

Abstract
Understanding how vegetation growth responds to climate change is a critical requirement for projecting future ecosystem dynamics. Parts of North America (NA) have experienced a spring cooling trend over the last three decades, but little is known about the response of vegetation growth to this change. Using observed climate data and satellite-derived Normalized Difference Vegetation Index (NDVI) data from 1982 to 2006, we investigated changes in spring (April–May) temperature trends and their impact on vegetation growth in NA. A piecewise linear regression approach shows that the trend in spring temperature is not continuous through the 25-year period. In the northwestern region of NA, spring temperature increased until the late 1980s or early 1990s, and stalled or decreased afterwards. In response, a spring vegetation greening trend, which was evident in this region during the 1980s, stalled or reversed recently. Conversely, an opposite phenomenon occurred in the northeastern region of NA due to different spring temperature trends. Additionally, the trends of summer vegetation growth vary between the periods before and after the turning point (TP) of spring temperature trends. This change cannot be fully explained by summer drought stress change alone and is partly explained by changes in the trends of spring temperature as well as those of summer temperature. As reported in previous studies, summer vegetation browning trends have occurred in the northwestern region of NA since the early 1990s, which is consistent with the spring and summer cooling trends in this region during this period.
Keywords: climate change, spring cooling
Vegetation plays an important role in regulating climate through the exchange of energy, water vapor, and momentum between the land surface and the atmosphere (1, 2). Vegetation also controls atmospheric CO2 and currently absorbs about one-third of anthropogenic fossil fuel emissions to the atmosphere. The magnitude of the gross flux of carbon taken up annually by photosynthesis is about 15 times the absolute value of fossil fuel emissions. Thus, even a small change in vegetation photosynthesis could have a large effect on the role of vegetation as a carbon sink (3). Long-term satellite observations from National Oceanic and Atmospheric Administration (NOAA) Advanced Very High Resolution Radiometer (AVHRR) have shown that vegetation greenness in the Northern Hemisphere (NH) significantly increased between the early 1980s and the late 1990s (4, 5). This greening has been interpreted using terrestrial biosphere models as increased photosynthesis and growth (6, 7). Thus, the observation that the NH vegetation has become greener helps to account for the increase of NH carbon sink during the 1980s and 1990s (ref. 8, pp. 511–538; 9). Intriguingly, this greening trend seems to have stalled, or even reversed in some regions, during the current decade (e.g., 10–13). Understanding the mechanisms behind this phenomenon is important for projections of future vegetation growth and feedbacks to climate change.
Vegetation growth in NH mid to high latitudes is very sensitive to temperature changes, particularly in spring (14, 15). Spring temperature has risen by 1.1 °C since the 1980s (16). Warmer spring temperature will generally enhance vegetation productivity by extending the growing season. Rather than responding to changes in continental mean temperature, vegetation growth is evidently more sensitive to spatially heterogeneous changes in local temperature (17). In fact, not a single region exactly follows the change in continental mean temperature (18). For instance, despite a remarkable overall warming in spring at the continental scale, a trend toward cooler spring conditions has occurred from 1979 to 2005 in a few regions of North America (NA). Such a spring cooling should exert negative impacts on regional vegetation growth and could possibly contribute to the recent pause of greening observed in some parts of the NH (e.g., 10). Because warming dominates on the NH scale, however, little attention has been paid to regions that experienced spring cooling and to the impact of cooling on local vegetation photosynthesis.
The objective of this study is to investigate regional-scale spring temperature changes and their impacts on vegetation growth over NA. Using observed temperature data from 1982 to 2006 (19), we characterize spatio-temporal changes of spring temperature trends over NA (north of 25 °N) based on a piecewise linear regression algorithm, which detects the potential turning point (TP) at which the trend changes at each point in space. We next assess the temperature trends for each location on both sides of each turning point. Then we analyzed how distinct regional spring temperature trends have influenced the trends of vegetation greenness index during the period 1982–2006. We infer vegetation growth from satellite-derived Normalized Difference Vegetation Index (NDVI) (20).
Results and Discussion
Spring Temperature Trends.
Fig. 1A provides the spatial pattern of spring temperature trends, based on Climatic Research Unit (CRU) data, during the period 1982–2006 (see Methods and Datasets). The majority of NA experienced a positive trend of spring temperature during 1982–2006, the average rate being of 0.047 ± 0.050 °C yr-1. However, some regions experienced a decrease in spring temperature (16% of the continent area). These cooling regions are mainly a west-to-east band at around 50 °N and southeastern Canada (Quebec). This result is consistent with findings of the IPCC fourth assessment report (ref. 8, pp. 247–258).
Fig. 1.

Spatial distribution of spring (April and May) temperature changes in North America. (A) Spring temperature trend during the whole study period 1982–2006, (B) the year corresponding to the turning point (TP) of spring temperature trend estimated by the piecewise linear regression approach, (C) spring temperature trend before the TP of spring temperature trend, and (D) spring temperature trend after the TP of spring temperature trend. White grids in B show area with insignificant (P > 0.10) TP. The insets show the frequency distributions of corresponding trends. The explanation (R2) of piecewise linear regression on the change in spring temperature is significant (P < 0.10) in 43% of the continent with the average R2 of 19%, which is about two times of average explanation (8%) derived from simple linear regression (26% of the area show significant explanation).
Further analyses on spring temperature data from CRU (Fig. 1 B–D) show, however, that the trend of spring temperature is not temporally homogeneous during the entire 25 years. Using the piecewise linear regression and t-test (see Methods and Datasets), we infer that the trend in spring temperature changed significantly (P < 0.10) in more than 40% of the entire study region, particularly north of 40 °N, and that the TP year of spring temperature trend varied dramatically across the continent (Fig. 1B). For example, spring temperature in the northwestern region of NA increased until the early 1990s (Fig. 1 B and C) and then decreased afterwards (Fig. 1D). In contrast, the northeast region of the continent experienced a decreasing trend of spring temperature before the TP of spring temperature trend (Fig. 1C) and an increasing trend of spring temperature more recently (Fig. 1D). Overall, spring temperature in about 76% of the North American territory (about 48% with a significant increase) increased before the TP (Fig. 1C), and the magnitude of this earlier warming trend was much larger than the trend during the whole study period (Fig. 1A). This result implies that rising spring temperature is stalled or reversed, cancelling out spring warming in an earlier period across most of this region (except in the northeast). Indeed, more than half of the continent (about 5% with a significant decrease) experienced a spring cooling trend after the TP, especially in the northwest, such as most parts of Alaska, Yukon, and British Columbia, where spring temperature decreased by > 0.05 °C yr-1 (Fig. 1D).
We applied the same regression analyses (linear regression and piecewise approach) to the three other gridded temperature datasets. The patterns of 25 years’ spring temperature trend are similar across these datasets (Fig. S1 A–D). An overall warming trend but some cooling in the area between 40 °N to 60 °N was observed, despite some differences in whether decreasing temperature is more evident in the east or in the west, particularly between the two National Centers for Environmental Prediction (NCEP) datasets (see Methods and Datasets). The piecewise regression detects similar patterns of changing trends of spring temperature (Fig. S1 E–P) in the areas we discussed with evident spring temperature changes, particularly in the northwest, west, and northeast of the continent, except for one dataset [dataset of NCEP/DOE Reanalysis 2 (NCEP-DOE)], which showed different patterns in western and northern Canada. These results suggest the changing trend of spring temperature is not likely to be artifacts of data processing by CRU. The CRU dataset has advantages in several aspects: First, it is derived from instrumental records instead of modeling; second, it covers the entire study period without years missing; third, it has finer spatial resolution than other observed temperature data. Considering the similar results regarding trends and TP among the datasets and the advantages of the CRU dataset, we will use the CRU temperature data in the following analyses.
Spring NDVI Trends.
Because vegetation growth in high latitude regions is limited primarily by temperature (5), a change in the spring temperature trend should influence vegetation growth in these regions. Fig. 2A shows spatial distribution of the spring NDVI trends calculated over the 1982–2006 period. Over that period, the spring NDVI trend is relatively small and fragmented across NA (Fig. 2A), with an average value of (0.17 ± 1.7) × 10-3 yr-1. Regions where the absolute value of spring NDVI trend is found to be larger than 2 × 10-3 yr-1 (< -2 × 10-3 yr-1 or > 2 × 10-3 yr-1) are limited to only 22% of the total area (Fig. 2A).
Fig. 2.

Spatial distribution of spring (April and May) NDVI change during different periods according to TP of spring temperature. (A) Spring NDVI trend during the whole study period 1982–2006, (B) spring NDVI trend before the TP of spring temperature trend, and (C) spring NDVI trend after the TP of spring temperature trend. The insets show the frequency distributions of corresponding trends.
The reason spring NDVI has changed so little in many regions over the entire observation period can be explained by calculating the trends before and after the TP year of spring temperature change (Fig. 2 B and C). Most regions (68% and 15% with a significant increase) show a pronounced NDVI greening (increasing) trend before the TP of spring temperature trend, particularly in the northwestern region of NA where the increasing trend of spring NDVI is larger than 5 × 10-3 yr-1 (Fig. 2B). In comparison, over the northeastern region—Quebec and Newfoundland and Labrador—where cooling or small changes of spring temperature were observed before the TP (Fig. 1C), a decrease or stalling of spring NDVI is observed. After the TP of spring temperature, the trends of spring NDVI over these regions also reversed (Fig. 2C), almost exactly mirroring the spring temperature trends. Specifically, the northeastern region of North America (Quebec and Newfoundland and Labrador, see Fig. 2 B and C) has greened after the TP year, whereas the larger northwestern part of the continent experienced browning (decreasing NDVI). This dipole of NDVI trends is remarkably coherent with the spring temperature trends in these two regions (Fig. 1 C and D). Similar patterns of the trend in the average NDVI during May and June were also observed (Fig. S2), further confirming that the vegetation growth in the early growing season has stalled or reversed recently in response to changes in spring temperature trends.
For the entire period 1982–2006, the spring NDVI change is less well connected with temperature changes (Figs. 1A and 2A). However, if we compare instead temperature and NDVI trend patterns before and after the TP of spring temperature (Figs. 1 C and D and 2 B and C), then the spatial pattern of the spring NDVI trends clearly matches that of the spring temperature trends (Figs. 1 C and D and 2 B and C). The match of patterns between the trends of both winter and spring precipitation and those of spring NDVI (Figs. S3 A and B and S4 A and B) and the match between trends of the spring Palmer Drought Severity Index (PDSI) and those of spring NDVI (Fig. S4 C and D) seem to be much weaker.
To further evaluate the linkage between spring NDVI trends and spring temperature trends, we perform a similar analysis but starting from the NDVI dataset. We calculate spring NDVI and spring temperature trends on each side of the TP estimated from a piecewise regression of the spring NDVI (Fig. 3A). Doing this, we still see remarkably similar spatial pattern between spring temperature and spring NDVI trends before and after the TP defined from spring NDVI time series. Furthermore, the spatial distribution of trends of spring temperature and spring NDVI before and after the TP of spring NDVI are similar to those calculated before and after TP of spring temperature. A large area (68% and 31% with a significant increase), mainly in Alaska, western Canada, and around the Great Lakes, greened before the TP of spring NDVI. At the same time, spring warming dominated the continent (82% and 24% with a significant increase) and most evidently (> 0.1 °C/yr) in those above-mentioned regions with spring greening. After the TP of spring NDVI, both browning and cooling are observed, particularly in the above-mentioned regions. Such negative effects of spring cooling on vegetation growth of northern ecosystems may be not only associated with the delayed vegetation greening (21–23) but also related to the increasing risk of spring frost or extreme low temperature (24–26). It has been suggested that spring frost may severely damage vegetation growth, particularly when it took place after budburst (24). However, because freezing effects are sensitive to high-frequency low-temperature extremes rather than monthly means, we have too little knowledge about the potential contribution of historical change in spring frost to the vegetation growth across NA due to the lack of spatially and temporally explicit frost data, and further studies are needed.
Fig. 3.

Spatial distribution of spring (April and May) NDVI change and temperature change during different periods in North America according to TP of spring NDVI. (A) The year corresponding to the turning point (TP) of spring NDVI trend estimated by the piecewise linear regression approach (B) spring NDVI trend before the TP of spring NDVI trend, (C) spring NDVI trend after the TP of spring NDVI trend, (D) spring temperature trend before the TP of spring NDVI trend, and (E) spring temperature trend after the TP of spring NDVI trend. The insets show the frequency distributions of corresponding trends. White grids in A show area with insignificant (P > 0.10) TP.
These remarkable spatial correlations of trends suggest that temperature is a dominant factor governing the trend of spring vegetation growth (6, 17, 21, 27). Also, a strong positive correlation between spring NDVI and spring temperature has been observed in the mid to high latitudes of the NH (56% of NA with a significant positive correlation) (Fig. S5A) (28, 29). Furthermore, this correlation is stronger than the correlation of spring NDVI with both winter and spring precipitation (Figs. S3C and S5B), which show negative correlation across most areas, implying that spring vegetation growth of northern ecosystems is probably not limited by water supply. Thus the recent shift of spring NDVI trend in mid to high latitudes of NA seems to be driven by recent change of spring temperature trend.
Summer NDVI Trends.
Results from observed atmospheric CO2 concentrations (30, 31), eddy-covariance carbon flux measurements (22, 23), remote sensing data (21, 32), and ecosystem models (33, 34) have all indicated that the lengthening of growing-season duration, driven by rising temperature, can enhance vegetation productivity of northern terrestrial ecosystems. Forexample, Myneni et al. (21) suggested that an extended growing season, particularly an earlier beginning, leads to an increase in annual maximum vegetation activity. In order to explore if changes in spring vegetation growth, driven by changes in spring temperature, have some influences on summer NDVI, we analyzed summer NDVI trends on each side of the spring temperature TP year (Fig. 4 A and B). Although previous studies pointed to an overall greening trend of North America from 1982 to 1999 (4, 35), our analyses show that most regions of North America, particularly the high latitude regions, have experienced a decrease in summer NDVI during the entire period 1982–2006 (Fig. S6). Some of this decrease can be explained by the negative trend of summer NDVI after the TP of spring temperature trend (Fig. 4B). As shown in Fig. 4A, summer NDVI over 63% of the continent shows an increasing trend before the TP of spring temperature trend. In contrast, although the direct physiological effects of rising atmospheric CO2 are expected to cause a uniform increase of vegetation greenness (7, 36, 37), a decreasing trend of summer NDVI after the TP of spring temperature trend has occurred over most of North America (67% and 28% with a significant decrease) except over northern Alaska and southern Saskatchewan (Fig. 4B).
Fig. 4.

Spatial distribution of summer (June to August) NDVI change and climate change during different periods in North America. (A) Summer NDVI trend before the TP of spring temperature trend, (B) summer NDVI trend after the TP of spring temperature trend, (C) summer temperature trend before the TP of spring temperature trend, (D) summer temperature trend after the TP of spring temperature change, (E) summer precipitation trend before the TP of spring temperature trend, (F) summer precipitation trend after the TP of spring temperature trend, (G) summer PDSI trend before TP of spring temperature, and (H) summer PDSI trend after TP of spring temperature. The insets show the frequency distributions of corresponding trends.
Several mechanisms can contribute to the recent summer vegetation browning in NA. Currently, the most widely accepted explanation is the increasing drought stress, driven by rising summer temperature (10) or decreasing precipitation (38). However, drought may not be the only climate driver of the summer browning. As Fig. 4 F and H shows, there are some regions, such as east Alaska and Yukon, that experienced an ease of drought stress implied by summer PDSI increase and summer precipitation increase but still showed summer browning after the TP of spring temperature. In fact, summer precipitation over the entire period does not support the idea of a general drying trend in regions such as the southeastern United States, Alaska, and most of Canada, though the rate varies spatially. The precipitation change in different periods (Fig. 4 E and F) further suggests that decreasing precipitation was more evident in the earlier period than the recent period. Thus, we speculate that change in drought stress may not be the only driving factor causing recent summer vegetation browning in North America and that recent spring cooling contributes to the decrease of summer NDVI.
Because vegetation metabolism is temperature-limited, vegetation growth in most boreal ecosystems is primarily sensitive to change in temperature (5–7, 39). For example, Lucht et al. (6) found that the warming signal alone largely explained the vegetation greening trend in the boreal region from 1982 to 1998. As shown in Figs. 1 and 4, the increasing temperature trends in both spring and summer before the TP of spring temperature trend were stalled or reversed after the TP of spring temperature trend over the high latitude regions (except for the summer temperature trend in the northwestern United States and southwest Alaska). This halt in spring and summer warming is spatially consistent with the change of summer NDVI trend between the two periods (Fig. 4 A and B), implying the important role of temperature change in controlling vegetation growth in these regions.
In addition to climate change, fire has been recognized as a major disturbance to vegetation dynamics in burned area of boreal NA (40, 41). But the widespread recent decreasing trend in NDVI in Alaska and western Canada is unlikely to be mainly due to fire because burned area is only a small proportion of this region (42, 43), though fire events are having increasing frequency (44). And the decreasing NDVI is still observed in the unburned area (40, 42). Besides climate and fire, there are some other factors that may influence vegetation growth, including insect and disease damage (45, 46) as well as anthropogenic activities such as land management, afforestation, and deforestation. For instance, the current increasing trend of summer NDVI in southern Saskatchewan and the Dakotas may be partly related to irrigation practices (47).
3. Conclusions
The results presented in this study not only suggest that vegetation growth in mid to high latitudes of NA is very sensitive to temperature change but also imply that in addition to increasing drought stress proposed by the previous study of Angert et al. (10), both spring and summer cooling could contribute to summer browning trend occurred recently in NA. Separating the relative roles of these climate factors on the current vegetation growth change is a challenge for accurately predicting potential vegetation growth in response to changing future climate regimes. In addition, the implication of the current temperature change observed in this study for the net carbon balance of NA remains unknown. Besides vegetation productivity, carbon release from organic matter decomposition is also enhanced by the rising temperature, and thus the net carbon exchange between terrestrial ecosystems and the atmosphere does not necessarily benefit from a warming trend (16). Current terrestrial ecosystem modeling estimated a remarkably decreasing trend in net ecosystem productivity (NEP) in NA over the last three decades (48), despite the simulated positive effects of rising atmospheric CO2 (7). Further studies need to investigate the linkage between current temperature change and the net carbon balance over NA.
Methods and Datasets
Datasets.
NDVI, defined as the ratio of the difference between near-infrared reflectance and red visible reflectance to their sum, is a remote-sensed vegetation index widely used to measure vegetation greenness (21). Here, we use the biweekly NDVI data with an 8-km spatial resolution covering the period from 1982 to 2006 from Global Inventory Monitoring and Modeling Studies (GIMMS) group (20). The data are derived from NOAA AVHRR imagery using the NOAA satellite series 7, 9, 11, 12, and 16. This version of the GIMMS NDVI dataset is corrected through a series of processing steps to alleviate known limitations of the AVHRR measurements induced by intersensor calibration, orbital drift, cloud cover, solar angle differences, volcanic eruptions, and other atmospheric contaminations (4, 49). The excellent spatial coverage and relatively long-term observations by this NDVI dataset enable trend analyses. Earlier versions of this NDVI dataset have been widely used in detecting vegetation growth change (e.g., 4, 5, 10, 21).
The monthly temperature and precipitation dataset used in this study, the CRU TS 3.0 climate dataset obtained from the Climatic Research Unit (CRU) at the University of East Anglia (19), spans the twentieth century and hence covers our study period (1982–2006). This gridded dataset, with a spatial resolution of 0.5° × 0.5°, was based on climate observations from more than 4,000 meteorological stations, with a particularly dense coverage in North America and West Europe. In order to assess whether the trends of spring temperature are robust across different datasets, we also used three other air temperature datasets: the Goddard Institute for Space Science (GISS) dataset (18), the North America Regional Reanalysis dataset by the National Centers for Environmental Prediction (NCEP NARR) (50), and the NCEP/DOE Reanalysis 2 (NCEP-DOE) dataset (51), respectively. The GISS temperature dataset (18) was also based on instrumental data for the land area, with a spatial resolution of 2° × 2° covering the period of 1982 to 2005. NCEP NARR data (50) is a state-of-art model reanalysis providing air temperature data at high spatial resolution (32 km, i.e., ∼0.25°) across North America. NCEP-DOE Reanalysis 2 (51) is an alternative global model reanalysis including air temperature data at the resolution about 2°. The NCEP NARR and NCEP-DOE reanalyses span about 30 yr and hence cover the 1982 to 2006 study period.
The Palmer Drought Severity Index (PDSI) is one of the most used drought indices (52). It is based upon a two-layer soil model to calculate the demand and supply of soil water based on climate variables. Though comparison of PDSI between different locations should be treated with caution (53), PDSI still serves well to indicate temporal variations of drought (e.g., 13, 54). The PDSI dataset we used was produced by Dai et al. (55), who calculate monthly PDSI with a spatial resolution of 2.5° × 2.5°. This dataset has been updated to 2005, but data for 2006 are not yet available. In addition, PDSI data during the period of 2003 to 2005 are not available in some parts of high latitude regions (e.g., in part of Alaska and Yukon Territory).
Analysis.
In contrast to earlier observations of a significant spring warming in North America before the 1990s (56), only a small spring temperature change is observed during the period of 1980 to 2002 (16). This observation not only indicates that the spring temperature trend is different among different periods, but it also implies that spring warming has slowed down in some regions of NA. Because the primary object in this study is to detect if there is a TP in spring temperature trend as well as its implications in the change of vegetation growth, a piecewise linear regression approach with one TP (Eq. 1) (57) was applied to spring temperature time series for each grid point across NA from 1982 to 2006.
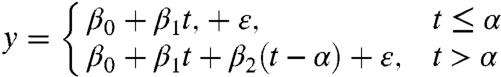 |
[1] |
where t is year; y is spring (April and May) temperature; α is the TP of spring temperature trend; and β0, β1, and β2 are regression coefficients. ε is the residual of the fit. The spring linear temperature trend is β1 before the turning point, β1 + β2 after it (Fig. S7). α is determined by least square error methods. We also confined α to within the period 1986 to 2002 in order to avoid linear regression in one period with too few data points. To evaluate the necessity of introducing TP, a t-test was applied to test the null hypothesis “β2 is not different from zero.” Similar analyses have also been applied independently to the spring NDVI. A P value < 10% was considered significant.
In order to describe the response of vegetation growth to changes in spring temperature trends, we analyzed spring and summer NDVI trends before and after the TP of spring temperature trend. Because NDVI has finer spatial resolution than climate data, TP of the nearest temperature grid cell was used to separate the two periods. Because the maximum value composite (MVC) NDVI (a maximum daily NDVI value during the 15-day period) minimizes atmospheric effects and cloud contamination effects (58), we used the larger 15-day MVC NDVI for a month to produce monthly NDVI datasets. Like in a previous study (5), spring is here defined as the period of April and May, and summer is defined as the period from June to August. To reduce the impact of bare and sparsely vegetated pixels on the NDVI trend, all pixels with growing-season average NDVI (April to October) smaller than 0.05 over the 25 yr were excluded from the analysis, similarly to Slayback et al. (49).
Supplementary Material
Acknowledgments.
This study was supported by the National Natural Science Foundation of China (grants 30970511 and 82641955) and the European Community’s Seventh Framework Programme (FP7/2007-2013) under grant agreement 24316.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1014425108/-/DCSupplemental.
References
- 1.Pielke R, et al. Interactions between the atmosphere and terrestrial ecosystems: influence on weather and climate. Glob Change Biol. 1998;4:461–475. [Google Scholar]
- 2.Wang W, et al. Feedbacks of vegetation on summertime climate variability over the North American grasslands. Part I: Statistical analysis. Earth Interact. 2006;10 (Paper 17) [Google Scholar]
- 3.Luyssaert S, et al. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob Change Biol. 2007;13:2509–2537. [Google Scholar]
- 4.Zhou L, et al. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J Geophys Res-Atmos. 2001;106:20069–20083. [Google Scholar]
- 5.Nemani R, et al. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science. 2003;300:1560–1563. doi: 10.1126/science.1082750. [DOI] [PubMed] [Google Scholar]
- 6.Lucht W, et al. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science. 2002;296:1687–1689. doi: 10.1126/science.1071828. [DOI] [PubMed] [Google Scholar]
- 7.Piao S, Friedlingstein P, Ciais P, Zhou L, Chen A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys Res Lett. 2006;33:L23402. 10.1029/2006GL028205. [Google Scholar]
- 8.Solomon SD, et al., editors. IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK, and New York: Cambridge Univ Press; 2007. [Google Scholar]
- 9.Piao S, et al. Spatiotemporal patterns of terrestrial carbon cycle during the 20th century. Global Biogeochem Cy. 2009;23:GB4026. 10.1029/2008GB003339. [Google Scholar]
- 10.Angert A, et al. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proc Natl Acad Sci USA. 2005;102:10823–10827. doi: 10.1073/pnas.0501647102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lotsch A, Friedl MA, Anderson BT, Tucker CJ. Response of terrestrial ecosystems to recent Northern Hemispheric drought. Geophys Res Lett. 2005;32:L06705. 10.1029/2004GL022043. [Google Scholar]
- 12.Park HS, Sohn BJ. Recent trends in changes of vegetation over East Asia coupled with temperature and rainfall variations. J Geophys Res. 2010;115:D14101. 10.1029/2009JD012752. [Google Scholar]
- 13.Zhao M, Running SW. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science. 2010;329:940–943. doi: 10.1126/science.1192666. [DOI] [PubMed] [Google Scholar]
- 14.Menzel A, et al. European phenological response to climate change matches the warming pattern. Glob Change Biol. 2006;12:1969–1976. [Google Scholar]
- 15.Schwartz MD, Ahas R, Aasa A. Onset of spring starting earlier across the Northern Hemisphere. Glob Change Biol. 2006;12:343–351. [Google Scholar]
- 16.Piao SL, et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature. 2008;451:49–53. doi: 10.1038/nature06444. [DOI] [PubMed] [Google Scholar]
- 17.Walther GR, et al. Ecological responses to recent climate change. Nature. 2002;416:389–395. doi: 10.1038/416389a. [DOI] [PubMed] [Google Scholar]
- 18.Hansen J, et al. Global temperature change. Proc Natl Acad Sci USA. 2006;103:14288–14293. doi: 10.1073/pnas.0606291103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mitchell TD, Jones PD. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int J Climatol. 2005;25:693–712. [Google Scholar]
- 20.Tucker CJ, et al. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int J Remote Sens. 2005;26:4485–4498. [Google Scholar]
- 21.Myneni R, Keeling C, Tucker CJ, Asrar G, Nemani R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature. 1997;386:698–702. [Google Scholar]
- 22.Niemand C, Koster B, Prasse H, Grunwald T, Bernhofer C. Relating tree phenology with annual carbon fluxes at Tharandt forest. Meteorol Z. 2005;14:197–202. [Google Scholar]
- 23.Richardson A, et al. Influence of spring and autumn phonological transitions on forest ecosystem productivity. Philos T R Soc B. 2010;365:3227–3246. doi: 10.1098/rstb.2010.0102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gu L, et al. The 2007 eastern US spring freeze: Increased cold damage in a warming world? Bioscience. 2008;58:253–262. [Google Scholar]
- 25.Rigby JR, Porporato A. Spring frost risk in a changing climate. Geophys Res Lett. 2008;35:L12703. 10.1029/2008GL033955. [Google Scholar]
- 26.Bennie J, Kubin E, Wiltshire A, Huntley B, Baxter R. Predicting spatial and temporal patterns of bud-burst and spring frost risk in north-west Europe: the implications of local adaptation to climate. Glob Change Biol. 2010;16:1503–1514. [Google Scholar]
- 27.Tanja S, et al. Air temperature triggers the recovery of evergreen boreal forest photosynthesis in spring. Glob Change Biol. 2003;9:1410–1426. [Google Scholar]
- 28.Ichii K, Kawabata A, Yamaguchi Y. Global correlation analysis for NDVI and climatic variables and NDVI trends: 1982–1990. Int J Remote Sens. 2002;23:3873–3878. [Google Scholar]
- 29.Gong D, Shi P. Northern hemispheric NDVI variations associated with large-scale climate indices in spring. Int J Remote Sens. 2003;24:2559. [Google Scholar]
- 30.Keeling CD, Chin JFS, Whorf TP. Increased activity of northern vegetation in inferred from atmospheric CO2 measurements. Nature. 1996;382:146–149. [Google Scholar]
- 31.Randerson JT, Field CB, Fung IY, Tans PP. Increases in early season ecosystem uptake explain recent changes in the seasonal cycle of atmospheric CO2 at high northern latitudes. Geophys Res Lett. 1999;26:2765–2768. [Google Scholar]
- 32.Kimball JS, McDonald KC, Running SW, Frolking SE. Satellite radar remote sensing of seasonal growing seasons for boreal and sub-alpine evergreen forests. Remote Sens Environ. 2004;90:243–258. [Google Scholar]
- 33.White MA, Thomton PE, Running SW. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Global Biogeochem Cy. 1997;11:217–234. [Google Scholar]
- 34.Piao SL, Friedlingstein P, Ciais P, Viovy N, Demarty J. Growing season extension and its effects on terrestrial carbon flux over the last two decades. Global Biogeochem Cy. 2007;21:GB3018. 10.1029/2006GB002888. [Google Scholar]
- 35.Tucker C, et al. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int J Biometeorol. 2001;45:184–190. doi: 10.1007/s00484-001-0109-8. [DOI] [PubMed] [Google Scholar]
- 36.Bazzaz FA, Jasienski M, Thomas SC, Wayne P. Microevolutionary responses in experimental populations of plants to CO2-enriched environments—parallel results from 2 model systems. Proc Natl Acad Sci USA. 1995;92:8161–8165. doi: 10.1073/pnas.92.18.8161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Norby RJ, et al. Forest response to elevated CO2 is conserved across a broad range of productivity. Proc Natl Acad Sci USA. 2005;102:18052–18056. doi: 10.1073/pnas.0509478102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang X, et al. Drought-induced vegetation stress in southwestern North America. Environ Res Lett. 2010;5:024008. 10.1088/1748-9326/5/2/024008. [Google Scholar]
- 39.Braswell BH, Schimel DS, Linder E, Moore B. The response of global terrestrial ecosystems to interannual temperature variability. Science. 1997;278:870–873. [Google Scholar]
- 40.Goetz SJ, Bunn AG, Fiske GJ, Houghton RA. Satellite-observed photosynthetic trends across boreal North America associated with climate and fire disturbance. Proc Natl Acad Sci USA. 2005;102:13521–13525. doi: 10.1073/pnas.0506179102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bond-Lamberty B, Peckham SD, Ahl DE, Gower ST. Fire as the dominant driver of central Canadian boreal forest carbon balance. Nature. 2007;450:89–92. doi: 10.1038/nature06272. [DOI] [PubMed] [Google Scholar]
- 42.Verbyla D. The greening and browning of Alaska based on 1982–2003 satellite data. Global Ecol Biogeogr. 2008;17:547–555. [Google Scholar]
- 43.Gillett NP, Weaver AJ, Zwiers FW, Flannigan MD. Detecting the effect of climate change on Canadian forest fires. Geophys Res Lett. 2004;31:L18211. 10.1029/2004GL020876. [Google Scholar]
- 44.Stocks BJ, et al. Large forest fires in Canada, 1959–1997. J Geophys Res. 2002;107:8149. 10.1029/2001JD000484. [Google Scholar]
- 45.Ayres MP, Lombardero MJ. Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci Total Environ. 2000;262:263–286. doi: 10.1016/s0048-9697(00)00528-3. [DOI] [PubMed] [Google Scholar]
- 46.Soja AJ, et al. Climate-induced boreal forest change: Predictions versus current observations. Global Planet Change. 2007;56:274–296. [Google Scholar]
- 47.Neigh CSR, Tucker CJ, Townshend JRG. North American vegetation dynamics observed with multi-resolution satellite data. Remote Sens Environ. 2008;112:1749–1772. [Google Scholar]
- 48.Le Quere C, et al. Trends in the sources and sinks of carbon dioxide. Nat Geosci. 2009;2:831–836. [Google Scholar]
- 49.Slayback DA, Pinzon JE, Los SO, Tucker CJ. Northern hemisphere photosynthetic trends 1982–99. Glob Change Biol. 2003;9:1–15. [Google Scholar]
- 50.Mesinger F, et al. North American regional reanalysis. B Am Meteorol Soc. 2006;87:343–360. [Google Scholar]
- 51.Kanamitsu M, et al. NCEP-DOE AMIP-II Reanalysis (R-2) B Am Meteorol Soc. 2002;83:1631–1643. [Google Scholar]
- 52.Keyantash J, Dracup JA. The quantification of drought: An evaluation of drought indices. B Am Meteorol Soc. 2002;83:1167–1180. [Google Scholar]
- 53.Guttman NB, Wallis JR, Hosking JRM. Spatial comparability of the Palmer Drought Severity Index. J Am Water Resour As. 1992;28:1111–1119. [Google Scholar]
- 54.Seneviratne SI, et al. Investigating soil moisture-climate interactions in a changing climate: A review. Earth-Sci Rev. 2010;99:125–161. [Google Scholar]
- 55.Dai AG, Trenberth KE, Qian TT. A global dataset of Palmer Drought Severity Index for 1870–2002: Relationship with soil moisture and effects of surface warming. J Hydrometeorol. 2004;5:1117–1130. [Google Scholar]
- 56.Jones PD, Briffa KR. Global surface air temperature variations during the twentieth century: Part 1, spatial, temporal and seasonal details. The Holocene. 1992;2:165–179. [Google Scholar]
- 57.Toms JD, Lesperance ML. Piecewise regression: A tool for identifying ecological thresholds. Ecology. 2003;84:2034–2041. [Google Scholar]
- 58.Holben BN. Characteristics of maximum-value composite images from temporal AVHRR data. Int J Remote Sens. 1986;7:1417–1434. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.