

Abstract
Social isolation can exacerbate the negative consequences of stress and increase the risk of developing psychopathology. However, the influence of living alone on experiences generally considered to be beneficial to the brain, such as physical exercise, remains unknown. We report here that individual housing prevents the positive influence of short-term running on adult neurogenesis in the hippocampus of rats and, in the presence of additional stress, suppresses the generation of new neurons. Individual housing also influenced corticosterone levels – runners in both housing conditions had elevated corticosterone during the active phase but individually housed runners had higher levels of the hormone in response to stress. Moreover, lowering corticosterone levels converts the influence of short-term running on neurogenesis in individually housed animals from negative to positive. These results suggest that, in the absence of social interaction, normally beneficial experiences can exert a potentially deleterious influence on the brain.
The physiological consequences of stressful experiences can range dramatically from one person to the next.1,2 Differential outcomes can depend on psychological factors related to the stressor itself, such as whether or not it is predictable or controllable.3,4 However, it is clear that even identical perturbations can elicit varying responses depending on the individual. Numerous studies indicate that individual differences in stress responses are determined by the environment. In this regard, social context has been shown to influence stress responsiveness. In rats, group housing can buffer the influence of some types of stress on hypothalamic pituitary adrenal (HPA) axis activity.5-7 Likewise, social support lessens the negative impact of stress on the brain and appears to protect against psychiatric illness in humans.8,9
Exercise is an experience that is associated with health benefits and yet it results in physiological changes that are indicative of stress. Running is known to activate the HPA axis, an effect that persists even after physical fitness is achieved.10 Despite large and continual elevations in stress hormones, running is generally linked to positive influences on the brain and behavior. For example, running has been associated with elevated expression of brain derived neurotrophic factor, increased synaptic plasticity and improved performance on spatial navigation learning tasks.11-13 Numerous studies have also shown that running increases the number of new neurons in the dentate gyrus of adult rats.12-15 These results may seem paradoxical since glucocorticoids have been shown to suppress adult neurogenesis.16-21 The majority of studies on running and neurogenesis have examined animals living in groups,12-15 and the possibility that social interaction buffers the brains of runners from the negative effects of elevated glucocorticoids remains unexplored.
Here we report that social isolation prevents the positive effects of short-term exercise on adult neurogenesis in the dentate gyrus. Enhancement of adult neurogenesis eventually occurs in individually housed runners, but only after a considerably longer period of running. Both isolated and group-housed runners exhibit elevations in glucocorticoids during the active period but individually housed runners experience higher levels of corticosterone at a different point in the circadian rhythm as well as in response to stress. Reducing corticosterone levels reverses the effects of running on adult neurogenesis only in animals living alone. These findings suggest that social experience buffers the runner from the negative actions of elevated glucocorticoids by blunting stress reactivity and by preventing corticosterone from suppressing adult neurogenesis. By contrast, isolation renders the hippocampus more susceptible to the suppressive actions of glucocorticoids on adult neurogenesis, and delays the positive consequences of physical activity.
Results
Effects of short-term running on adult neurogenesis depend on the social context
To determine whether social isolation affects the response of adult neurogenesis to physical activity, animals were exposed to 12 days of running or inactivity, while housed in groups or alone. Animals were injected with BrdU (50mg/kg), a marker of DNA synthesis, once daily for 10 days beginning on day 2 of the experiment. Runners housed in groups exhibited a significant increase in the number of BrdU labeled cells in the dentate gyrus, compared to group-housed controls (for the main effect of housing; F1,20=37.15, p= .001; Fig. 1, 2A-B). By contrast, individually-housed runners showed the opposite effect; a significant decrease in the number of BrdU labeled cells, compared to individually-housed controls (for the interaction of housing and physical activity, F1,20=16.68, p=.006; Fig.1, 2A-B). No significant differences in BrdU labeled cell number were seen between group- and individually-housed controls. Similar to the effect seen with BrdU labeling, the number of cells stained for Ki67, an endogenous marker of cell proliferation, was increased in group-housed runners relative to group-housed controls, and decreased in individually-housed runners relative to individually-housed controls (for the interaction of housing and physical activity, F1,20=36.35, p=.001; Fig. 1, 2C-D).
Figure 1. Social isolation reverses the positive influence of short-term running on adult neurogenesis.

The top panel outlines the BrdU injection protocol for these studies. The animals received daily injections of BrdU on days 2-11 and were perfused 24 hours after the last injection. (Top graph) The number of BrdU labeled cells is lower in individually-housed short-term runners compared to individually-housed controls. By contrast, the number of BrdU labeled cells in group-housed animals is elevated compared to group-housed controls. (Bottom graph) Similar results were obtained with the endogenous marker of cell proliferation, Ki67 – individually-housed runners have fewer Ki67 labeled cells, while group-housed runners have more Ki67 labeled cells, when compared with their respective control conditions. Asterisks indicate significance from controls following 2-way ANOVA and Tukey post hoc comparisons (p<.05).
Figure 2. Social context mediates the running effect on cell proliferation - newly generated cells express neuronal, glial, and endothelial markers.

[A-B], The number of BrdU labeled cells in the dentate gyrus of an individually-housed runner (A) is lower than that of a group-housed runner (B). [C-D], The number of Ki67-labeled cells in an individually-housed runner (C) is lower than that of a group-housed runner (D). [E-G], High magnification examples of proliferating cells in the dentate gyrus of a long-term runner – (E) BrdU labeled cells; (F) Ki67-labeled cells and; (G) phosphorylated histone H3 labeled cell that appears to be undergoing cytokinesis. [H-K], Immunofluorescence double-labeling for cell type specific markers (green) and BrdU (red). (H) cell double labeled with BrdU and a marker of mature neurons, NeuN; (I) cell double labeled with BrdU and a marker of immature and mature neurons, Tuj1; (J) cell double labeled with BrdU and a marker of astroglia, GFAP; and (K) cell double labeled with BrdU and a marker of endothelial cells, vWF . Arrows indicate labeled [A-G], or double-labeled [H-K] cells. Scale bars = 10μm.
Analysis of the phenotype of BrdU labeled cells revealed that the majority of new cells expressed neuronal markers, Tuj1 (~80%) and NeuN (~66%), at similar percentages across conditions. Much smaller percentages of BrdU labeled cells stained for an astroglial marker, GFAP (~7%) or an endothelial marker, von Willebrand factor (~6%) (Fig. 2H-K). These findings suggest that changes in cell proliferation with running primarily result in changes in adult neurogenesis.
Social housing and physical activity
It is unlikely that the differential effects of running on adult neurogenesis in group- versus individually-housed animals are the result of group-housed animals engaging in higher levels of activity because the total amount of running (for group-housed animals, daily km/3 – rats were housed 3 per cage) was similar for animals in both social conditions (t8=0.6136, ns; Supplemental Fig. 1A). In addition, no differences were observed in the frequency or duration of running bouts between social conditions (for the frequency of running bouts, t8=1.866, p=.099; for the duration of running bouts, t8=0.576, p=.58). Finally, during the active phase, individually housed animals engaged in slightly more locomotor activity when not in the running wheels than group housed animals. It is also unlikely that the differential effects of running on adult neurogenesis across social conditions are the result of variation in the amount of other behaviors, since no differences were observed between these groups during either the inactive or active phase in the amount of time spent eating, drinking, grooming or inactivity/sleeping.
Social housing alters corticosterone levels
The differential effects of running on adult neurogenesis observed with social condition may be associated with alterations in levels of corticosterone. To assess this possibility, we measured corticosterone in trunk blood collected following decapitation at two different times: at the onset of the dark period (7pm), and 4hr after lights-out (11pm). There is a diurnal rhythm in corticosterone – levels are known to be highest around the onset of the dark period. As previously reported,10 we observed substantially greater elevations in circulating corticosterone in runners at 7PM, the beginning of the active phase (for the main effect of physical activity, F1,20=17.21, p=.0005, Fig.3). No differences were observed between individual and group housed runners in this response (for the interaction of physical activity and housing, F1,20=1.67, p=.2110). In contrast, at the 11 PM time point, significant differences in corticosterone levels were observed with social condition – both controls and runners housed individually had higher levels of corticosterone than those housed in groups (for the main effect of housing, F1,20=8.909, p=.0073, Fig. 3).
Figure 3. Glucocorticoid levels are altered by running and social housing.

(A) Running increases corticosterone levels in both individually housed and group housed animals at the beginning of the active phase (7PM), when corticosterone levels are at their circadian peak. Asterisk (*) indicates significant difference from controls following 2-way ANOVA. (B) Group housing is associated with lower levels of corticosterone at a later time period during the active phase (11PM). Note also the overall decline in corticosterone levels between 7PM and 11PM. Asterisk (*) indicates significant difference from socially isolated animals (C) Group housing and running interact to buffer the stress response. Group housed runners fail to show a stress-induced increase in corticosterone, when tested during the inactive phase (10AM). A stress-induced increase in corticosterone levels was observed in all other conditions. Asterisks equal significant difference from baseline within each experimental condition (p<.05).
To determine whether social housing alters the response of the HPA axis to stress in runners, we examined baseline, stress and recovery corticosterone levels during the inactive phase from controls and runners housed either individually or in groups. In controls, restraint stress elevated corticosterone levels 30 min later – these levels returned to baseline 2 hours after the cessation of stress in both social conditions. However, within runners, substantial differences in the corticosterone response to stress were observed between the social conditions. Physical activity and group housing interact to alter the corticosterone response following restraint stress (F2,18=3.828, p=.041). Specifically, group housed runners showed no elevation in corticosterone levels 30 min after restraint stress (Fig.3). These data suggest that compared to individual housing, group housing may reduce the overall exposure to corticosterone, particularly in runners.
Glucocorticoids and socially isolated runners
To investigate whether adrenal steroids play a role in the different effects of running on adult neurogenesis in group versus individually housed animals, we repeated the same experiment on animals that were either sham-operated or adrenalectomized and administered low-dose corticosterone in the drinking water (25μg/ml in 0.9% saline). Prevention of the running-induced increase in glucocorticoid levels converted the effect of running on neurogenesis from negative to positive in individually-housed animals (for the effect of glucocorticoid status, F1,38=8.89, p=.005; for the interaction of running, glucocorticoid status, and housing, F1,38=7.75, p=.008, Fig. 4). Lowering corticosterone levels did not alter the positive effects on adult neurogenesis in group-housed runners. Thus, runners housed either individually or in groups exhibited increased neurogenesis when glucocorticoid levels are clamped. By contrast, sham-operated rats showed the same pattern of results observed with intact animals – for individually-housed rats, 12 days of running decreased the number of BrdU labeled cells, while for group-housed animals, the same period of running had the opposite effect.
Figure 4. Lowering glucocorticoid levels reverses the suppression of neurogenesis in isolated runners.

The top panel describes the experimental design for this study. Controls or runners were housed in groups or individually and subjected to sham operation or bilateral adrenalectomy (ADX+CORT). ADX animals received 25μg/ml of exogenous corticosterone in drinking water. The BrdU injection protocol was identical to the one described for Fig. 1. (Graph) BrdU labeling in the dentate gyrus of sham-operated (SHAM) controls and runners exhibits the same pattern seen in intact animals. Individually-housed runners have fewer labeled cells, while group-housed runners have more labeled cells, than controls. Lowering glucocorticoid levels by removing the adrenal glands and providing a low dose of corticosterone in the drinking water (ADX+CORT) permits the enhancement of neurogenesis in individually-housed runners and does not change the running effect on neurogenesis in the group-housed animals. Asterisks indicate significant difference from controls following 2-way ANOVA with Tukey HSD post hoc tests (p<.05).
No significant differences in the amount of running were observed between group and individually housed animals with or without ADX and corticosterone replacement (for the effect of housing, F1,44=0.71, ns, for the effect of glucocorticoid status, F1,44=0.99, ns; Supplemental Fig. 1B). Radioimmunoassay verified the reduction of corticosterone levels in adrenalectomized animals across runners and controls housed in groups or individually, and no significant differences were observed among these conditions.
Longer periods of running in social isolation
Since the initial period of running in previously sedentary animals is likely to be more demanding physiologically than running over the long-term,22 we examined whether the influence of exercise on adult neurogenesis in individually-housed animals changed over time. To do this, we measured cell proliferation in individually-housed animals that ran for 3, 6, 12, 24, or 48 days and received a single injection of BrdU (300 mg/kg) 2hr prior to perfusion. We found no change in BrdU labeling until the latest time point examined, 48 days (for the effect of running, F1,47=5.32, p=.0256; for the interaction of running and time, F4,47=9.85, p=.0001, Fig. 2E, 5). Increases in cell proliferation following 48 days of running were confirmed with two endogenous markers of cell proliferation, Ki67 or phospho-histone H3 (for Ki67, t10=4.25, p=.0004; for phospho-histone H3, t10=3.07, p=.0221, Fig. 2F,G, 4). These results indicate that socially isolated animals ultimately exhibit enhanced cell proliferation, but only with relatively long periods of running.
Figure 5. A longer duration of physical activity enhances cell proliferation in the dentate gyrus of socially isolated animals.

The top panel describes the experimental design for this study – separate cohorts of individually-housed animals ran for 3, 6, 12, 24, or 48 days before being injected once with BrdU (300 mg/kg) and perfused 2 hr later. (Top graph) In socially isolated animals, the number of BrdU labeled cells in the dentate gyrus is increased by 48 days of running but not at earlier time points examined. (Bottom graph) After 48 days of running, similar increases were observed in the numbers of cells that stained for the endogenous markers of cell proliferation Ki67 and phospho-histone H3. Error bars represent the standard error of the mean; Asterisks indicate significant difference from control following 2×5 ANOVA and Tukey HSD post hoc comparisons, p<.05, or unpaired t-test comparing Ki67 and phospho-histone H3 data from runners and controls at the 48-day time point.
Following 48 days of running, the number of BrdU labeled cells at the two week post-BrdU survival time was greater in runners (3788.0 ± 412.2 for individually-housed controls, 5979.0 ± 662.2 for individually-housed 48 day runners; t12=2.58, p=.024). Long-term running also increased the overall volume of the dentate gyrus (for the effect of running, F1,46=6.85, p=.012; Supplemental Figure 2). The majority of BrdU-labeled cells in both groups expressed neuronal markers, Tuj1 (~82%) and NeuN (~75%), while a much smaller proportion stained for the astroglial marker GFAP (~6%) or the endothelial marker von Willebrand factor (~5%). No change was noted in the proportion of cells staining for any of the markers, suggesting that differentiation of new cells was not influenced by long-term running. Changes in the amount of running per 24 hr are probably not responsible for the increase in adult neurogenesis following 48 days of running, because a plateau in daily running distance was reached considerably before (~21 days) we observed positive effects on neurogenesis.
To determine whether the positive influence of long-term running in individually housed animals is comparable to that of group-housed runners, we repeated the 48 day running experiment with animals in both social conditions. We found that 48 days of physical activity stimulates cell proliferation to a similar extent in group- and individually-housed animals (for the effect of running, F1,19=9.81, p=.005, Supplementary Fig. 3). Moreover, long-term running increases dentate gyrus volume in individually housed and group housed rats (F1,20=8.258, p=.0094; Supplementary Fig. 2). Again, group- and individually-housed animals did not differ in the overall amount of running activity (t8=0.8211,ns).
Social housing buffers the negative effects of stress
Unlike the pattern observed in our initial experiments (Fig. 1), socially isolated animals exposed to 12 days of running in the time course experiment (Fig. 5) did not exhibit decreased cell proliferation. This difference may be due to the fact that the animals in our first studies were subjected to the stress of daily handling and injection,23 while those in the current experiment were injected only once on the last day of the study. Daily handling and injection significantly elevated baseline levels of circulating corticosterone in socially isolated runners (19.04±2.83 ng/ml for undisturbed animals, 32.60±3.40 ng/ml for injected animals; t6=3.069, p=.020). Social isolation in the absence of additional stressors appears to prevent the positive influence of short-term running on adult neurogenesis such that the number of BrdU labeled cells is similar to that of sedentary controls. In the presence of additional stressors (handling and injection), however, the influence of short-term running is negative, resulting in a net decrease in the number of new cells.
To more directly test the possibility that the decreases in adult neurogenesis observed in individually housed runners exposed to multiple injections were the result of an interaction among isolation, running and stress, we repeated the 12 day social housing experiment, but replaced the daily BrdU injections with a brief cold swim stress. This stressor was of comparable duration to that experienced during restraint and BrdU injection (60-90 sec). These animals received only a single BrdU injection, on the last day of the experiment. We found that individually housed runners subjected to daily cold water stress exhibit decreased cell proliferation compared to individually housed controls. By contrast, group housed runners exposed to daily cold water stress had enhanced cell proliferation (Fig.6). These results strongly suggest that the negative effects of running on neurogenesis in our first study with multiple BrdU injections are the result of the combined action of individual housing, running and daily stress. Moreover, they suggest that group housed runners are buffered from the negative effects of stress on cell proliferation.
Figure 6. Daily stress interacts with running and social housing to alter cell proliferation.

The top panel represents the experimental design for this study. Animals were subject to brief (60-90sec) daily swim stress in cold water. On the morning of the last day, animals were injected once with BrdU (300 mg/kg) and perfused 2hr later. The graph shows that daily cold swim stress resulted in a decrease in cell proliferation in individually housed runners compared to individually housed control animals. In contrast, daily cold swim stress did not prevent the enhancement of cell proliferation in group-housed runners. Asterisks equal significant difference from control following 2-way ANOVA and Tukey HSD post hoc comparisons, p<0.05.
Duration of isolation may influence the effects of running
To determine whether the negative effects of running on adult neurogenesis are dependent on living alone for a certain time prior to gaining access to the running wheel, we compared animals that were moved directly from group housed conditions to the running wheel with those that spent 1 wk living alone prior to running. We found that the duration of isolation prior to running influenced the results – a decrease in cell proliferation was only observed in the runners with 1 wk of isolation prior to running but not those with no prior isolation experience (for the effect of prior isolation, F1,20=12.69, p=.002; for the interaction of physical activity and prior isolation, F1,20=14.27, p=.0012; Fig. 7).
Figure 7. The duration of previous isolation affects the response of cell proliferation to running alone.
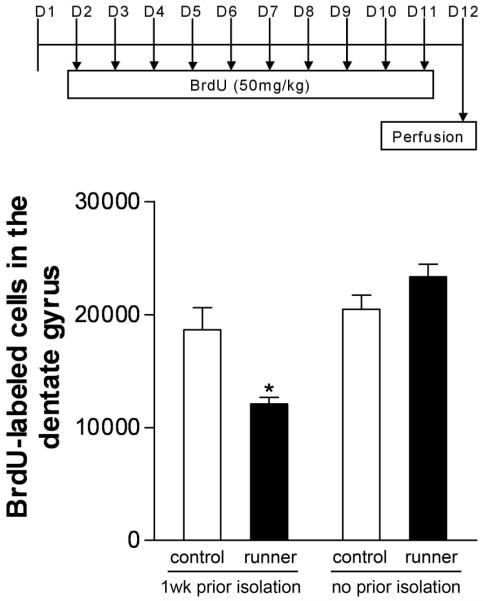
The BrdU injection protocol is identical to the one used in Figures 1 and 4. Prior to the beginning of the study, some animals were individually housed in standard cages, while other animals were housed in groups of three until the beginning of the experiment, when they were individually housed with or without a running wheel. One week of isolation prior to gaining access to the running wheel was associated with decreased cell proliferation in the dentate gyrus of runners. A shorter duration of individual housing (12 days as opposed to 19 days) resulted in a return to baseline levels of cell proliferation. This suggests a time-dependent effect of social isolation on the response of adult neurogenesis to running. Asterisk equals significant difference from control, 1 wk prior isolation – p<0.05.
Discussion
The results of these experiments demonstrate that the social context in which an experience occurs can dramatically influence the central response to stress hormones. When experienced in a group setting, running stimulates adult neurogenesis. However, when running occurs in social isolation, these positive effects are suppressed. Although runners housed in either social condition experience elevations in corticosterone during one part of the active phase, only those that run alone are vulnerable to the negative influence of glucocorticoids on neurogenesis in the hippocampus. Preventing the elevation in glucocorticoid levels in socially isolated runners stimulates neurogenesis, while group housed animals respond to short-term running with enhanced neurogenesis regardless of their glucocorticoid status.
These paradoxical effects may be the result of a competitive interaction between glucocorticoids, which suppress neurogenesis16-21 and an unidentified factor(s), which positively regulates neurogenesis, and is altered by both running and social housing. Although several candidate mechanisms likely exist; one interesting possibility is the neurotransmitter serotonin which, through its 5HT1A receptor subtype, enhances neurogenesis.24 Levels of serotonin have been shown to increase in the hippocampus with exercise,25 while expression of the 5HT1A receptor has been shown to decrease with both social isolation,26 and with prolonged elevation of corticosterone.27 The extent to which serotonin participates in interactions among social housing, running, glucocorticoids and neurogenesis, however, remains to be determined.
Prolonged exercise ultimately enhanced neurogenesis in the dentate gyrus of animals living in social isolation. This change occurred some time between 24 and 48 days of running, and was accompanied by a significant increase in the volume of the brain region. The basis for these changes in adult neurogenesis and dentate gyrus volume are unknown, but may involve alterations in the vascular structure. Angiogenesis supports cell proliferation in the subgranular zone, possibly through increased expression of neurotrophic factors28, 29 and physical activity has been shown to promote extension of the vasculature.30
It is noteworthy that socially isolated runners in the social housing study, but not those of the time course study exhibited decreased levels of adult neurogenesis. This differential response of socially isolated runners to short-term physical activity may be related to the added stress of daily injections.23 Indeed, daily handling and injection was found to significantly elevate levels of circulating corticosterone. Moreover, replacing the injections with cold water stress resulted in virtually identical effects on cell proliferation. Thus, in the presence of additional stressors, socially isolated runners exhibit decreased neurogenesis compared to controls. In the absence of additional stressors, socially isolated runners have levels of adult neurogenesis that are similar to sedentary controls. These results are consistent with a study showing that footshock-induced decreases in neurogenesis can be eliminated by social housing.31 Our data suggest that social housing alters the HPA axis response to stress- while isolated runners experience robust elevations in corticosterone in response to stress, group housed runners show no such increase in corticosterone. Collectively, these results suggest that socially isolated runners may be exposed to higher levels of corticosterone under certain conditions and furthermore, that elevations in adrenal steroids are involved in both the negative effects and prevention of the positive effects on neurogenesis since preventing elevations in glucocorticoid levels not only eliminated the negative effects but enabled the positive effects on neurogenesis to emerge.
Numerous studies have reported enhanced neurogenesis following voluntary running in animals housed in groups.12-15,32 Similar results have been obtained using forced running paradigms33,34. These latter studies also examined animals housed in groups (J. Trejo, personal communication; C-J Kim, personal communication). Some studies, however, have examined the effects of running on adult neurogenesis in individually-housed animals; most of these findings are consistent with our data, in that the duration of running used to enhance adult neurogenesis was relatively long.35-37 However, there are a few reports of increased cell proliferation in socially isolated adult rats following short bouts of running. Two of these studies examined individually-housed Spontaneously Hypertensive Rats (SHR).38,39 Since these experiments were conducted in female animals, which show estrous cycle dependent fluctuations in neurogenesis40 and running activity, and this rat strain differs from the Sprague-Dawley in the rate of hippocampal cell proliferation under basal conditions,41 it is not surprising that the physiological response to having access to a running wheel would also differ. In contrast to our results, Eadie et al42 have reported increased cell proliferation in the dentate gyrus of individually-housed adult male Sprague-Dawley rats following ~17 days of running, considerably earlier than we observed an increase in neurogenesis. This discrepancy raises the possibility that the duration of living in social isolation may be an important determinant of the effect, as these investigators housed their animals in isolation for 3 days, as opposed to the 7 day acclimation period we used, prior to the initiation of running. Our findings that the duration of individual housing alters the response of running on cell proliferation suggest this may be the case.
Running is a natural rodent behavior that appears to have a hedonic component. Naïve laboratory reared rodents will readily engage in wheel running, even without additional incentives, and can be trained to bar press for wheel access.43 Conversely, rodents accustomed to running will experience physiological and behavioral signs of withdrawal when suddenly denied access to a wheel.44,45 Our results suggest that engaging in an internally motivated, apparently pleasurable, activity can exert radically different influences on neural plasticity depending on the context in which it occurs. In a socially deprived environment, the enhancement of neurogenesis by running is delayed, and in the presence of additional stressors, neurogenesis is actually suppressed. By contrast, running in a group setting elevates neurogenesis – despite HPA axis activation, and in the context of additional stressors, such as injection or cold water stress. Although it is possible that different social contexts may alter the hedonic value of running, our behavioral data indicate no differences in the duration, frequency or distance run across social groups. Collectively, these results indicate that social isolation can both delay the enhancement of plasticity by physical exercise, and exacerbate the consequences of stressful stimuli. Structural plasticity and, in particular, adult neurogenesis in the hippocampus, have been linked to anxiety regulation and the therapeutic actions of antidepressants.46,47 Our findings present a potential mechanism whereby social isolation may predispose the organism to a negative outcome in the presence of physiologically ambiguous experiences.
Methods
Rats and Housing Conditions
Adult male Sprague-Dawley rats from Taconic (Germantown, NY; 250-300g, n=6 for group housing studies, n=5-8 in time course studies) were housed either individually or in groups of three, with or without access to a running wheel. All animals received unlimited food and water and were maintained on a 12hr light/dark schedule (lights on at 7:00am). After arrival, all animals were group-housed and acclimated to the colony for at least a week before any manipulations were started. After this time, the animals were randomly assigned to experimental conditions.
For all experiments but one, individually housed animals were moved from the standard group-housed condition 1wk prior to the start of each study. In one additional experiment, rats were transferred directly from group to individual housing with or without a running wheel – these animals were compared to similar groups of runners and controls with a 1 wk period of individual housing.
Physical Activity and Behavioral Monitoring
Distance was recorded daily, between 8 and 10am, from counters attached to the running wheels. Wheel circumference (111.76cm) was converted to kilometers. For group-housed animals, the total distance run every 24 hrs was divided by the number of animals in each cage (n=3). This number may underestimate the total distance run per animal since rats were occasionally observed running in pairs. Additionally, one cohort of group- and individually-housed runners was videotaped to evaluate possible differences in the duration and frequency of running bouts. These animals were marked with black ink for identification. Behavioral analysis was carried out for 10 min blocks at 7AM and 7PM each on days 3, 6, 9, and 11 of the experiment. These time points were selected because they are the times of lights on and lights off respectively. 7PM marks an extremely active phase, when animals are highly likely to engage in running. Digital videos were scored for the duration and frequency of running bouts, as well as behavior when not in the wheel, including rearing, eating, drinking, grooming, inactive/sleeping and non-wheel locomotion.
BrdU Administration and Perfusion
All animals received injections of the DNA synthesis marker bromodeoxyuridine (BrdU) between 9-11 AM. For multiple injection experiments, 50 mg/kg BrdU was administered daily for 10 consecutive days. These animals were perfused 24 hr after the last injection. This dose and treatment regimen was modeled after the initial studies of running enhanced neurogenesis.13,14 For experiments using single injections, animals were administered BrdU at a higher dose of 300mg/kg, and perfused 2hr or 2wk later. For experiments with a 2wk post-injection survival, the animals continued to have access to a running wheel during this period.
Group-housed animals were removed from the cage sequentially for injection; typical duration of restraint for BrdU administration was 60-90sec. For perfusion, all animals were deeply anaesthetized with Nembutal (Pentobarbital, 100 mg/kg IP) and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer.
Cold water swim stress
To assess the effects of a daily stressor on the response of individually housed animals to running, some animals received brief daily cold-water swim stress. These animals were removed from the cage, and placed in a bucket of ice water for 60 seconds before being towel-dried and returned to the home cage. Group-housed animals were administered swim stress in sequence. The total duration of stress (beginning with removal from the home cage until return to the home cage) was ~ 90 sec.
Corticosterone sampling and radioimmunoassay
To assess the influence of running on corticosterone levels in individually and group housed animals, blood samples were obtained from separate groups of unanesthetized animals either from the tail vein or by decapitation. Blood samples were not taken from animals included in the BrdU studies since stress is known to affect cell proliferation. For individually and group housed runners and controls, corticosterone levels were assessed at two points during the dark phase – 7PM and 11PM. Two time points during the dark phase were examined since a previous study showed a circadian effect of running on corticosterone levels with elevations observed only at the time of lights out (7PM) but not several hours later (11PM) when corticosterone levels are naturally declining.10
To assess the response of the HPA axis to stress in runners and controls housed in different social conditions, baseline, restraint stress and recovery corticosterone levels were obtained during the inactive phase, around the time when BrdU injections were normally given. Beginning at 10AM, animals were rapidly restrained and blood was obtained in less than 3 min from the tail vein. Thirty minutes later, a stress blood sample was obtained from restrained animals. The animals were then returned to their home cages where they remained for 2 hr, after which they were rapidly captured and a blood sample collected in less than 3 min. Plasma was obtained from these blood samples and corticosterone levels were determined by use of radioimmunoassay using an RIA kit (Diagnostic Products Corp, Los Angeles, CA).
Adrenalectomy and Corticosterone Replacement
To determine whether glucocorticoids are involved in the differential effects of running on neurogenesis in individually and socially housed animals, rats were subject to bilateral adrenalectomy or sham operation. Adrenalectomized animals received low-dose corticosterone replacement (25 μg/ml in 0.9% saline; Sigma) in the drinking water to both lower and normalize glucocorticoid levels. Providing corticosterone in the drinking water maintains the diurnal rhythm of glucocorticoids, and this dose has previously been shown to be sufficient to maintain the survival of mature hippocampal granule neurons.17 At the time of perfusion, blood samples were obtained from anesthetized animals to verify the efficacy of adrenalectomy. Corticosterone levels were determined from these samples via radioimmunoassay as indicated above.
Immunohistochemistry
Brain sections 40 um thick were cut on a Vibratome into a bath of PBS. For BrdU, slide-mounted or free floating sections were heated in 0.1M citric acid followed by incubation in trypsin and then 2N HCl:PBS. For endogenous markers of cell proliferation, the HCl step was omitted. The tissue was incubated overnight with primary antibody (mouse monoclonal anti-BrdU (1:200), mouse monoclonal anti-Ki67 (1:750, Novocastra) or rabbit polyclonal anti-phosphorylated histone H3 (1:200, Santa Cruz Biotech). The tissue was then rinsed and incubated with biotinylated anti-mouse or anti-rabbit (1:200, Vector) for 1hr, rinsed, incubated in avidin-biotin-HRP complex and then reacted with 0.01% diaminobenzidine (DAB, Sigma). Following this, the slides were counterstained with cresyl violet and coverslipped with Permount (Fisher Scientific).
Additional tissue sections were processed for double-labeling immunofluorescence for BrdU and cell type markers. Brain sections were treated with 2N HCl:TBS and incubated overnight with rat anti-BrdU (1:250, with 0.5% Tween-20; Accurate Chemical). Following rinses, the tissue was incubated for 1 hr in biotinylated goat anti-rat (1:250, Vector), rinsed and then incubated with streptavidin Alexa 568 (1:1000, Molecular Probes). Sections were then incubated with one of the following primary antibodies: mouse anti-NeuN (1:500, Chemicon), mouse anti-TuJ1 (1:500, Covance), guinea pig anti-GFAP (1:250, Advanced Immunochemical), or rabbit anti-vWF (1:200, Sigma) for 24 hours at 4°C. Alexa 488 secondary antibodies conjugated to the appropriate species were applied for 1 hour (Molecular Probes). Following rinses, sections were mounted and coverslipped with 90% glycerol:TBS.
Microscopic data analysis
Slides were coded until completion of the data analysis. Stereological estimates of the total number of labeled cells were determined from peroxidase stained tissue for BrdU, Ki67 or phospho-histone H3 on a 1:12 series of brain sections throughout the dentate gyrus. The volume of the dentate gyrus was estimated using Cavalieri’s principle and Image-Pro Plus software (Media Cybernetics, San Diego, CA).
A Zeiss Axiovert 510 LSM confocal microscope was used to examine tissue processed for immunofluorescence (HeNe and Argon lasers). A series of 1-um thick sections throughout the extent of each labeled cell were analyzed. For each marker in each brain, 25 BrdU-labeled cells were examined throughout the rostrocaudal extent of the dentate gyrus.
Statistical Analysis
For all studies with more than two groups, analysis of variance was performed with Tukey HSD post hoc tests. For the separate studies of 48 day runners and controls, two-tailed t tests were used to analyze cells labeled with BrdU, Ki67 and phospho-histone H3. In addition, mean distance run over each experiment was calculated for animals in each group and the results were analyzed either with analysis of variance or two-tailed t tests, depending on the number of groups.
Supplementary Material
Acknowledgements
This work was supported by NRSA predoctoral fellowship to A.S. and National Institutes of Mental Health grant to E.G.
Footnotes
Competing interests statement. The authors declare that they have no competing financial interests.
Supplementary Information accompanies this paper.
References
- 1.Selye H. The stress of life. McGraw-Hill; New York: 1976. [Google Scholar]
- 2.Roy MP, Steptoe A, Kirschbaum C. Life events and social support as moderators of individual differences in cardiovascular and cortisol reactivity. J Pers Soc Psychol. 1998;75:1273–1281. doi: 10.1037//0022-3514.75.5.1273. [DOI] [PubMed] [Google Scholar]
- 3.Weiss JM. Effects of coping behavior with and without a feedback signal on stress pathology in rats. J Comp Physiol Psychol. 1971;77:22–30. doi: 10.1037/h0031581. [DOI] [PubMed] [Google Scholar]
- 4.Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci. 2005;8:365–371. doi: 10.1038/nn1399. [DOI] [PubMed] [Google Scholar]
- 5.Weiss IC, Pryce CR, Jongen-Relo AL, Nanz-Bahr NI, Feldon J. Effect of social isolation on stress-related behavioral and neuroendocrine state in the rat. Beh Brain Res. 2004;152:279–295. doi: 10.1016/j.bbr.2003.10.015. [DOI] [PubMed] [Google Scholar]
- 6.Bartolomucci A, Palanza P, Sacerdote P, Ceresini G, Chirieleison A, et al. Individual housing induces altered immuno-endocrine responses to psychological stress in male mice. Psychoneuroendocrinology. 2002;28:540–558. doi: 10.1016/s0306-4530(02)00039-2. [DOI] [PubMed] [Google Scholar]
- 7.Ruis MAW, te Brake JHA, Buwalda B, De Boer SF, Meerlo P, et al. Housing familiar male wildtype rats together reduces the long-term adverse behavioral and physiological effects of social defeat. Psychoneuroendocrinology. 1999;24:285–300. doi: 10.1016/s0306-4530(98)00050-x. [DOI] [PubMed] [Google Scholar]
- 8.Seeman TE, McEwen BS. Impact of social environment characteristics on neuroendocrine regulation. Psychosom Med. 1996;58:459–471. doi: 10.1097/00006842-199609000-00008. [DOI] [PubMed] [Google Scholar]
- 9.McEwen BS. Allostasis and allostatic load: implications for neuropsychopharmacology. Neuropsychopharmacology. 2000;22:108–24. doi: 10.1016/S0893-133X(99)00129-3. [DOI] [PubMed] [Google Scholar]
- 10.Droste SK, Gesing A, Ulbricht S, Muller MB, Linthorst ACE, et al. Effects of long-term voluntary exercise on the mouse hypothalamic-pituitary-adrenocortical axis. Endocrinology. 2003;144:3012–3023. doi: 10.1210/en.2003-0097. [DOI] [PubMed] [Google Scholar]
- 11.Neeper SA, Gomez-Pinilla F, Choi J, Cotman CW. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res. 1996;726:49–56. [PubMed] [Google Scholar]
- 12.Farmer J, Zhao X, Gage FH, Christie BR. Effects of voluntary exercise on synaptic plasticity and gene expression in the dentate gyrus of adult male Sprague-Dawley rats in vivo. Neuroscience. 2004;124:71–79. doi: 10.1016/j.neuroscience.2003.09.029. [DOI] [PubMed] [Google Scholar]
- 13.van Praag H, Christie BR, Sejnowski TJ, Gage FH. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc Natl Acad Sci USA. 1999;96:13427–13431. doi: 10.1073/pnas.96.23.13427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.van Praag H, Kempermann G, Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci. 1999;2:266–270. doi: 10.1038/6368. [DOI] [PubMed] [Google Scholar]
- 15.Fabel K, Fabel K, Palmer TD. VEGF is necessary for exercise-induced neurogenesis. Eur J Neurosci. 2003;18:2803–2812. doi: 10.1111/j.1460-9568.2003.03041.x. [DOI] [PubMed] [Google Scholar]
- 16.Gould E, Cameron HA, Daniels DC, Woolley CS, McEwen BS. Adrenal hormones suppress cell division in the adult rat dentate gyrus. J Neurosci. 1992;12:3642–3650. doi: 10.1523/JNEUROSCI.12-09-03642.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cameron HA, Gould E. Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience. 1994;61:203–209. doi: 10.1016/0306-4522(94)90224-0. [DOI] [PubMed] [Google Scholar]
- 18.Tanapat P, Hastings NB, Rydel TA, Galea LA, Gould E. Exposure to fox odor inhibits cell proliferation in the hippocampus of adult rats via an adrenal hormone-dependent mechanism. J Comp Neurol. 2001;437:496–504. doi: 10.1002/cne.1297. [DOI] [PubMed] [Google Scholar]
- 19.Montaron MF, Drapeau E, Dupret D, Kitchener P, Aurousseau C, et al. Lifelong corticosterone level determines age-related decline in neurogenesis and memory. Neurobiol Aging. doi: 10.1016/j.neurobiolaging.2005.02.014. in press. [DOI] [PubMed] [Google Scholar]
- 20.Cameron HA, McKay RD. Restoring production of hippocampal neurons in old age. Nat Neurosci. 1999;2:894–897. doi: 10.1038/13197. [DOI] [PubMed] [Google Scholar]
- 21.Ambrogini P, Orsini L, Mancini C, Ferri P, Barbanti I, et al. Persistently high corticosterone levels but not normal circadian fluctuations of the hormone affect cell proliferation in the adult rat dentate gyrus. Neuroendocrinology. 2002;76:366–372. doi: 10.1159/000067581. [DOI] [PubMed] [Google Scholar]
- 22.Allen DL, Harrison BC, Maas A, Bell ML, Byrnes WC, et al. Cardiac and skeletal muscle adaptations to voluntary wheel running in the mouse. J Appl Physiol. 2001;90:1900–1908. doi: 10.1152/jappl.2001.90.5.1900. [DOI] [PubMed] [Google Scholar]
- 23.Cassano WJ, Jr, D’mello AP. Acute stress-induced facilitation of the hypothalamic-pituitary-adrenal axis: evidence for the roles of stressor duration and serotonin. Neuroendocrinology. 2001;74:167–177. doi: 10.1159/000054683. [DOI] [PubMed] [Google Scholar]
- 24.Banasr M, Hery M, Printemps R, Daszuta A. Serotonin-induced increases in adult cell proliferation and neurogenesis are mediated through different and common 5HT receptor subtypes in the dentate gyrus and the subventricular zone. Neuropsychopharmacology. 2004;29:450–460. doi: 10.1038/sj.npp.1300320. [DOI] [PubMed] [Google Scholar]
- 25.Gomez-Merino D, Bequet F, Berthelot M, Chennaoui M, Guezennec CY. Site-dependent effects of an acute intensive exercise on extracellular 5-HT and 5-HIAA levels in rat brain. Neurosci Lett. 2001;301:143–6. doi: 10.1016/s0304-3940(01)01626-3. [DOI] [PubMed] [Google Scholar]
- 26.Schiller L, Jahkel M, Kretzschmar M, Brust P, Oehler J. Autoradiographic analyses of 5HT1A and 5HT2A receptors after social isolation in mice. Brain Res. 2003;980:169–178. doi: 10.1016/s0006-8993(03)02832-4. [DOI] [PubMed] [Google Scholar]
- 27.Chalmers DT, Kwak SP, Mansour A, Akil H, Watson SJ. Corticosteroids regulate brain hippocampal 5HT1A receptor mRNA expression. J Neurosci. 1993;13:914–923. doi: 10.1523/JNEUROSCI.13-03-00914.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Palmer TD, Willhoite AR, Gage FH. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol. 2000;425:479–494. doi: 10.1002/1096-9861(20001002)425:4<479::aid-cne2>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- 29.Leventhal C, Rafii S, Rafii D, Shahar A, Goldman SA. Endothelial trophic support of neuronal production and recruitment from the adult mammalian subependyma. Mol Cell Neurosci. 1999;13:450–464. doi: 10.1006/mcne.1999.0762. [DOI] [PubMed] [Google Scholar]
- 30.Lopez-Lopez C, LeRoith D, Torres-Aleman I. Insulin-like growth factor I is required for vessel remodeling in the adult brain. Proc Natl Acad Sci USA. 2004;101:9833–9838. doi: 10.1073/pnas.0400337101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Westenbroek C, Den Boer JA, Veenhuis M, Ter Horst G. Chronic stress and social housing differentially affect neurogenesis in male and female rats. Brain Res Bull. 2004;64:303–308. doi: 10.1016/j.brainresbull.2004.08.006. [DOI] [PubMed] [Google Scholar]
- 32.Brown J, Cooper-Kuhn CM, Kempermann G, van Praag H, Winkler J, et al. Enriched environment and physical activity stimulate hippocampal but not olfactory bulb neurogenesis. Eur J Neurosci. 2003;17:2042–2046. doi: 10.1046/j.1460-9568.2003.02647.x. [DOI] [PubMed] [Google Scholar]
- 33.Trejo JL, Carro E, Torres-Aleman I. Circulating IGF-1 mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J Neurosci. 2001;21:1628–1634. doi: 10.1523/JNEUROSCI.21-05-01628.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kim Y-P, Kim H, Shin MS, Chang H-K, Jang M-H, et al. Age-dependence of the effect of treadmill exercise on cell proliferation in the dentate gyrus of rats. Neurosci Lett. 2004;355:152–154. doi: 10.1016/j.neulet.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 35.Rhodes JS, van Praag H, Garland T, Gage FH. Exercise increases hippocampal neurogenesis to high levels but does not improve spatial learning in mice bred for increased voluntary wheel-running. Behav Neurosci. 2003;117:1006–1016. doi: 10.1037/0735-7044.117.5.1006. [DOI] [PubMed] [Google Scholar]
- 36.Holmes MM, Galea LA, Mistlberger RE, Kempermann G. Adult hippocampal neurogenesis and voluntary running activity: circadian and dose-dependent effects. J Neurosci Res. 2004;76:216–222. doi: 10.1002/jnr.20039. [DOI] [PubMed] [Google Scholar]
- 37.Bjornebekk A, Mathe AA, Brene S. The antidepressant effect of running is associated with increased hippocampal cell proliferation. Int J Neuropsychopharm. 2005;8:1–12. doi: 10.1017/S1461145705005122. [DOI] [PubMed] [Google Scholar]
- 38.Persson AI, Naylor AS, Jonsdottir IH, Nyberg F, Eriksonn PS, et al. Differential regulation of hippocampal cell proliferation by opioid receptor antagonists in running and non-running spontaneously hypertensive rats. Euro J Neurosci. 2004;19:1847–1855. doi: 10.1111/j.1460-9568.2004.03268.x. [DOI] [PubMed] [Google Scholar]
- 39.Naylor AS, Persson AI, Erikkson PS, Jonsdottir IH, Thorlin T. Extended voluntary running inhibits exercise induced adult hippocampal progenitor proliferation in the spontaneously hypertensive rat. J Neurophysiol. 2004;93:2406–2414. doi: 10.1152/jn.01085.2004. [DOI] [PubMed] [Google Scholar]
- 40.Tanapat P, Hastings NB, Reeves AJ, Gould E. Estrogen stimulates a transient increase in the number of new neurons in the dentate gyrus of the adult female rat. J Neurosci. 1998;19:5792–801. doi: 10.1523/JNEUROSCI.19-14-05792.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Perfilieva E, Risedal A, Nyberg J, Johansson BB, Eriksson PS. Gender and strain influence on neurogenesis in dentate gyrus of young rats. J Cereb Blood Flow Metab. 2001;21:211–7. doi: 10.1097/00004647-200103000-00004. [DOI] [PubMed] [Google Scholar]
- 42.Eadie BD, Redila VA, Christie BR. Voluntary exercise alters the cytoarchitecture of the adult dentate gyrus by increasing cellular proliferation, dendritic complexity, and spine density. J Comp Neurol. 2005;486:39–47. doi: 10.1002/cne.20493. [DOI] [PubMed] [Google Scholar]
- 43.Iversen IH. Techniques for establishing schedules with wheel running as reinforcement in rats. J. Exp. Anal. Behav. 1993;60:219–238. doi: 10.1901/jeab.1993.60-219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hoffmann P, Thorén P, Ely D. Effect of voluntary exercise on open-field behavior and on aggression in the spontaneously hypertensive rat (SHR) Behav Neural Biol. 1987;47:346–355. doi: 10.1016/s0163-1047(87)90461-4. [DOI] [PubMed] [Google Scholar]
- 45.Widenfalk J, Olson L, Thoren P. Deprived of habitual running, rats downregulate BDNF and TrkB messages in the brain. Neurosci Res. 1999;34:125–32. doi: 10.1016/s0168-0102(99)00051-6. [DOI] [PubMed] [Google Scholar]
- 46.Santarelli L, Saxe M, Gross C, Surget A, Battallia F, et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science. 2003;301:805–809. doi: 10.1126/science.1083328. [DOI] [PubMed] [Google Scholar]
- 47.Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci. 2000;20:9104–9110. doi: 10.1523/JNEUROSCI.20-24-09104.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.