

Abstract
The essential biological role of erythropoietin (EPO) in maintaining erythrocyte mass has been well understood for many years. Although EPO is required for the maturation of red cells, it also has strong procoagulant effects on the vascular endothelium and platelets, which limit erythrocyte losses after hemorrhage. Like other members of the type 1 cytokine superfamily, EPO has multiple biological activities. For the past 10 years, multiple investigators have shown that EPO acts as a locally produced antagonist of proinflammatory cytokines that are generated by the innate immune response in response to infection, trauma, or metabolic stress. Specifically, EPO inhibits apoptosis of cells surrounding a locus of injury, reduces the influx of inflammatory cells, and recruits tissue-specific stem cells and endothelial progenitor cells. Available evidence suggests that these multiple, nonerythropoietic effects of EPO are mediated by a tissue protective receptor (TPR) that is distinct from the homodimeric receptor responsible for erythropoiesis. Notably, activation of the TPR requires a higher concentration of EPO than is needed for maximal erythropoiesis. Unfortunately, these higher concentrations of EPO also stimulate hematopoietic and pro-coagulant pathways, which can cause adverse effects and, therefore, potentially limit the clinical use of EPO for tissue protection. To circumvent these problems, the EPO molecule has been successfully modified in a variety of ways to interact only with the TPR. Early clinical experience has shown that these compounds appear to be safe, and proof of concept trials are ready to begin.
Keywords: inflammation, cytoprotection, apoptosis, cytokines, innate immune response
OVERVIEW: TISSUE PROTECTIVE EFFECTS OF ERYTHROPOIETIN
Erythropoietin (EPO) was first identified as a hormone that stimulates erythropoiesis by blocking the programmed cell death of red cell precursors (reviewed by Fisher1). In its endocrine role, EPO is produced in the adult kidney in response to hypoxia and released into the circulation. After binding to the homodimeric EPO receptor (EPOR)2 at picomolar concentrations, it facilitates the survival of proerythroblasts within the bone marrow (Fig. 1). Because erythrocyte precursors are continuously produced, a sustained, low concentration of EPO within the circulation is required to replace senescent red cells.
FIGURE 1.

Erythropoietin signals via 2 distinct receptor isoforms. The hematopoietic receptor is a high affinity homodimer that mediates hematological and vascular effects. In contrast, current evidence suggests that the TPR is a lower affinity heteromer composed of EPOR and βCR (CD131).
Unexpectedly, in 1990, investigators observed that EPO also stimulated growth and migration of endothelial cells in culture.2 Notably, the concentrations of EPO required for these effects were substantially higher (nanomolar) than that required for erythropoiesis. Follow-on studies showed neuronal-like PC12 cells also expressed a receptor for EPO with a lower affinity for EPO3 that was biochemically distinct from the (EPOR)2. Avariety of recent evidence has shown that this alternative receptor for EPO is a heteromer, composed of EPOR monomer subunits and the beta common receptor (CD131) that is also used for signaling by the type I cytokines granulocyte macrophage colony-stimulating factor, interleukin 3 (IL-3), and IL-5.4 Recently, Sautina et al.5 have proposed that the VEGF receptor also associates with CD131 in vascular endothelial cells.
The existence of receptor isoforms suggested that the EPO has other biological functions in addition to erythropoiesis. Indeed, for the past decade, additional work has shown that EPO is a principal component of a paracrine-autocrine system that mediates tissue protection. In this mode, EPO is locally produced in response to tissue injury but with a significant time delay. This pool of EPO subsequently signals through the heteromeric receptor (tissue protective receptor; TPR) to prevent secondary injury by suppressing proinflammatory cytokines activated by the innate immune response and blocks cellular apoptosis. In a variety of tissues,6,7 EPO antagonizes both the production and the effects of proinflammatory cytokines and limits ischemia-reperfusion injury. Expression of the TPR peaks several hours after infection or injury but before EPO production. In addition, proinflammatory cytokines suppress endogenous EPO but stimulate TPR expression. It is the relative excess of the TPR with respect to EPO that provides the rationale for administration of exogenous EPO for tissue protection.
Historically, the cytoprotective effects of EPO were first demonstrated in the nervous system, which remains the most extensively studied tissue to date. Our group expanded relevance of the newly discovered neuroprotective effects of EPO into ischemic, traumatic, and inflammatory lesions in a series of publications8 beginning in 2000 and as reviewed by Brines and Cerami.6 Furthermore, we showed that systemically administered EPO could cross the blood-brain barrier (BBB) in therapeutically relevant concentrations and significantly reduce the volume of injury in animal models of stroke or traumatic brain injury (TBI). The discovery that EPO passes through the intact BBB and is active not only in ischemia but in a variety of injury models greatly accelerated interest in using recombinant human EPO (rhEPO) to prevent brain injury. Since then, we and many other investigators have shown that EPO possesses tissue protective effects in diverse tissues and organs, including the heart, kidney, gut, lung, and skin (reviewed by Arcasoy9). Here, we will highlight in moderate detail some of the significant findings in the nervous system as examples of EPO-mediated tissue protection.
ERYTHROPOIETIN IN CENTRAL AND PERIPHERAL NERVOUS SYSTEM INJURIES
Stroke
Many laboratories have assessed the effects of systemically administered rhEPO in animal models of nonhemorrhagic ischemic stroke. Recently, 2 meta-analyses of these animal studies were published. Minnerup et al.10 analyzed 16 preclinical studies and concluded that when rhEPOs (or its analogues, as discussed later) were administered after the onset of ischemia, infarct size was reduced by about one-third and was associated with improved neurobehavioral outcomes. A second meta-analysis11 examined 19 studies involving 346 animals for infarct size and 425 animals for neurobehavioral outcome and found that rhEPO improved infarct size by 30.0% (95% confidence interval [CI], 21.3–38.8) and neurobehavioral outcome by 39.8% (33.7–45.9). These authors also note that studies that randomized the animals to treatment group or that blinded assessment of outcome showed lower efficacy.11
Results of the first attempt to translate these preclinical findings into a patient population suffering from nonhemorrhagic stroke in the territory of the middle cerebral artery suggested that rhEPO might be beneficial for the treatment of patients.12 However, a larger double-blind, placebo-controlled, randomized multicenter EPO Stroke Trial (NCT00604630), designed to evaluate efficacy and safety of rhEPO in stroke, did not demonstrate a therapeutic effect. In fact, the overall death rate was increased in the rhEPO-treated group: 16.4% (n = 42 of 256) versus 9.0% (n = 24 of 266) in the placebo group (odds ratio, 1.98; 95% CI, 1.16–3.38; P = 0.01).13 The major difference between these 2 studies was that the later one included a significant number of patients that also received thrombolytic treatment. Results of a recent preclinical rodent stroke model provide a plausible explanation for the different outcome of these trials: Zechariah et al.14 have shown that tissue plasminogen activator amplifies EPO-mediated metalloproteinase production, leading to a breakdown of the BBB and increased tissue damage.
Traumatic Brain and Spinal Cord Injury
In experimental models, rhEPO improves outcome after a TBI.8,15,16 At high doses, rhEPO has been shown to have neuroprotective effects when given early after injury and to have effects that enhance neurological recovery even when given with a significant temporal delay.17–23 The time window for EPO-induced neuroprotection has not been well defined, but EPO given as long as 24 hours post-injury has been reported to have significantly enhanced neurological recovery in TBI models. A clinical trial evaluating the effects of rhEPO in patients with TBI (NCT00313716) is ongoing at the Baylor College of Medicine and the University of Texas Health Science Center at Houston/Memorial Hermann Hospital. An additional multicenter trial is planned, but not yet initiated, by the Australian and New Zealand Intensive Care Research Centre (NCT00987454). Multiple laboratories have also demonstrated that rhEPO is neuroprotective after spinal cord injury in animal models.18–23 There is one active trial (NCT00561067) evaluating the efficacy of rhEPO in patients with traumatic spinal cord injury.
Cerebral Malaria
Cerebral malaria (CM) is a devastating neurological syndrome arising from an extreme production of proinflammatory cytokines in response to parasite-infected erythrocytes within the brain vasculature. Two laboratories have demonstrated the protective effect of rhEPO in murine models of CM.24,25 The importance of endogenous EPO in human CM is underscored by a recent clinical study of 124 Kenyan children with CM, in which high endogenous plasma levels of EPO correlated with an 80% reduction in risk of developing neurological sequelae.26 A clinical trial evaluating the use of rhEPO as an adjunct treatment for children with CM is currently active. This study (NCT00697164) aims to enroll 200 children of 6 months to 14 years of age admitted to the hospital with CM but without severe anemia or pulmonary involvement. The children will be treated with standard antimalarial treatment (quinine) and a 3-day course of high-dose rhEPO. The main endpoint of this trial is survival at 5 days after admission, and the study is powered to detect a 25% reduction of mortality.
Hypoxic-Ischemic Encephalopathy
Recombinant human EPO confers neuronal protection when administered systemically to animals subjected to global brain ischemia. A trial of rhEPO treatment for 153 term infants with moderate hypoxic-ischemic encephalopathy observed a reduced disability.27 Death or moderate/severe disability occurred in 35 (43.8%) of 80 infants in the control group and 18 (24.6%) of 73 infants in the EPO group (P = 0.017) at 18 months. In addition, a retrospective study of 82 preterm infants given 6 weeks of rhEPO therapy to prevent anemia identified an association between rhEPO doses and higher scores on the Bayley Scale of Infant Development, Psychomotor Developmental, and Mental Developmental Index at 12 months of age.28
Many additional in vivo and in vitro models of nervous system injury have been evaluated including diabetic or toxic neuropathies, experimental autoimmune encephalitis,29–31 and subarachnoid hemorrhage32,33 as recently reviewed by Brines and Cerami.6 A small number of pilot clinical studies have also been performed in normal volunteers and in patients with depression that show effects of EPO on cognition and mood34,35 or in patients with schizophrenia,36 amyotrophic lateral sclerosis,37 or multiple sclerosis.38
MANY TISSUES EXPRESS EPO IN RESPONSE TO INJURY
The role of EPO as a modulator of the innate immune response is not limited to the nervous system. Virtually, all tissues examined to date respond to metabolic stress or injury by activating a local EPO response.
Myocardial Injury
The heart was the first tissue outside of the brain for which cytoprotective effects of EPO were confirmed.39 Subsequent work performed by numerous investigators showed that similar to the effects within the nervous system, rhEPO protects the myocardium from ischemic, infectious, traumatic, or toxic injuries.40–45 Endogenous EPO is produced locally by myocardial tissue, and as expected of a paracrine/autocrine system, a receptor for EPO is expressed by neonatal and adult rat cardiomyocytes,46,47 ventricular myocytes, and endothelial cells.48 Preclinical studies have shown that administration of rhEPO after ischemia/reperfusion decreases infarct size in part by inhibiting cardiomyocyte apotosis. Recombinant human EPO and endogenous EPO may also reduce subacute ischemia by inducing neovascularization, either by stimulating endothelial cells in situ or by mobilizing endothelial progenitor cells from the bone marrow.
Clinical studies have demonstrated a beneficial effect of rhEPO administration in patients with chronic heart failure. Unlike patient populations with chronic kidney disease and cancer-related anemia (discussed later), treatment with erythropoiesis-stimulating agents (ESAs) has not been associated with higher mortality rates in patients with chronic heart failure. In fact, a recent meta-analysis of 7 randomized controlled clinical trials with 650 patients (363 and 287 patients treated with ESAs and placebo, respectively) showed a decreased risk of hospitalization for heart failure in the ESA-treated group and no difference in mortality between the 2 groups.49 Although one clinical study has demonstrated that rhEPO improved cardiac function without increasing hematocrit,50 in most, it has been impossible to determine if these beneficial effects on the human cardiovascular system occur independently of increases in hemoglobin levels.
In addition, in contrast to trials in which ESAs were administered chronically, acute dosing has been evaluated in the setting of revascularization procedures after myocardial infarction. Several small studies have been published that present opposite results. Binbrek et al.51 observed no protective effect of EPO when used in the setting of thrombolysis. In contrast, when administered after stent placement, EPO was observed to confer benefit by reducing infarct size.52
It is important to appreciate that a number of observations derived from animal studies suggest that EPO’s cardioprotective properties are independent of its erythropoietic activities. First, a single dose of rhEPO that does not increase the hematocrit is protective in rodent39 and rabbit53 models of cardiac infarction. Second, isolated cardiomyocytes and isolated perfused hearts are more tolerant to hypoxia when treated with rhEPO.39 Third, EPO modified to possess no erythropoietic activity (discussed later) retains its cardioprotective properties in permanent ischemia and ischemia/reperfusion.54 Fourth, EPO is not protective for cardiomyocytes subjected to metabolic stress that have been obtained from βCR (CD131) knockout mice that possess a fully intact (EPOR)2.4
HOW DOES EPO PROTECT TISSUES?
Erythropoietin’s mechanisms of protective action have been most extensively evaluated in nervous tissue for which rhEPO has been shown to possess neurotrophic,55 antiapoptotic, and angiogenic properties56 and to promote neural regeneration.57 These effects are dose dependent58 and, in some cases, depend in part on other known molecules, for example, brain-derived nerve growth factor.59 In animal models, the expression of EPO and EPOR in the brain after cerebral ischemia/infarction occurs with a strict temporal sequence. Histopathological changes identify a pneumbra around the lesion, that is, tissue at risk for further injury. Within the penumbra, endothelial cells, intravascular inflammatory cells, and neurons all express EPO, whereas EPOR is expressed by neurons, astrocytes, and endothelial cells.57,60,61 Because the expression of EPOR precedes the up-regulation of EPO synthesis,60 tissue sensitivity to EPO increases as a function of time.62 Also, EPO can increase cerebral blood flow by its effect on endothelial nitric oxide production63 and by preventing endothelial cell apoptosis and, in these ways, maintain the microcirculation within the penumbra. In the subacute phase, EPO can also improve tissue oxygenation by driving neoangiogenesis. These all tend to reduce infarct volume significantly.57,60,61 The persistent up-regulation of EPO production by astrocytes after hypoxia also provides rapidly available sources of EPO to enable neurons to tolerate otherwise damaging ischemia.61 Erythropoietin also strongly prevents the development of tissue edema by modulating the water transporter aquaporin in astrocytes.64
Receptor Signaling Cascade
Erythropoiesis occurs when EPO binds to the preformed (EPOR)2 and causes a conformational change that subsequently triggers Janus kinase/signal transducers and activators of transcription (STAT) activation. The resulting molecular cascade results in the up-regulation of antiapoptotic proteins (eg, Bcl-2) that inhibit apoptosis of erythrocyte precursors.65 In contrast, EPO-mediated tissue protection uses multiple signaling pathways activated in parallel and that vary by cell and tissue type. Cell signaling pathways that are known to be used by EPO and mediate tissue protection include (1) phosphorylation and activation of Janus kinase1/2, STAT3, or STAT5A (but not of STAT1alpha and STAT5B); (2) phosphorylation and activation of phosphoinositol 3-kinase and its downstream kinase Akt; (3) activation of protein kinase C, which results in a rise in free intra-cellular calcium; (4) RAS/mitogen-activated protein kinase (Raf, MEK1/2, p42/44 MAPK, and p38 MAPK); and (5) up-regulation of Sonic hedgehog and its downstream target mammalian achaetescute homolog 1 (Mash1).66
Individual specific tissue protective effects that are activated by EPO in particular cell types can depend upon different pathways. For example, EPO has both neurotrophic and antiapoptotic effects in neurons. For hippocampal neurons, EPO signals via both the PI3K/Akt and the STAT pathways to support neurite outgrowth, whereas the STAT pathway is not required for inhibition of apoptosis.67 In contrast, in the SH-SY5Y differentiated neuroblastoma cell line, both the Akt and STAT pathway activation is required to prevent apoptosis.68
A CONUNDRUM: THE HIGH DOSES OF RHEPO REQUIRED FOR TISSUE PROTECTION CAN CAUSE SERIOUS ADVERSE EFFECTS
Despite the many attractive features of rhEPO-mediated tissue protection observed in preclinical models, translation of rhEPO-dependent protection into the clinical setting for use in neurological, renal, or cardiac disorders has posed a dilemma. The high doses of rhEPO required for tissue protection and enhancement of recovery are associated with very real risks of significant clinical complications. In fact, even moderate doses of rhEPO administered chronically have been associated with adverse consequences in patients. For example, the very large Trial to Reduce Cardiovascular Events with Aranesp Therapy trial69 evaluated whether the use of an ESA to treat anemia in diabetic patients with renal insufficiency could reduce mortality and slow disease progression. In fact, the results showed an increased incidence of stroke within the ESA arm and no beneficial effects of rhEPO on disease progression.
Survival
Large-scale clinical studies involving higher doses of ESAs administered as treatment of anemia or to improve quality of life have shown that as a class ESAs reduce overall survival and/or increase the risk of tumor progression or recurrence in patients with breast, non-small cell lung, head and neck, lymphoid, and cervical cancer (http://www.fda.gov/Drugs/DrugSafety/PostmarketDrugSafetyInformationforPatientsandProviders/ucm200297.htm). A recent meta-analysis of randomized trials evaluating the association of recombinant ESAs with mortality in patients with cancer analyzed data from a total of 13,933 patients with cancer in 53 trials.70 Erythropoiesis-stimulating agents increased mortality during the active study period (combined hazard ratio [cHR], 1.17; 95% CI, 1.06–1.30) and reduced overall survival (cHR, 1.06; 95% CI, 1.00–1.12), with little heterogeneity between trials. The cHR for mortality during the active study period was 1.10 (range, 0.98–1.24), whereas it was 1.04 (range, 0.97–1.11) for overall survival. There was little evidence for a difference between trials of patients given different anticancer treatments (P value for interaction = 0.42). The mechanism by which ESAs affect these outcomes is not entirely clear but appears to arise from thrombosis and tumor growth.
Thrombosis
Multiple studies using both animal models and patients indicate that ESAs augment thrombopoeisis as well as platelet and endothelial cell activation.71–77 The resulting thrombotic complications of EPO are dose related.78–80 Almost all studies of ESAs administered to critically ill patients and in postoperative patients have shown an increased risk of thromboembolic events after ESA administration. For example, a randomized, double-blind, placebo-controlled, multicenter trial (N = 1460) in anemic critically ill patients demonstrated a 29-day survival benefit in the trauma subgroup receiving rhEPO (mortality, 6.7% vs 3.5%), although it did not result in fewer blood transfusions. Despite the improvement in the outcome of the rhEPO-treated patients, there was a significant increase in the incidence of thrombo-phlebitis, from 5.7% to 8.7% (P = 0.04).81 The most serious complications associated with rhEPO occurred in a recent trial of rhEPO, given at 40,000 IU/d for 3 days after stroke, in which rhEPO-treated patients had a higher rate of intracranial hemorrhage and a higher mortality rate.13 These results triggered the US Food Drug Administration to place all rhEPO neuroprotection trials in the United States on hold.
Tumor Growth
A second possible mechanism by which exogenously administered rhEPO may worsen outcome in patients with cancer is by promoting vasculogenesis and tumor growth. Erythropoietin has been shown to promote physiological angiogenesis,82,83 angiogenesis during wound healing,84 and estrogen-dependant cyclical angiogenesis in the uterus.85,86 However, it appears that EPO can also drive pathological neoangiogenesis. For example, 3 important studies in different mouse models of cancer have shown that EPO is an important angiogenic factor that regulates neovascularization of tumors.86–88 An additional concern is that EPO may act as a growth factor for certain types of cancer cells, although it must be noted that there is a great deal of controversy concerning this possibility. On the one hand, results of a number of in vitro studies show that rhEPO can induce proliferation, increase cellular migration and invasion, as well as confer resistance to chemoradiation in some tumor cells.89–95 On the other hand, studies using different tumor types have also concluded that rhEPO does not affect cellular proliferation or in vitro sensitivity to chemotherapeutic agents.95–97 It is notable that many workers in the field have demonstrated the expression of EPOR on tumor cells and responses to EPO (reviewed by Arcasoy9). In rebuttal, several papers recently published by scientists employed by Amgen (the largest manufacturer of ESAs) have concluded that the reagents used by other scientists are faulty98 and that tumor cells do not express a functional EPOR or respond biologically to rhEPO.99,100 Further work will obviously be required to clarify this controversial area.
A SOLUTION TO THESE PROBLEMS: DESIGNER EPOS THAT ARE ONLY TISSUE PROTECTIVE?
If the erythropoietic effects of EPO could be functionally separated from its tissue protective effects, more specific ligands could potentially be developed that avoid adverse effects that depend upon the EPOR homodimer. In 2003–2004, our group demonstrated for the first time that the hematopoietic and tissue protective effects of EPO are, in fact, independent biological properties101,102 (Table 1). This was accomplished using 2 approaches: (1) by capitalizing on the very different temporal characteristics of EPO signaling in erythropoiesis compared with tissue protection and (2) by preferentially interfering with the binding of EPO to the hematopoietic receptor.
TABLE 1.
Hematopoeitic and Neuroprotective Activities of rhEPO and Its Derivatives
| rhEPO Derivative | Hematopoeitic Activities (pM) |
Neuroprotective Activities (±SD for Compounds at 300 pM) |
||
|---|---|---|---|---|
| Erythropoietin Receptor Binding* (IC50) | UT7 Proliferation† (EC50) | % Neurons Protected‡ | % P19 Cells Protected§ | |
| Wild-type rhEPO | 10 | 10–30 | 78 ± 13 | 49 ± 12 |
| Asialo-rhEPO¶ | 14 | 10–30 | 71 ± 15 | n.d. |
| Carbamoylated rhEPO (CEPO)|| | > 10,000 | > 10,000 | 70 ± 9 | 49 ± 10 |
| Asialo-CEPO | > 10,000 | > 10,000 | 69 ± 16 | not done |
| S100E-rhEPO** | 100 | > 10,000 | 66 ± 9 | 55 ± 15 |
| R103E-rhEPO†† | > 10,000 | > 10,000 | 55 ± 13 | not done |
Inhibition of rhEPO binding to (EPOR)2.
UT7 is an EPO-sensitive leukemic cell line.
Percent rat hippocampal neurons protected after induction of apoptosis with the neurotoxin N-methyl-D-aspartate.
Percent P19 (neuronal precursor cells derived from murine teratocarcinoma cells) protected after induction of apoptosis by serum withdrawal.
Asialo-rhEPO was generated by total enzymatic desialylation of rhEPO, possesses a very short plasma half-life and is fully neuroprotective in vivo.
Carbamoylated EPO is a chemically modified rhEPO in which all of the lysines were transformed to homocitrulline by carbamoylation.
As discussed earlier, effective erythropoiesis requires EPO to be constantly available. In contrast, tissue protection is characterized by the appearance of molecules that trigger long-lasting effects, acting in effect as molecular switches for long-lasting gene expression programs. Recombinant human EPO possesses a relatively long plasma half-life (~5–8 hours) because of the presence of multiple oligosaccharides terminated by sialic acid. Removal of these sialic acid residues drastically increases plasma clearance, such that asialo-EPO has a reduced plasma half-life of only ~2 minutes. Although a single injection of asialo-EPO has no effect on hematopoiesis, it is as potent as native EPO for tissue protection.101
Another approach to the development of specific tissue protective ligands involves considerations of EPO structure-activity relationships with respect to the hematopoietic receptor. The molecular interaction of EPO with the (EPOR)2 complex has been extensively studied, and critical regions within EPO that interact (Fig. 2). Erythropoietin with (EPOR)2 have been identified103–106 binds to both subunits of the homodimer and, while in a bridging position, induces a conformational change that triggers receptor signaling. The specific regions of EPO involved are portions of helices A and C (site 2, Fig. 2) as well as helix D and the loop connecting helices A and B (site 1, Fig. 2).103–106 Chemical or mutational modifications of amino acid residues within these 2 regions of the EPO molecule prevent its binding to (EPOR)2. Remarkably, however, a number of these modified EPO molecules retain potent tissue protective properties.102 Using this approach, a number of EPO derivatives can be synthesized that are cytoprotective but are devoid of hematopoietic, procoagulant, and vasoconstrictive activities. Carbamoylated EPO (CEPO) is one extensively studied derivative that is produced by chemically converting the positively charged lysines within the molecule (EPO has 8) into the neutral amino acid homocitrulline. Erythropoietin and CEPO differ markedly in a number of biological properties. For example, although both recruit endothelial progenitor cells, only EPO stimulates the mitosis of more differentiated endothelial cells.107 Carbamoylated EPO has been extensively evaluated in multiple preclinical models in our laboratory54,102,107–110 as well as by others66,111–115 and confirmed to possess fully preserved tissue protective properties. For example, rodent cardiomyocytes undergo apoptosis when exposed to the toxin staurosporine in vitro. Inclusion of either CEPO or EPO in the exposure medium significantly attenuates this apoptosis (Fig. 3). Carbamoylated EPO is currently in clinical development for ischemic stroke (NCT00870844) and Friedreich’s ataxia (NCT01016366).
FIGURE 2.
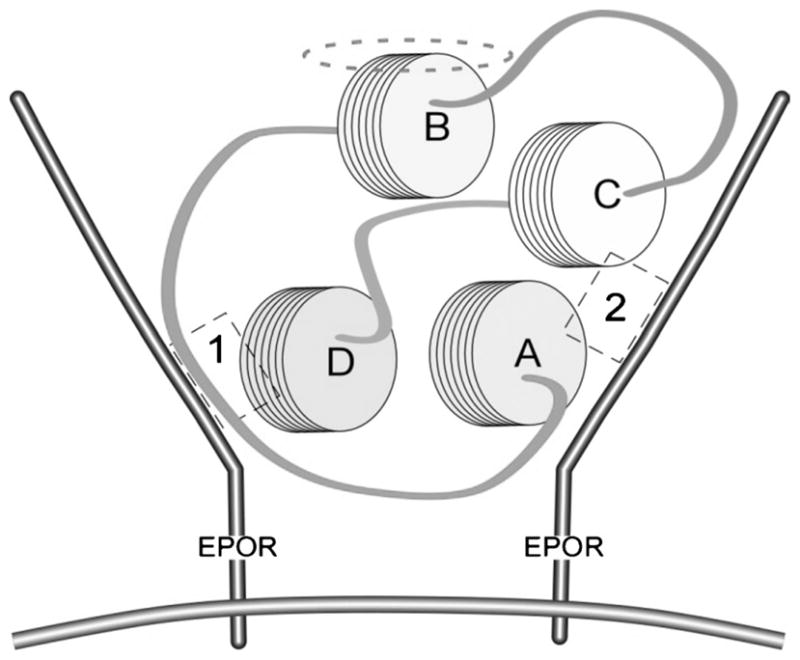
Erythropoietin binds to (EPOR)2 by bridging across an assembled homodimer. Helices A–D of EPO associate via hydrophobic interactions to form a compact, globular configuration. Sites 1 and 2 (indicated by dashed boxes) within the topography of the EPO molecule bind with high affinity to localized regions on each EPOR monomer. The aqueous face of the helix B is oriented away from the interior of the receptor, indicated by the dashed ellipse. Reprinted with permission from Proc Natl Acad Sci U S A.117
FIGURE 3.
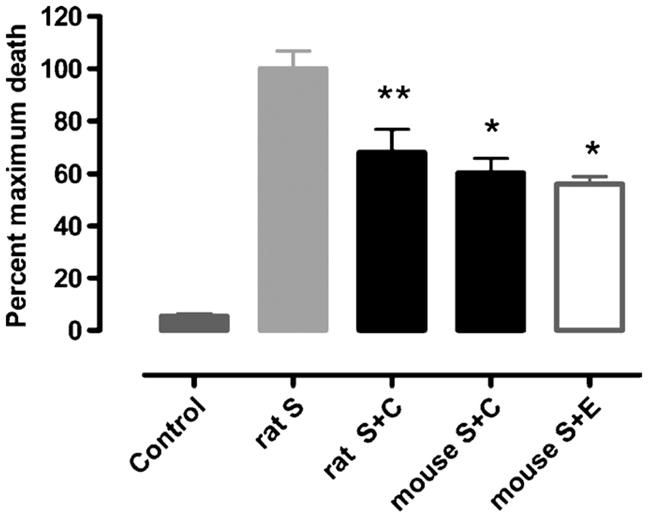
The nonhematopoietic derivative carbamoylated EPO protects cardiomyocytes from staurosporine-induced apoptosis in vitro. Mean percentage of myocytes having undergone apoptosis after isolation from rat or mouse hearts. Shown are data for rat control (n = 9), S (Staurosporine, n = 10), rat S plus C (S plus CEPO, n = 10), mouse S plus C (n = 4), and mouse S plus E (S plus EPO; n = 4). *, P < 0.05; **, P < 0.01 versus staurosporine. Reprinted with permission from Proc Natl Acad Sci U S A.54
Clearly, the observation of different biological effects of EPO and CEPO support the concept of receptor isoforms. As previously noted, the TPR is pharmacologically distinct from (EPOR)2. Specifically, it exhibits a lower affinity for EPO, forms distinct molecular species in cross-linking experiments,3 and when immunoprecipitated under nonreducing conditions, the EPOR monomer is covalently bound to the βCR (CD131). Previous work has shown that CD131 form complexes with EPOR116 and with the alpha receptor subunits specific for granulocyte macrophage colony-stimulating factor, IL-3, and IL-5. However, because normal hematopoiesis occurs in CD131 knockout mice, it was concluded that this association was not biologically relevant. Thus, there are 2 different EPOR complexes, one accounting for its hematopoietic and the other for its tissue-protective activities. As expected, in CD131 knockout mice, neither EPO nor CEPO is tissue protective. For example, Figure 4 shows that while wild-type mice respond well to CEPO in the setting of spinal cord injury, neither EPO nor CEPO is protective in CD131 knockout mice.
FIGURE 4.
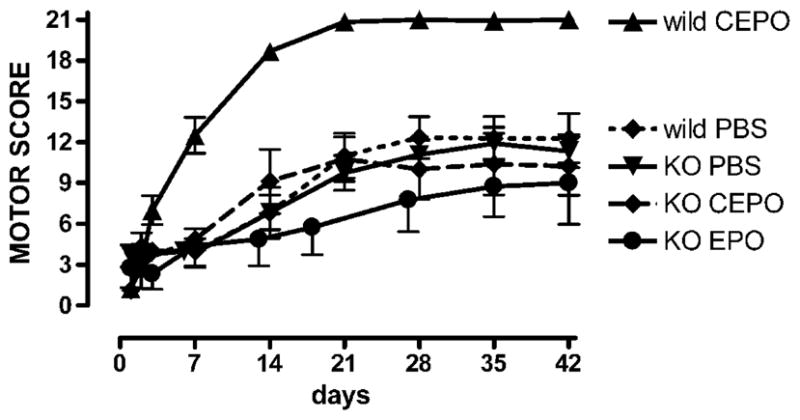
βCR knockout mice subjected to spinal cord compressive injury do not respond to either rhEPO or CEPO. Normal or CD131 knockout male mice of 8 to 16 weeks of age received a moderate compressive lesion of the spinal cord, followed immediately by a single intraperitoneal dose of rhEPO or CEPO (10 μg/kg of body weight: the equivalent of 1000 IU of EPO) and were subsequently evaluated for motor function for 6 weeks. Mortality was similar between groups (≈10%–20%). Wild-type mice responded to CEPO with a complete recovery within 4 weeks. In contrast, βcR knockout animals exhibited little recovery in motor function among the CEPO, rhEPO, or saline groups after 6 weeks.4 Reprinted with permission from Proc Natl Acad Sci U S A.4
In an effort to further characterize the structure-activity relationships of EPO, our team produced a series of peptides based on the primary sequence of EPO as well as peptides designed to mimic its 3-dimensional spatial structure. The most interesting peptides are derived from residues of EPO that are not involved in binding to the classical (EPOR)2 complex (ie, regions of EPO other than sites 1 and 2; Fig. 2). These peptides were then assayed for tissue/neuroprotective and hematopoeitic potency. As a result of these studies, helix B was identified as a tissue protective region of EPO. Of note, helix B does not contain lysine so it is not modified by carbamoylation. It is the only helix not involved with binding to the homodimeric receptor.
One of the peptides, ARA 290, is currently in clinical development by Araim Pharmaceuticals, a spin-off of Warren Pharmaceuticals. ARA 290 is a remarkable 11 amino acid peptide composed of the spatially adjacent amino acids on the portion of helix B that faces outward into the aqueous medium (Fig. 5). ARA 290 was designed to mimic the tertiary structure of helix B117 and thus acts as a ligand for the TPR. Despite the fact that ARA 290 has a short half-life (~2 minutes), it is at least equipotent on a molar basis to rhEPO, because like asialoEPO,101 it has a rapid effect on gene expression and, in this way, triggers protection responses.118 The tissue protective effects of ARA 290 have demonstrated using a large number of animal models, including models of sciatic nerve injury, middle cerebral artery ischemia, renal ischemia-reperfusion injury, and wound healing.117 For example, in mice subjected to 30 minutes of ischemia followed by reperfusion, administration of ARA 290 after reperfusion is associated with preservation of function (Fig. 6) and a reduction of tissue injury.117 In addition, like EPO, ARA 290 also has positive effects in some normal tissues, for example, effects on cognition.
FIGURE 5.

ARA 290 was synthesized based on the linear sequence of helix B indicated by the dashed ellipse in Figure 2 (boxed region; single letter amino acid code; U: pyroglutamate). Circled residues show those amino acid residues on the aqueous face of helix B and a linear peptide comprising only these residues was manufactured. Reprinted with permission from Proc Natl Acad Sci U S A.117
FIGURE 6.

ARA 290 improves renal ischemia-reperfusion injury. Renal function was assessed by measuring plasma creatinine in mice (n = 12 each group) 24 hours after being subjected to sham operation or renal ischemia–reperfusion injury (bilateral renal pedicle occlusion for 30 min). Phosphate-buffered saline (PBS) or ARA 290 (8.0 nmol/kg of body weight) was administered intraperitoneally 1 minute, 6 hours, and 12 hours into reperfusion. Data represent mean and SEM; ***, P < 0.001 versus PBS. Reprinted with permission from Proc Natl Acad Sci U S A.117
SAFETY OF ARA 290
Extensive preclinical safety studies have been carried out by Araim Pharmaceuticals. The results of these studies show that ARA 290 even at very high dosages has no effect on hematopoiesis or any toxic activities in rabbits, rats, or mice. Safety trials have been carried successfully through phase 1a and 1b. ARA 290 is currently poised to enter proof-of-concept studies.
CONCLUSIONS
Erythropoietin locally produced in response to cellular stress plays many roles in the prevention, attenuation, and termination of injury. Erythropoietin also promotes healing and restoration of function in diverse tissues. However, the strong suppressive effects of proinflammatory cytokines produced by activation of the innate immune response tend to limit the beneficial effects of locally produced EPO. In contrast, the TPR is strongly up-regulated in the vicinity of injury, providing the rationale for systemic exogenous EPO administration. Tissue protection requires a high local concentration of EPO because of the relatively low affinity of the TPR. Engineered molecules that bind only to TPR avoid hematopoietic complications and appear well suited for translation of EPO-mediated tissue protection into the clinic.
Acknowledgments
The authors thank Anthony Cerami, Michael Yamin, Thomas Coleman, Qiao-wen Xie, and many other co-workers for their critical contributions to this work. They apologize in advance to their many colleagues whose work they have not been able to discuss because of the limited space in this brief review.
Supported in part by a grant from the National Center for Research Resources (R13 RR023236).
Footnotes
C.C.H. is on the Scientific Advisory Board and owns stock in Warren Pharmaceuticals, which is pursuing the clinical development tissue protective cytokines and peptides.
M.B. is a director and shareholder of Warren Pharmaceuticals.
References
- 1.Fisher JW. Erythropoietin: physiology and pharmacology update. Exp Med Biol. 2003;228:1–14. doi: 10.1177/153537020322800101. [DOI] [PubMed] [Google Scholar]
- 2.Anagnostou A, Lee ES, Kessimian N, et al. Erythropoietin has a mitogenic and positive chemotactic effect on endothelial cells. Proc Natl Acad Sci U S A. 1990;87:5978–5982. doi: 10.1073/pnas.87.15.5978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Masuda S, Nagao M, Takahata K, et al. Functional erythropoietin receptor of the cells with neural characteristics. Comparison with receptor properties of erythroid cells. J Biol Chem. 1993;268:11208–11216. [PubMed] [Google Scholar]
- 4.Brines M, Grasso G, Fiordaliso F, et al. Erythropoietin mediates tissue protection through an erythropoietin and common beta-subunit heteroreceptor. Proc Natl Acad Sci U S A. 2004;101:14907–14912. doi: 10.1073/pnas.0406491101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sautina L, Sautin Y, Beem E, et al. Induction of nitric oxide by erythropoietin is mediated by the {beta} common receptor and requires interaction with VEGF receptor 2. Blood. 2009;115:896–905. doi: 10.1182/blood-2009-04-216432. [DOI] [PubMed] [Google Scholar]
- 6.Brines M, Cerami A. Erythropoietin-mediated tissue protection: reducing collateral damage from the primary injury response. J Int Med. 2008;264:405–432. doi: 10.1111/j.1365-2796.2008.02024.x. [DOI] [PubMed] [Google Scholar]
- 7.Brines M, Cerami A. Discovering erythropoietin’s extra-hematopoietic functions: biology and clinical promise. Kidney Int. 2006;70:246–250. doi: 10.1038/sj.ki.5001546. [DOI] [PubMed] [Google Scholar]
- 8.Brines ML, Ghezzi P, Keenan S, et al. Erythropoietin crosses the blood-brain barrier to protect against experimental brain injury. Proc Natl Acad Sci U S A. 2000;97:10526–10531. doi: 10.1073/pnas.97.19.10526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Arcasoy M. The non-haematopoietic biological effects of erythropoietin. Br J Haematol. 2008;141:14–31. doi: 10.1111/j.1365-2141.2008.07014.x. [DOI] [PubMed] [Google Scholar]
- 10.Minnerup J, Heidrich J, Rogalewski A, et al. The efficacy of erythropoietin and its analogues in animal stroke models: a meta-analysis. Stroke. 2009;40:3113–3120. doi: 10.1161/STROKEAHA.109.555789. [DOI] [PubMed] [Google Scholar]
- 11.Jerndal M, Forsberg K, Sena E, et al. A systematic review and meta-analysis of erythropoietin in experimental stroke. J Cereb Blood Flow Metab. 2010;30(5):961–968. doi: 10.1038/jcbfm.2009.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ehrenreich H, Hasselblatt M, Dembowski C, et al. Erythropoietin therapy for acute stroke is both safe and beneficial. Mol Med. 2002;8:495–505. [PMC free article] [PubMed] [Google Scholar]
- 13.Ehrenreich H, Weissenborn K, Prange H, et al. Recombinant human erythropoietin in the treatment of acute ischemic stroke. Stroke. 2009;40:e647–e656. doi: 10.1161/STROKEAHA.109.564872. [DOI] [PubMed] [Google Scholar]
- 14.Zechariah A, Elali A, Hermann D. Combination of tissue-plasminogen activator with erythropoietin induces blood-brain barrier permeability, extracellular matrix disaggregation, and DNA fragmentation after focal cerebral ischemia in mice. Stroke. 2010;41:1008–1012. doi: 10.1161/STROKEAHA.109.574418. [DOI] [PubMed] [Google Scholar]
- 15.Cherian L, Goodman JC, Robertson C. Neuroprotection with erythropoietin administration following controlled cortical impact injury in rats. J Pharmcol Exp Thera. 2007;322:789–794. doi: 10.1124/jpet.107.119628. [DOI] [PubMed] [Google Scholar]
- 16.Zhang Y, Xiong Y, Mahmood A, et al. Therapeutic effects of erythropoietin on histological and functional outcomes following traumatic brain injury in rats are independent of hematocrit. Brain Res. 2009;1294:153–164. doi: 10.1016/j.brainres.2009.07.077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lu D, Mahmood A, Qu C, et al. Erythropoietin enhances neurogenesis and restores spatial memory in rats after traumatic brain injury. J Neurotrauma. 2005;22:1011–1017. doi: 10.1089/neu.2005.22.1011. [DOI] [PubMed] [Google Scholar]
- 18.Celik M, Gökmen N, Erbayraktar S, et al. Erythropoietin prevents motor neuron apoptosis and neurologic disability in experimental spinal cord ischemic injury. Proc Natl Acad Sci U S A. 2002;99:2258–2263. doi: 10.1073/pnas.042693799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gorio A, Madaschi L, Di Stefano B, et al. Methylprednisolone neutralizes the beneficial effects of erythropoietin in experimental spinal cord injury. Proc Natl Acad Sci U S A. 2005;102:16379–16384. doi: 10.1073/pnas.0508479102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grasso G, Sfacteria A, Erbayraktar S, et al. Amelioration of spinal cord compressive injury by pharmacological preconditioning with erythropoietin and a nonerythropoietic erythropoietin derivative. J Neurosurg. 2006;4:310–318. doi: 10.3171/spi.2006.4.4.310. [DOI] [PubMed] [Google Scholar]
- 21.King VR, Averill SA, Hewazy D, et al. Erythropoietin and carbamylated erythropoietin are neuroprotective following spinal cord hemisection in the rat. Eur J Neurosci. 2007;26:90–100. doi: 10.1111/j.1460-9568.2007.05635.x. [DOI] [PubMed] [Google Scholar]
- 22.Vitellaro-Zuccarello L, Mazzetti S, Madaschi L, et al. Chronic erythropoietin-mediated effects on the expression of astrocyte markers in a rat model of contusive spinal cord injury. Neuroscience. 2008;151:452–466. doi: 10.1016/j.neuroscience.2007.11.004. [DOI] [PubMed] [Google Scholar]
- 23.Yoo JY, Won YJ, Lee JH, et al. Neuroprotective effects of erythropoietin posttreatment against kainate-induced excitotoxicity in mixed spinal cultures. J Neurosci Res. 2009;87:150–163. doi: 10.1002/jnr.21832. [DOI] [PubMed] [Google Scholar]
- 24.Kaiser K, Texier A, Ferrandiz J, et al. Recombinant human erythropoietin prevents the death of mice during cerebral malaria. J Infect Dis. 2006;193:987–995. doi: 10.1086/500844. [DOI] [PubMed] [Google Scholar]
- 25.Wiese L, Hempel C, Penkowa M, et al. Recombinant human erythropoietin increases survival and reduces neuronal apoptosis in a murine model of cerebral malaria. Malaria J. 2008;7:3. doi: 10.1186/1475-2875-7-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Casals-Pascual C, Idro R, Gicheru N, et al. High levels of erythropoietin are associated with protection against neurological sequelae in African children with cerebral malaria. Proc Natl Acad Sci U S A. 2008;105:2634–2639. doi: 10.1073/pnas.0709715105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhu C, Kang W, Xu F, et al. Erythropoietin improved neurologic outcomes in newborns with hypoxic-ischemic encephalopathy. Pediatrics. 2009;124:e218–e226. doi: 10.1542/peds.2008-3553. [DOI] [PubMed] [Google Scholar]
- 28.Brown M, Eichorst D, Lala-Black B, et al. Higher cumulative doses of erythropoietin and developmental outcomes in preterm infants. Pediatrics. 2009;124:e681–e687. doi: 10.1542/peds.2008-2701. [DOI] [PubMed] [Google Scholar]
- 29.Agnello D, Bigini P, Villa P, et al. Erythropoietin exerts an anti-inflammatory effect on the CNS in a model of experimental autoimmune encephalomyelitis. Brain Res. 2002;952:128–134. doi: 10.1016/s0006-8993(02)03239-0. [DOI] [PubMed] [Google Scholar]
- 30.Savino C, Pedotti R, Baggi F, et al. Delayed administration of erythropoietin and its non-erythropoietic derivatives ameliorates chronic murine autoimmune encephalomyelitis. J Neuroimmunol. 2006;172:27–37. doi: 10.1016/j.jneuroim.2005.10.016. [DOI] [PubMed] [Google Scholar]
- 31.Yuan R, Maeda Y, Li W, et al. Erythropoietin: a potent inducer of peripheral immuno/inflammatory modulation in autoimmune EAE. PLoS ONE. 2008;3:e1924. doi: 10.1371/journal.pone.0001924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Grasso G. Neuroprotective effect of recombinant human erythropoietin in experimental subarachnoid hemorrhage. J Neurosurg Sci. 2001;45:7–14. [PubMed] [Google Scholar]
- 33.Grasso G, Buemi M, Alafaci C, et al. Beneficial effects of systemic administration of recombinant human erythropoietin in rabbits subjected to subarachnoid hemorrhage. Proc Natl Acad Sci U S A. 2002;99:5627–5631. doi: 10.1073/pnas.082097299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Miskowiak K, Favaron E, Hafizi S, et al. Effects of erythropoietin on emotional processing biases in patients with major depression: an exploratory fMRI study. Psychopharm. 2009;207:133–142. doi: 10.1007/s00213-009-1641-1. [DOI] [PubMed] [Google Scholar]
- 35.Miskowiak K, O’Sullivan U, Harmer CJ. Erythropoietin reduces neural and cognitive processing of fear in human models of antidepressant drug action. Biol Psychiatry. 2007;62:1244–1250. doi: 10.1016/j.biopsych.2007.01.011. [DOI] [PubMed] [Google Scholar]
- 36.Ehrenreich H, Hinze-Selch D, Stawicki S, et al. Improvement of cognitive functions in chronic schizophrenic patients by recombinant human erythropoietin. Mol Psychiatry. 2006;12:206–220. doi: 10.1038/sj.mp.4001907. [DOI] [PubMed] [Google Scholar]
- 37.Lauria G, Campanella A, Filippini G, et al. Erythropoietin in amyotrophic lateral sclerosis: a pilot, randomized, double-blind, placebo-controlled study of safety and tolerability. Amyotroph Lateral Scler. 2009;10:410–415. doi: 10.3109/17482960902995246. [DOI] [PubMed] [Google Scholar]
- 38.Ehrenreich H, Fischer B, Norra C, et al. Exploring recombinant human erythropoietin in chronic progressive multiple sclerosis. Brain. 2007;130:2577–2588. doi: 10.1093/brain/awm203. [DOI] [PubMed] [Google Scholar]
- 39.Calvillo L, Latini R, Kajstura J, et al. Recombinant human erythropoietin protects the myocardium from ischemia-reperfusion injury and promotes beneficial remodeling. Proc Natl Acad Sci U S A. 2003;100:4802–4806. doi: 10.1073/pnas.0630444100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Baker J. Erythropoietin mimics ischemic preconditioning. Vasc Pharmacol. 2005;42:233–241. doi: 10.1016/j.vph.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 41.Cai Z, Manalo D, Wei G, et al. Hearts from rodents exposed to intermittent hypoxia or erythropoietin are protected against ischemia-reperfusion injury. Circulation. 2003;108:79–85. doi: 10.1161/01.CIR.0000078635.89229.8A. [DOI] [PubMed] [Google Scholar]
- 42.Cai Z, Semenza G. Phosphatidylinositol-3-kinase signaling is required for erythropoietin-mediated acute protection against myocardial ischemia/reperfusion injury. Circulation. 2004;109:2050–2053. doi: 10.1161/01.CIR.0000127954.98131.23. [DOI] [PubMed] [Google Scholar]
- 43.Rafiee P, Shi Y, Su J, et al. Erythropoietin protects the infant heart against ischemia-reperfusion injury by triggering multiple signaling pathways. Basic Res Cardiol. 2005;100:187–197. doi: 10.1007/s00395-004-0508-1. [DOI] [PubMed] [Google Scholar]
- 44.Abdelrahman M, Sharples EJ, McDonald MC, et al. Erythropoietin attenuates the tissue injury associated with hemorrhagic shock and myocardial ischemia. Shock. 2004;22:63–69. doi: 10.1097/01.shk.00001276869.21260.9d. [DOI] [PubMed] [Google Scholar]
- 45.Kim KH, Oudit GY, Backx PH. Erythropoietin protects against doxorubicin-induced cardiomyopathy via a phosphatidylinositol 3-kinase-dependent pathway. J Pharmacol Exp Ther. 2008;324:160–169. doi: 10.1124/jpet.107.125773. [DOI] [PubMed] [Google Scholar]
- 46.Tramontano A, Muniyappa R, Black A, et al. Erythropoietin protects cardiac myocytes from hypoxia-induced apoptosis through an Akt-dependent pathway. Biochem Biophys Res Commun. 2003;308:990–994. doi: 10.1016/s0006-291x(03)01503-1. [DOI] [PubMed] [Google Scholar]
- 47.Wright G, Hanlon P, Amin K, et al. Erythropoietin receptor expression in adult rat cardiomyocytes is associated with an acute cardioprotective effect for recombinant erythropoietin during ischemia-reperfusion injury. FASEB J. 2004;18:1031–1033. doi: 10.1096/fj.03-1289fje. [DOI] [PubMed] [Google Scholar]
- 48.Depping R, Kawakami K, Ocker H, et al. Expression of the erythropoietin receptor in human heart. J Thorac Cardiovasc Surg. 2005;130:877–878. doi: 10.1016/j.jtcvs.2004.12.041. [DOI] [PubMed] [Google Scholar]
- 49.van der Meer P, Groenveld HF, Januzzi JL, et al. Erythropoietin treatment in patients with chronic heart failure: a meta-analysis. Heart. 2009;95:1309–1314. doi: 10.1136/hrt.2008.161091. [DOI] [PubMed] [Google Scholar]
- 50.Mancini D, Katz S, Lang C, et al. Effect of erythropoietin on exercise capacity in patients with moderate to severe chronic heart failure. Circulation. 2003;107:294–299. doi: 10.1161/01.cir.0000044914.42696.6a. [DOI] [PubMed] [Google Scholar]
- 51.Binbrek AS, Rao NS, Al Khaja N, et al. Erythropoietin to augment myocardial salvage induced by coronary thrombolysis in patients with ST segment elevation acute myocardial infarction. Am J Cardiol. 2009;104:1035–1040. doi: 10.1016/j.amjcard.2009.05.050. [DOI] [PubMed] [Google Scholar]
- 52.Ferrario M, Arbustini E, Massa M, et al. High-dose erythropoietin in patients with acute myocardial infarction: a pilot, randomised, placebo-controlled study. Int J Cardiol. 2009 doi: 10.1016/j.ijcard.2009.10.028. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 53.Parsa C, Matsumoto A, Kim J, et al. A novel protective effect of erythropoietin in the infarcted heart. J Clin Invest. 2003;112:999–1007. doi: 10.1172/JCI18200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fiordaliso F, Chimenti S, Staszewsky L, et al. A nonerythropoietic derivative of erythropoietin protects the myocardium from ischemia-reperfusion injury. Proc Natl Acad Sci U S A. 2005;102:2046–2051. doi: 10.1073/pnas.0409329102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tabira T, Konishi Y, Gallyas F. Neurotrophic effect of hematopoietic cytokines on cholinergic and other neurons in vitro. Int J Develop. 1995;13:241–252. doi: 10.1016/0736-5748(94)00020-4. [DOI] [PubMed] [Google Scholar]
- 56.Yamaji R, Okada T, Moriya M, et al. Brain capillary endothelial cells express two forms of erythropoietin receptor mRNA. Eur J Biochem. 1996;239:494–500. doi: 10.1111/j.1432-1033.1996.0494u.x. [DOI] [PubMed] [Google Scholar]
- 57.Siren AL, Fratelli M, Brines M, et al. Erythropoietin prevents neuronal apoptosis after cerebral ischemia and metabolic stress. Proc Natl Acad Sci U S A. 2001;98:4044–4049. doi: 10.1073/pnas.051606598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Konishi Y, Chui DH, Hirose H, et al. Trophic effect of erythropoietin and other hematopoietic factors on central cholinergic neurons in vitro and in vivo. Brain Res. 1993;609:29–35. doi: 10.1016/0006-8993(93)90850-m. [DOI] [PubMed] [Google Scholar]
- 59.Viviani B, Bartesaghi S, Corsini E, et al. Erythropoietin protects primary hippocampal neurons increasing the expression of brain-derived neurotrophic factor. J Neurochem. 2005;93:412–421. doi: 10.1111/j.1471-4159.2005.03033.x. [DOI] [PubMed] [Google Scholar]
- 60.Bernaudin M, Marti HH, Roussel S, et al. A potential role for erythropoietin in focal permanent cerebral ischemia in mice. J Cereb Blood Flow Metab. 1999;19:643–651. doi: 10.1097/00004647-199906000-00007. [DOI] [PubMed] [Google Scholar]
- 61.Siren AL, Knerlich F, Poser W, et al. Erythropoietin and erythropoietin receptor in human ischemic/hypoxic brain. Acta neuropathologica. 2001;101:271–276. doi: 10.1007/s004010000297. [DOI] [PubMed] [Google Scholar]
- 62.Chin K, Yu X, Beleslin-Cokic B, et al. Production and processing of erythropoietin receptor transcripts in brain. Mol Brain Res. 2000;81:29–42. doi: 10.1016/s0169-328x(00)00157-1. [DOI] [PubMed] [Google Scholar]
- 63.Santhanam AV, Katusic ZS. Erythropoietin and cerebral vascular protection: role of nitric oxide. Acta Pharmacol Sin. 2006;27:1389–1394. doi: 10.1111/j.1745-7254.2006.00441.x. [DOI] [PubMed] [Google Scholar]
- 64.Gunnarson E, Song Y, Kowalewski J, et al. Erythropoietin modulation of astrocyte water permeability as a component of neuroprotection. Proc Natl Acad Sci U S A. 2009;106:1602–1607. doi: 10.1073/pnas.0812708106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wojchowski DM, Gregory RC, Miller CP, et al. Signal transduction in the erythropoietin receptor system. Exper Cell Res. 1999;253:143–156. doi: 10.1006/excr.1999.4673. [DOI] [PubMed] [Google Scholar]
- 66.Wang L, Zhang ZG, Gregg SR, et al. The Sonic hedgehog pathway mediates carbamylated erythropoietin-enhanced proliferation and differentiation of adult neural progenitor cells. J Biol Chem. 2007;282:32462–32470. doi: 10.1074/jbc.M706880200. [DOI] [PubMed] [Google Scholar]
- 67.Byts N, Samoylenko A, Fasshauer T, et al. Essential role for Stat5 in the neurotrophic but not in the neuroprotective effect of erythropoietin. Cell Death Differ. 2008;15:783–792. doi: 10.1038/cdd.2008.1. [DOI] [PubMed] [Google Scholar]
- 68.Um M, Gross A, Lodish H. A “classical” homodimeric erythropoietin receptor is essential for the antiapoptotic effects of erythropoietin on differentiated neuroblastoma SH-SY5Y and pheochromocytoma PC-12 cells. Cellular signalling. 2007;19:634–645. doi: 10.1016/j.cellsig.2006.08.014. [DOI] [PubMed] [Google Scholar]
- 69.Pfeffer MA, Burdmann EA, Chen C-Y, et al. A trial of darbepoetin alfa in type 2 diabetes and chronic kidney disease. N Engl J Med. 2009;361:2019–2032. doi: 10.1056/NEJMoa0907845. [DOI] [PubMed] [Google Scholar]
- 70.Bohlius J, Schmidlin K, Brillant C, et al. Recombinant human erythropoiesis-stimulating agents and mortality in patients with cancer: a meta-analysis of randomised trials. Lancet. 2009;373:1532–1542. doi: 10.1016/S0140-6736(09)60502-X. [DOI] [PubMed] [Google Scholar]
- 71.Haiden N, Cardona F, Schwindt J, et al. Changes in thrombopoiesis and platelet reactivity in extremely low birth weight infants undergoing erythropoietin therapy for treatment of anaemia of prematurity. Thromb Haemost. 2005;93:118–123. doi: 10.1160/TH04-02-0093. [DOI] [PubMed] [Google Scholar]
- 72.Kato S, Amano H, Ito Y, et al. Effect of erythropoietin on angiogenesis with the increased adhesion of platelets to the microvessels in the hind-limb ischemia model in mice. J Pharmacol Sci. 2010;112:167–175. doi: 10.1254/jphs.09262fp. [DOI] [PubMed] [Google Scholar]
- 73.Kirkeby A, Torup L, Bochsen L, et al. High-dose erythropoietin alters platelet reactivity and bleeding time in rodents in contrast to the neuroprotective variant carbamyl-erythropoietin (CEPO) Thromb Haemost. 2008;99:720–728. doi: 10.1160/TH07-03-0208. [DOI] [PubMed] [Google Scholar]
- 74.Stasko J, Galajda P, Ivanková J, et al. Soluble P-selectin during a single hemodialysis session in patients with chronic renal failure and erythropoietin treatment. Clinical and applied thrombosis/hemostasis. 2007;13:410–415. doi: 10.1177/1076029607303348. [DOI] [PubMed] [Google Scholar]
- 75.Stohlawetz PJ, Dzirlo L, Hergovich N, et al. Effects of erythropoietin on platelet reactivity and thrombopoiesis in humans. Blood. 2000;95:2983–2989. [PubMed] [Google Scholar]
- 76.Wolf RF, Gilmore LS, Friese P, et al. Erythropoietin potentiates thrombus development in a canine arterio-venous shunt model. Thromb Haemost. 1997;77:1020–1024. [PubMed] [Google Scholar]
- 77.Wolf RF, Peng J, Friese P, et al. Erythropoietin administration increases production and reactivity of platelets in dogs. Thromb Haemost. 1997;78:1505–1509. [PubMed] [Google Scholar]
- 78.Bohlius J, Schmidlin K, Brillant C, et al. Erythropoietin or Darbepoetin for patients with cancer–meta-analysis based on individual patient data. Cochrane Database Syst Rev. 2009;(3):CD007303. doi: 10.1002/14651858.CD007303.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Diskin C, Stokes T, Dansby L, et al. Beyond anemia: the clinical impact of the physiologic effects of erythropoietin. Sem Dialysis. 2008;21:447–454. doi: 10.1111/j.1525-139X.2008.00443.x. [DOI] [PubMed] [Google Scholar]
- 80.Fandrey J, Dicato M. Examining the involvement of erythropoiesis-stimulating agents in tumor proliferation (erythropoietin receptors, receptor binding, signal transduction), angiogenesis, and venous thromboembolic events. The Oncologist. 2009;14(suppl 1):34–42. doi: 10.1634/theoncologist.2009-S1-34. [DOI] [PubMed] [Google Scholar]
- 81.Napolitano L, Fabian T, Kelly K, et al. Improved survival of critically ill trauma patients treated with recombinant human erythropoietin. J Trauma. 2008;65:285–297. doi: 10.1097/TA.0b013e31817f2c6e. [DOI] [PubMed] [Google Scholar]
- 82.Kertesz N, Wu J, Chen THP, et al. The role of erythropoietin in regulating angiogenesis. Dev Biol. 2004;276:101–110. doi: 10.1016/j.ydbio.2004.08.025. [DOI] [PubMed] [Google Scholar]
- 83.Ribatti D, Presta M, Vacca A, et al. Human erythropoietin induces a pro-angiogenic phenotype in cultured endothelial cells and stimulates neovascularization in vivo. Blood. 1999;93:2627–2636. [PubMed] [Google Scholar]
- 84.Haroon Z, Amin K, Jiang X, et al. A novel role for erythropoietin during fibrin-induced wound-healing response. Am J Pathol. 2003;163:993–1000. doi: 10.1016/S0002-9440(10)63459-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Masuda S, Kobayashi T, Chikuma M, et al. The oviduct produces erythropoietin in an estrogen- and oxygen-dependent manner. Am J Physiol. 2000;278:E1038–E1044. doi: 10.1152/ajpendo.2000.278.6.E1038. [DOI] [PubMed] [Google Scholar]
- 86.Yasuda Y, Musha T, Tanaka H, et al. Inhibition of erythropoietin signalling destroys xenografts of ovarian and uterine cancers in nude mice. Br J Cancer. 2001;84:836–843. doi: 10.1054/bjoc.2000.1666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Hardee M, Cao Y, Fu P, et al. Erythropoietin blockade inhibits the induction of tumor angiogenesis and progression. PLoS ONE. 2007;2:e549. doi: 10.1371/journal.pone.0000549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Yasuda Y, Masuda S, Chikuma M, et al. Estrogen-dependent production of erythropoietin in uterus and its implication in uterine angiogenesis. J Biol Chem. 1998;273:25381–25387. doi: 10.1074/jbc.273.39.25381. [DOI] [PubMed] [Google Scholar]
- 89.Acs G, Acs P, Beckwith SM, et al. Erythropoietin and erythropoietin receptor expression in human cancer. Cancer Res. 2001;61:3561–3565. [PubMed] [Google Scholar]
- 90.Acs G, Zhang P, McGrath C, et al. Hypoxia-inducible erythropoietin signaling in squamous dysplasia and squamous cell carcinoma of the uterine cervix and its potential role in cervical carcinogenesis and tumor progression. Am J Pathol. 2003;162:1789–1806. doi: 10.1016/S0002-9440(10)64314-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fu P, Jiang X, Arcasoy M. Constitutively active erythropoietin receptor expression in breast cancer cells promotes cellular proliferation and migration through a MAP-kinase dependent pathway. Biochem Biophys Res Commun. 2009;379:696–701. doi: 10.1016/j.bbrc.2008.12.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kumar S, Yu H, Fong D, et al. Erythropoietin activates the phosphoinositide 3-kinase/Akt pathway in human melanoma cells. Melanoma Res. 2006;16:275–283. doi: 10.1097/01.cmr.0000222594.60611.c3. [DOI] [PubMed] [Google Scholar]
- 93.Mohyeldin A, Lu H, Dalgard C, et al. Erythropoietin signaling promotes invasiveness of human head and neck squamous cell carcinoma. Neoplasia. 2005;7:537–543. doi: 10.1593/neo.04685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Paragh G, Kumar SM, Rakosy Z, et al. RNA interference-mediated inhibition of erythropoietin receptor expression suppresses tumor growth and invasiveness in A2780 human ovarian carcinoma cells. Am J Pathol. 2009;174(4):1504–1514. doi: 10.2353/ajpath.2009.080592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Westenfelder C, Baranowski RL. Erythropoietin stimulates proliferation of human renal carcinoma cells. Kid Internat. 2000;58:647–657. doi: 10.1046/j.1523-1755.2000.00211.x. [DOI] [PubMed] [Google Scholar]
- 96.Liu WM, Powles T, Shamash J, et al. Effect of haemopoietic growth factors on cancer cell lines and their role in chemosensitivity. Oncogene. 2004;23:981–990. doi: 10.1038/sj.onc.1207294. [DOI] [PubMed] [Google Scholar]
- 97.Rosti V, Pedrazzoli P, Ponchio L, et al. Effect of recombinant human erythropoietin on hematopoietic and non-hematopoietic malignant cell growth in vitro. Haematologica. 1993;78:208–212. [PubMed] [Google Scholar]
- 98.Elliott S, Busse L, Bass M, et al. Anti-EPO receptor antibodies do not predict EPO receptor expression. Blood. 2006;107:1892–1895. doi: 10.1182/blood-2005-10-4066. [DOI] [PubMed] [Google Scholar]
- 99.Sinclair AM, Rogers N, Busse L, et al. Erythropoietin receptor transcription is neither elevated nor predictive of surface expression in human tumour cells. Br J Cancer. 2008;98:1059–1067. doi: 10.1038/sj.bjc.6604220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Swift S, Ellison A, Kassner P, et al. Absence of functional EpoR expression in human tumor cell lines. Blood. 2010;115(21):4254–4263. doi: 10.1182/blood-2009-10-248674. [DOI] [PubMed] [Google Scholar]
- 101.Erbayraktar S, Grasso G, Sfacteria A, et al. Asialoerythropoietin is a nonerythropoietic cytokine with broad neuroprotective activity in vivo. Proc Natl Acad Sci U S A. 2003;100:6741–6746. doi: 10.1073/pnas.1031753100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Leist M, Ghezzi P, Grasso G, et al. Derivatives of erythropoietin that are tissue protective but not erythropoietic. Science. 2004;305:239–242. doi: 10.1126/science.1098313. [DOI] [PubMed] [Google Scholar]
- 103.Boissel JP, Lee WR, Presnell SR, et al. Erythropoietin structure-function relationships. Mutant proteins that test a model of tertiary structure. J Biol Chem. 1993;268:15983–15993. [PubMed] [Google Scholar]
- 104.Cheetham JC, Smith DM, Aoki KH, et al. NMR structure of human erythropoietin and a comparison with its receptor bound conformation. Nature Struct Biol. 1998;5:861–866. doi: 10.1038/2302. [DOI] [PubMed] [Google Scholar]
- 105.Elliott S, Lorenzini T, Chang D, et al. Mapping of the active site of recombinant human erythropoietin. Blood. 1997;89:493–502. [PubMed] [Google Scholar]
- 106.Wen D, Boissel JP, Showers M, et al. Erythropoietin structure-function relationships. Identification of functionally important domains. J Biol Chem. 1994;269:22839–22846. [PubMed] [Google Scholar]
- 107.Coleman TR, Westenfelder C, Togel FE, et al. Cytoprotective doses of erythropoietin or carbamylated erythropoietin have markedly different procoagulant and vasoactive activities. Proc Natl Acad Sci U S A. 2006;103:5965–5970. doi: 10.1073/pnas.0601377103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Erbayraktar S, de Lanerolle N, de Lotbiniere A, et al. Carbamylated erythropoietin reduces radiosurgically-induced brain injury. Mol Med. 2006;12:74–80. doi: 10.2119/2006-00042.Erbayraktar. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Fantacci M, Bianciardi P, Caretti A, et al. Carbamylated erythropoietin ameliorates the metabolic stress induced in vivo by severe chronic hypoxia. Proc Natl Acad Sci U S A. 2006;103:17531–17536. doi: 10.1073/pnas.0608814103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Mennini T, De Paola M, Bigini P, et al. Nonhematopoietic erythropoietin derivatives prevent motoneuron degeneration in vitro and in vivo. Mol Med. 2006;12:153–160. doi: 10.2119/2006-00045.Mennini. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Fu Z-Q, Shao Q-L, Shen J-L, et al. Effect of carbamylated erythropoietin on major histocompatibility complex expression and neural differentiation of human neural stem cells. J Neuroimmunol. 2010;221:15–24. doi: 10.1016/j.jneuroim.2010.01.016. [DOI] [PubMed] [Google Scholar]
- 112.Imamura R, Okumi M, Isaka Y, et al. Carbamylated erythropoietin improves angiogenesis and protects the kidneys from ischemia-reperfusion injury. Cell Transplant. 2008;17:135–141. doi: 10.3727/000000008783907044. [DOI] [PubMed] [Google Scholar]
- 113.Mahmood A, Lu D, Qu C, et al. Treatment of traumatic brain injury in rats with erythropoietin and carbamylated erythropoietin. J Neurosurg. 2007;107:392–397. doi: 10.3171/JNS-07/08/0392. [DOI] [PubMed] [Google Scholar]
- 114.Wang Y, Zhang ZG, Rhodes K, et al. Post-ischemic treatment with erythropoietin or carbamylated erythropoietin reduces infarction and improves neurological outcome in a rat model of focal cerebral ischemia. British journal of pharmacology. 2007;151:1377–1384. doi: 10.1038/sj.bjp.0707285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Xu X, Cao Z, Cao B, et al. Carbamylated erythropoietin protects the myocardium from acute ischemia/reperfusion injury through a PI3K/Akt-dependent mechanism. Surgery. 2009;146:506–514. doi: 10.1016/j.surg.2009.03.022. [DOI] [PubMed] [Google Scholar]
- 116.Jubinsky PT, Krijanovski OI, Nathan DG, et al. The beta chain of the interleukin-3 receptor functionally associates with the erythropoietin receptor. Blood. 1997;90:1867–1873. [PubMed] [Google Scholar]
- 117.Brines M, Patel NS, Villa P, et al. Nonerythropoietic, tissue-protective peptides derived from the tertiary structure of erythropoietin. Proc Natl Acad Sci U S A. 2008;105:10925–10930. doi: 10.1073/pnas.0805594105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Sakanaka M, Wen TC, Matsuda S, et al. In vivo evidence that erythropoietin protects neurons from ischemic damage. Proc Natl Acad Sci U S A. 1998;95:4635–4640. doi: 10.1073/pnas.95.8.4635. [DOI] [PMC free article] [PubMed] [Google Scholar]