

Abstract
Human cord blood stem cells (hCBSCs) have been reported to generate hepatocyte-like cells and thus hold promise for repairing damaged liver. However, the frequency of hCBSC-derived hepatocytes varies tremendously between different studies, and it is still controversial as to whether hCBSC-derived cells can transdifferentiate into hepatocytes or simply fuse to recipient hepatocytes. We used the β-glucuronidase–deficient nonobese diabetic/severe combined immunodeficient/mucopolysaccharidosis type VII (NOD/SCID/MPSVII) mouse model for better identification of engrafted cells. We transplanted lineage-depleted human umbilical cord blood–derived cells with high aldehyde dehydrogenase activity (ALDHhiLin−) into irradiated NOD/SCID/MPSVII mice followed by carbon tetrachloride administration to induced liver damage. ALDHhiLin− cells were efficiently engrafted in the recipient mouse livers and improved recovery of the mice from toxic insult. The percentage of human cells in these livers ranged between 3% and 14.2% using quantitative real-time polymerase chain reaction. Furthermore, human-originated cells expressing liver-specific α1-antitrypsin messenger RNA, albumin and hepatocyte nuclear factor 1 protein were detected in the recipient livers. Interestingly, human versus murine centromeric fluorescent in situ hybridization analysis on the liver sections demonstrated that most human cells were not fused to mouse cells. However, the majority of the human originated albumin-expressing cells also carried mouse genetic material, hence were the product of cell fusion.
Conclusion
hCBSCs or their progeny may home to the injured liver and release trophic factors that hasten tissue repair, whereas fusion of these cells with hepatocytes may occur rarely and contribute to a lesser extent to liver repair.
Liver transplantation remains the only therapeutic option for many acute and chronic end-stage liver diseases. However, this approach is limited by a serious shortage of donor organs required for transplantation. Several studies have suggested that bone marrow stem cells (BMSCs) could generate hepatocytes (reviewed by Thorgeirsson and Grisham1). The most successful liver repopulation with bone marrow– derived cells was observed in mice with fumarylactoacetate hydrolase deficiency resulting in functional correction of liver disease.2,3 We previously showed that highly purified CD34+CD38−CD7− human cord blood stem cells (hCBSCs) could give rise to hepatocyte-like cells in immunodeficient mice.4 However, the frequency of hepatocyte-like cells derived from hematopoietic cells in most mouse models was rare (≤10−4).1 The mechanisms responsible for these phenomena are controversial. Both direct transdifferentiation5–7 of BMSCs to hepatocytes and fusion between BMSC-derived cells and recipient hepatocytes have been reported.3,8–13 Therefore, critical issues have yet to be resolved in this field of research.
To further evaluate the contribution of hCBSCs to the repair of damaged liver and to further evaluate the responsible mechanism, we used the β-glucuronidase (GUSB)-deficient nonobese diabetic/severe combined immunodeficient/mucopolysaccharidosis type VII (NOD/SCID/MPSVII) mouse model for better identification of engrafted human cells. Simultaneous detection of tissue-specific markers and the origin of cells has been one of the major challenges in the field of stem cell fate studies. One commonly used approach is to transplant donor cells into sex-mismatched recipients followed by fluorescence in situ hybridization (FISH) analysis of the X and Y chromosome to trace donor-derived cells. However, FISH is not a trivial technique, and detection of the Y chromosome may be error-prone.1 Another approach is to transduce donor cells with green fluorescent protein to label them. However, this method involves undesired genetic manipulation of donor cells. Our GUSB-deficient NOD/SCID/MPSVII mouse model allows the sensitive detection of donor cells across species with endogenous levels of GUSB in engrafted tissues.14
We and others have previously shown that lineage-depleted (Lin−) human umbilical cord blood– derived cells with high aldehyde dehydrogenase activity (ALDHhi) are enriched for primitive cells, primarily hematopoietic stem cells.15–17 In the current studies, irradiated NOD/SCID/MPSVII mice were transplanted with ALDHhiLin− cells and treated with CCl4 to induce liver damage. We found that ALDHhiLin− cells were efficiently engrafted in the recipient mouse livers. Furthermore, cells expressing human liver-specific markers were also found in these livers. Rare fusions between human and mouse cells were responsible for the appearance of these hepatocyte-like cells derived from hCBSCs.
Materials and Methods
Human Cell Purification by ALDH Activity
Human umbilical cord blood Lin− cells with either high or low ALDH activity were isolated as described.17
NOD/SCID/MPSVII Mouse Model
The NOD/SCID/MPSVII−/− mouse was bred and genotyped as described14 in our colony at Washington University and later at the University of California Davis, in compliance with all regulatory committees. ALDHloLin− (2 to 4 × 105) or ALDHhiLin− (2 to 4 × 105) cells were transplanted via tail vein injection into 8- to 12-week-old, sublethally irradiated (300 cGy) NOD/SCID/MPS-VII−/− mice. Mice injected with phosphate-buffered saline (PBS) served as nontransplanted controls. Four weeks later, these mice were injected intraperitoneally with 0.2 μL of CCl4 dissolved in 100 μL corn oil twice a week for 3 weeks to induce liver damage. Mice not injected with CCl4 also served as controls. All mice were analyzed 7 weeks after transplantation.
Fluorescence-Activated Cell Sorting Analysis of Human Cell Engraftment in Transplanted Mice
Cells from livers, spleens, bone marrow, and peripheral blood were collected from transplanted and control mice and analyzed via fluorescence-activated cell sorting for the presence of human CD45 (BD, San Jose, CA) as described.17
Immunohistochemistry, Immunofluorescence, and Terminal Deoxynucleotidyl Transferase–Mediated dUTP Nick-End Labeling
Mouse livers were frozen in optimal cutting temperature embedding medium (Sakura, Torrance, CA) and serial sections were taken at 10-μm thickness. Slides were stained for GUSB activity and counterstained with methyl green as described.18 For immunostaining, frozen sections were fixed for 20 minutes in 4% paraformaldehyde, washed with PBS, and penetrated for 30 minutes with 0.2% Triton-X100 in PBS. These slides were then blocked for 1 hour with mouse-on-mouse reagent (Vector Laboratories, CA) or 1% bovine serum albumin and incubated for 1 to 2 hours with primary antibodies. Following washing with PBS, slides were incubated with a secondary antibody (goat anti-rabbit IgG-Alexa Fluor 595, Invitrogen, CA) for 1 hour if the primary antibody was not labeled with a fluorochrome. Slides were washed with PBS and mounted using 4,6-diaminidino-2-phenylindole (DAPI)-containing mounting medium (Vector Laboratories). Fluorescein isothiocyanate– conjugated anti-human albumin antibody (Dako, CA), rabbit anti– hepatocyte nuclear factor 1 (HNF1) (Santa Cruz Biotechnology, CA), and Alexa fluor 488 – conjugated phosphohistone H3 (Cell Signaling Technology, MA) antibodies were diluted at 1:100, 1:50, or 1:25, respectively. Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling of the liver sections was performed using the DeadEnd Fluorometric TUNEL System (Promega, CA) according to the manufacturer’s specifications.
Fluorescence In Situ Hybridization
Frozen sections from recipient mouse livers were co-stained with pan-centromeric chromosome probes for either mouse or human (Cambio, Dry Drayton, Cambridge) according to the manufacturer’s specifications with the following minor modifications. Slides were incubated for 20 minutes in 10 ng/mL proteinase K. To denature DNA, slides were immersed in 70% formamide in 2×SSC at 66°C for 5 minutes. For colocalization of human albumin protein (ALB) and FISH, slides were first stained with anti-human albumin antibody and later subjected to FISH as described above.
Polymerase Chain Reaction (PCR) and Quantitative Real-time PCR
Mouse liver cells were mechanically dissociated. Genomic DNA was extracted from these cells as described.19 Amplification of the human alu gene via PCR using primers as described20 was under the following conditions: 94°C for 5 minutes, 25 cycles of 94°C for 30 seconds, 62°C for 45 seconds, and 72°C for 45 seconds. Quantitative real-time PCR was performed using the ABI 7300 system under default conditions (Applied Biosystems, CA). The primers and probe for the human ERV-3 gene have been described.21 The following primers and probe were used for the mouse glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene: forward primer, accacgagaaatatgacaactca; reverse primer, cccactgcctacataccatgagc; probe, 6FAM tcagcaatgcatcctgcaccaccaact TAMRA. Absolute quantification standard curves for human or mouse were plotted using DNA from human cord blood mononucleated cells and nontransplanted mouse liver, respectively. The copy numbers were calculated. All samples and standards were assayed in duplicate reactions, and averages were taken for calculation.
Reverse-Transcription PCR
Mouse livers were mechanically disassociated, and the total RNA was isolated from these cells using RNeasy kits (Qiagen, CA). First-strand complementary DNA was generated using reverse transcriptase (Invitrogen, CA) according to the manufacturer’s instructions. Complementary DNA was denatured at 94°C for 4 minutes and subsequently submitted to various PCR amplification cycles composed of 94°C for 30 seconds, 60°C for 45 seconds, and 72°C for 60 seconds. Primers for GAPDH were described.22 The primers for human α1-antitrypsin (AAT) were gctgaagaccttagtgatgc and ctttgaagtcaaggacaccg.
Results
Human ALDHhiLin− Cells Efficiently Engrafted into Damaged Mouse Livers
To assess whether human cord blood ALDHhiLin− or ALDHloLin− cells could efficiently engraft to the liver, we injected these populations into sublethally irradiated (300 cGy) NOD/SCID/MPS-VII mice via the tail vein. Because we and others have shown that CCl4-induced liver damage greatly enhances recruitment of hematopoietic stem cells to the liver,4,5,23,24 CCl4 (0.2 μL/mouse) was administered twice a week for 3 weeks to induce liver damage, starting 1 month after transplantation. This dose of CCl4 was much less than the dose used previously for NOD/SCID mice (20 μL/mouse) due to the fragility of the NOD/SCID/MPS-VII mice,4 but it still caused 67% lethality in nontransplanted mice (Fig. 1), though there were no obvious necrotic areas in the mouse liver (Fig. 2B). The lethality of mice remained at the same rate when the mice were transplanted with ALDHloLin− cells but was significantly reduced to 39% when the mice were transplanted with ALDHhiLin− cells (Fig. 1). Although proliferation and apoptosis was slightly increased in CCl4-treated mice compared with nontreated mice (Table 1), no significant increase in proliferation or decrease in apoptosis was detected in mice transplanted with ALDHhiLin− cells, in comparison to the mice that had received ALDHloLin− cells 5 days (data not shown) or 7 weeks (Table 1) prior to analysis.
Fig. 1.
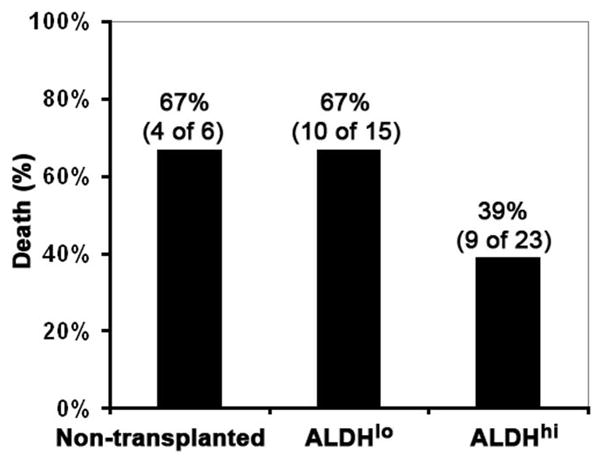
Mortality of nontransplanted mice or mice transplanted with either human ALDHloLin− or ALDHhiLin− cells after CCl4 (0.2 μL/mouse) was administered for 3 weeks.
Fig. 2.

Engraftment of human ALDHhiLin− cells into mouse livers. (A) Representative flow cytometric analysis of livers from mice transplanted with human cord blood ALDHloLin− or ALDHhiLin− cells. (B) Liver sections from transplanted mice were stained for GUSB (red) and counterstained with methyl green (green).
Table 1.
Quantification of Proliferation and Apoptotic Cells in Mouse Livers
| Mouse Group | Number of Proliferation Cells/Total Cells (%) | Number of Apoptotic Cells/Total Cells (%) |
|---|---|---|
| Goup A: transplanted with ALDHhiLin− (CCl4-treated) | ||
| No. 1 | 62/20,000 (0.31) | 297/10,000 (2.97) |
| No. 2 | 65/20,000 (0.33) | 151/10,000 (1.51) |
| No. 3 | 69/20,000 (0.35) | 300/10,000 (3) |
| No. 4 | 63/20,000 (0.32) | 380/10,000 (3.8) |
| No. 5 | 84/20,000 (0.42) | 313/10,000 (3.13) |
| Av ± SD | 68.6 ± 9 (0.34 ± 0.05) | 288 ± 84 (2.88 ± 0.84) |
| P (versus B) | 0.25 | ≤0.02 |
| P (versus C) | ≤0.001 | ≤0.003 |
| Group B: transplanted with ALDHloLin− (CCl4-treated) | ||
| No. 1 | 64/20,000 (0.32) | 120/10,000 (1.2) |
| No. 2 | 53/20,000 (0.27) | 167/10,000 (1.67) |
| No. 3 | 11/20,000 (0.06) | 155/10,000 (1.55) |
| Av ± SD | 42.7 ± 28 (0.21 ± 0.14) | 147 ± 24 (1.47 ± 0.24) |
| P (versus C) | 0.24 | ≤0.007 |
| Group C: nontransplanted (no CCl4) | ||
| No. 1 | 14/20,000 (0.07) | 65/10,000 (0.65) |
| No. 2 | 12/20,000 (0.06) | 28/10,000 (0.28) |
| No. 3 | 10/20,000 (0.05) | 41/10,000 (0.41) |
| No. 4 | 29/20,000 (0.15) | 49/10,000 (0.49) |
| Av ± SD | 16.3 ± 8.7 (0.08 ± 0.04) | 46 ± 15 (0.46 ± 0.15) |
Liver sections were stained with phosphohistone 3 antibody or terminal deoxynucleotidyl transferase–mediated dUTP nick-end labeling.
Human CD45+ cells were detected in the livers of mice transplanted with ALDHhiLin− cells (2.1 ± 1.1%) but not in mice transplanted with ALDHloLin− cells (Fig. 2A) via flow cytometric analysis. ALDHhiLin− cells also gave rise to robust hematopoietic reconstitution in bone marrow (71% ± 13.1), peripheral blood (12.8% ± 4%), and spleen (10.7% ± 8.8%), whereas ALDHloLin− cells failed to show hematopoietic engraftment (data not shown). Although mice transplanted with ALDHhiLin− cells without CCl4 treatment showed similar human CD45 cell engraftment in bone marrow (74.4 ± 7.9%), nearly one-fold less engraftment of CD45+ cells in the liver (1.2 ± 0.3%) was found in these nontreated mice.25 To detect all engrafted human cells in the recipient livers independent of their initial expression of hematopoietic stem cell surface markers, liver sections of the recipient mice were stained for their GUSB activity, absent in this mouse model but present in any human cells.14 Livers from mice transplanted with ALDHloLin− cells were completely devoid of GUSB-positive cells (Fig. 2B). In contrast, a substantial number of GUSB-positive cells were detected in the recipient livers from mice that received ALDHhiLin− cells (Fig. 2B).
Using PCR, we next detected the presence of human ALU DNA in all mice transplanted with ALDHhiLin− cells but not in nontransplanted mice or in mice transplanted with ALDHloLin− cells (Fig. 3A). Furthermore, quantitative real-time PCR measurement of the copy numbers of the human ERV-3 gene and the mouse GAPDH gene in the livers of mice transplanted with ALDHhiLin− cells showed that the percentage of human cells in these livers was between 3% and 14.2%, with an average of 8.7% (Fig. 3B). Therefore, our data demonstrated that human cord blood ALDHhiLin− cells, but not ALDHloLin− cells, can efficiently engraft recipient murine livers after damage.
Fig. 3.
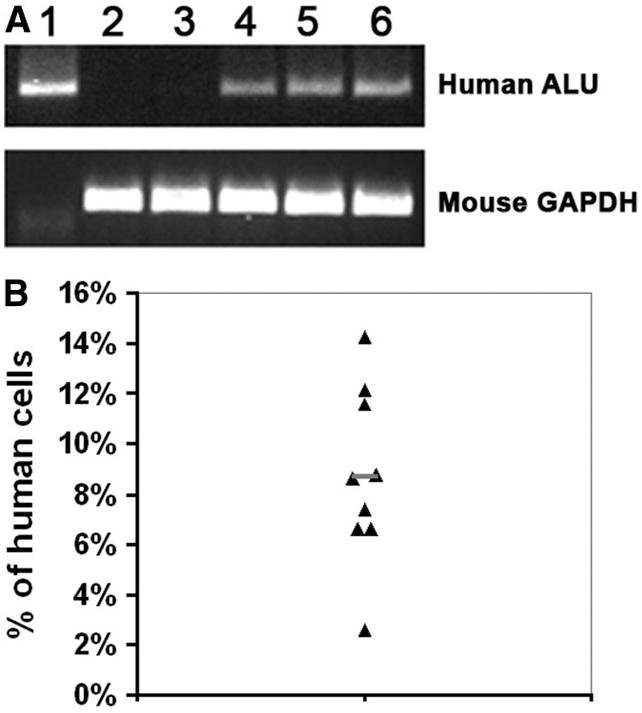
Presence of human DNA in recipient mouse livers. (A) PCR amplification of the human ALU gene and the mouse GAPDH gene in DNA samples from human mononuclear cells (lane 1), nontransplanted mice (lane 2), and mice receiving ALDHloLin− cells (lane 3) or ALDHhiLin− cells (lanes 4–6). (B) Quantitative real-time PCR analysis for the human ERV-3 gene and the mouse GAPDH gene in the recipient livers of mice transplanted with ALDHhiLin− cells. Data represent the percentage of human cells in each mouse analyzed, with the average indicated by a short line.
Human ALDHhiLin− Cells Generated Hepatocyte-Like Cells In Vivo
To test whether the engrafted human cells became liver cells, we first used reverse-transcription PCR to analyze the recipient livers for the presence of human AAT messenger RNA (mRNA), which is normally expressed in hepatocytes. We detected human AAT mRNA in the livers of all five tested mice transplanted with ALDHhiLin− cells but not in the livers of nontransplanted mice or mice transplanted with ALDHloLin− cells (Fig. 4). ALB, a marker for mature hepatocytes, was detected in the livers of mice transplanted with ALDHhiLin− cells, but not ALDHloLin− cells (Fig. 5A,C, Table 2). The frequency of human ALB+ cells in those mice was 0.05 ± 0.03%, whereas mice transplanted with the same type of cells without CCl4 treatment was lower (0.016 ± 0.01%) but not significantly so (Table 2). These ALB+ cells existed as either isolated single cells or small clusters of cells (usually 2 to 5 cells) at equivalent levels (Table 3), suggesting some of the ALB+ cells in clusters may have resulted from proliferation. We further detected that approximately 20% (6 out of 31) of the ALB-positive cells expressed GUSB (Fig. 5A,B). Those ALB+GUSB+ cells were all found in the same clusters with ALB+GUSB− cells, suggesting that many of the ALB+ cells had lost GUSB expression after proliferation. However, isolated single ALB+ cells had also lost GUSB expression. It is therefore not clear whether proliferation was required for the loss of GUSB expression in the ALB+ cells. On the other hand, most of the GUSB-positive cells did not express human ALB, suggesting that most engrafted cells were not becoming hepatocytes. The ALB+ cells present also expressed the hepatocyte-specific transcription factor HNF1,26 further proving that they were hepatocytes (Fig. 5C, Table 3). Furthermore, cells positive for both HNF1 and GUSB were also found in the recipient liver (Fig 5D). Thus, our data demonstrated that hCBSCs can give rise to hepatocyte-like cells at low levels.
Fig. 4.

Detection of human AAT mRNA in the recipient murine liver. Total RNA was isolated from HepG2 cells and livers from nontransplanted mice or mice transplanted with human cord blood ALDHloLin− or ALDHhiLin− cells. Reverse-transcription PCR was conducted for human AAT mRNA and both mouse and human GAPDH mRNA.
Fig. 5.
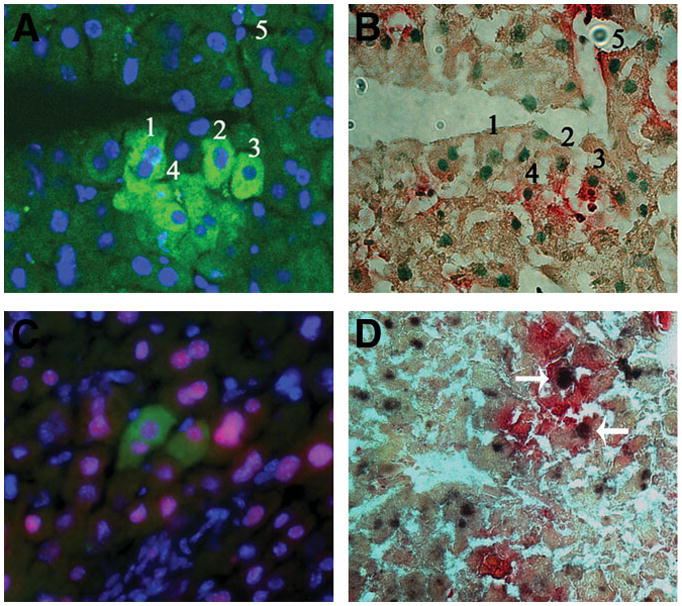
Identification of human hepatocyte-like cells in mouse livers receiving ALDHhiLin− cells. (A) Liver sections were immunostained with anti-human ALB antibody (green) and DAPI (blue). (B) These same sections were subsequently stained for GUSB activity (red) and methyl green (green nuclei). The numbers correspond to the same individual cells. (C) Liver sections were immunostained with human ALB (green), HNF1 for both mouse and human (red), and DAPI (blue). (D) Liver sections were stained for HNF1 (black) and GUSB (red). Arrows indicate HNF1+GUSB+ cells.
Table 2.
Quantification of Human ALB+ Cells in Transplanted Mouse Livers with or without CCl4 Treatment
| Mouse Group | Number of ALB+ Human | |
|---|---|---|
| Cells/Total Cells | % Human ALB+ | |
| Group A: transplanted with ALDHhiLin− (CCl4-treated) | ||
| No. 1 | 5/12,500 | 0.04% |
| No. 2 | 9/12,500 | 0.072% |
| No. 3 | 10/12,500 | 0.08% |
| No. 4 | 2/12,500 | 0.016% |
| No. 5 | 3/12,500 | 0.024% |
| Av ± SD | 5.8 ± 3.6 | 0.05 ± 0.03% |
| P (versus B) | ≤0.03 | |
| P (versus C) | 0.07 | |
| Group B: transplanted with ALDHloLin− (CCl4-treated) | ||
| No. 1 | 0/12,500 | 0% |
| No. 2 | 0/12,500 | 0% |
| No. 3 | 0/12,500 | 0% |
| No. 4 | 0/12,500 | 0% |
| No. 5 | 0/12,500 | 0% |
| Av ± SD | 0/12,500 | 0% |
| Group C: transplanted with ALDHhiLin− (no CCl4) | ||
| No. 1 | 2/12,500 | 0.016% |
| No. 2 | 0/12,500 | 0% |
| No. 3 | 3/12,500 | 0.024% |
| No. 4 | 2/12,500 | 0.016% |
| No. 5 | 3/12,500 | 0.024% |
| Av ± SD | 2 ± 1/12,500 | 0.016 ± 0.01% |
Liver sections were stained with anti-human ALB antibody.
Table 3.
Analysis of ALB+ Human Cells in Mice Transplanted with Cord Blood ALDHhiLin− Cells
| Total ALB+ | ALB+ Single | ALB+ Cluster | ALB+ HNF+ | ALB+ HNF− | ALB+ No Nucleus | |
|---|---|---|---|---|---|---|
| Number | 108 | 53 | 55 | 100 | 1 | 7 |
| Percentage | 49% | 51% | 93% | 1% | 6% |
Liver sections were immunostained with HNF1 and anti-human ALB antibodies.
Human Albumin-Expressing Cells Generated from Fusion of hCBSC-Derived Cells with Recipient Murine Hepatocytes
We next assessed whether ALDHhi-Lin− cells or their progeny directly became hepatocytes or had fused to murine hepatocytes. We used pan-centromeric probes specific for human or mouse chromosomes simultaneously to perform FISH analysis on liver sections. Images were taken using a confocal microscope. The specificity of the probes for either human or mouse was confirmed (Fig. 6A,B). Consistent with our previous data, there were significant numbers of human nuclei found in the recipient livers in mice that had received ALDHhiLin− cells (Fig. 6C). Most human cells were not fused to mouse cells. Occasionally, however, human nuclei were found to contact mouse nuclei, suggesting that fusion might occur (Fig. 6E, Supporting Video). To directly assess the type of nuclei that human ALB-expressing cells contained, the liver sections were sequentially immunostained with human ALB antibody and then subjected to FISH assay for human and mouse chromosomes. Cells without expression of human ALB primarily contained intact human nuclei, with a few exceptions where fusion between human and mouse cells could be detected (Fig. 6D,E, Supporting Video). Interestingly, the human ALB-expressing cells (more than 200 ALB+ cells were analyzed) contained mouse chromosomes with no detectable (90%) or only trace amounts (10%) of human chromosomes (Fig. 6D–F, Supporting Video). Our data suggest that hCBSCs or their progeny can fuse with host hepatocytes and that the fused cells gradually lose human genetic material after fusion.
Fig. 6.
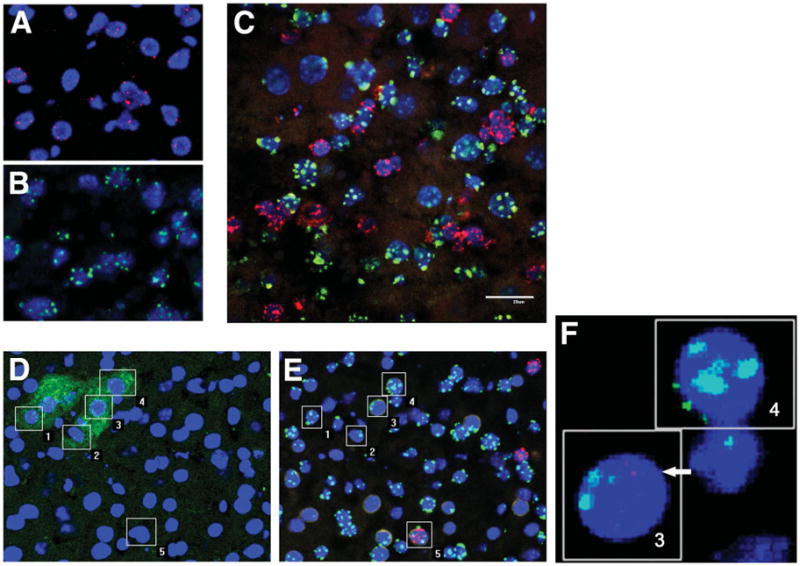
FISH analysis for human and mouse centromeres in liver sections. Liver sections from (A) human subjects, (B) nontransplanted mice, and (C) mice that had received ALDHhiLin− cells were probed with both human (red) and mouse (green) centromeric probes. (D–F) Liver sections were (D) immunostained with human ALB antibody (green) and then (E,F) probed with both human (red) and mouse (green) centromeric probes. Nuclei were counterstained with DAPI (blue) for all. The numbers correspond to the same individual cells. 1, 2, 4: Human ALB+ cells with mouse nuclei. 3: Human ALB+ cell with primarily mouse chromosomes and fragments of human chromosomes. 5: Human Alb-negative cell with both human and mouse chromosomes.
Discussion
We and others have shown that CCl4-induced liver damage greatly enhances recruitment of hematopoietic stem cells to the liver.4,5,23,24 NOD/SCID/MPSVII mice only tolerate very low doses of CCl4. This dose of CCl4 only marginally increased the proliferation and apoptosis in the mouse liver, and slightly increased the liver engraftment of donor cells. Consistently, only a small increase in donor-derived hepatocytes was observed in CCl4-treated mice compared with those in nontreated mice. ALDHhiLin− cells failed to engraft in the recipient livers when transplanted into mice preconditioned with CCl4 but without irradiation (data not shown). Similarly, Mallet et al.27 reported that Bcl-2– expressing bone marrow cells failed to generate hepatocyte-like cells when transplanted into mice without irradiation. Therefore, signals provided by irradiation (such as SDF-1 up-regulation) could be necessary to alter liver niches for donor cell recruitment. Alternatively, stem cells may need to home first to their natural niches—in this case, bone marrow—and thus provide a continuous supply of progeny to later be recruited to the damage site. When ALDHhiLin− cells are directly injected into the bloodstream, only a very limited number of progenitor cells may home to liver, thus presenting technical difficulty in detecting them. This latter explanation, but not the first one, is supported by a recent finding that fusion between blood cells and Purkinje neurons can be generated by parabiosis without irradiation, given that parabiosis also provides a constant supply of committed blood cells.28
Cell fusion has been implicated in apparent instances of cell fate switch. Derivation of hepatocyte-like cells by the fusion of a hematopoietic cell with a host hepatocyte has been demonstrated in Fah−/− mice3,8,9 and in other mouse models.10–13 However, direct transdifferentiation of BMSCs to hepatocytes has been reported by several other studies.5–7 We found many GUSB+ cells that did not express human ALB, and were likely hematopoietic. More interestingly, many of the human ALB-expressing cells lacked the expression of GUSB (Fig. 4), suggesting that only a partial human genome might be present. Furthermore, the human ALB-expressing cells contained primarily mouse chromosomes with only trace amounts of or no detectable human chromosomes. Therefore, they had arisen from fusion of human cells to host hepatocytes, and upon fusion, donor genetic material may be eliminated over time in the xenograft models. Loss of donor genetic material in fused cells has also been reported by other groups.12 However, whether the donor DNA will be lost in allografted human patients treated with cellular therapy for liver disease is not yet known.
The fusion partner from the donor side is still not completely understood. Myeloid cells or macrophages and recently lymphoid cells have been suggested to be the fusion partner.9,10,29,30 Our previous lineage analysis of ALDHhiLin−-derived cells in vivo revealed the postengraftment presence of mature myeloid cells, granulocytes, and mature B-lymphoid cells, but not T-lymphoid cells in transplanted NOD/SCID mice.17 The cord blood ALDHhi population contains rare mesenchymal stem cells (MSCs) (1 in 1500 cells [Capoccia et al, manuscript in preparation]), and it has been suggested that MSCs give rise to hepatocytes.31–33 Myeloid or lymphoid cells or their progenitors rather than MSCs are likely responsible for the observed donor-derived hepatocytes, because MSCs are rare in the ALDHhi population.
To our knowledge, this is the first time cells containing detectable human and mouse chromosomes without expression of ALB have been detected in recipient livers (Fig. 6D,E). Although our data do not exclude the possibility that cell fate switch precedes fusion, they suggest that at least some of the hematopoietic cells had fused to damaged host hepatocytes, and potentially murine hepatic-specific transcription factors activated the expression of ALB and AAT transcription from the human genes in the hybrid cells. Further studies are required to understand the nature of fusion between hematopoietic cells and hepatocytes in a system with higher frequencies of cell fusion, and to determine whether such low levels of cell fusion are safe.
Although our study sought to examine the possibility of promoting liver regeneration by hCBSC transplantation, very limited numbers (0.05%) of hepatocyte-like cells derived from hCBSCs were detected in the recipient livers, while significant numbers of donor cells, mostly hematopoietic, were engrafted. Similar low frequencies of bone marrow derived– hepatocytes have been reported by others1 and thus represent an obstacle to clinical application. However, hCBSC-derived cells may contribute indirectly to liver regeneration. Hematopoietic cells are capable of producing cytokines (such as interleukin-6, tumor necrosis factor α, hepatocyte growth factor, and others) to stimulate hepatocyte proliferation.34,35 It also has been reported that bone marrow– derived MSCs can partially revert CCl4-induced fibrosis of the mouse liver through the activation of metalloproteinase-9.36 Accordingly, we observed significantly higher levels of survival in mice that had been transplanted with human ALDHhiLin− stem cells, but not ALDHloLin− cells, which represent committed progenitors. Although we did not detect increased proliferation or decreased apoptosis in mice transplanted with ALDHhiLin− cells compared with ALDHloLin− cells at the time points taken, we cannot rule out the possibility that these effects may occur at other time points, or that the engrafted cells may enhance the function of liver in other ways.
In conclusion, we demonstrated that human cord blood ALDHhiLin− cells can efficiently engraft into the recipient livers and improve recovery of the mice from toxic insult. Donor-derived cells expressing hepatocyte-specific mRNA (AAT) and protein (ALB, HNF1) were evident in recipient livers. However, hepatocyte-like cells derived from hCBSCs were rare, and mostly, if not exclusively, arose from fusion of hCBSC-derived cells with the recipient hepatocytes. Our data suggest that hCBSCs may home to the injured liver and release trophic factors that hasten tissue repair, whereas direct contribution to hepatocyte regeneration occurs to a lesser extent.
Supplementary Material
Acknowledgments
Supported by Grant R01DK61848 from the National Institute of Diabetes and Digestive and Kidney Diseases (J. N.), Grant R01HL073256 from the National Heart, Lung, and Blood Institute (J. N.), and University of California Davis Stem Cell Program Start-up funding.
Abbreviations
- AAT
α1-antitrypsin
- ALB
albumin protein
- ALDH
aldehyde dehydrogenase
- ALDHhi
high activity of ALDH
- ALDHlo
low activity of ALDH
- BMSC
bone marrow stem cell
- DAPI
4,6-diaminidino-2-phenylindole
- FISH
fluorescence in situ hybridization
- GAPDH
glyceraldehyde 3-phosphate dehydrogenase
- GUSB
β-glucuronidase
- hCBSC
human cord blood stem cells
- HNF1
hepatocyte nuclear factor 1
- Lin−
lineage-depleted
- mRNA
messenger RNA
- MSC
mesenchymal stem cell
- NOD/SCID/MPSVII
nonobese diabetic/severe combined immunodeficient/mucopolysaccharidosis type VII
- PBS
phosphate-buffered saline
- PCR
polymerase chain reaction
Footnotes
Potential conflict of interest: Nothing to report.
Additional Supporting Information may be found in the online version of this article.
References
- 1.Thorgeirsson SS, Grisham JW. Hematopoietic cells as hepatocyte stem cells: a critical review of the evidence. Hepatology. 2006;43:2–8. doi: 10.1002/hep.21015. [DOI] [PubMed] [Google Scholar]
- 2.Lagasse E, Connors H, Al-Dhalimy M, Reitsma M, Dohse M, Osborne L, et al. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med. 2000;6:1229–1234. doi: 10.1038/81326. [DOI] [PubMed] [Google Scholar]
- 3.Vassilopoulos G, Wang PR, Russell DW. Transplanted bone marrow regenerates liver by cell fusion. Nature. 2003;422:901–904. doi: 10.1038/nature01539. [DOI] [PubMed] [Google Scholar]
- 4.Wang X, Ge S, McNamara G, Hao QL, Crooks GM, Nolta JA. Albumin-expressing hepatocyte-like cells develop in the livers of immune-deficient mice that received transplants of highly purified human hematopoietic stem cells. Blood. 2003;101:4201–4208. doi: 10.1182/blood-2002-05-1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jang YY, Collector MI, Baylin SB, Diehl AM, Sharkis SJ. Hematopoietic stem cells convert into liver cells within days without fusion. Nat Cell Biol. 2004;6:532–539. doi: 10.1038/ncb1132. [DOI] [PubMed] [Google Scholar]
- 6.Harris RG, Herzog EL, Bruscia EM, Grove JE, Van Arnam JS, Krause DS. Lack of a fusion requirement for development of bone marrow-derived epithelia. Science. 2004;305:90–93. doi: 10.1126/science.1098925. [DOI] [PubMed] [Google Scholar]
- 7.Muraca M, Ferraresso C, Vilei MT, Granato A, Quarta M, Cozzi E, et al. Liver repopulation with bone marrow derived cells improves the metabolic disorder in the Gunn rat. Gut. 2007;56:1725–1735. doi: 10.1136/gut.2007.127969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang X, Willenbring H, Akkari Y, Torimaru Y, Foster M, Al-Dhalimy M, et al. Cell fusion is the principal source of bone-marrow-derived hepatocytes. Nature. 2003;422:897–901. doi: 10.1038/nature01531. [DOI] [PubMed] [Google Scholar]
- 9.Camargo FD, Finegold M, Goodell MA. Hematopoietic myelomonocytic cells are the major source of hepatocyte fusion partners. J Clin Invest. 2004;113:1266–1270. doi: 10.1172/JCI21301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Alvarez-Dolado M, Pardal R, Garcia-Verdugo JM, Fike JR, Lee HO, Pfeffer K, et al. Fusion of bone-marrow-derived cells with Purkinje neurons, cardiomyocytes and hepatocytes. Nature. 2003;425:968–973. doi: 10.1038/nature02069. [DOI] [PubMed] [Google Scholar]
- 11.Sharma AD, Cantz T, Richter R, Eckert K, Henschler R, Wilkens L, et al. Human cord blood stem cells generate human cytokeratin 18-negative hepatocyte-like cells in injured mouse liver. Am J Pathol. 2005;167:555–564. doi: 10.1016/S0002-9440(10)62997-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kashofer K, Siapati EK, Bonnet D. In vivo formation of unstable heterokaryons after liver damage and hematopoietic stem cell/progenitor transplantation. Stem Cells. 2006;24:1104–1112. doi: 10.1634/stemcells.2005-0405. [DOI] [PubMed] [Google Scholar]
- 13.Fujino H, Hiramatsu H, Tsuchiya A, Niwa A, Noma H, Shiota M, et al. Human cord blood CD34+ cells develop into hepatocytes in the livers of NOD/SCID/gamma(c)null mice through cell fusion. FASEB J. 2007;21:3499–3510. doi: 10.1096/fj.06-6109com. [DOI] [PubMed] [Google Scholar]
- 14.Hofling AA, Vogler C, Creer MH, Sands MS. Engraftment of human CD34+ cells leads to widespread distribution of donor-derived cells and correction of tissue pathology in a novel murine xenotransplantation model of lysosomal storage disease. Blood. 2003;101:2054–2063. doi: 10.1182/blood-2002-08-2597. [DOI] [PubMed] [Google Scholar]
- 15.Jones RJ, Collector MI, Barber JP, Vala MS, Fackler MJ, May WS, et al. Characterization of mouse lymphohematopoietic stem cells lacking spleen colony-forming activity. Blood. 1996;88:487–491. [PubMed] [Google Scholar]
- 16.Storms RW, Trujillo AP, Springer JB, Shah L, Colvin OM, Ludeman SM, et al. Isolation of primitive human hematopoietic progenitors on the basis of aldehyde dehydrogenase activity. Proc Natl Acad Sci U S A. 1999;96:9118–9123. doi: 10.1073/pnas.96.16.9118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hess DA, Meyerrose TE, Wirthlin L, Craft TP, Herrbrich PE, Creer MH, et al. Functional characterization of highly purified human hematopoietic repopulating cells isolated according to aldehyde dehydrogenase activity. Blood. 2004;104:1648–1655. doi: 10.1182/blood-2004-02-0448. [DOI] [PubMed] [Google Scholar]
- 18.Hofling AA, Devine S, Vogler C, Sands MS. Human CD34+ hematopoietic progenitor cell-directed lentiviral-mediated gene therapy in a xenotransplantation model of lysosomal storage disease. Mol Ther. 2004;9:856–865. doi: 10.1016/j.ymthe.2004.03.013. [DOI] [PubMed] [Google Scholar]
- 19.Laird PW, Zijderveld A, Linders K, Rudnicki MA, Jaenisch R, Berns A. Simplified mammalian DNA isolation procedure. Nucleic Acids Res. 1991;19:4293. doi: 10.1093/nar/19.15.4293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fukuchi Y, Miyakawa Y, Kizaki M, Umezawa A, Shimamura K, Kobayashi K, et al. Human acute myeloblastic leukemia-ascites model using the human GM-CSF- and IL-3-releasing transgenic SCID mice. Ann Hematol. 1999;78:223–231. doi: 10.1007/s002770050506. [DOI] [PubMed] [Google Scholar]
- 21.Yuan CC, Miley W, Waters D. A quantification of human cells using an ERV-3 real time PCR assay. J Virol Methods. 2001;91:109–117. doi: 10.1016/s0166-0934(00)00244-5. [DOI] [PubMed] [Google Scholar]
- 22.Zhou P, Kitaura H, Teitelbaum SL, Krystal G, Ross FP, Takeshita S. SHIP1 negatively regulates proliferation of osteoclast precursors via Akt-dependent alterations in D-type cyclins and p27. J Immunol. 2006;177:8777–8784. doi: 10.4049/jimmunol.177.12.8777. [DOI] [PubMed] [Google Scholar]
- 23.Kollet O, Shivtiel S, Chen YQ, Suriawinata J, Thung SN, Dabeva MD, et al. HGF, SDF-1, and MMP-9 are involved in stress-induced human CD34+ stem cell recruitment to the liver. J Clin Invest. 2003;112:160–169. doi: 10.1172/JCI17902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Quintana-Bustamante O, Alvarez-Barrientos A, Kofman AV, Fabregat I, Bueren JA, Theise ND, et al. Hematopoietic mobilization in mice increases the presence of bone marrow-derived hepatocytes via in vivo cell fusion. Hepatology. 2006;43:108–116. doi: 10.1002/hep.21005. [DOI] [PubMed] [Google Scholar]
- 25.Hess DA, Craft TP, Wirthlin L, Hohm S, Zhou P, Eades WC, et al. Widespread nonhematopoietic tissue distribution by transplanted human progenitor cells with high aldehyde dehydrogenase activity. Stem Cells. 2008;26:611–620. doi: 10.1634/stemcells.2007-0429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Swenson ES, Guest I, Ilic Z, Mazzeo-Helgevold M, Lizardi P, Hardiman C, et al. Hepatocyte nuclear factor-1 as marker of epithelial phenotype reveals marrow-derived hepatocytes, but not duct cells, after liver injury in mice. Stem Cells. 2008;26:1768–1777. doi: 10.1634/stemcells.2008-0148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mallet VO, Mitchell C, Mezey E, Fabre M, Guidotti JE, Renia L, et al. Bone marrow transplantation in mice leads to a minor population of hepatocytes that can be selectively amplified in vivo. Hepatology. 2002;35:799–804. doi: 10.1053/jhep.2002.32530. [DOI] [PubMed] [Google Scholar]
- 28.Johansson CB, Youssef S, Koleckar K, Holbrook C, Doyonnas R, Corbel SY, et al. Extensive fusion of haematopoietic cells with Purkinje neurons in response to chronic inflammation. Nat Cell Biol. 2008;10:575–583. doi: 10.1038/ncb1720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Willenbring H, Bailey AS, Foster M, Akkari Y, Dorrell C, Olson S, et al. Myelomonocytic cells are sufficient for therapeutic cell fusion in liver. Nat Med. 2004;10:744–748. doi: 10.1038/nm1062. [DOI] [PubMed] [Google Scholar]
- 30.Nygren JM, Liuba K, Breitbach M, Stott S, Thorén L, Roell W, et al. Myeloid and lymphoid contribution to non-haematopoietic lineages through irradiation-induced heterotypic cell fusion. Nat Cell Biol. 2008;10:584–592. doi: 10.1038/ncb1721. [DOI] [PubMed] [Google Scholar]
- 31.Schwartz RE, Reyes M, Koodie L, Jiang Y, Blackstad M, Lund T, et al. Multipotent adult progenitor cells from bone marrow differentiate into functional hepatocyte-like cells. J Clin Invest. 2002;109:1291–1302. doi: 10.1172/JCI15182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sato Y, Araki H, Kato J, Nakamura K, Kawano Y, Kobune M, et al. Human mesenchymal stem cells xenografted directly to rat liver are differentiated into human hepatocytes without fusion. Blood. 2005;106:756–763. doi: 10.1182/blood-2005-02-0572. [DOI] [PubMed] [Google Scholar]
- 33.Banas A, Teratani T, Yamamoto Y, Tokuhara M, Takeshita F, Quinn G, et al. Adipose tissue-derived mesenchymal stem cells as a source of human hepatocytes. Hepatology. 2007;46:219–228. doi: 10.1002/hep.21704. [DOI] [PubMed] [Google Scholar]
- 34.Selzner N, Selzner M, Odermatt B, Tian Y, Van Rooijen N, Clavien PA. ICAM-1 triggers liver regeneration through leukocyte recruitment and Kupffer cell-dependent release of TNF-alpha/IL-6 in mice. Gastroenterology. 2003;124:692–700. doi: 10.1053/gast.2003.50098. [DOI] [PubMed] [Google Scholar]
- 35.Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology. 2006;43:S45–S53. doi: 10.1002/hep.20969. [DOI] [PubMed] [Google Scholar]
- 36.Sakaida I, Terai S, Yamamoto N, Aoyama K, Ishikawa T, Nishina H, et al. Transplantation of bone marrow cells reduces CCl4-induced liver fibrosis in mice. Hepatology. 2004;40:1304–1311. doi: 10.1002/hep.20452. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.