantification of total TUNEL positive cells per section in allantois and yolk sac (E) and chorionic ectoderm (F). Error bars indicate s.d.; n=embryos and s=sections quantified. ys=yolk sac, ch=chorion. Scale bar represents 100μm.
Sagittal sections of E8.5 wild type (A,G) and class II chato (B,H) embryos labeled with anti-VCAM (A,B; green) or anti-α4 integrin (G,H; green) antibodies and DAPI (red) counterstaining. C–F,I–L, are magnifications of boxed regions in A,B,G,H, respectively (E–F,K–L; green channel only). (M–P) Sagittal sections of wild type (M, O) and class II chato mutant embryos (N, P) stained with anti-phospho-histone H3 antibodies (PH3, red) and Phalloidin (green) (M–N) or with TUNEL (green) and DAPI (blue) (O–P). all, allantois; ch, chorion; VE, visceral endoderm. Arrowheads in O,P point to TUNEL positive nuclei in allantois. Scale bars in A,G,M,O represent 100μm.
(A–D) Raw images of the sagittal sections of E8.5 wild type (A) and chato (B–D) embryos shown in Figure 3. (E–G) Sagittal sections of X-gal stained tetraploid chimeras obtained from aggregation of wild type ES cells (control tetraploidwt ↔ ES cellwt chimera) (E) and chato mutant ES cells (tetraploidwt ↔ ES cellchato chimeras) (F,G). We found that chato tetraploid chimeras (n=8) reproduced the characteristic defects of class II (F, 75%) and class III chato mutants (G, 25%). The Rosa26 allele in the ES cell lines used to generate these chimeras served as a lineage marker to differentiate embryonic-derived cells (embryo and extraembryonic mesoderm, Xgal positive) from VE and trophoblast tissues (Xgal negative). (H, I) Whole mount in situ hybridizations with a Fgfr2 probe in wild type (H) and chato embryos (I). amn, amnion; ch, chorion; exc, exocoelomic cavity; ec, ectoplacental cavity. Red arrowheads point to chorion. Double-arrowed line in D points to the enlarged ectoplacental cavity. Brackets in C and D indicate yolk sac bubbles concentrated at the embryonic-extraembryonic boundary. Scale bars in A,E,H represent 100μm.
Whole mount in situ hybridizations of E8.5 wild type (A,E,I, M) and chato mutant (B, F, J, N, P) embryos with Pl1 (A–B, O–Q), Tpbpa (E–F), and Ascl2 (I–J) probes. (C–D, G–H, K–L) Sagittal sections of the trophoblast region of embryos in A–B, E–F, I–J, respectively. The trophoblast populations marked by these probes are indicated in the bottom margin. (M–N) High magnifications of E8.5 wild type (I) and chato mutant (J) sagittal sections showing Gcm1 expression in clusters of chorionic cells (arrowheads). In chato embryos, Gcm1 clusters are smaller. (O–Q) Whole mount in situ hybridizations of E8.5 wild type (O), chato (P) and Hand1 mutant (Q) embryos with a Pl1 probe. Hand1 mutants have a severe reduction in the number of Pl1-expressing trophoblast giant cells (Riley et al., 1998). In contrast, the number of Pl1-expressing trophoblast giant cells in Zfp568null embryos is similar to that of wild type littermates. Brackets in F and J mark yolk sac ruffles. Scale bars represent 100μm.
Sections of whole mount in situ hybridizations of wild type (A–C) and Zfp568null (D) embryos at E8.0 (A) and E8.5 (B,C,D) with a Zfp568 probe. Zfp568 expression was not detected in control Zfp568null embryos (D). bc, blood cells; exc, exocoelomic cavity; TE, trophectoderm; VE, visceral endoderm; Exm, extraembryonic mesoderm. Scale bars represents 100μm.
(A) The Zfp568 reversible allele (Zfp568rGT-mutant) was generated from a German Gene Trap Consortium ES cell clone containing a FlipROSAbgeo genetrap vector inserted into the third intron of the Zfp568 locus. The genetrap cassette in Zfp568rGT-mutant disrupted the Zfp568 transcript and generated a truncated ZFP568 protein fused to βgeo (http://tikus.gsf.de/ggtc/). This truncated protein, which originated from splicing between the second exon of Zfp568 and the splicing acceptor site in the genetrap cassette, contained only 11 N-terminal amino acids of ZFP568 and lacked all ZFP568 functional domains (KRAB domains and Zinc finger motifs). Complementation tests between Zfp568rGT-mutant, Zfp568chato and Zfp568null, as well as RT-PCR experiments, all confirmed that Zfp568rGT-mutant genetrap insertion generates a null allele for Zfp568. The reversible capabilities of Zfp568rGT-mutant reside in the loxP and FRT recombination sites (red and green circles) flanking the FlipROSAβgeo genetrap cassette. These sites mediate the inversion of the genetrap cassette when Cre or FLP recombinases are present and release the effect of the genetrap cassette on Zfp568, therefore restoring wild type function to the Zfp568 locus. (B–D) Complementation tests between Zfp568rGT-restored and Zfp568null alleles revealed that 7 out of 13 Zfp568rGT-restored/null embryos had a smooth yolk sac and wild type embryo morphology at E9.5 (C), whereas 6/13 embryos (D) had all the characteristic defects of chato mutants, including yolk sac ruffling. All other littermates were wild type in appearance (B). These results suggested that the inversion of the cassette did not fully restore wild type ZFP568 function and that Zfp568rGT-restored was likely a hypomorphic allele of Zfp568. Because the targeted insertion of vectors in intronic sequences has been previously reported to affect gene transcription (Meyers et al., 1998), we hypothesized that the presence of the genetrap allele in Zfp568rGT-restored, even in the wild type configuration, interferes with the normal splicing of the Zfp568 transcript. (E) Quantification of the levels of splicing through the two exons flanking the genetrap insertion site using qRT-PCR on E9.5 embryonic tissues showed that Zfp568rGT-restored/+ embryos have a 40% reduction in the levels of full length Zfp568 transcript. Zfp568rGT-restored/null embryos only express 10% the level of full length transcript compared to wild type embryos. These results confirmed Zfp568rGT-restored is a hypomorphic allele of Zfp568. ***p=0.0007 compared to wt. Error bars represent s.d. SA, splice acceptor; pA, polyadenylation signal.
(A–C) Graphic representation of results from yolk sac explant assays plated onto Lab-Tek™ II (Nunc) slides coated with Fibronectin (FN), no coating, Gelatin, or Laminin (LAM). The regions containing migrating cells for each wt (A, grey) and Zfp568 mutant (B, pink) explant were represented as shaded areas on different layers at 30% transparency, then scaled so that the size of the initial explant size (central white area) was constant across all explants. Darker areas represent regions most frequently populated by migrating cells. Overlays of wt and Zfp568 mutant explants (C) highlight the reduced migration from mutant explants in the absence of Fibronectin-coating. (D) Table of average values ± standard deviation obtained in yolk sac explant assays. n = number of explants with migrating cells that were quantified. Explant area = area occupied by explant in μm2. (B) Migration area = area occupied by cells migrating from the explant measured 16 hours after plating in μm2.
Immunohistochemistry of sagittal sections from E8.5 wild type (A, E) and chato mutant (B, F) embryos with anti-Fibronectin (green, A–D) and anti-Laminin (green, E–H) antibodies. C–D and G–H are high magnifications of boxed areas in A–B and E–F, respectively. Both wild type and chato mutants contained Fibronectin and Laminin in the basement membranes of the yolk sac (ys bm), the chorion (ch bm) and the amnion (amn bm), as well as in the area surrounding blood islands (bi). Scale bars in A and E represent 100μm.
Acknowledgments
We thank Kathryn Anderson, Joaquim Culi, Juan Modolell, Mark Roberson, John Schimenti, Sol Sotillos, members of the laboratory and anonymous reviewers for helpful discussions and comments on the manuscript; Drs. Ruth Arkell, Kathleen McGrath, Deborah Chapman, James Cross, Kat Hadjantonakis, Janet Rossant and Leif Lundh for mice and reagents; Ke-Yu Deng, personnel at Cornell’s Center for Materials Research and CARE for technical help. The conditional genetrap Zfp568 allele was obtained from the German Gene Trap Consortium (GGTC). This work was supported by Basil O’Connor Starter Scholar Research Award #5-FY06-589 from the March of Dimes Foundation and by NIH grant HD060581.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Argraves WS, Drake CJ. Genes critical to vasculogenesis as defined by systematic analysis of vascular defects in knockout mice. Anat Rec A Discov Mol Cell Evol Biol. 2005;286:875–84. doi: 10.1002/ar.a.20232. [DOI] [PubMed] [Google Scholar]
- Baldwin HS, Shen HM, Yan HC, DeLisser HM, Chung A, Mickanin C, Trask T, Kirschbaum NE, Newman PJ, Albelda SM, et al. Platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31): alternatively spliced, functionally distinct isoforms expressed during mammalian cardiovascular development. Development. 1994;120:2539–53. doi: 10.1242/dev.120.9.2539. [DOI] [PubMed] [Google Scholar]
- Baron MH. Embryonic origins of mammalian hematopoiesis. Exp Hematol. 2003;31:1160–9. doi: 10.1016/j.exphem.2003.08.019. [DOI] [PubMed] [Google Scholar]
- Barron M, McAllister D, Smith SM, Lough J. Expression of retinol binding protein and transthyretin during early embryogenesis. Dev Dyn. 1998;212:413–22. doi: 10.1002/(SICI)1097-0177(199807)212:3<413::AID-AJA9>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- Bartholin L, Melhuish TA, Powers SE, Goddard-Leon S, Treilleux I, Sutherland AE, Wotton D. Maternal Tgif is required for vascularization of the embryonic placenta. Dev Biol. 2008;319:285–97. doi: 10.1016/j.ydbio.2008.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basyuk E, Cross JC, Corbin J, Nakayama H, Hunter P, Nait-Oumesmar B, Lazzarini RA. Murine Gcm1 gene is expressed in a subset of placental trophoblast cells. Dev Dyn. 1999;214:303–11. doi: 10.1002/(SICI)1097-0177(199904)214:4<303::AID-AJA3>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- Becker S, Wang ZJ, Massey H, Arauz A, Labosky P, Hammerschmidt M, St-Jacques B, Bumcrot D, McMahon A, Grabel L. A role for Indian hedgehog in extraembryonic endoderm differentiation in F9 cells and the early mouse embryo. Dev Biol. 1997;187:298–310. doi: 10.1006/dbio.1997.8616. [DOI] [PubMed] [Google Scholar]
- Bohnsack BL, Lai L, Dolle P, Hirschi KK. Signaling hierarchy downstream of retinoic acid that independently regulates vascular remodeling and endothelial cell proliferation. Genes Dev. 2004;18:1345–58. doi: 10.1101/gad.1184904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breier G, Albrecht U, Sterrer S, Risau W. Expression of vascular endothelial growth factor during embryonic angiogenesis and endothelial cell differentiation. Development. 1992;114:521–32. doi: 10.1242/dev.114.2.521. [DOI] [PubMed] [Google Scholar]
- Burdsal CA, Damsky CH, Pedersen RA. The role of E-cadherin and integrins in mesoderm differentiation and migration at the mammalian primitive streak. Development. 1993;118:829–44. doi: 10.1242/dev.118.3.829. [DOI] [PubMed] [Google Scholar]
- Byrd N, Becker S, Maye P, Narasimhaiah R, St-Jacques B, Zhang X, McMahon J, McMahon A, Grabel L. Hedgehog is required for murine yolk sac angiogenesis. Development. 2002;129:361–72. doi: 10.1242/dev.129.2.361. [DOI] [PubMed] [Google Scholar]
- Chapman DL, Garvey N, Hancock S, Alexiou M, Agulnik SI, Gibson-Brown JJ, Cebra-Thomas J, Bollag RJ, Silver LM, Papaioannou VE. Expression of the T-box family genes, Tbx1-Tbx5, during early mouse development. Dev Dyn. 1996;206:379–90. doi: 10.1002/(SICI)1097-0177(199608)206:4<379::AID-AJA4>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- Chawengsaksophak K, de Graaff W, Rossant J, Deschamps J, Beck F. Cdx2 is essential for axial elongation in mouse development. Proc Natl Acad Sci U S A. 2004;101:7641–5. doi: 10.1073/pnas.0401654101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coffinier C, Thepot D, Babinet C, Yaniv M, Barra J. Essential role for the homeoprotein vHNF1/HNF1beta in visceral endoderm differentiation. Development. 1999;126:4785–94. doi: 10.1242/dev.126.21.4785. [DOI] [PubMed] [Google Scholar]
- Cross JC. Genetic insights into trophoblast differentiation and placental morphogenesis. Semin Cell Dev Biol. 2000;11:105–13. doi: 10.1006/scdb.2000.0156. [DOI] [PubMed] [Google Scholar]
- Damert A, Miquerol L, Gertsenstein M, Risau W, Nagy A. Insufficient VEGFA activity in yolk sac endoderm compromises haematopoietic and endothelial differentiation. Development. 2002;129:1881–92. doi: 10.1242/dev.129.8.1881. [DOI] [PubMed] [Google Scholar]
- Dickson MC, Martin JS, Cousins FM, Kulkarni AB, Karlsson S, Akhurst RJ. Defective haematopoiesis and vasculogenesis in transforming growth factor-beta 1 knock out mice. Development. 1995;121:1845–54. doi: 10.1242/dev.121.6.1845. [DOI] [PubMed] [Google Scholar]
- Downs KM, Hellman ER, McHugh J, Barrickman K, Inman KE. Investigation into a role for the primitive streak in development of the murine allantois. Development. 2004;131:37–55. doi: 10.1242/dev.00906. [DOI] [PubMed] [Google Scholar]
- Dyer MA, Farrington SM, Mohn D, Munday JR, Baron MH. Indian hedgehog activates hematopoiesis and vasculogenesis and can respecify prospective neurectodermal cell fate in the mouse embryo. Development. 2001;128:1717–30. doi: 10.1242/dev.128.10.1717. [DOI] [PubMed] [Google Scholar]
- Dziadek MA, Andrews GK. Tissue specificity of alpha-fetoprotein messenger RNA expression during mouse embryogenesis. EMBO J. 1983;2:549–54. doi: 10.1002/j.1460-2075.1983.tb01461.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Hashash AH, Warburton D, Kimber SJ. Genes and signals regulating murine trophoblast cell development. Mech Dev. 2010;127:1–20. doi: 10.1016/j.mod.2009.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrington SM, Belaoussoff M, Baron MH. Winged-helix, Hedgehog and Bmp genes are differentially expressed in distinct cell layers of the murine yolk sac. Mech Dev. 1997;62:197–211. doi: 10.1016/s0925-4773(97)00664-3. [DOI] [PubMed] [Google Scholar]
- Ferkowicz MJ, Yoder MC. Blood island formation: longstanding observations and modern interpretations. Exp Hematol. 2005;33:1041–7. doi: 10.1016/j.exphem.2005.06.006. [DOI] [PubMed] [Google Scholar]
- Firulli AB, McFadden DG, Lin Q, Srivastava D, Olson EN. Heart and extra-embryonic mesodermal defects in mouse embryos lacking the bHLH transcription factor Hand1. Nat Genet. 1998;18:266–70. doi: 10.1038/ng0398-266. [DOI] [PubMed] [Google Scholar]
- Flamme I, Risau W. Induction of vasculogenesis and hematopoiesis in vitro. Development. 1992;116:435–9. doi: 10.1242/dev.116.2.435. [DOI] [PubMed] [Google Scholar]
- Fraser ST, Baron MH. Embryonic fates for extraembryonic lineages: New perspectives. J Cell Biochem. 2009;107:586–91. doi: 10.1002/jcb.22165. [DOI] [PubMed] [Google Scholar]
- Friedrich G, Soriano P. Promoter traps in embryonic stem cells: a genetic screen to identify and mutate developmental genes in mice. Genes Dev. 1991;5:1513–23. doi: 10.1101/gad.5.9.1513. [DOI] [PubMed] [Google Scholar]
- Garcia-Garcia MJ, Anderson KV. Essential role of glycosaminoglycans in Fgf signaling during mouse gastrulation. Cell. 2003;114:727–37. doi: 10.1016/s0092-8674(03)00715-3. [DOI] [PubMed] [Google Scholar]
- Garcia-Garcia MJ, Shibata M, Anderson KV. Chato, a KRAB zinc-finger protein, regulates convergent extension in the mouse embryo. Development. 2008;135:3053–62. doi: 10.1242/dev.022897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George EL, Baldwin HS, Hynes RO. Fibronectins are essential for heart and blood vessel morphogenesis but are dispensable for initial specification of precursor cells. Blood. 1997;90:3073–81. [PubMed] [Google Scholar]
- Gladdy RA, Nutter LM, Kunath T, Danska JS, Guidos CJ. p53-Independent apoptosis disrupts early organogenesis in embryos lacking both ataxia-telangiectasia mutated and Prkdc. Mol Cancer Res. 2006;4:311–8. doi: 10.1158/1541-7786.MCR-05-0258. [DOI] [PubMed] [Google Scholar]
- Glukhova MA, Thiery JP. Fibronectin and integrins in development. Semin Cancer Biol. 1993;4:241–9. [PubMed] [Google Scholar]
- Goh KL, Yang JT, Hynes RO. Mesodermal defects and cranial neural crest apoptosis in alpha5 integrin-null embryos. Development. 1997;124:4309–19. doi: 10.1242/dev.124.21.4309. [DOI] [PubMed] [Google Scholar]
- Goumans MJ, Zwijsen A, van Rooijen MA, Huylebroeck D, Roelen BA, Mummery CL. Transforming growth factor-beta signalling in extraembryonic mesoderm is required for yolk sac vasculogenesis in mice. Development. 1999;126:3473–83. doi: 10.1242/dev.126.16.3473. [DOI] [PubMed] [Google Scholar]
- Guillemot F, Nagy A, Auerbach A, Rossant J, Joyner AL. Essential role of Mash-2 in extraembryonic development. Nature. 1994;371:333–6. doi: 10.1038/371333a0. [DOI] [PubMed] [Google Scholar]
- Gurtner GC, Davis V, Li H, McCoy MJ, Sharpe A, Cybulsky MI. Targeted disruption of the murine VCAM1 gene: essential role of VCAM-1 in chorioallantoic fusion and placentation. Genes Dev. 1995;9:1–14. doi: 10.1101/gad.9.1.1. [DOI] [PubMed] [Google Scholar]
- Hashimoto K, Fujimoto H, Nakatsuji N. An ECM substratum allows mouse mesodermal cells isolated from the primitive streak to exhibit motility similar to that inside the embryo and reveals a deficiency in the T/T mutant cells. Development. 1987;100:587–98. doi: 10.1242/dev.100.4.587. [DOI] [PubMed] [Google Scholar]
- Hayashi S, Lewis P, Pevny L, McMahon AP. Efficient gene modulation in mouse epiblast using a Sox2Cre transgenic mouse strain. Mech Dev. 2002;119(Suppl 1):S97–S101. doi: 10.1016/s0925-4773(03)00099-6. [DOI] [PubMed] [Google Scholar]
- Hu D, Cross JC. Development and function of trophoblast giant cells in the rodent placenta. Int J Dev Biol. 54:341–54. doi: 10.1387/ijdb.082768dh. [DOI] [PubMed] [Google Scholar]
- Hu D, Cross JC. Development and function of trophoblast giant cells in the rodent placenta. Int J Dev Biol. 2010;54:341–54. doi: 10.1387/ijdb.082768dh. [DOI] [PubMed] [Google Scholar]
- Hunter PJ, Swanson BJ, Haendel MA, Lyons GE, Cross JC. Mrj encodes a DnaJ-related co-chaperone that is essential for murine placental development. Development. 1999;126:1247–58. doi: 10.1242/dev.126.6.1247. [DOI] [PubMed] [Google Scholar]
- Inman KE, Downs KM. Brachyury is required for elongation and vasculogenesis in the murine allantois. Development. 2006;133:2947–59. doi: 10.1242/dev.02454. [DOI] [PubMed] [Google Scholar]
- Inman KE, Downs KM. The murine allantois: emerging paradigms in development of the mammalian umbilical cord and its relation to the fetus. Genesis. 2007;45:237–58. doi: 10.1002/dvg.20281. [DOI] [PubMed] [Google Scholar]
- Jollie WP. Development, morphology, and function of the yolk-sac placenta of laboratory rodents. Teratology. 1990;41:361–81. doi: 10.1002/tera.1420410403. [DOI] [PubMed] [Google Scholar]
- Kwee L, Baldwin HS, Shen HM, Stewart CL, Buck C, Buck CA, Labow MA. Defective development of the embryonic and extraembryonic circulatory systems in vascular cell adhesion molecule (VCAM-1) deficient mice. Development. 1995;121:489–503. doi: 10.1242/dev.121.2.489. [DOI] [PubMed] [Google Scholar]
- Larsson J, Goumans MJ, Sjostrand LJ, van Rooijen MA, Ward D, Leveen P, Xu X, ten Dijke P, Mummery CL, Karlsson S. Abnormal angiogenesis but intact hematopoietic potential in TGF-beta type I receptor-deficient mice. Embo J. 2001;20:1663–73. doi: 10.1093/emboj/20.7.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechleider RJ, Ryan JL, Garrett L, Eng C, Deng C, Wynshaw-Boris A, Roberts AB. Targeted mutagenesis of Smad1 reveals an essential role in chorioallantoic fusion. Dev Biol. 2001;240:157–67. doi: 10.1006/dbio.2001.0469. [DOI] [PubMed] [Google Scholar]
- Lescisin KR, Varmuza S, Rossant J. Isolation and characterization of a novel trophoblast-specific cDNA in the mouse. Genes Dev. 1988;2:1639–46. doi: 10.1101/gad.2.12a.1639. [DOI] [PubMed] [Google Scholar]
- Linzer DI, Fisher SJ. The placenta and the prolactin family of hormones: regulation of the physiology of pregnancy. Mol Endocrinol. 1999;13:837–40. doi: 10.1210/mend.13.6.0286. [DOI] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods. 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- Lu CC, Brennan J, Robertson EJ. From fertilization to gastrulation: axis formation in the mouse embryo. Curr Opin Genet Dev. 2001;11:384–92. doi: 10.1016/s0959-437x(00)00208-2. [DOI] [PubMed] [Google Scholar]
- Luo J, Sladek R, Bader JA, Matthyssen A, Rossant J, Giguere V. Placental abnormalities in mouse embryos lacking the orphan nuclear receptor ERR-beta. Nature. 1997;388:778–82. doi: 10.1038/42022. [DOI] [PubMed] [Google Scholar]
- Mahlapuu M, Ormestad M, Enerback S, Carlsson P. The forkhead transcription factor Foxf1 is required for differentiation of extra-embryonic and lateral plate mesoderm. Development. 2001;128:155–66. doi: 10.1242/dev.128.2.155. [DOI] [PubMed] [Google Scholar]
- McGrath KE, Koniski AD, Malik J, Palis J. Circulation is established in a stepwise pattern in the mammalian embryo. Blood. 2003;101:1669–76. doi: 10.1182/blood-2002-08-2531. [DOI] [PubMed] [Google Scholar]
- Meyers EN, Lewandoski M, Martin GR. An Fgf8 mutant allelic series generated by Cre- and Flp-mediated recombination. Nat Genet. 1998;18:136–41. doi: 10.1038/ng0298-136. [DOI] [PubMed] [Google Scholar]
- Morikawa Y, Cserjesi P. Extra-embryonic vasculature development is regulated by the transcription factor HAND1. Development. 2004;131:2195–204. doi: 10.1242/dev.01091. [DOI] [PubMed] [Google Scholar]
- Nagy A. Manipulating the mouse embryo: a laboratory manual. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, N.Y: 2003. [Google Scholar]
- Oshima M, Oshima H, Taketo MM. TGF-beta receptor type II deficiency results in defects of yolk sac hematopoiesis and vasculogenesis. Dev Biol. 1996;179:297–302. doi: 10.1006/dbio.1996.0259. [DOI] [PubMed] [Google Scholar]
- Papadaki C, Alexiou M, Cecena G, Verykokakis M, Bilitou A, Cross JC, Oshima RG, Mavrothalassitis G. Transcriptional repressor erf determines extraembryonic ectoderm differentiation. Mol Cell Biol. 2007;27:5201–13. doi: 10.1128/MCB.02237-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rashbass P, Cooke LA, Herrmann BG, Beddington RS. A cell autonomous function of Brachyury in T/T embryonic stem cell chimaeras. Nature. 1991;353:348–51. doi: 10.1038/353348a0. [DOI] [PubMed] [Google Scholar]
- Rhee JM, Pirity MK, Lackan CS, Long JZ, Kondoh G, Takeda J, Hadjantonakis AK. In vivo imaging and differential localization of lipid-modified GFP-variant fusions in embryonic stem cells and mice. Genesis. 2006;44:202–18. doi: 10.1002/dvg.20203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, Parsons JT, Horwitz AR. Cell migration: integrating signals from front to back. Science. 2003;302:1704–9. doi: 10.1126/science.1092053. [DOI] [PubMed] [Google Scholar]
- Riley P, Anson-Cartwright L, Cross JC. The Hand1 bHLH transcription factor is essential for placentation and cardiac morphogenesis. Nat Genet. 1998;18:271–5. doi: 10.1038/ng0398-271. [DOI] [PubMed] [Google Scholar]
- Rossant J, Cross JC. Placental development: lessons from mouse mutants. Nat Rev Genet. 2001;2:538–48. doi: 10.1038/35080570. [DOI] [PubMed] [Google Scholar]
- Rossant J, Tam PP. Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. Development. 2009;136:701–13. doi: 10.1242/dev.017178. [DOI] [PubMed] [Google Scholar]
- Rossant J, Tam PPL. Mouse development: patterning, morphogenesis, and organogenesis. Academic; San Diego, Calif. London: 2002. [Google Scholar]
- Scott IC, Anson-Cartwright L, Riley P, Reda D, Cross JC. The HAND1 basic helix-loop-helix transcription factor regulates trophoblast differentiation via multiple mechanisms. Mol Cell Biol. 1999;20:530–41. doi: 10.1128/mcb.20.2.530-541.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solloway MJ, Robertson EJ. Early embryonic lethality in Bmp5;Bmp7 double mutant mice suggests functional redundancy within the 60A subgroup. Development. 1999;126:1753–68. doi: 10.1242/dev.126.8.1753. [DOI] [PubMed] [Google Scholar]
- Tallquist MD, Soriano P. Epiblast-restricted Cre expression in MORE mice: a tool to distinguish embryonic vs. extra-embryonic gene function. Genesis. 2000;26:113–5. doi: 10.1002/(sici)1526-968x(200002)26:2<113::aid-gene3>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- Tian Y, Lei L, Cammarano M, Nekrasova T, Minden A. Essential role for the Pak4 protein kinase in extraembryonic tissue development and vessel formation. Mech Dev. 2009;126:710–20. doi: 10.1016/j.mod.2009.05.002. [DOI] [PubMed] [Google Scholar]
- Tremblay KD, Dunn NR, Robertson EJ. Mouse embryos lacking Smad1 signals display defects in extra-embryonic tissues and germ cell formation. Development. 2001;128:3609–21. doi: 10.1242/dev.128.18.3609. [DOI] [PubMed] [Google Scholar]
- Tzu J, Marinkovich MP. Bridging structure with function: structural, regulatory, and developmental role of laminins. Int J Biochem Cell Biol. 2008;40:199–214. doi: 10.1016/j.biocel.2007.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urrutia R. KRAB-containing zinc-finger repressor proteins. Genome Biol. 2003;4:231. doi: 10.1186/gb-2003-4-10-231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uy GD, Downs KM, Gardner RL. Inhibition of trophoblast stem cell potential in chorionic ectoderm coincides with occlusion of the ectoplacental cavity in the mouse. Development. 2002;129:3913–24. doi: 10.1242/dev.129.16.3913. [DOI] [PubMed] [Google Scholar]
- Watson ED, Cross JC. Development of structures and transport functions in the mouse placenta. Physiology (Bethesda) 2005;20:180–93. doi: 10.1152/physiol.00001.2005. [DOI] [PubMed] [Google Scholar]
- Welsh IC, O’Brien TP. Loss of late primitive streak mesoderm and interruption of left-right morphogenesis in the Ednrb(s-1Acrg) mutant mouse. Dev Biol. 2000;225:151–68. doi: 10.1006/dbio.2000.9814. [DOI] [PubMed] [Google Scholar]
- Winnier G, Blessing M, Labosky PA, Hogan BL. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995;9:2105–16. doi: 10.1101/gad.9.17.2105. [DOI] [PubMed] [Google Scholar]
- Xu X, Weinstein M, Li C, Naski M, Cohen RI, Ornitz DM, Leder P, Deng C. Fibroblast growth factor receptor 2 (FGFR2)-mediated reciprocal regulation loop between FGF8 and FGF10 is essential for limb induction. Development. 1998;125:753–65. doi: 10.1242/dev.125.4.753. [DOI] [PubMed] [Google Scholar]
- Yamaguchi TP, Dumont DJ, Conlon RA, Breitman ML, Rossant J. flk-1, an flt-related receptor tyrosine kinase is an early marker for endothelial cell precursors. Development. 1993;118:489–98. doi: 10.1242/dev.118.2.489. [DOI] [PubMed] [Google Scholar]
- Yang JT, Rayburn H, Hynes RO. Cell adhesion events mediated by alpha 4 integrins are essential in placental and cardiac development. Development. 1995;121:549–60. doi: 10.1242/dev.121.2.549. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A–F) In situ hybridizations in E8.5 wild type (A, C, E) and chato mutants (B, D, F) with Ttr (A,B) Hnf1b (C,D) and ®H1 globin (E,F) probes. High magnifications of sagittal sections through the yolk sac region are shown. (B) class III and (D, F) class II chato mutants (G–J) Wild type (G) and chato mutant sagittal sections were stained with anti-CDH1 antibody (green) and counterstained with DAPI (blue). (I,J) higher magnifications of boxed areas in G, H respectively. (H, J) show a class II chato mutant. (K–L) Immunohistochemistry with anti-phospho-histone H3 antibodies (PH3, red) counterstained with Phalloidin (green) as shown on a class III chato mutant (M) Quantification of the number of PH3 positive cells per section in the VE of wild type (orange; 7.97±3.18) and chato (blue; 6.48±2.38) embryos (p=0.0655). Error bars indicate s.d.; n=embryos and s=sections quantified. VE, visceral endoderm; amn, amnion; TE, trophectoderm; Exm, extraembryonic mesoderm; ch, chorion. Scale bars represent 25μm.
(A–D) TUNEL (green) and DAPI (blue) staining on sagittal sections of wild type (A) and a class II chato (B) embryo. (C,D) Higher magnifications of boxed regions in A, B. Arrowheads point to TUNEL positive nuclei. (E,F) Quantification of total TUNEL positive cells per section in allantois and yolk sac (E) and chorionic ectoderm (F). Error bars indicate s.d.; n=embryos and s=sections quantified. ys=yolk sac, ch=chorion. Scale bar represents 100μm.
Sagittal sections of E8.5 wild type (A,G) and class II chato (B,H) embryos labeled with anti-VCAM (A,B; green) or anti-α4 integrin (G,H; green) antibodies and DAPI (red) counterstaining. C–F,I–L, are magnifications of boxed regions in A,B,G,H, respectively (E–F,K–L; green channel only). (M–P) Sagittal sections of wild type (M, O) and class II chato mutant embryos (N, P) stained with anti-phospho-histone H3 antibodies (PH3, red) and Phalloidin (green) (M–N) or with TUNEL (green) and DAPI (blue) (O–P). all, allantois; ch, chorion; VE, visceral endoderm. Arrowheads in O,P point to TUNEL positive nuclei in allantois. Scale bars in A,G,M,O represent 100μm.
(A–D) Raw images of the sagittal sections of E8.5 wild type (A) and chato (B–D) embryos shown in Figure 3. (E–G) Sagittal sections of X-gal stained tetraploid chimeras obtained from aggregation of wild type ES cells (control tetraploidwt ↔ ES cellwt chimera) (E) and chato mutant ES cells (tetraploidwt ↔ ES cellchato chimeras) (F,G). We found that chato tetraploid chimeras (n=8) reproduced the characteristic defects of class II (F, 75%) and class III chato mutants (G, 25%). The Rosa26 allele in the ES cell lines used to generate these chimeras served as a lineage marker to differentiate embryonic-derived cells (embryo and extraembryonic mesoderm, Xgal positive) from VE and trophoblast tissues (Xgal negative). (H, I) Whole mount in situ hybridizations with a Fgfr2 probe in wild type (H) and chato embryos (I). amn, amnion; ch, chorion; exc, exocoelomic cavity; ec, ectoplacental cavity. Red arrowheads point to chorion. Double-arrowed line in D points to the enlarged ectoplacental cavity. Brackets in C and D indicate yolk sac bubbles concentrated at the embryonic-extraembryonic boundary. Scale bars in A,E,H represent 100μm.
Whole mount in situ hybridizations of E8.5 wild type (A,E,I, M) and chato mutant (B, F, J, N, P) embryos with Pl1 (A–B, O–Q), Tpbpa (E–F), and Ascl2 (I–J) probes. (C–D, G–H, K–L) Sagittal sections of the trophoblast region of embryos in A–B, E–F, I–J, respectively. The trophoblast populations marked by these probes are indicated in the bottom margin. (M–N) High magnifications of E8.5 wild type (I) and chato mutant (J) sagittal sections showing Gcm1 expression in clusters of chorionic cells (arrowheads). In chato embryos, Gcm1 clusters are smaller. (O–Q) Whole mount in situ hybridizations of E8.5 wild type (O), chato (P) and Hand1 mutant (Q) embryos with a Pl1 probe. Hand1 mutants have a severe reduction in the number of Pl1-expressing trophoblast giant cells (Riley et al., 1998). In contrast, the number of Pl1-expressing trophoblast giant cells in Zfp568null embryos is similar to that of wild type littermates. Brackets in F and J mark yolk sac ruffles. Scale bars represent 100μm.
Sections of whole mount in situ hybridizations of wild type (A–C) and Zfp568null (D) embryos at E8.0 (A) and E8.5 (B,C,D) with a Zfp568 probe. Zfp568 expression was not detected in control Zfp568null embryos (D). bc, blood cells; exc, exocoelomic cavity; TE, trophectoderm; VE, visceral endoderm; Exm, extraembryonic mesoderm. Scale bars represents 100μm.
(A) The Zfp568 reversible allele (Zfp568rGT-mutant) was generated from a German Gene Trap Consortium ES cell clone containing a FlipROSAbgeo genetrap vector inserted into the third intron of the Zfp568 locus. The genetrap cassette in Zfp568rGT-mutant disrupted the Zfp568 transcript and generated a truncated ZFP568 protein fused to βgeo (http://tikus.gsf.de/ggtc/). This truncated protein, which originated from splicing between the second exon of Zfp568 and the splicing acceptor site in the genetrap cassette, contained only 11 N-terminal amino acids of ZFP568 and lacked all ZFP568 functional domains (KRAB domains and Zinc finger motifs). Complementation tests between Zfp568rGT-mutant, Zfp568chato and Zfp568null, as well as RT-PCR experiments, all confirmed that Zfp568rGT-mutant genetrap insertion generates a null allele for Zfp568. The reversible capabilities of Zfp568rGT-mutant reside in the loxP and FRT recombination sites (red and green circles) flanking the FlipROSAβgeo genetrap cassette. These sites mediate the inversion of the genetrap cassette when Cre or FLP recombinases are present and release the effect of the genetrap cassette on Zfp568, therefore restoring wild type function to the Zfp568 locus. (B–D) Complementation tests between Zfp568rGT-restored and Zfp568null alleles revealed that 7 out of 13 Zfp568rGT-restored/null embryos had a smooth yolk sac and wild type embryo morphology at E9.5 (C), whereas 6/13 embryos (D) had all the characteristic defects of chato mutants, including yolk sac ruffling. All other littermates were wild type in appearance (B). These results suggested that the inversion of the cassette did not fully restore wild type ZFP568 function and that Zfp568rGT-restored was likely a hypomorphic allele of Zfp568. Because the targeted insertion of vectors in intronic sequences has been previously reported to affect gene transcription (Meyers et al., 1998), we hypothesized that the presence of the genetrap allele in Zfp568rGT-restored, even in the wild type configuration, interferes with the normal splicing of the Zfp568 transcript. (E) Quantification of the levels of splicing through the two exons flanking the genetrap insertion site using qRT-PCR on E9.5 embryonic tissues showed that Zfp568rGT-restored/+ embryos have a 40% reduction in the levels of full length Zfp568 transcript. Zfp568rGT-restored/null embryos only express 10% the level of full length transcript compared to wild type embryos. These results confirmed Zfp568rGT-restored is a hypomorphic allele of Zfp568. ***p=0.0007 compared to wt. Error bars represent s.d. SA, splice acceptor; pA, polyadenylation signal.
(A–C) Graphic representation of results from yolk sac explant assays plated onto Lab-Tek™ II (Nunc) slides coated with Fibronectin (FN), no coating, Gelatin, or Laminin (LAM). The regions containing migrating cells for each wt (A, grey) and Zfp568 mutant (B, pink) explant were represented as shaded areas on different layers at 30% transparency, then scaled so that the size of the initial explant size (central white area) was constant across all explants. Darker areas represent regions most frequently populated by migrating cells. Overlays of wt and Zfp568 mutant explants (C) highlight the reduced migration from mutant explants in the absence of Fibronectin-coating. (D) Table of average values ± standard deviation obtained in yolk sac explant assays. n = number of explants with migrating cells that were quantified. Explant area = area occupied by explant in μm2. (B) Migration area = area occupied by cells migrating from the explant measured 16 hours after plating in μm2.
Immunohistochemistry of sagittal sections from E8.5 wild type (A, E) and chato mutant (B, F) embryos with anti-Fibronectin (green, A–D) and anti-Laminin (green, E–H) antibodies. C–D and G–H are high magnifications of boxed areas in A–B and E–F, respectively. Both wild type and chato mutants contained Fibronectin and Laminin in the basement membranes of the yolk sac (ys bm), the chorion (ch bm) and the amnion (amn bm), as well as in the area surrounding blood islands (bi). Scale bars in A and E represent 100μm.