

1. Structure
Galectins are a family of animal lectins defined by two criteria: affinity towards β-galactosides and the presence of at least one evolutionary conserved carbohydrate-binding domain (Dumic, J. et al., 2006). They are found in all metazoans examined, from sponges and fungi, to both invertebrates and vertebrates. As many as 15 galectins have been identified to-date in mammals, and they are widely distributed among different types of cells and tissues.
Human galectin-3 is a 35-kDa protein coded by a single gene, LGALS3, located on chromosome 14. LGALS3 is composed of six exons and five introns, spanning a total of ∼17 kilobases (Fig. 1A). Exons IV to VI encode for the C-terminal domain containing the carbohydrate recognition domain (CRD). Exon III and 18 bp of exon II encode for a long and flexible N-terminal domain that contains sites for serine phosphorylation and other determinants important for the nonclassical secretion of the protein. Because of the presence of an N-terminal domain adjacent to the CRD, galectin-3 is classified as a chimera-type galectin.
Figure 1.
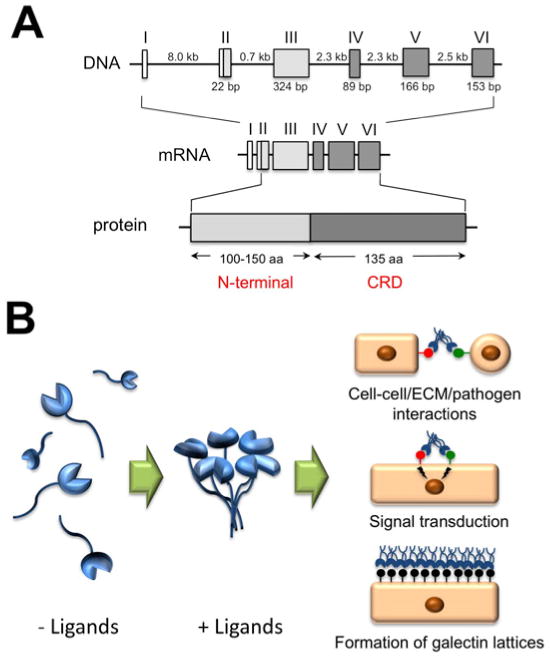
A. Genomic organization and protein structure of galectin-3. The Roman numerals indicate the position of the exons. The size of introns and polypeptide coding sequences are given in kb and bp, respectively. Galectin-3 consists of an N-terminal domain, 100-150 amino acids according to species of origin, and a C-terminal domain of about 135 amino acids, containing one carbohydrate recognition domain (CRD). B. Galectin-3 assembles in pentamer formation in the presence of multivalent ligands, in a process mediated through the N-terminal domain. At the cell surface, galectin-3 participates in cell-cell and cell-extracellular matrix (ECM) interactions, receptor clustering and signal transduction, and in the formation of glycoprotein-galectin lattices.
Galectin-3, like most members of the galectin family, binds glycoconjugates containing N-acetyllactosamine, but its affinity toward ligands is modulated by the presence of additional saccharides in proximity to the galactose residue. Differential recognition of cell surface glycans by different galectins correlates well with their distinct biological and signaling activities.
2. Function
Galectins are probably unique among all types of animal lectins in that they can be found in the nucleus, cytoplasm, cell surface, extracellular matrix, and biological fluids. Although galectin-3 exists as a monomer in solution, it can self-associate through intermolecular interactions involving the N-terminal domain when bound to a multivalent ligand and, therefore, can mediate crosslinking of glycoproteins (Fig. 1B).
The effects of galectin-3 are complex; intracellular forms typically protect cells against apoptosis through carbohydrate-independent mechanisms. Extracellularly, the lectin mediates cell–cell and cell–matrix interactions and promotes apoptosis by binding to lactosamine-containing cell surface glycoconjugates via the CRD. In recent years, several studies have revealed that galectin-3, by binding and cross-linking glycans on cell surface receptors, modulates signal transduction by novel carbohydrate-based recognition systems. For example, galectin-3: (i) promotes corneal epithelial cell migration by cross-linking complex N-glycans on α3β1 integrin, and inducing lamellipodia formation by activating the α3β1 integrin-Rac1 signaling pathway (Saravanan, C. et al., 2009); (ii) modulates VEGF- and bFGF-mediated angiogenesis by binding, via its CRD, to N-glycans on integrin αvβ3; and (iii) modulates the function of EGF and TGFβ receptors (Partridge, E.A. et al., 2004). Additionally, galectin-3, through interactions with mucin O-glycans at the apical membrane of corneal epithelial cells, forms a cell surface lattice important to the barrier function of the ocular surface (Argueso, P. et al., 2009).
In addition to cornea, galectin-3 has been detected in conjunctiva, trabecular meshwork, retina, and in the lens where it plays a role in cell differentiation and adhesion of fiber cells through interaction with MP20, a member of the tetraspanin superfamily of integral membrane proteins. Studies in non-ocular tissues have also shown that galectin-3 is expressed in inflammatory cells like monocytes, macrophages, dendritic cells, netrophils, and mast cells, and has been implicated in both innate and adaptive immune responses.
3. Disease involvement
Several studies have implicated galectin-3 in disease processes. It has been demonstrated that the rate of re-epithelialization of corneal wounds is significantly reduced in Gal3-/-mice compared to Gal3+/+ mice, and that the lectin plays a key role in integrin signaling that modulates wound closure following injury. Interestingly, the glycosyltransferases involved in the biosynthesis of galectin-3 counter receptors are altered in dry eye and wound healing, supporting the concept that disruption of galectin-ligand interactions leads to ocular surface epithelial dysfunction.
Findings that galectin-3 is an important player in VEGF and bFGF-mediated angiogenic response have broad implications and are suggestive of a putative role of galectin-3 in the pathogenesis of corneal neovascularization, corneal graft rejection, diabetic retinopathy, and wet age-related macular degeneration (AMD). Recent proteomic analyses of the Bruch membrane/choroid complex have revealed that galectin-3 is the most significantly elevated protein in advanced dry AMD, adding support to the concept that galectin-3 and its receptors, such as advanced glycation end products (AGE), contribute to AMD. Studies in animal models have shown that prevention of AGE formation or genetic deletion of galectin-3 can effectively prevent acute diabetic retinopathy, indicating that inner blood-retinal barrier dysfunction in diabetes is modulated by the presence of galectin-3.
4. Future studies
Our understanding of the roles of different members of the galectin family in the pathophysiology of the eye is in its infancy. Recent studies unraveling the role of galectins in important disease processes, such as wound healing, epithelial barrier dysfunction, infection, immune response, immune tolerance, and angiogenesis, will no doubt provide impetus to interrogate the functions of galectins in blinding diseases. Galectin-3 has been shown to regulate myofibroblast activation and fibrosis in a number of non-ocular tissues including lung, liver and kidney, but it remains to be determined whether galectin-3 modulates fibrosis and scar formation following trauma caused by surgery or disease in the eye. Also, studies aimed at determining whether exogenous galectin-3 can be used to treat patients with nonhealing epithelial defects, dry eye, and chronic inflammation are likely to prove to be rewarding. Likewise, it would be important to determine whether the complications of diabetic retinopathy and choroidal neovascularization in wet AMD can be ameliorated by specific inhibitors of galectin-3.
Acknowledgments
Supported by NIH/NEI Grant Nos. R01EY014847 (to PA), R01EY07088 (to NP), Massachusetts Lions Eye Research fund, and a Research to Prevent Blindness unrestricted award to the New England Eye Center. We apologize to authors whose work cannot be cited due to limits for reference citation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Argueso P, Guzman-Aranguez A, Mantelli F, Cao Z, Ricciuto J, Panjwani N. Association of cell surface mucins with galectin-3 contributes to the ocular surface epithelial barrier. J Biol Chem. 2009;284:23037–23045. doi: 10.1074/jbc.M109.033332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumic J, Dabelic S, Flogel M. Galectin-3: an open-ended story. Biochim Biophys Acta. 2006;1760:616–635. doi: 10.1016/j.bbagen.2005.12.020. [DOI] [PubMed] [Google Scholar]
- Partridge EA, Le Roy C, Di Guglielmo GM, Pawling J, Cheung P, Granovsky M, Nabi IR, Wrana JL, Dennis JW. Regulation of cytokine receptors by Golgi N-glycan processing and endocytosis. Science. 2004;306:120–124. doi: 10.1126/science.1102109. [DOI] [PubMed] [Google Scholar]
- Saravanan C, Liu FT, Gipson IK, Panjwani N. Galectin-3 promotes lamellipodia formation in epithelial cells by interacting with complex N-glycans on alpha3beta1 integrin. J Cell Sci. 2009;122:3684–3693. doi: 10.1242/jcs.045674. [DOI] [PMC free article] [PubMed] [Google Scholar]