

Abstract
A popular hypothesis regarding legged locomotion is that humans and other large animals walk and run in a manner that minimizes the metabolic energy expenditure for locomotion. Here, using numerical optimization and supporting analytical arguments, I obtain the energy-minimizing gaits of many different simple biped models. I consider bipeds with point-mass bodies and massless legs, with or without a knee, with or without a springy tendon in series with the leg muscle and minimizing one of many different ‘metabolic cost’ models—correlated with muscle work, muscle force raised to some power, the Minetti–Alexander quasi-steady approximation to empirical muscle metabolic rate (from heat and ATPase activity), a new cost function called the ‘generalized work cost’ Cg having some positivity and convexity properties (and includes the Minetti–Alexander cost and the work cost as special cases), and generalizations thereof. For many of these models, walking-like gaits are optimal at low speeds and running-like gaits at higher speeds, so a gait transition is optimal. Minimizing the generalized work cost Cg appears mostly indistinguishable from minimizing muscle work for all the models. Inverted pendulum walking and impulsive running gaits minimize the work cost, generalized work costs Cg and a few other costs for the springless bipeds; in particular, a knee-torque-squared cost, appropriate as a simplified model for electric motor power for a kneed robot biped. Many optimal gaits had symmetry properties; for instance, the left stance phase was identical to the right stance phases. Muscle force–velocity relations and legs with masses have predictable qualitative effects, if any, on the optima. For bipeds with compliant tendons, the muscle work-minimizing strategies have close to zero muscle work (isometric muscles), with the springs performing all the leg work. These zero work gaits also minimize the generalized work costs Cg with substantial additive force or force rate costs, indicating that a running animal's metabolic cost could be dominated by the cost of producing isometric force, even though performing muscle work is usually expensive. I also catalogue the many differences between the optimal gaits of the various models. These differences contain information that might help us develop models that better predict locomotion data. In particular, for some biologically plausible cost functions, the presence or absence of springs in series with muscles has a large effect on both the coordination strategy and the absolute cost; the absence of springs results in more impulsive (collisional) optimal gaits and the presence of springs leads to more compliant optimal gaits. Most results are obtained for specific speed and stride length combinations close to preferred human behaviour, but limited numerical experiments show that some qualitative results extend to other speed-stride length combinations as well.
Keywords: legged locomotion, walking and running, optimization and optimal control, minimize energy, gaits, metabolic cost
1. Introduction
Human walking and running gaits are so stereotypical [1] that many simple, albeit subjective, descriptions of these gaits have been devised by locomotion researchers over the years. Inverted pendulum walking and impulsive running are two such idealized gait descriptions involving impulsive leg work. In inverted pendulum walking [2–5], the hip moves in a series of circular arcs, vaulting over a straight leg. The transition from one circular arc to the next is accomplished by an impulsive push-off by the trailing leg, followed by an impulsive heel-strike by the leading leg. Other versions of inverted pendulum walking have also been proposed, with various degrees of overlap between heel-strike impulse and push-off impulse [4]. In impulsive running [4–6], the hip describes a series of parabolic free-flight arcs, interrupted by impulsive elastic-looking bounces off a near-vertical leg.
More recently, the centre of mass motion of human running and walking gaits has been described a little better by the so-called spring-mass models [7–12]. These spring-mass models embody the observation that during walking and running, the leg performs mechanical work more gently than in the ‘impulsive gaits’ described above, undergoing some compression and restitution as if the whole leg were a linear spring.
Why do humans walk and run in the manner approximately described by these simple descriptions? More generally, why do humans and other animals move the way they do? An ancient hypothesis, dating back at least to a contemporary of Galileo and Newton [13], is that animals move in a manner that minimizes effort, perhaps as quantified by metabolic cost per distance travelled [14–16]. Many researchers have attempted to compute the consequences of such energetic optimality for biped models of varying complexity and realism with varying predictive success (e.g. [5,17–27]). Despite considering relatively different models and assumptions, many of these calculations have resulted in gaits that resemble those of a human in some manner. But it is hard to determine the specific reasons for such agreement as the models are often relatively complex. Also, such model complexity prevents us from appreciating the simple structure of the optimal gaits hiding just beneath the surface, as it were.
Here, using careful numerical optimization and some supporting analytical arguments, I examine biped models with four different leg architectures, minimizing a number of different ‘metabolic cost’ models—correlated with muscle work, muscle force raised to some power, a quasi-steady approximation to empirical metabolic rates, and generalizations and combinations thereof. I find that the optimal gaits for many of these model variations are similar to each other, with the qualitative differences often predictable simply. For instance, I define a class of metabolic cost functions with some positivity and convexity properties called the generalized work cost Cg, which includes as a special case both the quasi-steady empirical model and the work cost. Minimizing cost functions from this class of functions gives optimal solutions similar to simply minimizing muscle mechanical work, even though some of these cost functions have large costs for isometric force. Also, I consider biped models that have zero, small or considerable tendon compliance, so the effect of tendon compliance on the optimal gaits can be seen clearly.
A central problem in locomotion biomechanics is the determination of a simple cost function, if it exists, which when minimized for a simple biped model with appropriate task constraints, correctly and quantitatively predicts many aspects of human locomotor behaviour. This paper contributes to the solution of this inverse optimization problem by organizing, to some extent, the consequences of various model assumptions to the optimal gaits. The specific results of the various gait optimizations for the various biped model variations are described in detail in §§7–10. In §12, these results are summarized into a number of observations (about 15 in number) about the structure of energy optimal gaits for these simple biped models. In this section, it is argued that a number of features (gait structure, walk–run transition, various symmetries, walking on straight legs, apparent minimization of work, passive dynamics, linear spring-like leg behaviour, unavoidability of joint work, etc.) that characterize human gait might follow from energy minimization. Other significant results and observations (e.g. the ‘optimality of holding still’ and the structure of optimal leg-swing strategies) are noted elsewhere in the paper in appropriate sections and appendices.
2. Four biped models
The four biped models shown in figure 1 are most closely related to those used by Alexander [20,21] and Minetti & Alexander [23]. All four biped models have point-mass bodies and massless legs, embodying the observation that the upper body is usually much more massive than the legs (a human leg is about 15% body mass; [28]). The four biped models differ only in the properties of the leg.
Figure 1.

A sequence of simple biped models with different leg architectures. In each biped model, only one leg is shown; the second leg is identical to the one shown. All models have point-mass upper bodies and massless legs. (a) The leg is a telescoping actuator (muscle) with no spring (tendon) in series. (b) The leg has a knee joint, articulated by a muscle without a springy tendon in series. The two segments are of equal length. (c) The leg is a telescoping actuator (muscle) in series with a spring (tendon). (d) The leg has a knee joint, articulated by a muscle in series with a springy tendon. (e–h) Visual abbreviations for each of the four biped models are shown below the corresponding model. These visual abbreviations will be used in other figures to indicate what model those figures refer to.
The simplest model is shown in figure 1a, first introduced by Alexander [20] and elaborated later by Srinivasan & Ruina [5,29] and Srinivasan [30]. In this model, the leg is a telescoping actuator with no elastic spring in series with it; here, the telescoping actuator is identified with the sum total of all muscles articulating all joints, assuming that the muscles have no springy tendons in series.
In the model of figure 1b, the massless leg has two segments of equal length connected at a knee, with a uniarticular extensor muscle, without a springy tendon in series. This muscle has a constant moment arm about the joint, so that the joint torque is a constant multiple of the muscle force.
Animals have springy tendons in series with their muscles. These tendons are capable of storing elastic energy when stretched and releasing the energy while shortening with little loss of stored energy, thus potentially lowering the metabolic cost [31,32]. The biped models in figure 1c,d attempt to capture this feature by adding a ‘linear’ spring in series with the muscles (actuators) in the previous two models.
The properties of the telescoping actuators and muscles in these biped models are discussed in §5. Figure 1e–h show visual abbreviations for each of the four models to be used in other figures in this article.
2.1. Equations of motion for the bipeds
All the equations (figures) in this paper are written (drawn) in terms of non-dimensional quantities. All quantities are non-dimensionalized by dividing by appropriate combinations of the maximum leg length lmax, the body mass m and the acceleration due to gravity g. This non-dimensionalization, same as in Srinivasan & Ruina [5] and Srinivasan [30] with slightly different notation, is equivalent to choosing units such that the leg length, the body mass and the acceleration due to gravity are all equal to 1 in their respective units.
The most general non-dimensional equations describing the motion of our biped models are
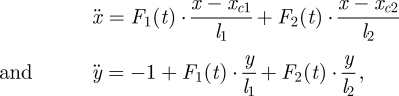 |
2.1 |
where (x, y) is the non-dimensionalized position of the point-mass upper body (centre of mass) in the sagittal plane, as shown in figure 1. These equations apply directly to the double support phase, when both feet contact the ground. Here, F1(t) and F2(t) are the compressive leg forces as functions of time, acting along the lines joining the feet and the upper body, xc1 and xc2 are the x-positions of the two feet when in ground contact and 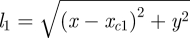 and
and 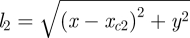 are the lengths of the two legs when they are both in ground contact. The equations of motion when only one foot touches the ground (single-stance phase) or when neither foot touches the ground (flight phase) are obtained by setting one or both leg forces to zero in equation (2.1).
are the lengths of the two legs when they are both in ground contact. The equations of motion when only one foot touches the ground (single-stance phase) or when neither foot touches the ground (flight phase) are obtained by setting one or both leg forces to zero in equation (2.1).
The non-dimensional stride period is T, the non-dimensional average speed is V = [x(T) − x(0)]/T and the non-dimensional stride length is D = [x(T) − x(0)]. If vfwd is the dimensional average speed and dstride is the dimensional stride length, then the non-dimensional speed is 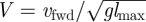 (essentially the Froude number; [33]) and the non-dimensional stride length is D = dstride/lmax.
(essentially the Froude number; [33]) and the non-dimensional stride length is D = dstride/lmax.
3. Gait optimization problem statement in brief
For each of the four biped models, I seek periodic gaits with at most one stance phase per leg per stride, having a given average speed V, stride length D and stride period T = D/V (all non-dimensional), such that the metabolic cost over the stride period is as small as possible.
The metabolic cost minimized is one of many metabolic cost models discussed in §5. The leg forces and/or muscle forces are constrained either by simple bounds or more elaborate force–velocity bounds. The non-dimensional leg length l(t) is constrained to be at most 1. The goal is to determine the optimal values for the leg forces as functions of time F1(t) and F2(t), appropriate initial conditions, the time periods over which each leg contacts the ground, and the foot–ground contact points.
See the electronic supplementary material for a more detailed formulation of the optimization problems, and the numerical methods used in their solution. The numerical methods are similar to those used earlier by Srinivasan [5,30], with modifications required by the changes in the biped models and cost functions. The electronic supplementary material has brief notes on handling the combinatorial structure of possible bipedal gaits and the regularization of non-smooth functions and kinematic singularities.
Each optimization is performed for a given [V, D] pair, so ideally one would like to perform gait optimizations for a large number of [V, D] pairs, thereby determining the dependence of the optimal gait on [V, D] as in Srinivasan & Ruina [5] and Srinivasan [30]. But because many biped models with many model variations are considered here, it is feasible to explore only a few different [V, D] pairs in detail. For the gait optimizations described in this paper, I chose [V, D] pairs mostly on the preferred human speed–stride length relation, given by the simple expression D = 2.5V0.6, as used by Alexander [20,21]. However, for many of the models, I also explored a few other [V, D] pairs not close to this preferred human behaviour and briefly allude to the results from these calculations in §12. Other [V, D] pairs are also implicitly described in the analytical calculations of appendix C.
Finally, it is thought that humans and animals move in a manner that roughly minimizes the total metabolic cost per unit distance travelled [14,16,34,35] when not in a hurry. However, when both speed V and stride length D are specified, as in the optimization calculations here, minimizing the metabolic cost per unit time Ėm, the metabolic cost per unit distance Ėm/V, and the metabolic cost per stride Ėm·D/V are all mathematically equivalent, and will necessarily give the same optimal gaits (see [16] for a discussion).
4. Total metabolic cost of locomotion
For simplicity, I model the total metabolic cost for locomotion as being separable into three additive parts: (i) resting cost, (ii) stance cost, and (iii) leg-swing cost [2,29,36].
Resting cost. The resting cost Ėr is a constant equal to the nominal metabolic rate at rest. For most of this paper, the objective is to determine gaits that minimize the energetic cost per unit distance travelled at specified average speeds V and stride lengths D. When both the speed and stride length are specified, the resting cost per unit distance (Ėr/V speed) only contributes a constant to the total cost per distance, and, therefore, drops out of the optimization. An aside: in many models without a resting cost, the total cost per distance is a strictly increasing function of the forward speed, making the optimal walking speed zero; the resting cost makes the optimal speed non-zero [37,38], but as described above, is not relevant when the speed is specified. See [16] for an account of issues related to optimal walking and running speeds.
Stance cost. The stance cost over a stride is the metabolic expenditure of the legs, either the telescoping actuator or the knee muscle in figure 1, as the legs exert force and perform work on the upper body while in contact with the ground (stance phase). This stance cost depends in detail on the motion of the centre of mass. For almost all of this paper, the goal is to determine gaits that minimize the stance cost for a given speed and stride length.
Swing cost. The possible cost of swinging the legs is neglected, as is consistent with massless legs. I constrain both the speed and the stride length in the optimizations because it is usually thought that a leg-swing cost is important for determining the optimal stride length when given only the forward speed [30,36]. The effect of adding legs with masses, with an attendant cost for leg swing [28], is briefly discussed in §11. As noted in Srinivasan & Ruina [5], if the leg-swing cost is a pure function of V, D and T (which it is not quite), it drops out of the optimization at any specified [V, D].
5. Muscle models
5.1. Metabolic cost models and force–velocity relations
One might expect the energy optimal gaits to depend somewhat on the specific properties of the muscles and actuators. A muscle model specifies how the muscle's length lm, shortening rate vm, activation a (0 ≤ a ≤ 1) and possibly other relevant muscle state variables (like the strain distribution of the bound cross-bridge population) affect the tensional muscle force Fm (≥0) and the metabolic energy expenditure of the muscle.
Work cost Cw. The simplest muscle model considered here expends energy at a rate proportional to a weighted sum of the positive and negative mechanical power produced by it (as in [5,20]):
| 5.1 |
where P = Fmvm is the mechanical power of the muscle, the positive part [P]+ of the power is defined as: [P]+ = P when P ≥ 0 and [P]+ = 0 when P < 0. The integral of [P]−(=[−P]+) gives the negative work. This cost model is motivated by the observation that a muscle's metabolic rate is somewhat correlated with mechanical power, but with different positive constants of proportionality b1 and b2 (inverse efficiencies) for positive and negative work, respectively [4,39]. The primary purpose of this simple model here is to serve as a reference for what ‘minimizing work’ entails, and will often be called the ‘work cost’. Note that isometric force generation (force without work) has no cost in this model and negative work cannot be stored in the muscle to perform positive work later.
Generalized work costCg. It is useful to define a (new) more general cost function Cg of the form:
| 5.2 |
where g(vm) is some positive and convex function of the muscle-shortening velocity, and the muscle force obeys Fm ≥ 0. The work cost Cw (equation (5.1)) is a special case of Cg with g(vm) = b1[vm]+ + b2[vm]−. Another simple special case is a linear combination of a work cost and the integral of muscle force, suggested by Alexander [40], with g(vm) = b1[vm]++b2[vm]− + b3. The function g(vm) is the ‘cost per unit force magnitude’ at the shortening rate vm. Cg is called the generalized work cost because minimizing Cg often gives optimal behaviour similar to minimizing Cw, even though it can have substantial costs for isometric force.
Convexity and positivity of g(vm) will be key determinants of the qualitative structure of the optimal gaits. In §6, I elaborate on what one means by convexity and show that convexity is necessary to ensure meaningful optimal solutions in a simple but fundamental example of ‘supporting a weight’. In appendix B, I discuss another important task that suggests positivity as a plausible property. In this paper, I will only use Cg with positive and convex g(vm) in the optimizations. The g(vm) for Cw is piecewise linear and (barely) convex.
Alexander–Minetti quasi-steady empirical costCAM. Next, I consider a simplified Hill-type model, borrowed from Alexander and Minetti [23,41,42]. In this model, the muscle force depends on muscle activation a and shortening velocity vm, but not directly on muscle length lm. In the following, vmax is the maximum shortening velocity at which a positive muscle force can be produced, and F0 is the isometric muscle force when fully activated (vm = 0, a = 1). Figure 2a,b shows the normalized force Fm/aF0 = ϕ(vm/vmax) and the normalized empirical cost rate ĊAM/aF0vmax = γ(vm/vmax) as a function of the normalized muscle-shortening velocity vm/vmax. This is a quasi-steady muscle metabolic cost model, an approximation to some roughly steady-state constant-vm experimental data on muscle heat generation and ATPase activity at full activation (reviewed in [43,44]), with an extra assumption of linear dependence of both the force and the cost rate on the muscle activation.
Figure 2.

A quasi-steady muscle model. Dependence of (a) the muscle force Fm and (b) the muscle metabolic rate ĊAM on the activation a and normalized shortening velocity vm/vmax, given the isometric force F0 and the maximum shortening velocity vmax at which the muscle can produce force. (c) The Alexander–Minetti empirical metabolic rate per unit muscle force based on heat and ATPase activity ĊAM/Fmvmax = g(vm/vmax). (d) The work cost per unit force Ċw/Fmvmax = (b1[vm]++b2[vm]−)/vmax. The b1 and b2 for (d) were selected so that it most closely resembled (c).
Observe that CAM can also be written in the form Cg (equation (5.2)). The g(vm) = ĊAM/Fm = vmaxγ/ϕ for CAM is depicted in figure 2c. A g(vm) for Cw with appropriately chosen b1 and b2 is shown in figure 2d to emphasize the similarity of Alexander–Minetti empirical metabolic cost CAM and the work cost Cw. This apparent similarity between the cost CAM and the work cost Cw, and the specific plots (figure 2c,d) were first suggested to me by A. Ruina (2008, personal communication). The g(vm) for CAM is convex or close-to-but-not-quite convex depending on the approximation of the experimental data.
Muscle force costs and robot electric motor costs. Another cost function commonly used in the gait optimization literature is:
| 5.3 |
I mainly consider β = 2, giving CF2. The β = 1 case is an exceptional case of Cg, with g(v) = constant (see §6 for comment). The CF2 cost or a related activation-squared cost has been used by numerous authors as a proxy for a (metabolic) cost function, especially in the context of inverse dynamics, when one wishes to compute the individual muscle forces consistent with an observed body motion when there are many more muscles than degrees of freedom in the model [24,45,46]. While this procedure often gives reasonable agreement with muscle EMG data, there is no direct biophysical basis for such a cost depending purely on the muscle force; it contradicts the so-called Fenn effect [47] and (equivalently) the muscle data that motivate the cost CAM just described. When the force is replaced by a torque, CF2 is also perhaps the simplest cost model for the energy cost for electric motors and therefore relevant for legged robots1 [48–51].
Cost combinations. The metabolic cost functions described so far (equations (5.1)–(5.3) and figure 2b) are only simple models. So I also consider convex combinations of some cost functions: for instance, C = λCw + (1 − λ)CF2, where 0 ≤ λ≤ 1. Such cost combinations are not meant as actual models for metabolic costs, but are considered mainly to examine the robustness of the optimal solutions to perturbations in the costs.
Force–velocity relations. Among the numerous optimizations performed for this paper, only a few involved a physiological force–velocity relation, such as that shown in figure 2a. For the rest of the optimizations, only a simple muscle force bound is used. Sometimes, a linear force–velocity relationship, such as Fm ≤ F0(1 − vm/vmax), completely specified by F0 and vmax, was used for simplicity of computation.
5.2. Muscle forces and length rates
The above metabolic cost functions and the force–velocity relations are expressed in terms of the muscle force Fm and the muscle-shortening rate vm. Because of the differences in body architectures, I now describe what specifically is meant by Fm and vm in each of the four models.
Springless telescoping biped. The muscle is identical to the telescoping actuator, so the muscle force is identical to the leg force F. The muscle-shortening velocity is equal to the leg lengthening rate (vm = l̇).
Springless kneed biped. The muscle force is proportional to the knee moment and the muscle length shortening rate is proportional to −α̇, the negative of the knee angle rate. The knee moment M is given by M = −(F/2)· sin(α/2), where F is the leg force and α is the knee angle. The knee angle by l = cos(α/2). The knee angular velocity α̇ is given by: l̇ = −α̇ sin(α/2)/2. The knee muscle force is Fm = |M|/rm, where rm is the constant moment arm of the muscle about the knee joint. Similarly, the knee muscle-shortening rate is vm = −rmα̇. The actual value of the moment arm rm plays no role in this paper, because all the calculations—the evaluations of the metabolic cost, the dynamics and the constraints—are expressed directly in terms of the knee moment M and the knee angular velocity α̇.
Springy telescoping biped. The muscle force is identical to the leg force F here. The total leg length l is the sum of two lengths: tendon spring's lspring and the telescoping actuator's lactuator. When the spring is linear with non-dimensional stiffness k, the leg force is equal to F = −k(lspring − l0), where l0 is the spring's stress-free length. The spring length rate is then l̇spring = −Ḟ/k. The actuator length rate is l̇actuator = l̇ − l̇spring. The muscle-shortening rate is vm = l̇actuator. In this paper, the quantities lactuator and lspring appear in calculations either as their derivatives or their sum l, so that their individual values are irrelevant and left undetermined up to a constant.
Springy kneed biped. The muscle force is proportional to the knee moment as before (Fm = |M|/rm). The knee flexion angle α is accommodated by changes in the spring length and the muscle length, which are quantified in terms of the knee angle changes they are responsible for: αspring and αmuscle. The spring deflection is related to the knee moment by M = −kαspring = −(F/2) · sin(α/2), assuming a torsional spring stiffness k. The spring angular rate is given by α̇spring = −Ṁ/k = (Ḟ/2k) sin(α/2) + (Fα̇/4k) cos(α/2). The muscle angular rate is then α̇muscle = α̇ − α̇spring. The muscle length shortening rate is vm = −rmα̇muscle. Again, the moment arm rm of the muscle drops out of all calculations here.
6. ‘Holding still’ is optimal for Cg if g(v) is convex
In §5, a class of candidate muscle cost functions called the generalized work cost, Cg = ∫Fg(v) dt, was introduced. Before this class of cost functions is used in the locomotion optimization problems, I use this cost function in the context of the much simpler task of ‘exerting a given average force’ using a muscle, with the least cost. This problem is described to show why g(v) must be strictly convex around v = 0 and what happens if g(v) were not convex. This section is perhaps slightly digressive (but fundamental), and the reader may skip to the next section without much loss of continuity.
Supporting an average force. Consider the situation shown in figure 3c. The goal is to use the muscle to support the mass (m = 1) at some vertical position on average, in the presence of gravity, while minimizing the cost Cg with a strictly convex g(v), over some long time period (or per unit time). I wish to show that holding the mass exactly at rest for all time (that is, holding still) is less expensive than, say, moving the mass about an average position periodically with any period however long.
Figure 3.

(a) A convex function. For the convex function, the function value (solid line) at some velocity v is less than or equal to any weighted average (dotted line) of the function values at two or more velocities (say v1 and v2) whose weighted average is v. (b) A non-convex function. For the non-convex function, there exist velocities v1 and v2 such that the aforementioned inequality is violated. (c) The objective is to keep the mass at a given position on average, with zero average velocity. (d) The objective is to exert a given average force on the wall, with possibly some periodic motion of the point P.
So consider, as an alternative, a time-varying periodic force F(t), producing a time-varying vertical velocity v =− ẏ. Say the time period of this oscillation is tp. The equation of motion is v̇ = F − 1 (gravity constant =1). The periodicity equations imply
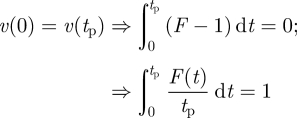 |
6.1 |
and
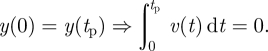 |
6.2 |
Definition: strictly convex function. A function g(v) is said to be strictly convex if for every v1 ≠ v2 in its domain, it satisfies
| 6.3 |
where 0 < μ< 1. In words, the ‘function value at a weighted average of two v's’ is less than the ‘weighted average of the function values at two v's’. More colloquially, convex functions are functions that are ‘bowl shaped’. Figure 3a,b shows examples of convex and non-convex functions. In figure 3a,b, if some point on the solid line represents the left-hand side of equation (6.3), the point on the dotted straight line with the same abscissa (v-value) represents the right-hand side of equation (6.3).
Proof of optimality. Strictly convex functions obey the so-called Jensen's inequality [52]:
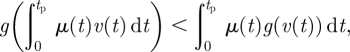 |
6.4 |
where μ(t) is any non-constant non-negative (piecewise continuous) function that satisfies ∫0tpμ(t) = 1 and v(t) is non-constant. Equation (6.4) is essentially a generalization of equation (6.3) in the sense that equation (6.4) involves weighted averages over a possibly continuous distribution of v, as opposed to averages over only two distinct v's in equation (6.3). Setting μ(t) = F(t)/tp in equation (6.3),
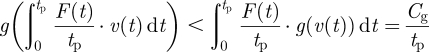 |
6.5 |
for non-constant F(t). The right-most equality with Cg/tp above assumes that F(t) is positive. Also, ∫0tpF(t)v(t) dt = 0 because this is the mechanical work done over one period and integrates to zero by periodicity. So, equation (6.5) simplifies to
| 6.6 |
That is, the cost Cg with any non-constant periodic F(t) over time tp will always be greater than the g(0)tp, which can only be attained by keeping the mass absolutely still: v(t) ≡ 0. Thus, ‘holding still’ is the unique optimal strategy (at least from among periodic motions of arbitrarily long periods) if g(v) is strictly convex.
Effect of non-convexity. If g(v) were not convex (concave) around v = 0, for instance, as in figure 3b, there exists a time-varying μ(t), and therefore F(t), for which the above inequalities are reversed; that is, Cg ≤ g(0)tp and a time-varying F(t) (‘tremor’) would be better than holding still. In the non-convex example of figure 3b, one would be able to reduce the effective g(v) to the dotted line, from the solid line, by operating between v1 and v2. It is not clear if such optimality of oscillation, as opposed to rest, is physiological. A provocative conjecture would be that muscle activation is slightly ‘noisy’ to take energetic advantage of the possible slight non-convexity in the cost function, a hypothesis I hope to examine elsewhere in greater detail. To keep things simple, g(v) is restricted to convex functions here.
Note that strict convexity of g(v) is sufficient but not necessary for the above result. Optimality of holding still only requires that the inequality (equation (6.3)) be strict when the weighted average of v evaluates to v = 0. For instance, functions like g(v) = |v|, while not strictly convex away from v = 0, do have holding to be optimal for this task. Most significantly, note also that the optimality of holding still does not depend at all on the minimum of g(v) being at v = 0; for instance, even if g(v) = 1 + (v − 1)2, holding still (v(t) ≡ 0) is optimal. Nor is it necessary that g(v) be positive.
Another situation. The optimality of holding still also applies to other situations; for instance, that shown in figure 3d, in which it is required that the muscle exert an average force of unit magnitude on the wall, as it perhaps oscillates against the spring. Set v = ẋa. By periodicity, xa(0) = xa(tp) and xs(0) = xs(tp), giving ∫0tp F(t)v(t) dt = 0 (no net work done). Jensen's inequality (equation (6.5)) again applies if g(v) is strictly convex, giving g(0)tp < Cg, demonstrating the optimality of holding still.
Positivity. In appendix B, a simple task analogous to swinging a leg is considered. In this leg swing task, positivity of g(v) is required to avoid some implausible optimal strategies. Positivity of g(v) is natural; it simply means that the metabolic rate is positive whether the muscle is performing positive or negative work. Thus, in this paper, g(v) is both positive and convex.
7. Springless telescoping biped
As the simplest in the series of four models (figure 1a–d), I discuss the optimal gaits of the springless telescoping biped (figure 1a) in greater detail than that of the other models. Srinivasan and Ruina [5,30] considered the simplest version of this model, determining the periodic gaits that minimized the work cost Cw at a range of speeds and step lengths [V, D], assuming that the leg forces can be unbounded if necessary. They discovered that inverted pendulum walking was optimal at low speeds, impulsive running was optimal at higher speeds and a third hybrid gait dubbed ‘pendular running’ was optimal at a range of intermediate speeds and large step lengths. However, this calculation did not allow double support, considered only a single step, assumed that each step is the same as every other step and did not minimize multiple cost models other than Cw. In the following, I rid the calculations of these simplifications. I assume bounded leg forces, allow for double-support phases, consider two steps (equal to one stride) instead of one step, with the possibility that the steps might be different, and consider multiple muscle metabolic cost models. In the rest of the paper, unless specified otherwise, the maximum possible leg force is 3 (i.e. three times body weight). This choice for the maximum leg force has no special significance except being sufficiently greater than 1 to support many gaits, but not unrealistically high. Values such as 2.5 or 3.5 give qualitatively the same answer for a range of [V, D].
Minimize work. Minimizing Cw with these (mostly) relaxed constraints does not produce any gaits qualitatively different from those found in Srinivasan & Ruina [5]. Mostly, I found that non-impulsive analogues of the three impulsive gaits—namely, inverted pendular walking, impulsive running and pendular running [5]—minimized Cw. That is, the infinite forces required during the impulses of the three previously optimal impulsive gaits are replaced by the maximum available leg forces (here, equal to 3) in these non-impulsive gaits.
For example, figure 4a–c shows the non-impulsive analogue of inverted pendular walking, optimal when V = 0.39 and D = 1.36. This optimal walking gait has negligible double-support phases, despite allowing for them. A substantial double support is optimal only when the maximum leg force available is too close to or less than 1, when a single leg is unable to support body weight. For most of the stance phase, the leg length is constant and no leg work is performed. Negative work is performed at the beginning of the stance phase, akin to a heel-strike. Positive work is performed at the end of each stance phase, akin to a push-off.
Figure 4.

(a–c) A non-impulsive analogue of an inverted pendulum walking with the springless telescoping model, minimizing the work cost Cw at V = 0.39, D = 1.36 and Fmax = 3. The gait consists of the body moving in circular arcs vaulting over a straight leg, with the transition from one circular arc to the next accomplished by finite-force impulses (with F ∼ Fmax) by the trailing leg and the leading leg, identified with the push-off and the heel-strike, respectively. (d–f) Optimal running with springless telescoping model, minimizing the work cost Cw at V = 1.2, D = 2.8 and Fmax = 3. The running stance phase has the leg force at Fmax identically. The graphs corresponding to the two legs' stance phases are overlaid in (b,c,e,f) and they appear indistinguishable, indicating left–right symmetry.
Figure 4d–f shows the non-impulsive analogue of impulsive running, found to be optimal at V = 1.2 and D = 2.8. Here, the leg force is equal to the maximum allowed leg force throughout the stance phase. Negative work is performed for the first half of the stance phase, and positive work during the second half, redirecting the centre of mass velocity from downward to upward.
The energy cost landscape appears to be relatively flat near the transition from walking to running. Figure 5a–c shows the non-impulsive analogue of pendular running [5], optimal at V = 0.9 and D = 1.17. Figure 5d–f shows a very different asymmetric gait, but with a cost only 1 per cent greater than that of the optimal pendular run at the same speed and stride length. The energy cost landscape in the [V, D] space with the finite leg forces is probably similar qualitatively to when the optimal gaits involved idealized impulses in Srinivasan & Ruina [5] and Srinivasan [30], so it is not plotted here.
Figure 5.

Two near-optimal gaits at V = 0.9, D = 2.34 and Fmax = 3 for the springless telescoping biped. (a–c) The non-impulsive analogue of pendular running. (d–f) A hybrid asymmetric gait, with one stance phase pendular and the other stance more compliant. The cost of the second gait is only about 1 per cent more than that of the first gait.
Adding a force–velocity relation. First, recall that all three optimal gaits obtained above with only a simple bound on the leg forces made use of the maximum available forces available to them at some point during the stance phase. So by continuation, when a non-trivial force–velocity relationship is imposed for the telescoping actuator, one still expects the optimal gaits to take advantage of the maximum available forces. However, now, the maximum available forces are dependent on the leg length rate—in particular, the maximum available force when the leg is performing negative work is greater than when the leg is performing positive work. This suggests that the leg force will cease to be symmetric about mid-stance. Figure 6a–c shows the effect of a linear force–velocity relation on the walking gait of figure 4a–c. As just argued, the leg force is slightly asymmetric about mid-stance—the push-off is slightly different from the heel-strike—but this difference has little effect on the centre of mass trajectory, which remains quite close to circular arcs. Similarly, figure 6d–f shows the optimal running gait (V = 1.2, D = 2.8) when constrained by a linear force–velocity relation. The leg force hugs the force–velocity constraint, just as when there was a simple bound on the force, the leg force was identically equal to the maximum force.
Figure 6.

Optimal gaits when the springless telescoping actuator has a linear force–velocity relation F ≤ F0(1 − l̇/vmax) with F0 = 3, and vmax = 2. (a–c) Walking gait minimizing the work cost Cw at V = 0.39 and D = 1.34, close to inverted pendulum walking. (d–f) Running gait minimizing the work cost Cw at V = 1.2 and D = 2.8. The leg force mostly takes advantage of the maximum allowed forces by the force–velocity relation, shown as a dotted line in (b,e). The plots corresponding to the two legs are overlaid and are indistinguishable in these plots, suggesting symmetry.
Minimize generalized work costsCgandCAM. As described in §5, the quasi-steady model CAM with a convex g(v) is a special case of Cg. Minimizing CAM with a force–velocity relationship (V = 0.39, D = 1.38) gave the gait depicted in figure 7a,b, again very similar to inverted pendulum walking.
Figure 7.

(a,b) Walking with the springless telescoping model, minimizing the convex empirical cost CAM at V = 0.39, D = 1.36, F0 = 3 and vmax = 0.8. (c,d) Walking with the springless telescoping model at V = 0.39 and D = 1.38, minimizing Cg with convex g(v) = v2. (e,f) Minimizing Cg with g(v) = (v − 0.5)2. All these three optimal walking gaits are quite close to inverted pendulum walking and each other.
In addition to CAM, I considered two more ad hoc examples of Cg. Figure 7c,d shows the optimal walking gait at V = 0.39, D = 1.38 for Cg with an g(v) = v2 and figure 7e,f shows the optimal walking gait for g(v) = (v − 0.5)2. Both are close to inverted pendulum walking. Numerical experiments with a few other convex and positive g(v) also result in qualitatively similar optimal gaits to that which minimizes the work cost Cw, in particular, inverted pendulum walking at low speeds. At higher speeds, running gaits (not shown) that involve leg forces that essentially reach the force bounds, be they simple force bounds or force–velocity relations, appear to be optimal. In this sense, these gaits are analogous to impulsive running. As before, when the leg force tracks the force–velocity relation, the leg force can be asymmetric about mid-stance. Overall, these results suggest that the optimal gaits for Cg are essentially similar to that for Cw.
With additive force costs. Consider a linear force penalty (i.e. CFβ with β = 1) added to the costs Cw, CAM or Cg. Then, the new metabolic power would be Fg(v) + b1 F = F(b1 + g(v)), which is also of the form Cg with a g(v) simply shifted up by the constant b1, still positive and convex. So, it is not surprising that essentially no qualitative changes are found in the optimal gaits for a linear additive force cost.
A quadratic force cost CF2 is qualitatively different, being a much stronger penalty on large forces and, less directly, force fluctuations. See appendix A for a simple example that informs the following discussion. Periodic locomotion, as a task, does not prescribe the average leg force—but periodicity does specify that the average vertical force on the body must equal the weight. At small stride lengths, when the legs do not make large angles with the vertical, this vertical force constraint is similar to an average leg force constraint. Thus, when one has a cost like CF2, one obtains gaits in which both the force fluctuations and the actual leg forces are minimized. The leg forces are reduced by having large double-support phases and the force fluctuations are reduced by having the leg forces relatively constant. In fact, at small step lengths, the optimal walking gait (not shown) tended to have mostly double support and very little single stance, an artefact that can perhaps be mitigated by having a substantial leg-swing cost. But even for a pure CF2 cost, it appears that for high enough speeds and step lengths, the optimal gait, at least locally, is one with a flight phase (figure 8).
Figure 8.

Running gait that minimizes the CF2 cost for the springless telescoping biped at V = 2, D = 3.78.
With explicit bounds on force rates. Thus far, it has been assumed that the leg force can rise from zero instantaneously if necessary. But muscles have internal dynamics that limit the rate of increase of the force they produce. To simply model this, I bound the rate of leg force change: −3 ≤ Ḟ ≤ 3, the specific values selected so the leg force rate bound is limiting, but not so limiting that a gait cannot be supported. It is further assumed that the forces must start and end at zero, and can change only continuously. Figure 9a–d shows the results of minimizing Cw at V = 0.39 and D = 1.36 with this force rate constraint. Here, the presence of double support is energetically optimal—that is, making the double-support phase zero increases the energy cost. Note that this double support is an explicit effect of bounding the force rates. Exactly the same minimization, in the absence of bounding the force rates, showed that having no double support was optimal (figure 4). Figure 9e shows the even smoother (and realistic) optimal leg forces when the second derivatives of the leg forces were appropriately bounded.
Figure 9.

(a–d) Walking gait that minimizes the work cost Cw for the springless telescoping biped, in the presence of bounds on the leg force rates, at V = 0.39 and D = 1.38, −4 ≤ Ḟ ≤ 4. (e) Leg forces for the optimal gait when the second derivative of leg force is bounded: −20 ≤ F¨ ≤ 20, without constraining F or Ḟ. It is found that a non-zero double support becomes optimal and using the maximum available leg force is no longer optimal. The specific bounds on Ḟ and F¨ determine the amount of double stance and the maximum leg forces that are optimal. The slight wiggles in (e) are an indication of near but imperfect convergence.
At higher speeds, a running gait with a unimodal (increasing–decreasing) leg force profile is obtained, with the increase and decrease in the leg force happening at the maximum available leg force rate (figure 10a–c). If the second derivative of the leg force is bounded instead, a running gait with a smoother (piecewise quadratic) unimodal leg force F(t) is obtained (figure 10d–f). Interestingly, for a range of such rate bounds, a graph between the leg force and the leg compression looks approximately linear—the so-called linear spring-like leg behaviour [9,10]—despite the presence of no springs in the calculations [53].
Figure 10.

Running gait that minimizes the work cost for the springless telescoping biped at V = 0.39 and D = 1.38, (a–c) in the presence of bounds on the leg force rates (−3 ≤ Ḟ ≤ 3) and (d–f) in the presence of bounds on the second derivative of forces (−100 ≤ F¨ ≤ 100). Both optimizations assumed a leg force bound as well (F ≤ 3). It is found that the leg force as a function of the leg compression, while nonlinear, is not far from linear on average, suggesting that the so-called linear spring-like leg behaviour at optimality requires no springs.
8. Springless kneed biped
The springless kneed biped is shown in figure 1b. The knee angle is α and the knee moment is M, positive when anti-clockwise on the upper leg. The leg length is then l = cos(α/2) and the leg length rate l̇ = −sin(α/2) · α̇/2. The compressive leg force F and the knee moment M are related as: M = −F sin (α/2). Thus, when α is small, a large leg force (F = −M/sin(α/2)) is possible with a small leg moment; also, although not explicitly accounted for by this formula, finite leg forces are possible at α = 0 with zero leg moments. A related singularity is that the knee angular velocity α̇ goes to infinity for non-zero leg length rate l̇ when α → 0; see the electronic supplementary material for how these ‘kinematic singularities’ are regularized during the optimization. The leg power is, of course, identical to the knee power: P = Fl̇ = Mα̇.
Minimizing work costCw. As just mentioned, the mechanical power P = Fl̇ of the leg is identical to the mechanical power Mα̇ of the knee muscle in the springless kneed biped. Therefore, the results of minimizing the work cost Cw for the kneed biped of figure 1b are identical to those obtained when Cw is minimized for the telescoping biped of figure 1a, in the absence of other constraints. Force constraints are now most naturally imposed on the knee moment (and, therefore, the muscle force) rather than the leg force. A simple bound on the knee moment, as long the bound is not too small, results in essentially the classic work-minimizing gaits from earlier (inverted pendulum walking, etc).
Minimizing ‘robot’ knee moment costCF2. As noted, the kinematic singularity at the knee allows a kneed biped to exert a finite leg force with very little knee moment when the leg is close to straight. Thus, when the metabolic cost is dominated by the CF2 term (here ∫M2 dt), one might superficially expect that the biped would operate near the kinematic singularity for as much of the stance phase as possible. Indeed, this is true of the optimal walking gait, for instance, as shown in figure 11 for V = 0.39 and D = 1.38. The leg is fully extended for almost all the stance, except for the beginning and the end of the stance phase, when push-offs and heel-strikes happen with large leg forces. In the limit of infinite allowed leg forces, this walking gait appears to approach the classic inverted pendulum gait described earlier. At higher speeds, it appears advantageous to break into a run (figure 12) and as the allowed leg forces are increased, the optimal gait approaches the impulsive running gait, even though the leg force seems somewhat U-shaped (figure 12b). Thus, remarkably, this force-based cost CF2, which is completely unrelated to the work cost Cw, gives essentially the same optimal gaits for this kneed biped, at least in the impulsive limit.
Figure 11.

Walking gait that minimizes the CF2 cost for a springless kneed biped with no force–velocity constraint, at V = 0.39, D = 1.38 and with Fmax = 3. Again, this gait is quite close to inverted pendulum walking, with no double support.
Figure 12.

Running gait that minimizes the CF2 cost for a springless kneed biped with no force–velocity constraint, at V = 1.2, D = 2.8 and with Fmax = 3. When the maximum allowed forces is increased, the running gait at this [V, D] approaches impulsive running, although the leg force pattern is still U-shaped.
With a force–velocity relation. The force–velocity relation is now a constraint on the knee moment M as a function of the knee angular velocity α̇. The simplest such constraint is linear. Noting that a positive α̇ corresponds to the knee muscle stretching, the linear force–velocity relation used is
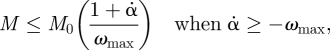 |
8.1 |
and M ≡ 0 when α̇ < ωmax. Again, M0 and ωmax were selected such that they support gaits, but are not too high, as can be gleaned from the figures referred to below.
Because of the kinematic singularity, at the end of stance, if l ≈ 1 and l̇ > 0, α̇ is negative and very large, the available knee moment M is zero, because of the force–velocity constraint equation (8.1), and unable to provide the necessary compressive leg force for the push-off. Thus, introduction of a force–velocity constraint implies that the biped might have to operate slightly away from the kinematic singularity, so that reasonable knee moments are available. Figure 13a–f shows the CF2-minimizing walking gait, when the biped is explicitly constrained to operate away from the kinematic singularity by setting lmax = 0.95 (and an extra leg force constraint Fmax = 3). The knee moment required now is well within the feasible set allowed by the force–velocity constraint. Figure 13g–l shows a Cw-minimizing running gait in the presence of force–velocity constraints on the knee muscle, constrained by Fmax = 3. When the force–velocity constraint is active, a slight asymmetry probably appears about midstance as for the telescoping model.
Figure 13.

(a–f) Walking gait that minimizes the CF2 cost for springless kneed biped with a linear force–velocity constraint on the knee moment, when V = 0.39, D = 1.38, lmax = 0.95 and Fmax = 3. Again, this gait is close to inverted pendulum walking. (g–l) Running gait that minimizes Cw for the springless kneed biped at V = 1.2 and D = 2.8, with a force–velocity relationship for the knee muscle. This gait shows more time-reversal asymmetry in the trajectory and the leg force profile than other gaits here on account of the force–velocity relation.
MinimizingCgcosts. In limited numerical explorations, minimizing Cg, including CAM and two other instances of Cg with g(v) = α̇2 and (α̇ − 0.5)2, gave rise to inverted pendulum walking and running-like gaits at low and high speeds, respectively.
9. Springy telescoping biped
Animals have compliant tendons in series with muscles. Such tendons can substantially affect the energetics of steady locomotion by storing work performed on it and returning it later during the stance. Figure 1c is perhaps the simplest biped model that has both a muscle (telescoping actuator) and a tendon (linear springs). The springs are linear, perfectly elastic and have no damping.
As a practical matter, because the muscle length rate and therefore the metabolic rate function depends directly on the muscle force rate for these springy bipeds (unlike the springless bipeds), the muscle force rates are bounded during the optimizations. In general, however, the force rate bounds were chosen to be so large (−20 ≤ Ḟ ≤ 20) that they are never taken advantage of in most of the optimal solutions described below. In this sense, it is as if no force rate bounds are used for most of the optimizations below, unless noted otherwise. Note on the other hand that most of the optimizations with the springless bipeds (except for one) involved no explicit force rate bounds.
Minimizing work costCw. Given the same constraints including leg force capabilities, this springy biped can perform every gait that can be performed by the springless biped of figure 1a and vice versa. Of particular interest are gaits that minimize the muscle work cost Cw for the springy biped. Figure 14 shows one such gait, with essentially zero work done by the actuator. All the positive and negative work is done by the series spring, while the series telescoping actuator remains at constant length (isometric contraction) and provides a force that is at all times equal to the spring's. The stiffness of the spring was chosen to be close to the ‘apparent stiffness’ of a human leg while running (non-dimensional stiffness is 16), but the actual value does not affect the qualitative conclusions, as long as the muscle is capable of sufficiently high forces and the stiffness is not too small ([12]; apparent stiffness is defined as the slope of the leg force versus leg compression curve and is not a true stiffness). The bound on the leg force was 3, higher than that required by the work-free gait—that is, this force bound is not used by the optimal gait, just as the force rate bound is also unused by the optimal gait.
Figure 14.

Springy telescoping biped: a gait that minimizes Cw for the springy telescoping model with V = 1 and D = 2.4. The muscle work is essentially equal to zero. This gait may be somewhat reminiscent of skipping gaits [11].
The work-free gaits (Cw = 0) of this biped are identical to the periodic gaits of the so-called spring-mass biped model [10,12,54,55], which has only a telescoping spring and no series actuator. For such unactuated spring-mass models, it can be shown that for a given speed, step length and spring stiffness within some bounds, there usually exists (that is, it is non-exceptional to find) one or more isolated walking and/or running gaits with left–right symmetry; see [12] and [30] for counting arguments. For an example of one such symmetric and muscle-work-free running gait, see figure 16 (although it was not obtained by minimizing Cw as explained below). If one allows that the left and the right stance phases need not be identical, there is a one-parameter family of work-free gaits—and therefore infinitely many equally optimal gaits—for specified speed, stride length and spring stiffness. The gait shown in figure 14 is one such asymmetric gait. Note that the stance phases for this gait are time-reversed versions of each other. The optimization converged to an asymmetric gait perhaps because the symmetric gait is a special case and asymmetry is generic (on the other hand, asymmetry seems ‘structurally unstable’ as noted below).
Figure 16.

Springy telescoping biped: running gait minimizing λCw + (1 − λ)CF2 for λ = 0.9, V = 1 and D = 2.4. Despite a substantial cost for isometric force, the optimal gait appears to still minimize Cw, requiring essentially zero work. The leg force versus leg compression shows a linear spring behaviour, as would be expected.
Figure 15 shows an essentially muscle-work-free walking gait for the springy telescoping biped; left–right symmetry was imposed for this optimization for simplicity and to emphasize similarity with human walking. One obtains a qualitatively similar walking gait—with double-humped leg force profile—for a range of spring stiffnesses.
Figure 15.

Walking with a springy telescoping model, with a relatively soft spring: a non-dimensional stiffness of 16, at V = 0.39 and D = 1.38, minimizing Cw with enforced left–right symmetry. The leg force rate is bounded: −20 ≤ Ḟ ≤ 20, but the bound is never used. Essentially, zero mechanical work is performed by the muscles and all the mechanical work is performed by the springs in series. The leg force versus leg compression behaves, not surprisingly, in a linear manner.
MinimizeCwwith a force penalty. That work-free gaits are obtained when Cw is minimized is to be expected, and is not surprising. But what if there is a strong cost penalty for muscle forces? Figure 16 shows a running gait that minimizes a linear combination of work cost Cw and a force cost CF2 : λCw + (1 − λ)CF2, for λ = 0.9. The muscle work performed is still essentially equal to zero, despite the metabolic cost having a substantial force cost. Thus, the same muscle-work-free gait (figure 16) minimizes the cost for both λ = 1 and λ = 0.9. More generally, it appears that the work-free gait is optimal for a range of λ near 1, another instance of the insensitivity of the optimum to changes in the cost function. This particular insensitivity of the optimum is explained in appendix D as being related to the non-smoothness of the cost at the optimum at λ = 0. Also, note that the gait in figure 16 corresponding to λ = 0.9 is symmetric, while the optimal gait (figure 14) corresponding to λ = 1 was generically asymmetric. It appears that the presence of the small force penalty CF2 makes symmetry optimal, while Cw alone has both symmetric and asymmetric optimal gaits. Figure 17 shows the optimal running gait for λ = 0.75, so that the weight on CF2 is higher than when λ = 0.9. This gait does involve some muscle mechanical work, even though work-free gaits exist, as demonstrated by figures 14 and 16. Thus, it seems like the work-free gaits are optimal for a range of λ near 1, but not optimal for lower λ, when the force penalty starts dominating the nature of the optimum.
Figure 17.

Running with the springy telescoping biped. Minimizing λCw + (1 − λ)CF2 for λ = 0.75, V = 1 and D = 2.4. As the force penalty is increased, the optimal gait moves away from being completely work-free. Negative and positive work is performed by the muscle.
MinimizeCgcosts. As §6 would suggest, minimizing Cg also results in a zero Cw gait. This is true whether or not even when there is a cost for isometric force (force with no work). As before, I used a few different g(v), including v2, (v − 0.5)2 and the g(v) for CAM. When Cg has a cost for isometric force, there is a tendency toward left–right gait symmetry.
High stiffness limit. If the leg force is not explicitly constrained, work-free gaits exist for any high stiffness of the linear tendon spring. At very high stiffnesses, the work-free running gait looks like the impulsive running gait, with parabolic free flights interrupted by brief bounces on the leg springs, with the series telescoping actuator remaining isometric.
However, such work-free gaits involve high leg forces and become infeasible when the leg forces or the leg force rates are bounded. Indeed, these force and force rate bounds become active constraints mainly in the high tendon stiffness limit. When the force rates are bounded and the optimal gaits are sought for given speed and stride length, the high stiffness limit approaches the limit of having no springs, namely the springless telescoping biped. Figure 18 shows the optimal walking gait at V = 0.39 and D = 1.38 when the stiffness is high (k = 500). Note that the gait is close to inverted pendulum walking, which is optimal in the absence of the spring, except made a little smoother by the presence of the spring. At a higher speed (V = 1.2 and D = 2.4), one obtains a running gait, in which the maximum available force rates were used, analogous to impulsive running.
Figure 18.

Walking with the springy telescoping model, with a relatively stiff spring: a non-dimensional stiffness of 500. V = 0.39 and D = 1.38. Because the spring stiffness is high, the biped largely behaves like it does not have a spring at all. This gait is close to inverted pendulum walking, with some artefacts associated with the presence of the spring. The muscle performs most of the work, and the spring performs very little.
10. Springy kneed biped
Similar to the springy telescoping biped, the springy kneed biped has work-free gaits in which the muscle remains exactly isometric throughout the stance phase, while the springy tendon performs negative work first (stretches) and positive work next (shortens), redirecting the centre of mass velocity from down to up. When there are no constraints on the forces and Cw is minimized, the optimization converges to one of the work-free gaits, corresponding to the specified speed and stride length. As before, work-free gaits continue to be optimal when the cost is slightly perturbed as λCw + (1 − λ)CF2 for a range of λ near 1. Also, the work-free gaits were found to be optimal for CAM and for a couple of other Cg's that were minimized. For example, figure 19 shows the gait-minimizing CAM (with the constant 0.5 added to its g(v)) and is essentially muscle-work-free. It is interesting to note that despite the kinematic singularity, the relation between leg force and leg length change is not far from linear.
Figure 19.

Running gait minimizing CAM with the integrand offset up by 0.5 to give a larger isometric cost with a force–velocity relation. Minimizing CAM also gives essentially the same gait. The gait is close to muscle-work-free. Also, despite the presence of the kinematic singularity, the relation between leg force and the leg compression is not far from linear. The force–velocity bound on the knee moment is not reached except briefly at the end of stance. The parameters are V = 1.2 and D = 2.8, isometric knee moment M0 = −2 and maximum knee angular rate for non-zero moment ωmax = 2 for the linear force–velocity relation. The leg length was bound by 0.97 so that the singularity is approached but never reached. The torsional stiffness for the knee tendon was taken to be 2.85, roughly consistent with what would give a duty factor near what humans use. Other nearby stiffnesses, as long as it is not too high or too low (see main text), also give muscle-work-free gaits, but with slightly different duty factors.
When the stiffness of the series spring is sufficiently high, the optimal gaits are qualitatively similar to those of the springless kneed biped, given that the leg forces are bounded in a manner that the work-free gaits are now not feasible. As an aside, a substantially different objective function of the form ∫M2(1 + α̇muscle2) gave essentially the same result, with a little more muscle work but still a small fraction of the total leg work.
Finally, when the cost is a pure function of the knee moment and not dependent on the muscle-shortening rate (for instance, CF2 = ∫M2dt), the springy kneed model and its optimization reduces to that of the springless model—as long as the force–velocity bound is not very constraining at the optimum.
While the springy kneed biped is perhaps the most realistic of all the bipeds considered in this paper, it is the least examined and discussed. I hope to expand on this model and its properties, especially as a function of the tendon stiffness, in a sequel to this paper.
11. Swinging a massive leg
In most legged locomotion, the swinging of the legs is not passive, but is powered by muscles and requires energy. On the other hand, throughout this paper, I have determined gaits that minimized only the so-called stance cost, the cost of moving and supporting the centre of mass by shortening and lengthening the leg. The effect of having to swing massive legs can be most naturally studied by endowing the model with massive legs, whose movements can be controlled by hip muscles and whose dynamics is coupled to the motion of the hip.
An alternative to considering a model with massive legs is to use an additive cost for swinging the leg (e.g. [2,36]). This additive cost could be a simple algebraic function of the leg-swing angle δs and the leg-swing time ts, the actual function being drawn from simple theory or from leg-swing experiments [28,30]. Simple expressions for a leg-swing cost can be derived by considering a rigid leg pivoting from an immovable upper body, articulated by uniarticular muscles. For swinging the leg faster than it would swing passively, the strategy that minimizes Cw (or Cg in general) has an accelerating impulse at the beginning of the swing and a decelerating impulse at the end of the swing, with the rest of the swing being essentially passive (see appendix B). The strategy that minimizes CF2, as would be relevant for legged robots, has the hip torque essentially decreasing linearly from an accelerating torque to a decelerating torque, in a manner that satisfies the swing time and swing amplitude constraint.
These additive costs were used with appropriate pre-factors in a couple of springless optimizations described so far, but the optimal gait did not change qualitatively when the speed and stride lengths were fixed. The leg-swing cost does affect the duty factor by imposing a penalty on too short a swing time, or conversely, too great a double-support phase. But inverted pendulum walking and impulsive running already have as great a swing time as possible for the given type of gait (walking versus running), with all swing times being equal by symmetry. Perhaps, this is the reason for the robustness of such optima to such additive swing costs.
To be sure, the separation of the total cost into additively separable stance and swing costs is a simplification. For instance, work performed to move the hip around (here called the stance cost) can potentially contribute to leg swing, making the separation into stance and swing costs, as defined above, generally impossible. But considering the leg swing as being separable for the purposes of understanding the structure of the optimal solution is supported in some cases by optimization results from a model with massive legs. I considered a model similar to figure 1a, except for legs having hip muscles and point-masses at their feet. I found that the Cw-minimizing gait was inverted pendulum walking with push-offs, heel-strikes and vaulting on circular arcs. And the leg swing was accomplished by hip impulses at the beginning and end of the swing. Thus, the work optimal gait for a model with massive legs consists of pasting together the optimal gait for a model with massless legs and the optimal leg-swing strategy with the legs considered in isolation. This separability of the structure of the optimal gait is probably untrue for strange animals whose body mass is dominated by their leg mass, especially if most of the leg mass is close to the feet.
12. Discussion: general observations about the optimal locomotion
The following observations about the mechanics and energetics of legged locomotion arise from the many calculations detailed in this paper and other general considerations. Many of these observations correspond to salient features of human terrestrial locomotion that the hypothesis of energy optimality is able to explain.
1. Unavoidability of a minimum amount of leg or joint work. In periodic legged locomotion on level ground, the total mechanical energy of the animal is the same at the beginning and the end of a gait period. Despite such overall energy neutrality, common experience and careful experiments tell us that our legs perform substantial work when we walk and run. Is such leg work unavoidable for legged locomotion? If so, why?
One reason for performing work is dissipation. However, the springless telescoping biped considered here had no dissipative mechanisms—no collisions and had only no-slip frictional contact.2 By minimizing Cw for the springless telescoping biped, I have shown—here and in Srinivasan & Ruina [5]—that travelling at non-infinitesimal speeds and step lengths requires non-infinitesimal positive leg work (that is, the optimal Cw does not go to zero) even in the absence of frictional or collisional dissipation, as long as the biped's body is constrained to never penetrate the ground. This minimum required positive leg work, given the lack of physical dissipation, must be exactly cancelled by the performance of an equal amount of negative leg work elsewhere during the gait.
Now imagine a more realistic biped—say, with more body segments, especially legs, with masses—with frictionless joints, slip-free frictional contact with the ground and plastic mechanical collisions that can in principle be avoided by stepping carefully. For such a more realistic biped, I believe (based on unpublished calculations and some arguments with additive leg-swing costs) that this work-unavoidability claim is even stronger: travelling at non-infinitesimal speeds requires non-infinitesimal leg work, or more generally, some joint work. That is, the condition of non-infinitesimal step lengths from the previous paragraph is unnecessary. See [56] for a related result for the special case of unactuated downhill walking ‘passive dynamic’ robots.
A potential source of confusion might be that in real human or robot walking, heel-strike collisions do appear to be a big source of energy loss, which must be made up by positive work elsewhere [4,57–59]. But this heel-strike collisional loss may be interpreted as just the animal's way of performing negative work (which is unavoidable), without using muscles and at the optimal time, i.e. at the end of stance, as in our optimal inverted pendulum walking gaits. In other words, if the motion involves collisions, the biped has at least that much energy to replace by positive work; and if the biped avoids collisions, the legs have to do both positive and negative work.
Two more clarifications are in order. First, the above claims are with respect to ‘leg or joint work’, as opposed to ‘actuator or muscle work’. Of course, locomotion is possible in principle with just spring work—as in Geyer et al. [10], O'Connor & Kuo [11] and Srinivasan & Holmes [12], and the springy bipeds here—without the series muscles performing work, but the spring work is part of the unavoidable leg work. Second, it has been conjectured by ([60]; also personal communication 2008) that it is possible to construct bipeds that have legs with closed kinematic chains that can overcome the unavoidability of leg work. My claim about the unavoidability of leg work does not apply to these unconventional bipeds; another such unconventional biped is McGeer's ‘silly wheel’ [61].
Roughly, the origins of this leg/joint work unavoidability is related to the assumption of (non-infinitesimal) finite gravity. In zero gravity, locomotion once begun—perhaps by pushing off a wall—can continue for ever with no additional leg work and therefore no cost, coasting ad infinitum. In finite gravity, at least for the simple biped models here with non-infinitesimal step lengths, the only two ways of going forward performing no work for extended periods of time during a single step is to either lock the leg joints (as in an inverted pendulum phase) or have zero leg force (as in a flight phase). Both these phases have a body trajectory that is strictly concave-downwards (d2y/dx2 < 0) because of the gravitational pull. But a periodic trajectory has to be horizontal on average, so it appears that the leg needs to do some finite amount of work to make the average value of d2y/dx2 equal zero. It remains an open mathematical problem to prove this intuition rigorously for any reasonably realistic class of bipeds with leg mass.
2. Optimality of inverted pendulum walking and impulsive running. Inverted pendulum walking and impulsive running are classic idealized descriptions of human walking and running, going back at least half a century [6,62]. Srinivasan & Ruina [5] showed that these originally subjective idealizations have a more objective existence as the work-minimizing gaits of the same simple biped models (figure 1a,b). These classic descriptions appear to minimize a vast number of cost functions for the models in figure 1a,b and the stiff-tendon limit of the models in figure 1c,d. For instance, this optimality result has been extended to minimizing a more general cost Cg, including the empirical cost CAM. These gaits also appear optimal for a vastly different cost function, namely the integral of squared knee moment M2, if the biped has a knee.
3. Double-humped vertical ground reaction force for walking. The vertical ground reaction force (GRF) for human walking has a double-humped (bimodal) profile. Somewhat analogous to this bimodal GRF, idealized and impulsive inverted pendulum walking has two large force spikes at the beginning and the end of a stance phase, and much smaller forces during the pendular part of the stance phase. When bounds are placed on how fast the leg force can increase from zero, as exists for a realistic muscle, the force spikes at the ends of inverted pendulum walking become smoother (figure 9), making the optimality of a bimodal GRF more apparent. Somewhat interestingly, the Cg-optimal gaits for the springy telescoping model also had a double-humped vertical GRF, for a range of tendon spring stiffnesses, from k = 16 to 500, at least for the specific V = 0.39 considered in detail. But note that for the springy models, work-free gaits with no flight phase but with a unimodal vertical GRF are also possible for some leg spring stiffnesses, similar to the ‘grounded running’ gaits of cockroaches and some birds [11,12].
4. ‘Bang-coast-bang’ structure of theCg-minimizing gaits in springless models. The optimal gaits for Cg for the springless models all had a simple structure. In these gaits, the mechanical power was close to zero for most of the time, either in pendular phases with l̇ = 0, or in free flight with F = 0. Thus, the gait is mostly energetically passive, with mechanical work being performed in brief episodes of large leg forces. Such a strategy is sometimes called bang-coast-bang by optimal control theorists [63]. In [5], where this structure was discovered as a consequence of minimizing Cw, it was surmised that minimizing Cw = ∫Fl̇ resulted in solutions that had large phases with F = 0 or l̇ = 0 perhaps because both these phases had the cost identically zero. However, such superficial speculation seems insufficiently general, now that such optimal gait structure has been shown to be a property of minimizing a wider class of functions Cg, especially because the optimal structure is preserved even when the minimum of g(v) is not at v = 0. In appendix B, I prove using elementary geometric arguments, the optimality of this bang-coast-bang structure for a problem related to swinging a leg through a given amplitude. And indeed it is known that human leg swing during walking has muscle activation mostly at the beginning and the end of the swing [28]. It must be pointed out that a true bang-coast-bang structure is, in detail, at odds with common experience because it seems like humans are not exerting muscles maximally during slow-speed walking. Note also that while the similarity of optimal behaviour for Cg and Cw has been formally established in simple examples (§6 and appendix B), I only considered special cases of Cg for the locomotion optimization problems, which therefore does not constitrue a complete proof that all Cg have similar optima. Finally, using the maximum available forces becomes non-optimal when there are force-derivative bounds—when using the maximum available force-derivative becomes optimal.
5. ‘Compliant’ walking and running gaits. Human gaits are relatively smooth and involve largely gradual growth of the various GRFs involved. Such smoothness has allowed these gaits to be approximated by ‘spring-mass models’, in which the leg is modelled as a massless linear spring, such as recently elaborated by a number of authors, notably [10] in the context of human walking [11,12]. The many ‘passive’ gaits obtained for these spring-mass models are all shown here to be work-optimal (and more generally Cg-optimal) in our springy actuated models.
On the other hand, when there are no springs in the biped's leg, the optimal gaits here were mostly non-impulsive analogues of gaits with impulses. However, these impulsive gaits do become smoother when muscles are modelled more realistically. For instance, a muscle's force cannot rise to some finite value instantaneously, but can only do so gradually, as allowed by its excitation–contraction dynamics. With such properties, the impulses get smoothed out, giving the gaits a compliant appearance.
6. Linear spring-like leg behaviour. Another feature, especially of human running gaits, is the roughly linear relationship between the leg force and leg compression. Interestingly, this roughly linear spring-like leg behaviour can be derived as a consequence of energy optimality in three vastly different situations. First, not surprisingly, the zero muscle work running gait of the running springy telescoping biped, in which the linear telescoping spring in series does all the leg work, has an exactly linear-spring-like leg behaviour in this limit (figure 16). More interestingly, the kneed springy biped also showed a linear-spring-like leg behaviour, but now only approximate because of the nonlinearity imposed by the knee kinematics (figure 19). At the other extreme, a springless telescoping biped or a springy biped with very high stiffness, now with a simple bound on the force rate or its derivatives, might have running gaits with a roughly linear spring-like leg behaviour—even in the absence of a spring (figure 10). Thus, a roughly linear spring-like leg behaviour is obtained in running whenever the leg force increases and decreases gradually and is unimodal—as in all these cases—independent of whether there is an actual spring involved.
7. Walk-to-run transition. One common theory of gait transition holds that animals switch to running because inverted pendulum walking requires high centripetal accelerations at high speeds, not achievable without tensional leg forces [2,30,64]. However, this theory is unsatisfactory as it does not explain why animals do not use other no-flight-phase gaits at higher speeds not requiring tensional forces, such as one with a horizontal centre of mass trajectory [30]. Here, as many others have also argued, it has been shown that energetics provide a logically satisfying explanation for gait transition.
For all the springless bipeds with the various cost functions and assumptions considered here, it was energetically optimal to transition from gaits with no flight phase at low speeds (walking) to gaits with a flight phase at higher speeds (running). This is a generalization of the analogous result in Srinivasan & Ruina [5]. In appendix C, I show why the gait transition happens at roughly the same speed-stride-length boundaries for a few different Cg-like cost functions.
The walk-to-run transition curves have not been explored well enough for different costs for the springy bipeds in this paper. But note that the springy bipeds can have work-free running at very slow speeds, if the leg forces are not constrained ([21]; also see fig. 5 of [12]). Therefore, a realistic walk-to-run transition is not predicted for work-minimizing springy bipeds for a range of not-too-high stiffnesses. But the walk–run transition claim does apply to springy bipeds with high enough spring stiffnesses and bounded leg forces, as they are similar to the springless bipeds. Also, Minetti & Alexander [23] obtained plausible gait transitions with a springy kneed model with CAM and leg mass.
8. Optimality of left–right symmetry. Human and many other bipedal animal walking and running gaits have a left–right symmetry. That is, their left stance phases are at least approximately similar to their right stance phases. This symmetry is probably explained by energetic optimality given a bilaterally symmetric body.
First, all of the optimal gaits computed here for the springless bipeds for various costs had their left stance phases essentially identical to their right stance phases, despite allowing for asymmetry. In appendix E, assuming small step lengths, it is shown that a symmetric inverted pendulum walking gait always requires less work than an inverted pendulum gait with asymmetric steps. On the other hand, I did find that the cost landscape for Cw was relatively flat near the gait transition for the springless telescoping biped, so that asymmetric gaits are close to but not quite optimal (suggesting that small perturbations of the model might make asymmetry optimal sometimes).
For springy bipeds with sufficiently compliant tendons, when muscle work Cw was minimized, it was found that both symmetric and asymmetric gaits (such as skipping; figure 14) were optimal. However, when the metabolic cost function is slightly modified with a cost for isometric force, it was often found that the flatness of the landscape was lost, the asymmetric gaits became non-optimal and the symmetric gait was picked out often as the optimal gait.
9. Time-reversal symmetry in stance phases. Many, but not all, of the optimal gaits discussed here had an additional symmetry property. Their stance phases had an approximate or an exact time-reversal symmetry. That is, these gaits look the same run forward or backward in time. Obviously, human gaits do not have this symmetry exactly, but they are not far away, especially if one looks at the GRFs and the centre of mass motion; it is this approximate symmetry that lets approximation of human running by the symmetric gaits of spring-mass models [9,10,12]. This approximate time-reversal symmetry in bipedal walking is therefore probably a consequence of energy optimality. Interestingly, note that this symmetry appears approximately true even when g(v) in Cg is far from an even function. Also, the optimal gaits for the springless models have another time-reversal symmetry, not demonstrated here: the gait that minimizes the cost Cg with g(v) is the time-reversed version of the gait that minimizes the cost Cg with g(−v), assuming the force–velocity bound is not active, neglected or also reversed.
Finally, considering models with extended as opposed to point feet will modify these time-reversal symmetries somewhat.
10. Horizontal-hip-trajectory walking. It is sometimes argued [62] that horizontal-hip-trajectory walking (sometimes called ‘level walking’; [5]) might reduce work requirements from the legs because it was thought that muscle mechanical work was required mainly to lift and lower the centre of mass. Here, when I minimized the work costs, I never discovered this horizontal-hip-trajectory gait as an optimum—this horizontal-hip-trajectory gait requires more positive leg work than the optimal gaits, as also confirmed by experiments for metabolic costs [65,66]. See [30,67] and [5] for simple cost expressions (Cw) for such level walking gaits; for constant speed level walking with double support, Cw = D2/24H; for non-constant speed level walking with no double support, Cw = D2/32H. Comparing these cost expressions with Cw for inverted pendulum walking (approx. V2D2/32), pendular running and impulsive running (approx. D2/32V2) computed accurately using careful numerics show that level walking never minimizes Cw [5,30]. Such horizontal-hip-trajectory gaits are non-optimal for most other metabolic cost functions considered here as well.
11. Walking on relatively straight legs. Almost all the optimal gaits obtained here make use of essentially all the maximum leg length available to the biped, at some part of the stance phase. For instance, in inverted pendulum walking, pendular running, impulsive running and their non-impulsive analogues, the leg is almost its maximum possible length for the whole of the stance phase. In the more compliant optimal gaits of the springy models, especially with a cost for isometric force, the leg length is maximal or close to maximal at the beginning and the end of stance phase. Classically, it has been argued that this tendency for straight legs, especially in human walking, is to reduce the muscle force requirements by taking the best advantage of the knee's kinematic singularity. However, it has been shown here (generalizing [5]) that even without a knee and even without a cost for force, it is useful to walk and run on straight legs for the springless models while minimizing a variety of metabolic costs. This trend is also clear from simple cost expressions for these idealized gaits.
12. Optimality of double support in walking. Minimizing Cw or Cg for springless bipeds at low speeds appears to produce walking gaits with no double support. That is, in these walking gaits, the push-off is entirely before the heel-strike with no overlap. This absence of double support appears to be true even when there is a not-too-low bound on the maximum leg force (say about 2–3). However, double support appears to become optimal when one has a bound on the rate of leg force (figure 9). This result shows that the optimality of push-off before heel-strike, advocated by some [3,4] is true for slow walking only when the leg forces can increase arbitrarily fast, as in a collisional impulse. Another reason to have a double support is if there is a strong cost on the leg force (say, CF2) without a knee singularity to reduce the muscle force—as for instance, is the case when the work is performed by the ankle joint. Finally, note that walking gaits with double support were also obtained as being optimal for the springy bipeds with substantial tendon compliance, while minimizing Cg-like costs.
13. Why might animals look like they minimize work instead of metabolic cost? Some animals have long compliant tendons in series with some of their muscles [40], so that during running, these tendons account for a large fraction of the positive and negative work of the musculo-tendon complex, while the muscle itself remains close to isometric [68,69]. This strategy is clearly muscle-work minimizing, at least for the muscles that are close to isometric. Even other animals appear to take advantage of the presence of tendons springs to reduce the amount of muscle mechanical work required [31,70–72]. Such running is literally close to the behavioural assumptions of the spring-mass model, although it is not clear if all major leg muscles are close to isometric (the proximal ones might not be). In any case, why care about ‘muscle work’ instead of metabolic cost? Here, it has been demonstrated that minimizing Cw is similar to minimizing a number of other costs. Thus, if animals appear to minimize work, it is probably because minimizing whatever is important to them (e.g. metabolic cost) ‘looks like’ minimizing work. Note again that minimizing work can be optimal even with substantial costs for isometric force.
14. Optimal robot gaits and biomimicry. While other researchers, especially roboticists [19,48,49,73] have determined the optimal gaits of models more complex than, but closely related to, the kneed springless model, it appears not to have been noted that a particular simplification of their models yields the classic inverted pendulum model and impulsive running as the optimum. (Note again that the M2 cost CF2 is a simple model of idealized motor electrical cost appropriate for robots). This similarity of idealized human gaits and idealized robot optimal gaits allows robots based on a blind mimicry of some aspects of human walking to be energetically economical even though the human metabolic cost function could be very different from a robot's electrical cost function. There is, of course, no a priori reason to expect that what is optimal for an animal would be optimal for a robot, but sometimes there might be coincidences such as those seen here. Note that I am not claiming that the corresponding optimal gaits are identical at any given [V, D], but only that a similar set of gaits seem optimal overall.
Thus, one could perhaps conjecture that the under-actuated level-ground walking robots [59], inspired by the so-called passive dynamic robots, inherit their energy economy from their imitation of energy optimal gaits here, which have large energetically passive phases. Of course, in general, having many largely passive phases does not imply energy economy (even though energy economy has been equated with passivity in the literature); this gait structure just happens to be good for the many costs discussed here.
15. When are springs energetically useful? Whether or not a spring in series with a muscle can be energetically useful depends on the stiffness of the spring, the maximum muscle forces, the time scale of the stance phase and the properties of the metabolic cost function. Springs in series with muscles are useful for reducing the muscle work requirements during legged locomotion. But if the muscle forces are bounded, then the spring in series can store only as much energy as this maximum muscle force lets it. So a spring that is much too stiff can store little energy in this case; so a sufficiently stiff spring would be indistinguishable from having no spring at all. At the other extreme, a spring with very low stiffness in series with the muscle can also be energetically expensive. Imagine replacing your tendons with a very thin rubber band; clearly such low stiffness would make locomotion at normal speeds essentially infeasible and if feasible, expensive (see [30] for a related discussion about leg swing; also [32]). Thus, given a motion with some time scale, it is likely that there is a range of optimal tendon stiffnesses for reducing muscle work requirements [42].
Note that springs do not only help reduce work requirements. Springs are useful in the reduction of the more general metabolic cost Cg as well. Even more generally, it seems likely that springs help minimize metabolic cost when the metabolic cost function depends strongly on the muscle contraction speed. On the other hand, if the cost were a pure function of muscle force, for instance, CF2, then clearly springs can have no direct effect on the cost—so the springy bipeds are the same as the springless bipeds. This means that robots with electrical motors with perfect regeneration might be less amenable to cost reduction using springs in series with the motors. This is also a critique of the use of CF2 or other pure functions of muscle force in predictive biomechanics optimizations—all such optimizations should find that the presence of springs have only little or no direct effect in reducing the cost (springs may have a small indirect effect on the coordination through the muscle force–velocity relation).
16. What dominates cost at an optimum? First, consider a simple concrete example. Consider a function f that is a weighted sum of two smooth functions: f(x) = λf1(x) + (1 − λ)f2(x), where f1(x) = (x − a)2 and f2(x) = (x − b)2. The optimal solution x* =λa + (1 − λ)b. By changing λ from 0 to 1, the optimum of f(x) shifts from x* = b, the minimum of f2, to x* = a, the minimum of f1. It can be shown that when λ ≈ 1, even though x* is determined by minimizing f1, the cost f is dominated by the (1 − λ)f2 term. On the other hand, when λ ≈ 0, even though x* is determined by minimizing f2, the cost f is dominated by the λf1 term. This overall result is generically true for a weighted sum of any two smooth functions.
Thus, the part of the cost function that mostly determines the optimum might be only a small part of the optimal cost. In other words, just because something costs you a lot at the optimum does not mean that this aspect of the cost is the most important determinant of the optimum strategy. Rather, it might well be the less important determinant of the optimum. The more important determinant might have a much smaller fraction at the optimum.
Thus, for instance, it might be possible for the cost of locomotion to be largely explained by the cost of isometric force (if it is, as sometimes argued; [74,75]), while the optimal gait is largely explained by the minimization of work, say. This feature is observed in many of the zero muscle work Cg-optimal gaits of the springy bipeds here.
17. Insensitivity to objective function. As noted in the introduction, one goal of research on legged locomotion is to determine a hopefully simple function that when minimized under appropriate task constraints for a simple-enough biped model predicts the observed locomotor behaviour [26]. In this paper, I have shown that the optimal gait is insensitive to some objective function perturbations. See appendix D for one simple explanation for such insensitivity involving a non-smooth minimum. Insensitivity of optima to the objective functions means that it may be harder to infer a unique objective function purely from the observed behaviour alone (if we cannot measure the cost function as well). On the other hand, once the insensitivity is better understood (or even if not), the insensitivity helps by letting us make good predictions with a relatively poor model of the objective function.
18. What about the predicted metabolic costs? In the models here and in the following discussion, I do not consider the cost of swinging the legs, which is thought to be a small fraction (perhaps about one-third) of the total cost [76]. In [5] and [30], I plotted certain aspects of the optimal work costs Cw for the springless biped models, and noted that the metabolic cost values estimated by Cw are within a factor of 2 of the metabolic cost estimated from VO2 experiments (e.g. figs 2.4 and 2.7 in [30] assuming b1 = 4 and b2 = 0.8). The leg work in these optimal gaits are also close to the so-called centre of mass work (sometimes called ‘external work’) computed from the animal's GRFs. Without changing the optimal gaits at all, much better agreement with metabolic data could presumably be achieved by considering the more general cost Cg and choosing g(v) appropriately for a springless biped, whether kneed or telescoping. This would be somewhat unsatisfactory as it is known that springs do have a role to play in the energetics.
On the other hand, the springy biped models, while producing gaits that are compliant and having more realistic looking GRFs, suffer from too low an estimate of the metabolic cost, especially if the series spring is sufficiently compliant and is performing all the leg work. In this work-free limit, Cw = 0 and Cg is mostly due to the integral of the muscle force. A slightly more realistic model with knees, tendon springs and leg mass owing to Minetti and Alexander also suffers from a similar underestimation of the metabolic cost while using CAM (see fig. 4d in [23]). While this cost underestimation might potentially be fixed by manipulating g(v), this manipulation might have to be substantially at odds with the data informing the CAM cost. Also, the perfectly muscle-work-free optimal gait structure is probably unlikely to be a good approximation anyway, as it is thought that only about 50 per cent of the leg work is due to the springs in the legs and the rest is due to muscle work [53]. Overall, I suspect that a generalization of the metabolic cost model Cg to something that looks like, say, ∫f(F)g(v) dt or ∫h(F,v) dt might be indicated, or perhaps as others have suggested, a cost function that includes force derivatives.
Summary. In this paper, I have explored many objective functions, many biped models, under many situations. This exploration resulted in many qualitative results, which I suspect apply more generally than just for the situations considered and described here. I have only explored a small section of a very large number of possible simple models, and even within those simple models, only specific parameter combinations have been considered. As the space of possible bipeds is explored more carefully and more exhaustively, it is likely to be found that some of the claims and conclusions here are insufficiently general and must be revised and subsumed under more general results. A more careful and exhaustive analysis of each of the four biped models in this article is indicated and might provide a foundation for the construction and understanding of more complex and more realistic models. Also, in this article, I mostly derive the optimal gaits using numerical optimization and they are sometimes buttressed by analytical reasoning where appropriate. It is likely that many of these optimization calculations can be understood a bit better by attempting proofs of optimality, perhaps in simplified cases such as in [29].
In summary, while no single model examined here has all the features that we would like of a predictive model of steady legged locomotion, it has been shown here that a large number of qualitative features of human bipedal locomotion can be explained by the hypothesis of energy optimality, even when applied to only relatively simple biped models. It seems likely that a model not much more complex than the ones presented here could be much more successful.
Acknowledgements
This work was supported partly by the Ohio State University and partly by NSF EF-0425878 FIBR grant, awarded to Philip Holmes at Princeton University. I thank P. Holmes and four anonymous reviewers for a close reading of and detailed comments on earlier versions of the manuscript. Andy Ruina first suggested the plots (figure 2c,d) and noted their similarity. He also independently suggested the representation of Cg. I thank A. Ruina and P. Holmes for numerous insightful discussions regarding all these topics, which have contributed immeasurably to this manuscript.
Appendix A. Minimizing F2 costs
Consider two actuators in parallel, exerting forces F1 and F2, respectively. Say that it is required that the two forces sum to F0. Then, F1 = F2 = F0/2 minimizes F12 + F22. Also, for a single actuator, if you want to produce an average force F while minimizing the integral of F2, it is best to have a constant force rather an oscillatory force.
The above observation is implicated in the following: especially for small step lengths when the leg forces are close to vertical, the CF2 optimal gait for the telescoping biped had a lot of double support; in fact, most of the stance phase tended to be double support. This is because a given force F over time T, produced by one leg roughly costs F2T and when produced by two legs costs 2 × (F/2)2 T = F2T/2. For higher speeds and step lengths, a running-like gait with flight phases and single-stance phases appears optimal (figure 8).
Appendix B. Minimizing Cg for a simple task resembling swinging a leg
Here I describe a simple task analogous to leg swing [30] for which it can be proved that minimizing Cg with a positive g(v) is identical to minimizing Cw.
Consider the mass m on a frictionless surface, whose horizontal position is denoted x (figure 20a). Say the task is to move the mass through a distance xmax in time tmax; in effect, an average speed of vavg = xmax/tmax is required. The mass must start at rest and end at rest (ẋ = 0). There is one actuator, able to apply forces F(t) of any magnitude.
Figure 20.

(a) The task is to move the mass from x = 0 to xmax in time tmax, starting and ending at rest. (b) The optimal strategy is OABC, OA is instantaneous, AB is constant speed and BC is instantaneous. The series of horizontal lines are the energy contours. A non-optimal path will necessarily have to intersect a higher energy contour than AB, thus requiring greater positive work.
What strategy minimizesCw for this task? I will show that the best strategy is to apply an impulse initially, so that the mass is immediately accelerated to a speed vavg = xmax/tmax, let the mass coast the distance xmax over time tmax, and then decelerate to zero speed instantaneously at time tmax. The positive work required for this bang-coast-bang strategy is mvavg2/2. A short outline of a proof of optimality follows. This claimed optimal strategy uses a constant forward velocity equal to the required average velocity vavg over essentially the whole time duration. An alternative strategy, with some finite duration spent at velocities less than vavg, will necessarily also have a duration in which the mass travels faster than vavg, to satisfy the average velocity constraint. This means that the mass has, at some point during its motion, an energy greater than mvavg2/2. This implies that the minimum required positive work (=∫ Fv dt with F > 0 and v > 0) for such an alternative strategy must be greater than mvavg2/2, proving the optimality of the impulsive strategy, which only requires mvavg2/2. Figure 20b shows the path OABC taken by the optimal strategy in the x − ẋ phase plane. See [30] and [29] for similar arguments in similar contexts.
What about minimizing Cg? It can be shown that exactly the same bang-coast-bang strategy, the path OABC of figure 20b, also minimizes Cg with a positive g(v) (where v = ẋ) for this task. The proof is the same as above, except now ‘energy’ in the above proof is replaced with the quantity G(v) = ∫0v g(v′) dv′, which is monotonically increasing with v when g(v) is positive. Note that G(v) equals the cost Cg when F > 0 and v > 0, and, therefore, has the same status that positive work has to Cw: Cg = ∫ Fg(v) dt = ∫v̇g(v) dt = ∫ g(v) dv. The constant mechanical energy contours in figure 20b now become constant G(v) contours, albeit with different spacing.
The similarity of the optimal strategies for the costs Cw and the more general Cg for this simple task foreshadows the similarity of the optimal walking and running strategies for the springless telescoping model. The sameness only requires that G(v) be strictly increasing, which is assured for positive g(v). Convexity of g(v), which was used in §6, was not required for the above proof. Also, in the above proofs, it was implicitly assumed that F could be positive or negative; it may be more physically plausible to have two muscles, one for acceleration (agonist) and another for deceleration (antagonist), but both the optimal strategy and the proof arguments would be essentially identical.
Appendix C. A gait-transition theory for the springless bipeds
In this section, I present simple expressions for the cost of the optimal gaits for the springless telescoping biped, minimizing a Cg-like cost—namely for inverted pendulum walking and impulsive running. Using these simple expressions, it is possible to determine when gait transitions should occur. The cost of transport is used to choose between different gaits; the cost of transport ct is the non-dimensional quantity defined as the metabolic cost per unit weight and distance travelled. For simplicity, the arguments below about gait transitions assume that the gait choice is only between inverted pendulum walking and impulsive running gaits, and show that transition between just these gaits should happen at roughly the same [V, D] boundaries for some different Cg's for small step lengths.
Impulses for inverted pendulum walking and impulsive running. Especially for Cw, all the cost is incurred at the impulses: the heel-strike and push-off impulses for pendular walking and running, and the single pseudo-elastic impulse for impulsive running. The pendular phases of these gaits have no cost, as these phases involve no work, and in the case of the kneed biped, requires no knee moment. So for completeness, I recall the impulse magnitudes for gaits inverted pendulum walking and impulsive running; see [30] and [4] for details.
For inverted pendular walking (and pendular running), the push-off and the heel-strike impulses are each equal to vc = v− tanθ, where v− is the velocity magnitude just before push-off, θ is the leg angle with vertical and 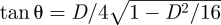 . The speed v− can be determined by numerical simulation or using approximations. A simple approximation, applicable for small D and not-too-small V is v− ≈ V, so that vc ≈ VD/4.
. The speed v− can be determined by numerical simulation or using approximations. A simple approximation, applicable for small D and not-too-small V is v− ≈ V, so that vc ≈ VD/4.
For impulsive running, the single vertical impulse can be broken up into a negative work impulse and a positive work impulse. The magnitude of each of these impulses is equal to the vertical velocity just before impact: vc = T/4 = D/4V. There are no approximations in this expression. Note again that in this paper, D stands for stride length, whereas in Srinivasan & Ruina [5] and Srinivasan [30], D stands for step length.
Work costCw. For both the springless telescoping biped and the kneed biped, it can be shown that the cost of transport ct for a positive work cost (Cw with b1 = 1 and b2 = 0) is given by:
| C 1 |
where vc is the non-dimensional magnitude of the push-off impulse [4,30]. Substituting the values for vc derived in the previous paragraphs, ct ≈ V2D/32 for inverted pendulum walking. For impulsive running, ct = D/32V2. If the choice were restricted to just between these two gaits, a comparison of these costs shows that walking is better for V < 1 and running for V > 1. This is a small-D approximation. The exact phase boundaries are in Srinivasan [30] and Srinivasan & Ruina [5]. The key observation here is that the cost expression (equation (C 1)) is a monotonic function of vc for a given D, and therefore, the gait transitions happen when the vc for one gait becomes larger than another. Such observations apply to the springless kneed biped as well.
Convex costCg. The Cg cost of the positive work impulse is given by ∫0vc Fg(v) dt = ∫0vc v̇g(v) dt = ∫0vcg(v) dv = G(vc) − G(0). The negative work impulse has a cost equal to G(0) − G(−vc). The cost of the pendular phase is equal to g(0)tpendular, where tpendular is the time spent in the pendular phase. The cost of transport will then be ct = [G(vc) − G(−vc) + g(0)tpendular]/D. If g(0) were zero, the cost ct would be a monotonic function of vc for a given D and, therefore, the gait transition boundaries will be exactly the same as in the earlier two cases. When g(0) is not equal to zero, the gait transition boundaries are likely to be a little different from the earlier cases. Note that g(0) for CAM is relatively close to zero (figure 2c).
Appendix D. Insensitivity of work minima to an additive CF2 cost
Consider the non-smooth function fλ(x) = |x| + λ(x − 1)2. For λ = 0, f(x) = |x| and is minimized at x = 0. Figure 21 shows fλ for a sequence of λ. The minimum remains exactly at x = 0 for a range of lambda around λ = 0 (a‘dead zone’) and then changes gradually from x = 0.
Figure 21.

Robustness of a non-smooth minimum. The function fλ(x) = |x| + λ(x − 1)2 is shown for a sequence of λ from 0 to 1. The minima of fλ(x) (shown as little circles) is remains exactly at x = 0 for λ = 0 to 0.5, and is greater than zero for λ > 0.5.
It can be shown that dead-zone behaviour for minimum around λ = 0 is generic for functions of the form fλ(x) = f1(x) + λf2(x) (x ∈ ℝn) when the f1(x) has a ‘non-smooth minimum’ at x = a such that the directional derivative of f1(x) along every feasible direction is positive (bounded away from zero). The proof is elementary: if the directional derivatives of f2(x) at x = a are bounded, the directional derivatives of fλ(x) at x = a remain positive for a range of λ around x = 0, thus making x = a remain a minimum for this range. The set of feasible directions is assumed independent of λ.
Arguments similar to these can presumably be used to show the robustness of optimality of the zero work strategies to small cost function perturbations of the form λ∫F2 dt for the springy bipeds. For a given speed, stride length and spring stiffness, there is only one (locally isolated) symmetric work-free gait. The work cost for this gait probably satisfies the conditions above: that any small perturbation to this gait, satisfying the speed and stride length constraints, will result in a small work cost whose size is the same order as the perturbation to the gait (say measured by changes in leg forces), thus showing the positivity of the directional derivatives in every direction, as required by the above proof of insensitivity.
Appendix E. Optimality of symmetric walking
Consider the asymmetric walking gait shown in figure 22a, in which the two steps that make up a stride are unequal. The two push-offs in a stride are accomplished with different leg orientations, namely with leg angles θ1 and θ2 as shown in the figure. For small leg angles, it can be shown that the positive work performed during the push-off scales like V2θ2/2 (see appendix C and [30]). Given θi ≈ Di/2, the total cost (positive work) over the two steps (one stride) would be V2(D12 + D22)/8. Given D1 + D2 = D, minimizing the cost for a given V and D is like minimizing D12 + D22, accomplished by setting D1 = D2 = D/2. The reasoning is identical to that in appendix A. Thus, symmetric inverted pendulum walking is best among all inverted pendulum walking gaits. This proof assumes small leg angles, but the claim is generally true.
Figure 22.

Idealized asymmetric gaits. (a) Asymmetric inverted pendulum walking, with two unequal step lengths in a stride. The leg length is constant during most of the stance phase, equal to the maximum possible leg length, except for infinitesimal extensions and contractions during push-off and heel-strike impulses. (b) Asymmetric impulsive running gait, with two unequal steps in a stride.
Similarly, it can be shown that symmetric impulsive running, with equally spaced vertical impulses, is better than the asymmetric impulsive running gait in figure 22b. Because the two costs (for symmetric and asymmetric impulsive running gaits) are the same up to leading order in D [4], the proof of optimality would be somewhat longer than the above proof for walking. But see [30] for a demonstration of the claim that a symmetric impulsive running gait is better than an asymmetric running gait in which the two impulses are close together, analogous to skipping.
Footnotes
Actually, the simplest model for electric motors would be a1F2 + a2P, the first term related to Joule heating losses and the second term allowing for regeneration of mechanical power. The power P gets integrated to zero when one considers periodic motions in the absence of physical dissipation, as is the case here, so the second term drops out of the optimizations here.
If the surface was frictionless, the biped could glide without performing work once motion is initiated somehow, say by pushing off a wall.
References
- 1.Daffertshofer A., Lamoth C. J. C., Meijer O. G., Beek P. J. 2004. PCA in studying coordination and variability: a tutorial. Clin. Biomech. 19, 415–428 10.1016/j.clinbiomech.2004.01.005 (doi:10.1016/j.clinbiomech.2004.01.005) [DOI] [PubMed] [Google Scholar]
- 2.Alexander R. McN. 1976. Mechanics of bipedal locomotion. Perspectives in experimental biology, vol. 1, pp. 493–504 New York, NY: Pergamon Press [Google Scholar]
- 3.Kuo A. D., Donelan J. M., Ruina A. 2005. Energetic consequences of walking like an inverted pendulum: step-to-step transitions. Exer. Sport Sci. Rev. 33, 88–97 10.1097/00003677-200504000-00006 (doi:10.1097/00003677-200504000-00006) [DOI] [PubMed] [Google Scholar]
- 4.Ruina A., Bertram J. E., Srinivasan M. 2005. A collisional model of the energetic cost of support work qualitatively explains leg-sequencing in walking and galloping, pseudo-elastic leg behavior in running and the walk-to-run transition. J. Theor. Biol. 14, 170–192 [DOI] [PubMed] [Google Scholar]
- 5.Srinivasan M., Ruina A. 2006. Computer optimization of a minimal biped model discovers walking and running. Nature 439, 72–75 10.1038/nature04113 (doi:10.1038/nature04113) [DOI] [PubMed] [Google Scholar]
- 6.Rashevsky N. 1948. On the locomotion of mammals. Bull. Math. Biophys. 10, 11–23 10.1007/BF02478326 (doi:10.1007/BF02478326) [DOI] [PubMed] [Google Scholar]
- 7.McMahon T. A., Cheng G. C. 1990. The mechanics of running: how does stiffness couple with speed? J. Biomech. 23(Suppl 1), 65–78 10.1016/0021-9290(90)90042-2 (doi:10.1016/0021-9290(90)90042-2) [DOI] [PubMed] [Google Scholar]
- 8.Farley C. T., Blickhan R., Saito J., Taylor C. R. 1991. Hopping frequency in humans: a test of how springs set stride frequency in bouncing gaits. J. Appl. Physiol. 71, 2127–2132 [DOI] [PubMed] [Google Scholar]
- 9.Blickhan R., Full R. J. 1993. Similarity in multilegged locomotion: bouncing like a monopode. J. Comp. Physiol. A. 173, 509–517 [Google Scholar]
- 10.Geyer H., Seyfarth A., Blickhan R. 2006. Compliant leg behaviour explains basic dynamics of walking and running. Proc. R. Soc. B 273, 2861–2867 10.1098/rspb.2006.3637 (doi:10.1098/rspb.2006.3637) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.O'Connor S. M., Kuo A. D. 2006. Dynanic biopedal locomotion on compliant legs. J. Biomech. 39(Suppl 1), S359 [Google Scholar]
- 12.Srinivasan M., Holmes P. 2008. How well can spring-mass-like telescoping leg models fit multi-pedal sagittal plane locomotion? J. Theor. Biol. 255, 1–7 10.1016/j.jtbi.2008.06.034 (doi:10.1016/j.jtbi.2008.06.034) [DOI] [PubMed] [Google Scholar]
- 13.Borelli J. A. 1680. On the movement of animals (De Motu Animalium, Pars prima) (Maquet P. transl. 1989), p. 152 Berlin, Germany: Springer-Verlag [Google Scholar]
- 14.Alexander R. McN. 1989. Optimization and gaits in the locomotion of vertebrates. Physiol. Rev. 69, 1199–1227 [DOI] [PubMed] [Google Scholar]
- 15.Alexander R. McN. 1996. Optima for animals. Princeton, NJ: Princeton University Press [Google Scholar]
- 16.Srinivasan M. 2009. Optimal speeds for walking and running, and walking on a moving walkway. Chaos 19, 026112. [DOI] [PubMed] [Google Scholar]
- 17.Chow C. K., Jacobson D. H. 1971. Studies of human locomotion via optimal programming. Math. Biosci. 10, 239–306 10.1016/0025-5564(71)90062-9 (doi:10.1016/0025-5564(71)90062-9) [DOI] [Google Scholar]
- 18.Hatze H. 1976. The complete optimization of the human motion. Math. Biosci. 28, 99–135 10.1016/0025-5564(76)90098-5 (doi:10.1016/0025-5564(76)90098-5) [DOI] [Google Scholar]
- 19.Beletskii V. V., Chudinov P. S. 1977. Parametric optimization in the problem of bipedla locomotion. Izv. An SSSR Mekhanika Tverdogo (Mech. Solids), pp. 25–35 [Google Scholar]
- 20.Alexander R. McN. 1980. Optimum walking techniques for quadrupeds and bipeds. J. Zool. Lond. 192, 97–117 10.1111/j.1469-7998.1980.tb04222.x (doi:10.1111/j.1469-7998.1980.tb04222.x) [DOI] [Google Scholar]
- 21.Alexander R. McN. 1992. A model of bipedal locomotion on compliant legs. Phil. Trans. R. Soc. Lond. B 338, 189–198 10.1098/rstb.1992.0138 (doi:10.1098/rstb.1992.0138) [DOI] [PubMed] [Google Scholar]
- 22.Davy D. T., Audu M. L. 1987. A dynamic optimization technique for predicting muscle forces in the swing phase of gait. J. Biomech. 20, 187–201 10.1016/0021-9290(87)90310-1 (doi:10.1016/0021-9290(87)90310-1) [DOI] [PubMed] [Google Scholar]
- 23.Minetti A. E., Alexander R. McN. 1997. A theory of metabolic costs for bipedal gaits. J. Theor. Biol 186, 467–476 10.1006/jtbi.1997.0407 (doi:10.1006/jtbi.1997.0407) [DOI] [PubMed] [Google Scholar]
- 24.Anderson F. C., Pandy M. G. 2001. Dynamic optimization of human walking. J. Biomech. Eng. 123, 381–390 10.1115/1.1392310 (doi:10.1115/1.1392310) [DOI] [PubMed] [Google Scholar]
- 25.Safonova A., Pollard N., Hodgins J. K. 2003. Optimizing human motion for the control of a humanoid robot. In Proc. Int. Conf. on Robotics and Automation, pp. 1390–1397 [Google Scholar]
- 26.Liu C. K., Hertzmann A., Popović Z. 2005. Learning physics-based motion style with nonlinear inverse optimization. ACM Trans. Graph. 24, 1071–1081 10.1145/1073204.1073314 (doi:10.1145/1073204.1073314) [DOI] [Google Scholar]
- 27.Ren L., Jones R., Howard D. 2007. Predictive modelling of human walking over a complete gait cycle. J. Biomech. 40, 1567–1574 10.1016/j.jbiomech.2006.07.017 (doi:10.1016/j.jbiomech.2006.07.017) [DOI] [PubMed] [Google Scholar]
- 28.Doke J., Donelan M. J., Kuo A. D. 2005. Mechanics and energetics of swinging the human leg. J. Exp. Biol. 208, 439–445 10.1242/jeb.01408 (doi:10.1242/jeb.01408) [DOI] [PubMed] [Google Scholar]
- 29.Srinivasan M., Ruina A. 2007. Idealized walking and running gaits minimize work. Proc. R. Soc. A 463, 2429–2446 10.1098/rspa.2007.0006 (doi:10.1098/rspa.2007.0006) [DOI] [Google Scholar]
- 30.Srinivasan M. 2006. Why walk and run: energetic costs and energetic optimality in simple mechanics-based models of a bipedal animal. PhD thesis, Cornell University, Ithaca [Google Scholar]
- 31.Cavagna G. A., Saibene F. P., Margaria R. 1964. Mechanical work in running. J. Appl. Physiol 19, 249–256 [DOI] [PubMed] [Google Scholar]
- 32.Alexander R. McN. 1988. Elastic mechanisms in animal movement. Cambridge, UK: Cambridge University Press [Google Scholar]
- 33.Alexander R. McN. 2003. Principles of animal locomotion. Princeton, NJ: Princeton University Press [Google Scholar]
- 34.Ralston H. J. 1958. Energy-speed relation and optimal speed during level walking. Eur. J. Appl. Physiol. (Int. Z. Angew. Physiol.) 17, 277–283 [DOI] [PubMed] [Google Scholar]
- 35.Hoyt D. F., Taylor C. R. 1981. Gait and the energetics of locomotion in horses. Nature 292, 239–240 10.1038/292239a0 (doi:10.1038/292239a0) [DOI] [Google Scholar]
- 36.Kuo A. D. 2001. A simple model predicts the step length-speed relationship in human walking. J. Biomech. Eng. 123, 264–269 10.1115/1.1372322 (doi:10.1115/1.1372322) [DOI] [PubMed] [Google Scholar]
- 37.Tucker V. A. 1975. The energetic cost of moving about. Am. Sci. 63, 413–419 [PubMed] [Google Scholar]
- 38.Radhakrishnan V. 1998. Locomotion: dealing with friction. Proc. Natl Acad. Sci. USA 95, 5448–5455 10.1073/pnas.95.10.5448 (doi:10.1073/pnas.95.10.5448) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Margaria R. 1976. Biomechanics and energetics of muscular exercise. Oxford, UK: Clarendon Press [Google Scholar]
- 40.Alexander R. McN. 2002. Work or force minimization as a criterion for the evolution of locomotion. In Fourth World Congress of Biomechanics, Calgary, Canada. [Google Scholar]
- 41.Alexander R. McN. 1997. A minimum energy cost hypothesis for human arm trajectories. Biol. Cybern. 76, 97–105 10.1007/s004220050324 (doi:10.1007/s004220050324) [DOI] [PubMed] [Google Scholar]
- 42.Alexander R. McN. 1997. Optimum muscle design for oscillatory movements. J. Theor. Biol. 184, 253–259 10.1006/jtbi.1996.0271 (doi:10.1006/jtbi.1996.0271) [DOI] [PubMed] [Google Scholar]
- 43.Woledge R. C., Curtin N. A., Homsher E. 1985. Energetic aspects of muscle contraction. London, UK: Academic Press; [PubMed] [Google Scholar]
- 44.Ma S. P., Zahalak G. I. 1991. A distribution-moment model of energetics in skeletal muscle. J. Biomech. 24, 21–35 10.1016/0021-9290(91)90323-F (doi:10.1016/0021-9290(91)90323-F) [DOI] [PubMed] [Google Scholar]
- 45.Crowninshield R. D. 1978. Use of optimization techniques to predict muscle forces. J. Biomech. Eng. 100, 88–92 [Google Scholar]
- 46.Crowninshield R. D., Brand R. A. 1981. A physiologically based criterion of muscle force prediction in locomotion. J. Biomech. 14, 793–801 10.1016/0021-9290(81)90035-X (doi:10.1016/0021-9290(81)90035-X) [DOI] [PubMed] [Google Scholar]
- 47.Fenn W. O. 1924. The relation between the work performed and the energy liberated in muscular contraction. J. Physiol. 58, 373–395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Roussel L., Canudas-De-Wit C., Goswami A. 1998. Generation of energy optimal complete gait cycles for biped robots. In Proc. 1998, IEEE International Conference on Robotics and Automation, vol. 3, pp. 2036–2041 [Google Scholar]
- 49.Chevallereau C., Aoustin Y. 2001. Optimal reference trajectories for walking and running of a biped robot. Robotica 19, 557–569 [Google Scholar]
- 50.Bhounsule P. 2008. Cornell ranger: computer simulation and experimental fits. Dynamic walking. Delft, The Netherlands: Holland [Google Scholar]
- 51.Bhounsule P., Ruina A. 2009. Cornell ranger: energy-optimal control. In Dynamic walking. Vancouver, Canada. [Google Scholar]
- 52.Rudin W. 1987. Real and complex analysis. New York, NY: McGraw-Hill [Google Scholar]
- 53.Alexander R. McN. 1997. Invited editorial on ‘interaction of leg stiffness and surface stiffness during human hopping’. J. Appl. Physiol. 82, 13–14 [DOI] [PubMed] [Google Scholar]
- 54.Blickhan R. 1989. The spring-mass model for running and hopping. J. Biomech. 22, 1217–1227 10.1016/0021-9290(89)90224-8 (doi:10.1016/0021-9290(89)90224-8) [DOI] [PubMed] [Google Scholar]
- 55.Ghigliazza R. M., Altendorfer R., Holmes P., Koditschek D. 2003. A simply stabilized running model. SIAM J. Appl. Dyn. Syst. 2, 187–218 10.1137/S1111111102408311 (doi:10.1137/S1111111102408311) [DOI] [Google Scholar]
- 56.Chatterjee A., Garcia M. 2000. Small slope implies low speed for Mcgeer's passive walking machines. Dyn. Stab. Syst. 15, 139–157 [Google Scholar]
- 57.Garcia M., Chatterjee A., Ruina A. 2000. Efficiency, speed, and scaling of two-dimensional passive-dynamic walking. Dyn. Stab. Syst. 15, 75–99 [Google Scholar]
- 58.Donelan J. M., Kram R., Kuo A. D. 2002. Mechanical work for step-to-step transitions is a major determinant of the metabolic cost of human walking. J. Exp. Biol. 205, 3717–3727 [DOI] [PubMed] [Google Scholar]
- 59.Collins S. H., Ruina A., Tedrake R., Wisse M. 2005. Efficient bipedal robots based on passive dynamic walkers. Science 307, 1082–1085 10.1126/science.1107799 (doi:10.1126/science.1107799) [DOI] [PubMed] [Google Scholar]
- 60.Gomes M., Ruina A. In preparation A walking model with no energy cost. [DOI] [PubMed] [Google Scholar]
- 61.McGeer T. 1989. Powered flight, child's play, silly wheels and walking machines. In Proc. IEEE Int. Conf. Robotics and Automation, vol. 3, pp. 1592–1597 10.1109/ROBOT.1989.100205 (doi:10.1109/ROBOT.1989.100205) [DOI] [Google Scholar]
- 62.Saunders J. B., Inman V. T., Eberhart H. D. 1953. The major determinants in normal and pathological gait. J. Bone Joint Surg. 35, 543–558 [PubMed] [Google Scholar]
- 63.Athans M., Falb P. L. 1969. Optimal control. New York, NY: McGraw Hill [Google Scholar]
- 64.Usherwood J. R. 2005. Why not walk faster? Biol. Lett. 1, 338–341 10.1098/rsbl.2005.0312 (doi:10.1098/rsbl.2005.0312) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ortega J. D., Farley C. T. 2005. Minimizing center of mass vertical movement increases metabolic cost in walking. J. Appl. Physiol. 99, 2099–2107 10.1152/japplphysiol.00103.2005 (doi:10.1152/japplphysiol.00103.2005) [DOI] [PubMed] [Google Scholar]
- 66.Gordon K. E., Ferris D. P., Kuo A. D. 2009. Metabolic and mechanical energy costs of reducing vertical center of mass movement during gait. Arch. Phys. Med. Rehabil. 90, 136–144 10.1016/j.apmr.2008.07.014 (doi:10.1016/j.apmr.2008.07.014) [DOI] [PubMed] [Google Scholar]
- 67.Alexander R. McN. 1991. Energy-saving mechanisms in walking and running. J. Exp. Biol. 160, 55–69 [DOI] [PubMed] [Google Scholar]
- 68.Roberts T. J., Marsh R. L., Weyand P. G., Taylor C. R. 1997. Muscular force in running turkeys: the economy of minimizing work. Science 275, 1113–1115 10.1126/science.275.5303.1113 (doi:10.1126/science.275.5303.1113) [DOI] [PubMed] [Google Scholar]
- 69.Fukunaga T., Kubo K., Kawakami Y., Fukashiro S., Kanehisa H., Maganaris C. N. 2001. In vivo behavior of human muscle tendon during walking. Proc. R. Soc. Lond. B 268, 229–233 10.1098/rspb.2000.1361 (doi:10.1098/rspb.2000.1361) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cavagna G. A., Heglund N. C., Taylor C. R. 1977. Mechanical work in terrestrial locomotion: two basic mechanisms for minimizing energy expenditure. Am. J. Physiol. 233, 243–261 [DOI] [PubMed] [Google Scholar]
- 71.Alexander R. McN. 1984. Elastic energy stores in running vertebrates. Am. Zool. 24, 85–94 [Google Scholar]
- 72.Biewener A. A. 1998. Muscle-tendon stresses and elastic energy storage during locomotion in the horse. Comp. Biochem. Physiol. B 120, 73–87 10.1016/S0305-0491(98)00024-8 (doi:10.1016/S0305-0491(98)00024-8) [DOI] [PubMed] [Google Scholar]
- 73.Beletskii V. V., Berbyuk V. E., Samsonov V. A. 1982. Parametric optimization of motions of a bipedal walking robot. Izv. An SSSR Mekhanika Tverdogo (Mech. Solids) 17, 28–40 [Google Scholar]
- 74.Kram R., Taylor C. R. 1990. Energetics of running: a new perspective. Nature 346, 265–267 10.1038/346265a0 (doi:10.1038/346265a0) [DOI] [PubMed] [Google Scholar]
- 75.Pontzer H. 2005. A new model predicting locomotor cost from limb length via force production. J. Exp. Biol. 208, 1513–1524 10.1242/jeb.01549 (doi:10.1242/jeb.01549) [DOI] [PubMed] [Google Scholar]
- 76.Marsh R. L., Ellerby D. J., Carr J. A., Henry H. T., Buchanan C. I. 2004. Partitioning the energetics of walking and running: swinging the legs is expensive. Science 303, 80–83 10.1126/science.1090704 (doi:10.1126/science.1090704) [DOI] [PubMed] [Google Scholar]