

Abstract
The generation of insulin-producing β-cells from human pluripotent stem cells is dependent on efficient endoderm induction and appropriate patterning and specification of this germ layer to a pancreatic fate. In this study, we elucidated the temporal requirements for TGFβ family members and canonical WNT signaling at these developmental stages and show that the duration of nodal/activin A signaling plays a pivotal role in establishing an appropriate definitive endoderm population for specification to the pancreatic lineage. WNT signaling was found to induce a posterior endoderm fate and at optimal concentrations enhanced the development of pancreatic lineage cells. Inhibition of the BMP signaling pathway at specific stages was essential for the generation of insulin-expressing cells and the extent of BMP inhibition required varied widely among the cell lines tested. Optimal stage-specific manipulation of these pathways resulted in a striking 250-fold increase in the levels of insulin expression and yielded populations containing up to 25% C-peptide+ cells.
Keywords: Endoderm differentiation, Human embryonic stem cells, Human induced pluripotent stem cells, β-Cell, Pancreas
INTRODUCTION
Modeling embryonic development in pluripotent stem cell (PSC) differentiation cultures has proven to be the most effective approach for the efficient generation of differentiated cells types in vitro (Murry and Keller, 2008). For pancreatic development, the crucial steps that need to be accurately modeled include the induction of definitive endoderm, the patterning and specification of endoderm to a pancreatic fate and the generation of mature endocrine/exocrine cells (Wells and Melton, 1999). Studies using different model systems have shown that induction of definitive endoderm is dependent on signaling through the nodal pathway (Conlon et al., 1994; Feldman et al., 1998; Gritsman et al., 2000; Jones et al., 1995; Lowe et al., 2001; Osada and Wright, 1999; Zhou et al., 1993). Once formed, definitive endoderm generates the gut tube, which is patterned into anterior and posterior fates by gradients of WNT, FGF and retinoic acid (RA) signaling (Apelqvist et al., 1999; Chung et al., 2010; Chung et al., 2008; Hebrok, 2003; Hebrok et al., 1998; Martin et al., 2005; Molotkov et al., 2005; Stafford and Prince, 2002; Wandzioch and Zaret, 2009). Once the pancreatic program has been established, signaling by TGFβ superfamily members appears to be dispensable as mice lacking functional Smad4 in the pancreatic epithelium generate a normal pancreas (Bardeesy et al., 2006).
By manipulating key pathways at specific stages, D'Amour et al. demonstrated that human embryonic stem cells (hESCs) could be differentiated along the pancreatic lineage to generate insulin-producing cells (D'Amour et al., 2006). These findings reinforced the importance of translating developmental biology to ESC differentiation and clearly demonstrated the potential to generate pancreatic derivatives from hESCs. Although the study represented an important step forward, the strategy outlined was not broadly applicable as the efficiency of differentiation for the different hESC lines tested varied widely. Modifications of this approach have been used by others to generate populations with similar characteristics from both hESCs and human induced pluripotent stem cells (hiPSCs) (Cai et al., 2009; Chen et al., 2009; Jiang, J. et al., 2007; Jiang, W. et al., 2007; Maehr et al., 2009; Mfopou et al., 2010; Tateishi et al., 2008; Zhang et al., 2009). The benefit of these modifications is, however, often difficult to assess, as quantification of end-stage populations and comparison of the differentiation status of the PSC-derived populations to fetal or adult pancreatic tissue often are not reported.
The low efficiency of differentiation and cell line variability have limited access to end-stage insulin-producing cells and, as a consequence, have hampered progress in characterizing these cells for their developmental status and functional potential. The suboptimal production of insulin+ cells in the differentiation cultures strongly suggests that key steps along the developmental pathway have not been optimized. To address this issue, we mapped the development of the pancreatic lineage from different PSCs in detail using clues from developmental biology to inform us on the optimal signaling pathways required to induce, pattern and specify definitive endoderm to a pancreatic fate. Here, we show that optimal induction of definitive endoderm, combined with appropriate anterior-posterior patterning and inhibition of TGFβ signaling enables the efficient differentiation of hPSCs to a pancreatic fate. End-stage populations generated through this approach consist of up to 25% C-peptide+ cells and display INS levels that approximate those found in the adult pancreas.
MATERIALS AND METHODS
HPSC culture and differentiation
38-2 hiPSC line was derived from fibroblasts (GM01390, Coriell Institute for Medical Research, NJ, USA) by using a novel pEYK3.1 retroviral vector containing four reprogramming factors [OCT4 (POU5F1 – Human Gene Nomenclature Database), SOX2, KLF4, MYC] in one backbone and removing transgenes by Cre recombinase (Koh et al., 2002) (I.-H.P. and G.Q.D., unpublished). INSGFP/w hESCs were generated by targeting sequences encoding GFP to the insulin locus of HES3 cells using previously described protocols (Costa et al., 2007) (S.J.M., X.L., A.G.E. and E.G.S., unpublished). HPSCs were maintained as described (Kennedy et al., 2007). Embryoid bodies (EBs) were cultured in low-cluster plates (Corning) in serum free differentiation (SFD) media (Gouon-Evans et al., 2006). BMP4 (10 ng/ml) was added for the first day of differentiation (d0-d1). At day 1, EBs were harvested and resuspended in medium A: basic fibroblast growth factor (bFGF; 2.5 ng/ml), activin A (100 ng/ml) and BMP4 (0.25 ng/ml). The medium was changed on day 4 and was supplemented with vascular endothelial growth factor (VEGF; 20 ng/ml) and bFGF (5 ng/ml) (medium B). EBs were harvested at day 5, dissociated to single cells and plated on gelatin-coated dishes at 4×105 cells/ml in media B with activin A at 50 ng/ml. Stage 2 medium consisted of SFD supplemented with FGF10 (50 ng/ml) in combination with the indicated concentrations of mouse Wnt3a, DKK1, dorsomorphin (Sigma) and noggin. Stage 3 medium consisted of Dulbecco's modified Eagle's medium (DMEM) with 1% vol/vol B27 supplement (Invitrogen), ascorbic acid (50 μg/ml; Sigma), KAAD-cyclopamine (0.25 μM; Toronto Research Chemicals, ON, Canada), all-trans retinoic acid (2 μM) (Sigma), noggin (50 ng/ml) and FGF10 (50 ng/ml). Stage 4 medium consisted of DMEM with 1% vol/vol B27 supplement, ascorbic acid (50 μg/ml), SB431542 (6 μM; Sigma) and noggin (50 ng/ml). Stage 5 medium consisted of SFD supplemented with d-glucose (40 mM; Sigma), SB431542 (6 μM), noggin (50 ng/ml) and the γ-secretase inhibitor L-685,458 (10 μM; Tocris). All cytokines were human and purchased from R&D Systems, unless stated otherwise. Cultures were maintained in a 5% CO2, 5% O2, 90% N2 environment. Monolayer differentiation was carried out as described previously (D'Amour et al., 2006). Fetal calf serum (FCS) was replaced by the cytokines present in medium B. Cultures differentiated in monolayer were maintained in a 5% CO2 air environment.
Flow cytometry and cell sorting
EB and monolayer cultures generated from hPSC differentiation experiments were dissociated with 0.25% trypsin in EDTA. Day 0-5 EB- and d3 monolayer-induced cells were stained with anti-CXCR4-phycoerythrin (BD; 1:100), anti-CD31-phycoerythrin (BD; 1:10), anti-CD117-allophycocyanin (Invitrogen; 1:100) and anti-KDR–allophycocyanin (R&D Systems; 1:10). Day 20-25 cells were stained with HPi3 (1:20), HPα2 (1:20), HPx1 (1:20), HPd1 (1:20) (Dorrell et al., 2008) and anti-mouse IgG-phycoerythrin (Jackson ImmunoResearch; 1:200). Intracellular antigens were detected by staining with goat anti-human SOX17 (R&D Systems; 1:40), goat anti-FOXA2 (clone M20, Santa Cruz; 1:50), rat anti-human C-peptide (AB1921, Beta Cell Biology Consortium; 1:300), mouse anti-GCG (Sigma; 1:500), donkey anti-goat IgG-Alexa 488 (Invitrogen; 1:400), donkey anti-rat IgG-Alexa 488 (Invitrogen; 1:400), goat anti-mouse allophycocyanin (BD; 1:200). For cell surface markers, staining was carried out in PBS with 10% FCS. For intracellular proteins, staining was carried out on cells fixed with 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA, USA) in PBS. Cells were permeabilized with 90% ice-cold methanol for 20 minutes for SOX17 and FOXA2 staining as previously described (Krutzik and Nolan, 2003). C-peptide staining was performed in PBS with 10% FCS and 0.5% saponin (Sigma). Stained cells were analyzed using an LSRII flow cytometer (BD). The cells were sorted using a FACSAria™II (BD) cell sorter (SickKids-UHN Flow Cytometry Facility, Toronto, ON, Canada). Data were analyzed using FlowJo software (Treestar, Ashland, OR, USA).
Immunostaining
Immunostaining was performed as described previously (Gouon-Evans et al., 2006) using the following antibodies: goat anti-FOXA2 (M20, Santa Cruz; 1:200), goat anti-PDX1 (gift of Dr C. Wright, Vanderbilt University, Nashville, TN, USA; 1:10,000), rat anti-human C-peptide (AB1921, Beta Cell Biology Consortium; 1:1000), goat anti-human glucagon (C-18, Santa Cruz; 1:500), mouse anti-SST (AB1985, Beta Cell Biology Consortium; 1:500), goat IgG (Sigma), mouse, rabbit or rat IgG (Jackson ImmunoResearch); concentrations of isotype controls were matched to primary antibodies. Secondary antibodies used were: goat anti-mouse IgG-Cy3 (Jackson ImmunoResearch; 1:400), donkey anti-rat IgG-Cy3 (Jackson ImmunoResearch; 1:400), donkey anti-goat IgG-Alexa 488 (Invitrogen; 1:400), donkey anti-rabbit IgG-Alexa 488 (Invitrogen; 1:400), rabbit anti-mouse Alexa 350 (Invitrogen; 1:200). DAPI was used to counterstain nuclei. The stained cells were visualized using a fluorescence microscope (Leica CTR6000) and images captured using the Leica Application Suite software.
Immunoblot analysis
Immunoblots were prepared as described previously (Lazzara et al., 2010). Primary antibodies anti-p-SMAD2, anti-p-SMAD1/5/8 and anti-β-ACTIN (ACTB – Human Gene Nomenclature Database) were purchased from Cell Signaling Technology. Secondary antibodies, IR 680-labeled anti-mouse IgG and IR 800-labeled goat anti-rabbit IgG (Rockland, Gilbertsville, PA, USA) were detected using an Odyssey reader (LI-COR, Lincoln, NE, USA).
Quantitative real-time PCR
Total RNA was prepared with the RNAqueous-Micro Kit (Ambion) and treated with RNase-free DNase (Ambion). 500 ng to 1 μg RNA was reverse transcribed into cDNA using random hexamers and Oligo (dT) with Superscript III Reverse Transcriptase (Invitrogen). QPCR was performed on a MasterCycler EP RealPlex (Eppendorf) using QuantiFast SYBR Green PCR Kit (Qiagen) as described previously (Nostro et al., 2008). Expression levels were normalized to the housekeeping gene TATA box binding protein (TBP). The oligonucleotide sequences are available on request. Three different lots of total human adult pancreas RNA were purchased from Clontech.
RESULTS
HESC-derived endoderm progenitors generated as embryoid bodies give rise to insulin-expressing cells
To generate definitive endoderm, we induced the HES2 hESC line with activin A (hereafter referred to as activin) using an embryoid body (EB) format. As shown previously with mouse ESCs (Gadue et al., 2006; Gouon-Evans et al., 2006), the emergence of definitive endoderm can be monitored by co-expression of CXCR4 and CD117 (KIT – Human Gene Nomenclature Database). CXCR4+ CD117+ cells were detected as early as day 3 of differentiation (not shown) in the HES2 cultures and by day 5, greater than 90% of cells (n=20) expressed these markers (Fig. 1A; see Fig. S1A in the supplementary material). More than 70% of day 5 EBs expressed SOX17 (74±6, n=4; Fig. 1A; see Fig. S1A in the supplementary material), suggesting that they were enriched for endoderm progenitors. Virtually no KDR+ or CD31 (PECAM1 – Human Gene Nomenclature Database)+ cells were detected in activin-induced EBs (not shown), indicating little contamination with mesoderm (Matthews et al., 1991; Vecchi et al., 1994). Molecular analyses confirmed the flow-cytometric data, showing that EBs expressed the endodermal genes FOXA2 and SOX17 (Fig. 1B) and only low levels of the pluripotency marker OCT4 (Nichols et al., 1998), the ectodermal gene SOX3 (Fig. 1B) (Collignon et al., 1996; Wood and Episkopou, 1999) and the mesodermal genes MEOX1 and MESP1 (see Fig. S1B in the supplementary material), (Candia et al., 1992; Saga et al., 1996). EBs cultured without activin contained few CXCR4+ CD117+ SOX17+ cells (see Fig. S1A in the supplementary material).
Fig. 1.

Endoderm induction in human embryonic stem cell (hESC)-derived embryoid bodies (EBs). (A) Flow-cytometric analysis of CXCR4, CD117 and SOX17 on hESCs [differentiation day (d)0] and d5 EB-derived cells differentiated in the presence of activin (d5+ACT) in serum free media demonstrating efficient induction of endoderm cells using the embryoid body, serum-free differentiation system. Percentage of cells within each quadrant is indicated. (B) Quantitative PCR (QPCR) analysis of OCT4, SOX3, FOXA2 and SOX17 in d0 and d5 populations. Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test. P=0.033 (OCT4), P=0.025 (SOX3), P=0.081 (FOXA2) and P=0.030 (SOX17). n=3.
Most previous studies induced endoderm from hPSCs as monolayers, using the protocol described by D'Amour et al. (Ameri et al., 2010; Chen et al., 2009; D'Amour et al., 2006; Johannesson et al., 2009; Maehr et al., 2009; Mfopou et al., 2010). A comparison of the two approaches revealed that induction in monolayers is faster than in EBs as the CXCR4+ CD117+ population emerged by day 3 of differentiation (see Fig. S2A in the supplementary material). Comparable kinetics were observed in the monolayer cultures if serum was substituted with BMP4, bFGF and VEGF during the induction step, demonstrating that monolayer differentiation can be achieved under serum-free conditions (see Fig. S2A in the supplementary material). OCT4, SOX3, FOXA2, SOX17, HEX (HHEX – Human Gene Nomenclature Database), HNF1B and HNF6 (ONECUT1 – Human Gene Nomenclature Database) expression was similar in d5 EBs [day 5 EB + growth factors (GF)] and d3 monolayers (day 3 monolayer (M) + GF and day 3 M + FCS), whereas HNF4A expression was higher in d5 EBs compared with d3 monolayers (see Fig. S2B in the supplementary material). Taken together, these results demonstrate that the CXCR4+ CD117+ population generated under serum-free conditions in EBs is comparable to that induced in monolayer serum-containing cultures. Although the monolayer cultures promote differentiation faster, the EB system offers the advantage of scalability required for the generation of large number of cells.
To verify that the EB-derived CXCR4+ CD117+ cells could differentiate to a pancreatic fate, we adopted a modified version of the protocol described by D'Amour et al. for the generation of insulin-producing cells (D'Amour et al., 2006). EBs (d5) were dissociated and cells were plated on gelatin-coated dishes and cultured for 20 days under the following conditions (Fig. 2A): (1) FGF10 for 3 days to generate the equivalent of the primitive gut tube; (2) noggin, KAAD-cyclopamine and retinoic acid (NCR) for 3 days to generate PDX1+ pancreatic progenitors; and (3) DMEM supplemented with B27 in the absence of added factors (to day 25) to promote maturation to pancreatic endocrine lineage cells. With this protocol, a large proportion of the population (~80%) retained FOXA2 expression over 20 days (Fig. 2B,C). NCR treatment induced the development of PDX1+ cells, which emerged as distinct clusters throughout the culture (Fig. 2D). Small numbers of C-peptide+ cells were routinely detected by day 25 of culture (Fig. 2E).
Fig. 2.

Generation of pancreatic cells from embryoid body (EB)-derived endoderm. (A) Schematic representation of the differentiation protocol. EBs were trypsinized at differentiation day (d)5 and plated as a monolayer in the presence of FGF10 in SFD media for three days to generate the equivalent of the primitive gut tube. Pancreatic endoderm was subsequently induced by treatment with noggin, KAAD-cyclopamine and retinoic acid (NCR) for 3 days in DMEM supplemented with B27. Cells were then cultured in DMEM supplemented with B27 up to d25 to generate endocrine cells. (B) Intracellular flow-cytometric analysis of FOXA2 at days 8, 11, 13 and 20 of differentiation showing maintenance of FOXA2 expression during this time. Percentage of cells within each quadrant is indicated. (C-E) Immunostaining of FOXA2 at d11, PDX1 at d13 and C-peptide at d25 of differentiation. (F) Quantitative PCR (QPCR) analysis of INS and ALB at d22 in cultures treated with or without noggin at the NCR stage (d8-d10). INS levels are compared with adult pancreas. Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test. P=0.025 (INS), P=0.03 (ALB). n=3.
Although previous studies have included the BMP inhibitor noggin during the induction of PDX1, the extent to which BMP inhibition impacts endocrine lineage development has not been established (Cai et al., 2009; Jiang, J. et al., 2007; Kroon et al., 2008; Mfopou et al., 2010; Tateishi et al., 2008; Zhang et al., 2009). We found that BMP inhibition was essential for pancreatic development because without it C-peptide+ cells were not detected (not shown) and the insulin (INS) levels were significantly diminished (Fig. 2F). The untreated cultures showed increased albumin (ALB) expression, suggesting that BMP signaling at this stage plays a key role in determining whether the cells adopt a ventral pancreatic or hepatic fate (Fig. 2F). These observations are consistent with findings from previous studies, which indicate that BMP signaling regulates hepatic specification from ventral foregut endoderm (Cai et al., 2009; Mfopou et al., 2010). Whereas noggin promoted pancreatic development, INS levels in end-stage cultures were ~300 times lower than those found in the adult pancreas (Fig. 2F).
Definitive endoderm specification and patterning
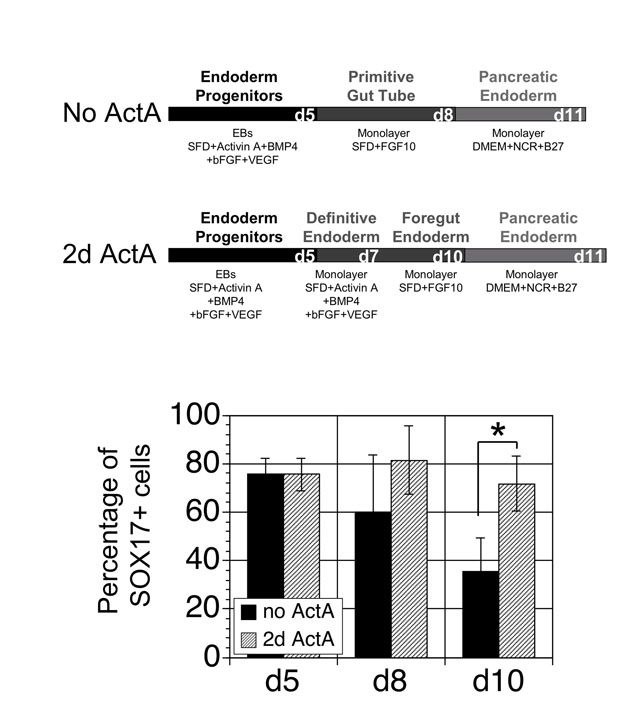
The small numbers of C-peptide+ cells and the low INS levels detected suggested that crucial aspects of pancreatic development have not been reproduced in our culture system. We therefore investigated the effect of manipulating specific signaling pathways at different developmental stages, focusing initially on the CXCR4+ CD117+ endoderm progenitor population (d5). As the duration of activin/nodal signaling plays a pivotal role in the generation of definitive endoderm in the mouse ESC model (Gadue et al., 2009; Gadue et al., 2006), we investigated whether or not extended signaling through this pathway impacts endoderm induction and pancreatic development in the hESC differentiation cultures (see Fig. S3 in the supplementary material). Support for this approach is provided by findings from an independent study that showed that treatment of day 5 CXCR4+ CD117+ cells with activin for an additional two days resulted in enhanced hepatic development (d10) (S.O. and G.K., unpublished). To determine if this manipulation improves pancreatic differentiation, we treated CXCR4+ CD117+ cells with activin for two days prior to FGF10 treatment. Although the additional activin step led to a significant increase in the proportion of SOX17+ cells detected at day 10 of differentiation (see Fig. S3 in the supplementary material; P=0.003, n=3), it did not result in an increase in the size of the C-peptide+ population at day 25 under these conditions. The fact that these early manipulations did not enhance development of the insulin-producing cell population suggests either that they are not relevant to pancreatic development or that additional steps regulating later crucial stages of development are not optimized.
Inhibition of TGFβ signaling is essential for endocrine lineage commitment
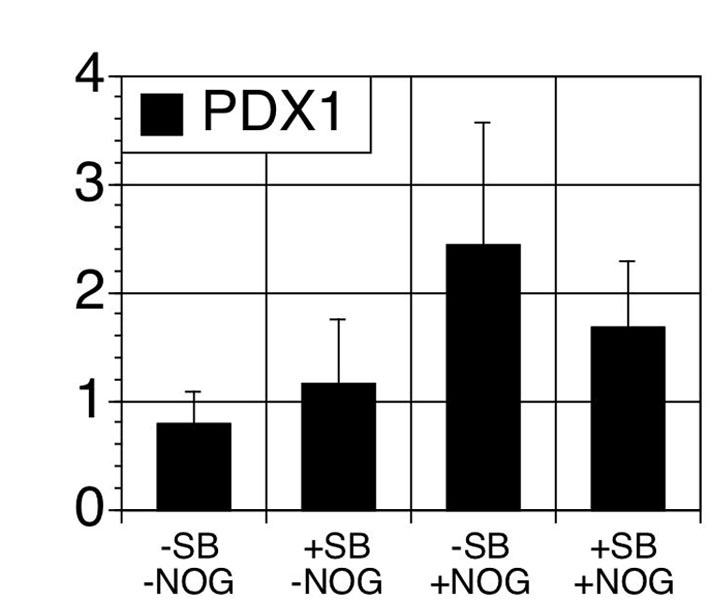
To address this issue, we focused our efforts on promoting the commitment of the PDX1 progenitor population to the endocrine lineage, as most protocols have not manipulated signaling pathways at this transition. The observation that inhibition of BMP signaling at the NCR step is essential for pancreatic specification (Fig. 2F) and the fact that signaling through TGFβ superfamily members is not required for pancreatic development following the emergence of the PDX1+ population (Bardeesy et al., 2006) suggested that stage-specific inhibition of these pathways might be important for endocrine commitment. To investigate this possibility, we inhibited the TGFβ/activin/nodal and BMP pathways by adding the small molecule ALK4/5/7 inhibitor SB431542 (SB) (Inman et al., 2002) and noggin immediately following PDX1 induction (days 13-17). The effectiveness of the inhibitors was evaluated by immunoblot analysis one day after treatment. In the absence of inhibitors, phosphorylated SMAD2 and SMAD1/5/8 were detected (Fig. 3A,B), indicating signaling by endogenous TGFβ superfamily members. Treatment with SB and noggin blocked SMAD2 and SMAD1/5/8 phosphorylation, respectively (Fig. 3A,B). As expected, the addition of both inhibitors simultaneously blocked both SMAD2 and SMAD1/5/8 phosphorylation.
Fig. 3.

Inhibition of TGFβ signaling is essential for endocrine lineage commitment. (A) Western blot analysis showing endogenous SMAD2 and SMAD1/5/8 phosphorylation at day 14 of cultures (−SB–NOG, lane 1). Treatment with SB431542 (SB; 6 μM) at day 13 inhibits SMAD2 phosphorylation (+SB, lane 2). Treatment with noggin (NOG; 50 ng/ml) at day 13 inhibits SMAD1/5/8 phosphorylation (+NOG, lane 3). Treatment with SB and noggin at day 13 inhibits SMAD2 and SMAD1/5/8 phosphorylation (+SB+NOG, lane 4). (B) Phospho-SMAD1/5/8 and SMAD2 normalized to β-ACTIN. Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test. Treatment with SB (± NOG) inhibits SMAD2 phosphorylation when compared with −SB-NOG (**P=0.0023). Treatment with SB+NOG inhibits SMAD1/5/8 phosphorylation when compared with −SB-NOG (*P=0.018). n=3. (C) Treatment with SB increases cell yield at day 17 of differentiation. Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test. P=0.002 (+SB) and P=0.001 (+SB+NOG) compared with −SB–NOG treated group. n=4 for −SB–NOG and +SB–NOG, n=5 for −SB+NOG, n=6 for SB+NOG group. (D) QPCR analysis for INS at day 22 and day 25 of monolayer and embryoid body (EB) differentiation, respectively. Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test of NOG treated groups compared with untreated group (−S-N). P<0.05. n=3. (E) Immunostaining for C-peptide at day 17 in the different treatment groups in EB-derived populations.
Four days following addition of inhibitors (day 17), cultures treated with SB consistently contained higher numbers of cells compared with non-treated control cultures or cultures treated with noggin alone (Fig. 3C). Molecular analysis revealed that treatment with noggin alone resulted in a significant (threefold) increase in INS expression measured on day 22 of culture. The combination of noggin and SB had an additive effect, resulting in a sixfold increase in INS expression over that observed in untreated cultures, although no difference was observed in PDX1 expression levels (Fig. 3D; see Fig. S4 in the supplementary material). Consistent with the expression analysis, immunostaining revealed larger C-peptide+ clusters in the cells treated with noggin alone or the combination of noggin and SB (Fig. 3E). The positive effects of noggin and SB were even more dramatic in cells induced in monolayer culture as the addition of both antagonists resulted in a 27-fold increase in INS expression. Taken together, these findings indicate that inhibition of TGFβ/activin/nodal and BMP signaling following induction of pancreatic progenitors does promote differentiation to the endocrine lineage.
As INS levels following these manipulations were significantly higher than those measured in our earlier studies, we re-evaluated the importance of the extended activin treatment under these conditions in cultures generated from EBs. Activin treatment had a dramatic effect, as CXCR4+ CD117+ cells treated for 2 days (d5-7) generated populations that expressed almost 15-fold more PDX1 and 67-fold more INS compared with non-treated populations (Fig. 4A,B). Populations generated from CXCR4+ CD117+ cells treated for 3 days expressed less INS than those from a 2-day treatment suggesting that the duration of activin/nodal signaling is crucial for generating the appropriate endoderm population (Fig. 4A,B).
Fig. 4.

Patterning with activin and Wnt3a is required for optimal pancreatic specification. (A,B) Quantitative PCR (QPCR) analysis evaluating PDX1 and INS expression at different stages of differentiation in cultures generated from populations treated without activin (No ACT) at day 5, with activin for 2 days (d5-d7; 2dACT) and with activin for three days (d5-d8; 3dACT). Cultures were treated with SB431542 (SB) and noggin (NOG) following NCR treatment and were terminated at day 15, 17 and 18 for No Act, 2dAct and 3dAct, respectively. Expression levels were normalized to TBP and are relative to the 2dACT sample at d17 (set at 100). Bars represent mean±s.d. Asterisks indicate statistical significance as determined by t-test. P=0.032 (PDX1) and P=0.027 (INS). n=3. (C,D) Day 17 cultures were analyzed by qPCR for the expression of intestinal (CDX2) and pancreatic (INS) genes following treatment with DKK1 or Wnt3a. Bars represent mean±s.d. Cultures were treated between days 7 and 9 with FGF10 (F) or with a combination of FGF10 and one of the following: DKK1 at 150 ng/ml (F+DKK), Wnt3a at 1 ng/ml (F+W1), Wnt3a at 3 ng/ml (F+W3), Wnt3a at 9 ng/ml (F+W9). CDX2 expression was significantly downregulated in F+DKK, INS expression was significantly upregulated in F+W3. *P<0.05, **P<0.01 determined by ANOVA with Tukey's HSD test. n=4.
Patterning by WNT enhances endocrine development
Patterning the appropriate endoderm population is a crucial step in the generation of derivative lineages and functional cell populations. As the pancreas develops from the foregut/midgut junction of the gut tube, it is important to identify conditions that promote the development of this population. Most protocols use FGF10 or KGF to pattern the activin-induced population to the posterior foregut fate. To determine whether additional morphogens would enhance pancreatic development, we examined the consequences of manipulating the canonical WNT pathway at this stage, as studies in Xenopus have shown that this pathway plays a pivotal role in establishing anterior-posterior fate (McLin et al., 2007). For these studies, day 7 cultures were treated for 3 days with FGF10 together with either DKK1 to block endogenous WNT signaling or with different concentrations of Wnt3a to activate the pathway. The cells were then induced with NCR for 3 days, cultured for an additional 4 days to promote maturation and then evaluated for expression of genes indicative of pancreatic (PDX1 and INS) and intestinal (CDX2) development.
Inhibition of WNT signaling with DKK1 significantly reduced the expression of the intestinal gene CDX2 (Fig. 4C). WNT signaling did not affect the levels of PDX1 (not shown), but did increase INS expression fourfold compared with control cultures and 15-fold compared with those treated with DKK1 (Fig. 4D). The positive effects on INS expression were lost with increasing concentration of agonist (9 ng/ml), indicating that optimal levels of WNT signaling were required for patterning the equivalent of the foregut/midgut population.
Kinetics of pancreatic lineage development
To evaluate further the impact of manipulating the TGFβ and WNT signaling pathways on the efficiency of pancreatic differentiation, we compared cells differentiated under these conditions with those differentiated using the modified version of the original protocol described by D'Amour (D'Amour et al., 2006) that included the addition of noggin during stage 3 (Kroon et al., 2008). For these studies HES2 hESCs were differentiated in monolayer culture using the following three sets of conditions (see Fig. S5 in the supplementary material): (1) the D'Amour/Kroon protocol (protocol 1), (2) our protocol (protocol 2) and (3) our protocol with 2 days extended activin (protocol 2+act). The extended activin protocol was included to determine if the optimal time for activin/nodal signaling was influenced by the culture format (EB versus monolayer). Cells were harvested at the end of each stage as defined by D'Amour (D'Amour et al., 2006) and analyzed for expression of endoderm and pancreatic lineage genes. As shown in Fig. 5A, genes indicative of endoderm induction (FOXA2, SOX17), posterior foregut specification (HNF1B) and pancreatic commitment (PDX1) were induced at comparable times and at similar levels under the three conditions. Following induction of pancreatic fate by NCR, however, striking differences in expression were observed. NGN3 (NEUROG3 – Human Gene Nomenclature Database), NEUROD1, INS and glucagon (GCG) were all expressed at higher levels in cells induced with protocol 2 (without extended activin) compared with those induced with protocol 1. Whereas the stage 4 and 5 populations induced with protocol 1 expressed very low levels of INS, those generated with protocol 2 expressed levels similar to that found in the adult pancreas. In contrast to INS and GCG, somatostatin (SST) was expressed at comparable levels in stage populations induced by both protocols. Interestingly, the 2 day extended activin treatment in the monolayer cultures was not beneficial as these populations (protocol 2+act) expressed lower levels of these genes than was found in cells that did not receive the additional induction.
Fig. 5.

Kinetics of pancreatic lineage development. (A) Quantitative PCR (QPCR) analysis for FOXA2, SOX17, HNF1B, NGN3, NEUROD1, GCG, INS and SST in the adult pancreas (P) and at stages 1 to 5 (S1-S5) following differentiation with three different protocols: the D'Amour protocol (D'Amour et al., 2006) supplemented with noggin at stage 3 (protocol 1) and our protocol (protocol 2) with or without extended activin (Act). The protocols are depicted in Fig. S5 in the supplementary material. Expression levels are normalized to TBP. Bars represent mean±s.d. *P<0.05, **P<0.01 determined by ANOVA with Tukey's HSD test. (B) Intracellular flow-cytometric analysis of C-peptide and GCG at days 14, 20 and 22 of differentiation for protocol 1, 2 and 2+ACT, respectively. **P<0.01 determined by ANOVA with Tukey's HSD test. (C) Representative flow cytometric analysis of C-peptide and GCG at day 14 and 20 of differentiation using protocol 1 and protocol 2 showing the higher efficiency of protocol 2 in inducing endocrine cells as compared with protocol 1. Percentage of cells within each quadrant is indicated.
As expected from the gene expression profiles, flow cytometric analyses revealed dramatic differences in the proportion of C-peptide cells generated at the end stage cultures under the three different sets of conditions. With monolayer induction in the absence of extended activin, we were able to routinely generate populations that contained 25% C-peptide+ cells by day 20 of culture. The extended activin treatment reduced this to 11% C-peptide+ cells, whereas populations generated with protocol 1 contained ~5% of these cells (Fig. 5B). Approximately 50% of the C-peptide+ cells co-expressed GCG, consistent with observations from others that at least a portion of the endocrine cells generated in these cultures are polyhormonal, possibly reflecting an immature stage of development. Although slightly more than 50% of the C-peptide cells (56±14%) that developed at this stage in our culture did not express GCG, they were not glucose-responsive (Fig. 5B,C; see Fig. S6C in the supplementary material). While the population generated using protocol 1 did not contain many C-peptide+ cells, the majority of those that were detected did not co-express GCG. Similarly, the C-peptide+ population induced with the extended activin protocol contained a lower proportion of GCG-expressing cells than those generated in the absence of the additional activin stimulation (Fig. 5B,C).
Pancreatic differentiation from other hPSC lines
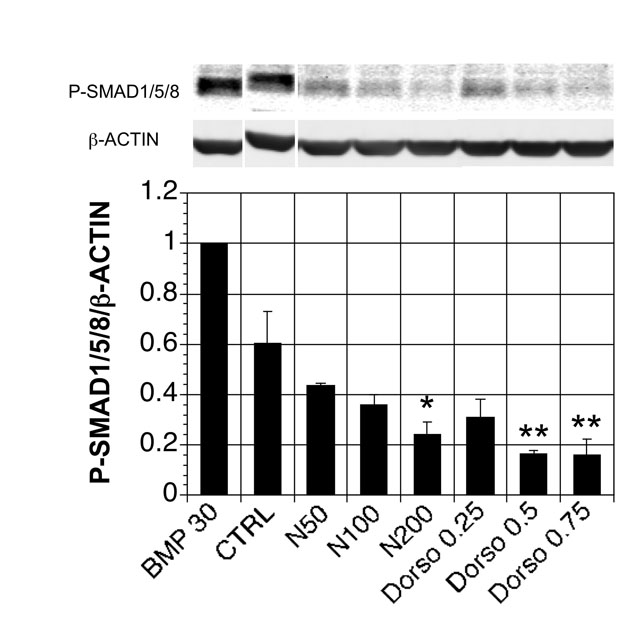
To determine whether our approach was broadly applicable, we evaluated the differentiation potential of three other hESC lines; H1, H9 and a reporter HES3 line carrying a green fluorescent protein (GFP) cDNA targeted to the INS locus (S.J.M., X.L., A.G.E. and E.G.S., unpublished) as well as a transgene-free hiPSC line (38-2) (I.-H.P. and G.Q.D., unpublished). All these lines responded well to activin and efficiently generated CXCR4+ CD117+ populations (not shown). Our initial attempts at producing pancreatic lineage cells from the H1 hESC-derived CXCR4+ CD117+ population were not successful; rather than differentiating along the pancreatic lineage, these cells gave rise to hepatic cells, as indicated by albumin (ALB) expression (Fig. 6A, ALB red line Ctrl). Hepatic lineage cells developed despite the fact that BMP signaling was blocked from stage 3 onwards, suggesting that commitment occurs earlier in culture. The presence of phosphorylated SMAD1/5/8 in H1-derived stage 1 cultures (Ctrl) (see Fig. S7 in the supplementary material) supports the concept that endogenous BMP signaling prior to the NCR step promotes hepatic differentiation. Addition of noggin or the small molecule BMP inhibitor dorsomorphin (Yu et al., 2008) to the cultures during stage 2 of differentiation inhibited SMAD1/5/8 phosphorylation (see Fig. S7 in the supplementary material). As dorsomorphin was more effective than noggin at inducing INS (see Fig. S8 in the supplementary material) and has been shown to induce β-cell development in zebrafish embryos (Chung et al., 2010), we used it to inhibit BMP signaling in H1-derived cultures. Cells treated with increasing concentrations of dorsomorphin (stage 2) displayed increasing levels of INS and decreasing levels of ALB (Fig. 6A). The dorsomorphin-treated cultures contained typical clusters of PDX1+ and C-peptide+ cells (Fig. 6B). Flow-cytometric analyses revealed that C-peptide+ cells represented ~6% of the entire dorsomorphin-treated population (5.7±0.8%, n=3; Fig. 6B,C).
Fig. 6.

Pancreatic differentiation of different human pluripotent stem cells (hPSCs). (A) Quantitative PCR (QPCR) analysis for INS (black histogram) and ALB (red line) at d25 of differentiation in H1-derived populations generated in the presence of FGF10 alone (d7-10; ctrl) or FGF10 and 0.25 μM (dorso 0.25), 0.50 μM (dorso 0.50) or 0.75 μM (dorso 0.75) dorsomorphin. Line and bars represent mean±s.d. n=3. (B) Immunostaining for C-peptide (red) and PDX1 (green) at d27 in H1-derived cultures. (C,D) Flow-cytometric analysis of C-peptide at d25 of differentiation in untreated (Ctrl) and dorsomorphin-treated cultures (Dorso) of H1-derived cells. Percentage represents the mean percentage cells expressing C-peptide (s.d.=0.8 for D, n=3). (E) Flow cytometric analysis measuring GFP at d22 of differentiation in INS:GFP-HES3-derived populations. Cells were differentiated in the presence of 0.75 μM dorsomorphin (Dorso). Percentage represents the mean percentage of cells expressing GFP (s.d.=2.3%, n=4). (F) Presence of GFP+ clusters in the d22 population generated from the INS:GFP-HES3 cell line differentiated in the presence of 0.75 μM dorsomorphin (Dorso). (G-I) Immunostaining for C-peptide in day 22 INS:GFP HES3-derived populations. (J) QPCR analysis for INS in d25 populations generated from the 38-2 iPS cell line. Cells were differentiated in the absence (Ctrl) or presence of 0.75 μM dorsomorphin (Dorso). Bars represent mean±s.d. Asterisk indicates statistical significance determined by t-test. P=0.048, n=4. (K) Immunostaining for C-peptide in d27 populations generated from the 38-2 iPS cell line. (L,M) Flow-cytometric analysis of C-peptide at d25 of differentiation in 38-2-derived untreated (Ctrl) and dorsomorphin-treated populations (Dorso). Percentages represent the mean percentage of cells expressing C-peptide (s.d.=1.3% for L, n=4; s.d.=0.6% for M, n=3).
Pancreatic development from the HES3 INS:GFP and H9 cell line also depended upon the addition of dorsomorphin at stage 2. As observed with H1 cells, the HES3 line differentiated along the hepatic lineage in the absence of dorsomorphin (not shown). Following addition of dorsomorphin, the cells differentiated towards the pancreatic lineage, yielding populations consisting of >6% INS:GFP+ cells at day 22 of culture (6.7±2.3%, n=4; Fig. 6E,F). No INS:GFP+ cells were detected in the cultures generated from cells that did not receive dorsomorphin (not shown). The INS:GFP+ cells co-expressed C-peptide (day 22) indicating that GFP expression is reflective of insulin production (Fig. 6G-I). H9 cells generated an average of 11±1.7% C-peptide+ cells by stage 5, when dorsomorphin was added during stage 2. Less than 4% C-peptide+ cells were detected in its absence (see Fig. S9A,B in the supplementary material). As observed with the HES2 cells, extended activin treatment diminished pancreatic differentiation of H9 cells in the monolayer format (see Fig. S9A,C in the supplementary material). H9 and HES3 INS:GFP cells differentiated with the D'Amour/Kroon protocol (protocol 1) generated <1% C-peptide+ cells (see Fig. S9A,D in the supplementary material) and populations (H9) that expressed only low levels of SST and no detectable INS and GCG (see Fig. S9C in the supplementary material). In contrast to H1, H9 and HES3 hESCs, the 38-2 hiPSC line did not require dorsomorphin during stage 2. In fact, addition of dorsomorphin at this stage inhibited the generation of insulin-expressing cells (Fig. 6J; P=0.048, n=4). By day 25 of differentiation, the 38-2 hiPSC line did generate clusters of C-peptide+ cells (Fig. 6K) that represented >3% of the culture (3.3±1.3%, n=4; Fig. 6L,M). A summary of the differentiation protocol highlighting the requirement for BMP inhibition by the different cell lines is depicted in Fig. S10 in the supplementary material.
Isolation and characterization of insulin-positive cells
To gain a better understanding of the end-stage population we stained cells with a panel of antibodies recognizing different cellular subsets of the adult pancreas (alpha, ductal, exocrine and pan-islet) (Dorrell et al., 2008). Approximately 30% (34±16%, n=3) of the day 22 population stained with the pan-islet (HPi3) antibody, whereas 43% (43±4%, n=3) stained with the pan-exocrine antibody (HPx1) (Fig. 7A,B). Most, but not all, INS:GFP+ cells were HPi3+ (Fig. 7A), whereas none of the INS:GFP+ cells stained with HPx1 (Fig. 7B). No cells stained with the alpha (HPα2)- or ductal (HPd1)-specific antibodies (not shown). The HPx1− INS:GFP+ (INS:GFP+), HPx1+INS:GFP− (HPx1+) and double negative cell (DN) populations depicted in Fig. 7B were isolated from day 22 cultures by FACS and analyzed by qPCR for expression of genes indicative of endocrine [INS, GCG, PPY (pancreatic polypeptide), SST, GHRL (ghrelin), NEUROD1, PDX1 and ARX] and exocrine [CELA1 (elastase 1) and PTF1A] development. The INS:GFP+ population expressed INS, SST and very high levels of GCG, whereas GHRL and PPY were not detected (Fig. 7C; data not shown). The level of INS is similar to that found in the adult pancreas, whereas SST and GCG expression exceeded that in the adult tissue. These endocrine genes were not expressed in the HPx1+ or DN populations. Immunocytochemistry for C-peptide, SST and GCG revealed that the majority of cells are polyhormonal expressing two (C-peptide and either GCG and SST) or three hormones (see Fig. S11 in the supplementary material). Consistent with the patterns of INS, SST and GCG expression, the endocrine-specific transcription factors NEUROD1 and ARX were expressed at highest levels in the INS:GFP+ fraction (Fig. 7D). PDX1 was detected in both the INS:GFP+ and HPx1+ fraction and was found at levels higher than in the adult pancreas (Fig. 7D). Genes indicative of exocrine development (CELA1 and PTF1A) were expressed at higher levels in the HPx1+ population than in the INS:GFP+ and DN cells. However, the expression level of these genes was variable between experiments and dramatically lower than the levels measured in the adult pancreas (Fig. 7E). To verify that a similar enrichment could be achieved with the pan-islet antibody on an untargeted cell line, we sorted pan-islet+ and pan-islet− cells from HES2-derived d22 cultures (Fig. 7F) and analyzed the two fractions, as well as the pre-sort (PS) population, for INS and GCG expression (Fig. 7F-G). Expression of INS and GCG segregated to the pan-islet+ fraction indicating that this antibody can be used to enrich endocrine-like cells from end-stage populations.
Fig. 7.

Isolation and characterization of endocrine populations. (A,B) Flow-cytometric analysis for GFP and the pan-Islet (HPi3; A) or the pan-Exocrine (HPx1; B) markers in INS:GFP-HES3-derived populations. (C-E) Quantitative PCR (QPCR) analysis for INS, SST, GCG, NEUROD1, PDX1, ARX, CELA1 and PTF1A in the pre-sort (PS), double negative (DN), INS:GFP+ and HPx1+ sorted populations. Values for adult pancreas (Ad pancreas) are shown for comparison. Bars represent mean±s.d., n=4. (F) Flow-cytometric analysis showing the presence of pan-Islet+ cells in a HES2-derived population at d22 of differentiation. (G) QPCR analysis for INS and GCG in the sorted populations: pre-sort (PS), pan-ISLET+ and pan-ISLET−. Bars represent mean±s.d., n=2.
DISCUSSION
Successful derivation of β-cells from PSCs depends on recapitulating key embryonic developmental steps within the differentiation cultures. Using this approach, both hESCs and hiPSCs can be differentiated to a pancreatic fate, resulting in the generation of insulin-expressing cells (Cai et al., 2009; Chen et al., 2009; D'Amour et al., 2006; Jiang, J. et al., 2007; Jiang, W. et al., 2007; Kroon et al., 2008; Maehr et al., 2009; Mfopou et al., 2010; Phillips et al., 2007; Shim et al., 2007; Tateishi et al., 2008; Zhang et al., 2009). Although these findings represent an important step forward, most studies reported low efficiency of differentiation and heterogeneity in the ability of different PSC lines to generate pancreatic lineage cells when treated under identical conditions (Borowiak and Melton, 2009; Chen et al., 2009; D'Amour et al., 2006; Maehr et al., 2009; Mfopou et al., 2010). In this study, we evaluated the role of TGFβ family members and WNT on the patterning and pancreatic specification of hPSC-derived definitive endoderm, as previous studies in different model systems have shown that stage-specific control of these pathways is essential for establishment of a pancreatic fate (Chung et al., 2010; McLin et al., 2007; Wandzioch and Zaret, 2009). Our findings demonstrate that these pathways are also key regulators of pancreatic fate in hPSC cultures and show that endogenous BMP signaling is a major contributing factor to the variability of pancreatic differentiation observed with different hPSCs. Through appropriate activation and inhibition of BMP, activin/nodal and WNT signaling, it is possible to generate populations consisting of greater than 25% C-peptide+ cells that express levels of insulin similar to those found in the adult pancreas.
Patterning of the hESC-derived definitive endoderm
The day 5 CXCR4+ CD117+ population generated with the EB approach contains an endoderm population that requires additional activin/nodal signaling to respond appropriately to patterning molecules. Based on observations in the mouse ESC model, we interpret this activin-dependent step as representing the induction of definitive endoderm from more immature anterior primitive streak (APS) progenitors (Gadue et al., 2009; Gadue et al., 2006). The dramatic increase in INS expression following the 2-day activin step is striking and demonstrates the importance of generating appropriate progenitor populations at early stages of differentiation. The observation that INS expression is significantly reduced if the induction period is maintained for an additional 24 hours, highlights the importance of timing and suggests that the duration of activin/nodal signaling is crucial for optimal pancreatic differentiation. The fact that the cells induced as a monolayer do not require the extended activin induction indicates that the culture format can impact the optimal timing of signaling through this pathway. As induction is more rapid in the monolayer cultures than in the EBs, nodal/activin signaling is probably more efficient under these conditions. Consequently, extended treatment with activin might not be necessary.
A key step in the generation of endoderm-derived cell types is patterning the appropriate region of the gut tube along the anteroposterior axis. Studies using different model systems have shown that during gastrulation, WNT signaling is restricted to the posterior region of the embryo and, together with FGF signaling, is responsible for the induction of a posterior phenotype (Dessimoz et al., 2006; Keenan et al., 2006; McLin et al., 2007; Pownall et al., 1996). Our findings show that WNT signaling functions in a similar manner to promote the development of a posterior phenotype in the hESC cultures. At appropriate concentrations, Wnt3a did induce a fourfold increase in the INS levels, indicating that WNT signaling plays a role in patterning endoderm to a pancreatic fate in hESC-derived populations.
Endocrine lineage commitment
Studies using zebrafish, mouse embryos and mouse ESCs have shown that BMP signaling is essential for hepatic specification (Chung et al., 2008; Gouon-Evans et al., 2006; Rossi et al., 2001), and that inhibition of this pathway can promote β-cell development (Chung et al., 2010; Wandzioch and Zaret, 2009). BMP inhibitors have been included in hESC pancreatic induction protocols prior to and during the retinoic acid induction step, presumably to suppress hepatic development (Cai et al., 2009; Kroon et al., 2008; Mfopou et al., 2010; Zhang et al., 2009). Our data show that the requirement for BMP inhibition extends beyond the NCR step, as blocking the pathway following PDX1 induction resulted in a dramatic 19-fold increase in INS expression. Although the addition of SB (which inhibits TGFβ/activin/nodal signaling) alone had a more modest effect on INS expression, the combination of both antagonists had an additive effect, resulting in a 27-fold increase in INS expression. In addition to enhancing INS expression, the addition of SB led to more than a threefold increase in total cell number within the cultures, indicating that endogenous signaling through these pathways inhibits cell proliferation at this stage. The promotion of pancreatic development by these inhibitors is consistent with findings in the mouse embryo, which show that TGFβ signaling can restrain specification of pancreatic progenitors (Wandzioch and Zaret, 2009). These findings are also supported by a recent study by Rezania and colleagues (Rezania et al., 2011) who also found that blocking TGFβ/activin/nodal signaling following pancreatic specification promoted endocrine specification from H1 hESCs. Collectively, these observations highlight the role played by endogenous signaling and the need to control both intrinsic and extrinsic factors in the differentiation cultures.
Importantly, we found that blocking BMP signaling prior to PDX1 induction was essential for some, but not all cell lines, suggesting that endogenous BMP levels vary between different hPSC lines and can dramatically affect the outcome of the pancreatic differentiation protocol (see Fig. S7 in the supplementary material). The generation of INS-expressing cells from the H1, HES3.INS:GFP and H9 cell lines was enhanced by inhibiting BMP (at stage 2) prior to PDX1 induction. In the absence of BMP inhibitors at this stage, the H1 hESCs differentiated to hepatic cells, probably owing to high levels of endogenous BMP signaling, as indicated by the presence of pSMAD1/5/8. These observations are consistent with recent studies reporting that inhibition of BMP signaling was required for the generation of PDX1+ cells from H1 and other hESC lines (Cai et al., 2009; Mfopou et al., 2010). In contrast to the H1, HES3 and H9 lines, induction of pancreatic lineage cells from HES2 cells or the iPSC line 38-2 did not require inhibition of BMP prior to PDX1 induction.
Through the combinatorial regulation of the signaling pathways outlined in this study (see Fig. S10 in the supplementary material), we are able to generate pancreatic populations consisting of 3-25% C-peptide+ cells from five different hPSC lines with corresponding INS levels ranging from 1.5% to 89% of that found in the adult pancreas. Although there is still some variability in these cultures, it is considerably less than that reported by D'Amour et al., who showed that the levels of INS expression differed by 1000- to 10,000-fold in cultures generated from different hESCs (D'Amour et al., 2006). A comparison of cells generated by the D'Amour/Kroon protocol with those generated by our protocol indicated that the early induction step to the PDX1 progenitor stage were comparable. Beyond this step, the populations differed considerably, as commitment to the endocrine lineage and development of INS- and GCG-expressing cells was significantly more efficient with our protocol, likely due to the inhibition of BMP and TGFβ/activin/nodal signaling following PDX1 induction. Inhibition of these pathways at this step was required for all cell lines tested to promote the development of C-peptide+ cells. It is possible that the CyT203 and CyT49 lines used by D'Amour/Kroon to develop the initial differentiation protocol display low levels of endogenous BMP and TGFβ/activin/nodal signaling, allowing commitment to the endocrine lineage without the need to add inhibitors of these pathways.
Only one other study to date has evaluated the proportion of C-peptide+ cells by showing 2-8% positive cells in end-stage H1-derived islet-like clusters (ILCs) (Jiang, J. et al., 2007). Several other groups have quantified differentiation by flow cytometry using an anti-insulin rather than an anti-C-peptide antibody and reported the production of 7% and 25% insulin+ cells from the Cyt203 and H1/H9 hESC lines, respectively (D'Amour et al., 2006; Zhang et al., 2009). Rezania and colleagues generated 65% INS+ GCG+ cells in their end-stage clusters, however, these clusters represent only a component of the stage 6 culture, and the frequency of INS+ cells in the entire differentiated population is not reported (Rezania et al., 2011). Although immunocytochemistry clearly documented the presence of insulin-producing cells in these cultures, quantification using an anti-insulin antibody can be difficult to interpret given that most media contain insulin, which can be internalized by the cultured cells (Hansson et al., 2004). The efficient development of insulin+ cells enabled us to use the targeted reporter INS:GFP line and anti-pancreatic antibodies to isolate populations enriched for endocrine potential (Dorrell et al., 2008). The fact that some of the antibodies generated against the adult pancreas react with the hESC-derived cells is encouraging and indicates that markers that define subsets of cells in the adult pancreas are present on the cells generated in the differentiation cultures.
Most, but not all, of the C-peptide+ cells generated in our study co-express GCG or GCG and SST. In contrast to what is reported during mouse pancreatic embryogenesis, a cell co-expressing INS, GCG and SST has been identified in human fetal pancreas, suggesting that our culture conditions might be recapitulating an embryonic stage of pancreatic development (Polak et al., 2000). As such, they might represent an intermediate stage of endocrine development or the in vitro equivalent of the first transition of insulin-producing cells, which in the mouse embryo do not contribute to the adult β-cell population (Herrera, 2000; Teitelman et al., 1993). Future studies will be required to define further the developmental status of the polyhormonal cells in hPSC cultures and to identify the regulatory pathways that could promote their differentiation to functional β-cells.
In conclusion, our findings clearly document the temporal requirements for TGFβ family members and canonical WNT signaling during differentiation of hPSCs to pancreatic lineage cells. Appropriate manipulation of these pathways results in populations that contain high frequencies of C-peptide+ cells and express levels of INS similar to those found in the adult pancreas. In addition to defining the role of these pathways in pancreatic differentiation, our study highlights several important aspects regarding lineage-specific differentiation of hPSCs. First, it demonstrates that signaling pathways that govern embryonic development can be used to control lineage development in hPSC cultures. Second, it clearly documents the importance of generating appropriate staged cells at each step of the developmental pathway to achieve the desired terminally differentiated state. Third, it shows that endogenous signaling molecules can compete with the activity of the key pro-differentiation pathways, giving rise to the variability in the differentiation potential observed with various hPSC lines. Given these variables, deciphering the complex signaling networks directing lineage-specific commitment during embryogenesis and properly manipulating these pathways in vitro are essential for the efficient derivation of mature cell types ex vivo.
Supplementary Material
Acknowledgements
We would like to thank members of the Keller laboratory, Dr Nadeem Moghal and Dr Benjamin Neel for comments on the manuscript. This work was supported by a grant from NIH/NIDDK (U01-DK072513) to G.K. M.C.N. was supported by a JDRF postdoctoral fellowship. A.G.E., E.G.S., X.L. and S.J.M. are supported by grants from the JDRF, the ASCC and the NHMRC (Australia). A.G.E. and E.G.S. are Senior Research Fellows of the NHMRC. Deposited in PMC for release after 12 months.
Footnotes
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material for this article is available at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.055236/-/DC1
References
- Ameri J., Stahlberg A., Pedersen J., Johansson J. K., Johannesson M. M., Artner I., Semb H. (2010). FGF2 specifies hESC-derived definitive endoderm into foregut/midgut cell lineages in a concentration-dependent manner. Stem Cells 28, 45-56 [DOI] [PubMed] [Google Scholar]
- Apelqvist A., Li H., Sommer L., Beatus P., Anderson D. J., Honjo T., Hrabe de Angelis M., Lendahl U., Edlund H. (1999). Notch signalling controls pancreatic cell differentiation. Nature 400, 877-881 [DOI] [PubMed] [Google Scholar]
- Bardeesy N., Cheng K. H., Berger J. H., Chu G. C., Pahler J., Olson P., Hezel A. F., Horner J., Lauwers G. Y., Hanahan D., et al. (2006). Smad4 is dispensable for normal pancreas development yet critical in progression and tumor biology of pancreas cancer. Genes Dev. 20, 3130-3146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borowiak M., Melton D. A. (2009). How to make beta cells? Curr. Opin. Cell Biol. 21, 727-732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai J., Yu C., Liu Y., Chen S., Guo Y., Yong J., Lu W., Ding M., Deng H. (2009). Generation of homogeneous PDX1(+) pancreatic progenitors from human ES cell-derived endoderm cells. J. Mol. Cell Biol. 2, 50-60 [DOI] [PubMed] [Google Scholar]
- Candia A. F., Hu J., Crosby J., Lalley P. A., Noden D., Nadeau J. H., Wright C. V. (1992). Mox-1 and Mox-2 define a novel homeobox gene subfamily and are differentially expressed during early mesodermal patterning in mouse embryos. Development 116, 1123-1136 [DOI] [PubMed] [Google Scholar]
- Chen S., Borowiak M., Fox J. L., Maehr R., Osafune K., Davidow L., Lam K., Peng L. F., Schreiber S. L., Rubin L. L., et al. (2009). A small molecule that directs differentiation of human ESCs into the pancreatic lineage. Nat. Chem. Biol. 5, 258-265 [DOI] [PubMed] [Google Scholar]
- Chung W. S., Shin C. H., Stainier D. Y. (2008). Bmp2 signaling regulates the hepatic versus pancreatic fate decision. Dev. Cell 15, 738-748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung W. S., Andersson O., Row R., Kimelman D., Stainier D. Y. (2010). Suppression of Alk8-mediated Bmp signaling cell-autonomously induces pancreatic beta-cells in zebrafish. Proc. Natl. Acad. Sci. USA 107, 1142-1147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collignon J., Sockanathan S., Hacker A., Cohen-Tannoudji M., Norris D., Rastan S., Stevanovic M., Goodfellow P. N., Lovell-Badge R. (1996). A comparison of the properties of Sox-3 with Sry and two related genes, Sox-1 and Sox-2. Development 122, 509-520 [DOI] [PubMed] [Google Scholar]
- Conlon F. L., Lyons K. M., Takaesu N., Barth K. S., Kispert A., Herrmann B., Robertson E. J. (1994). A primary requirement for nodal in the formation and maintenance of the primitive streak in the mouse. Development 120, 1919-1928 [DOI] [PubMed] [Google Scholar]
- Costa M., Dottori M., Sourris K., Jamshidi P., Hatzistavrou T., Davis R., Azzola L., Jackson S., Lim S. M., Pera M., et al. (2007). A method for genetic modification of human embryonic stem cells using electroporation. Nat. Protoc. 2, 792-796 [DOI] [PubMed] [Google Scholar]
- D'Amour K. A., Bang A. G., Eliazer S., Kelly O. G., Agulnick A. D., Smart N. G., Moorman M. A., Kroon E., Carpenter M. K., Baetge E. E. (2006). Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat. Biotechnol. 24, 1392-1401 [DOI] [PubMed] [Google Scholar]
- Dessimoz J., Opoka R., Kordich J. J., Grapin-Botton A., Wells J. M. (2006). FGF signaling is necessary for establishing gut tube domains along the anterior-posterior axis in vivo. Mech. Dev. 123, 42-55 [DOI] [PubMed] [Google Scholar]
- Dorrell C., Abraham S. L., Lanxon-Cookson K. M., Canaday P. S., Streeter P. R., Grompe M. (2008). Isolation of major pancreatic cell types and long-term culture-initiating cells using novel human surface markers. Stem Cell Res. 1, 183-194 [DOI] [PubMed] [Google Scholar]
- Feldman B., Gates M. A., Egan E. S., Dougan S. T., Rennebeck G., Sirotkin H. I., Schier A. F., Talbot W. S. (1998). Zebrafish organizer development and germ-layer formation require nodal-related signals. Nature 395, 181-185 [DOI] [PubMed] [Google Scholar]
- Gadue P., Huber T. L., Paddison P. J., Keller G. M. (2006). Wnt and TGF-beta signaling are required for the induction of an in vitro model of primitive streak formation using embryonic stem cells. Proc. Natl. Acad. Sci. USA 103, 16806-16811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gadue P., Gouon-Evans V., Cheng X., Wandzioch E., Zaret K. S., Grompe M., Streeter P. R., Keller G. M. (2009). Generation of monoclonal antibodies specific for cell surface molecules expressed on early mouse endoderm. Stem Cells 27, 2103-2113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gouon-Evans V., Boussemart L., Gadue P., Nierhoff D., Koehler C. I., Kubo A., Shafritz D. A., Keller G. (2006). BMP-4 is required for hepatic specification of mouse embryonic stem cell-derived definitive endoderm. Nat. Biotechnol. 24, 1402-1411 [DOI] [PubMed] [Google Scholar]
- Gritsman K., Talbot W. S., Schier A. F. (2000). Nodal signaling patterns the organizer. Development 127, 921-932 [DOI] [PubMed] [Google Scholar]
- Hansson M., Tonning A., Frandsen U., Petri A., Rajagopal J., Englund M. C., Heller R. S., Hakansson J., Fleckner J., Skold H. N., et al. (2004). Artifactual insulin release from differentiated embryonic stem cells. Diabetes 53, 2603-2609 [DOI] [PubMed] [Google Scholar]
- Hebrok M. (2003). Hedgehog signaling in pancreas development. Mech. Dev. 120, 45-57 [DOI] [PubMed] [Google Scholar]
- Hebrok M., Kim S. K., Melton D. A. (1998). Notochord repression of endodermal Sonic hedgehog permits pancreas development. Genes Dev. 12, 1705-1713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrera P. L. (2000). Adult insulin- and glucagon-producing cells differentiate from two independent cell lineages. Development 127, 2317-2322 [DOI] [PubMed] [Google Scholar]
- Inman G. J., Nicolas F. J., Callahan J. F., Harling J. D., Gaster L. M., Reith A. D., Laping N. J., Hill C. S. (2002). SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 62, 65-74 [DOI] [PubMed] [Google Scholar]
- Jiang J., Au M., Lu K., Eshpeter A., Korbutt G., Fisk G., Majumdar A. S. (2007). Generation of insulin-producing islet-like clusters from human embryonic stem cells. Stem Cells 25, 1940-1953 [DOI] [PubMed] [Google Scholar]
- Jiang W., Shi Y., Zhao D., Chen S., Yong J., Zhang J., Qing T., Sun X., Zhang P., Ding M., et al. (2007). In vitro derivation of functional insulin-producing cells from human embryonic stem cells. Cell Res. 17, 333-344 [DOI] [PubMed] [Google Scholar]
- Johannesson M., Stahlberg A., Ameri J., Sand F. W., Norrman K., Semb H. (2009). FGF4 and retinoic acid direct differentiation of hESCs into PDX1-expressing foregut endoderm in a time- and concentration-dependent manner. PLoS ONE 4, e4794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones C. M., Kuehn M. R., Hogan B. L., Smith J. C., Wright C. V. (1995). Nodal-related signals induce axial mesoderm and dorsalize mesoderm during gastrulation. Development 121, 3651-3662 [DOI] [PubMed] [Google Scholar]
- Keenan I. D., Sharrard R. M., Isaacs H. V. (2006). FGF signal transduction and the regulation of Cdx gene expression. Dev. Biol. 299, 478-488 [DOI] [PubMed] [Google Scholar]
- Kennedy M., D'Souza S. L., Lynch-Kattman M., Schwantz S., Keller G. (2007). Development of the hemangioblast defines the onset of hematopoiesis in human ES cell differentiation cultures. Blood 109, 2679-2687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koh E. Y., Chen T., Daley G. Q. (2002). Novel retroviral vectors to facilitate expression screens in mammalian cells. Nucleic Acids Res. 30, e142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroon E., Martinson L. A., Kadoya K., Bang A. G., Kelly O. G., Eliazer S., Young H., Richardson M., Smart N. G., Cunningham J., et al. (2008). Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat. Biotechnol. 26, 443-452 [DOI] [PubMed] [Google Scholar]
- Krutzik P. O., Nolan G. P. (2003). Intracellular phospho-protein staining techniques for flow cytometry: monitoring single cell signaling events. Cytometry A 55, 61-70 [DOI] [PubMed] [Google Scholar]
- Lazzara M. J., Lane K., Chan R., Jasper P. J., Yaffe M. B., Sorger P. K., Jacks T., Neel B. G., Lauffenburger D. A. (2010). Impaired SHP2-mediated extracellular signal-regulated kinase activation contributes to gefitinib sensitivity of lung cancer cells with epidermal growth factor receptor-activating mutations. Cancer Res. 70, 3843-3850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe L. A., Yamada S., Kuehn M. R. (2001). Genetic dissection of nodal function in patterning the mouse embryo. Development 128, 1831-1843 [DOI] [PubMed] [Google Scholar]
- Maehr R., Chen S., Snitow M., Ludwig T., Yagasaki L., Goland R., Leibel R. L., Melton D. A. (2009). Generation of pluripotent stem cells from patients with type 1 diabetes. Proc. Natl. Acad. Sci. USA 106, 15768-15773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin M., Gallego-Llamas J., Ribes V., Kedinger M., Niederreither K., Chambon P., Dolle P., Gradwohl G. (2005). Dorsal pancreas agenesis in retinoic acid-deficient Raldh2 mutant mice. Dev. Biol. 284, 399-411 [DOI] [PubMed] [Google Scholar]
- Matthews W., Jordan C. T., Gavin M., Jenkins N. A., Copeland N. G., Lemischka I. R. (1991). A receptor tyrosine kinase cDNA isolated from a population of enriched primitive hematopoietic cells and exhibiting close genetic linkage to c-kit. Proc. Natl. Acad. Sci. USA 88, 9026-9030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLin V. A., Rankin S. A., Zorn A. M. (2007). Repression of Wnt/beta-catenin signaling in the anterior endoderm is essential for liver and pancreas development. Development 134, 2207-2217 [DOI] [PubMed] [Google Scholar]
- Mfopou J. K., Chen B., Mateizel I., Sermon K., Bouwens L. (2010). Noggin, retinoids, and fibroblast growth factor regulate hepatic or pancreatic fate of human embryonic stem cells. Gastroenterology 138, 2233-2245 [DOI] [PubMed] [Google Scholar]
- Molotkov A., Molotkova N., Duester G. (2005). Retinoic acid generated by Raldh2 in mesoderm is required for mouse dorsal endodermal pancreas development. Dev. Dyn. 232, 950-957 [DOI] [PubMed] [Google Scholar]
- Murry C. E., Keller G. (2008). Differentiation of embryonic stem cells to clinically relevant populations: lessons from embryonic development. Cell 132, 661-680 [DOI] [PubMed] [Google Scholar]
- Nichols J., Zevnik B., Anastassiadis K., Niwa H., Klewe-Nebenius D., Chambers I., Scholer H., Smith A. (1998). Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 95, 379-391 [DOI] [PubMed] [Google Scholar]
- Nostro M. C., Cheng X., Keller G. M., Gadue P. (2008). Wnt, activin, and BMP signaling regulate distinct stages in the developmental pathway from embryonic stem cells to blood. Cell Stem Cell 2, 60-71 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osada S. I., Wright C. V. (1999). Xenopus nodal-related signaling is essential for mesendodermal patterning during early embryogenesis. Development 126, 3229-3240 [DOI] [PubMed] [Google Scholar]
- Phillips B. W., Hentze H., Rust W. L., Chen Q. P., Chipperfield H., Tan E. K., Abraham S., Sadasivam A., Soong P. L., Wang S. T., et al. (2007). Directed differentiation of human embryonic stem cells into the pancreatic endocrine lineage. Stem Cells Dev. 16, 561-578 [DOI] [PubMed] [Google Scholar]
- Polak M., Bouchareb-Banaei L., Scharfmann R., Czernichow P. (2000). Early pattern of differentiation in the human pancreas. Diabetes 49, 225-232 [DOI] [PubMed] [Google Scholar]
- Pownall M. E., Tucker A. S., Slack J. M., Isaacs H. V. (1996). eFGF, Xcad3 and Hox genes form a molecular pathway that establishes the anteroposterior axis in Xenopus. Development 122, 3881-3892 [DOI] [PubMed] [Google Scholar]
- Rezania A., Riedel M. J., Wideman R. D., Karanu F., Ao Z., Warnock G. L., Kieffer T. J. (2011). Production of functional glucagon-secreting alpha cells from human embryonic stem cells. Diabetes 60, 239-247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi J. M., Dunn N. R., Hogan B. L., Zaret K. S. (2001). Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm. Genes Dev. 15, 1998-2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saga Y., Hata N., Kobayashi S., Magnuson T., Seldin M. F., Taketo M. M. (1996). MesP1: a novel basic helix-loop-helix protein expressed in the nascent mesodermal cells during mouse gastrulation. Development 122, 2769-2778 [DOI] [PubMed] [Google Scholar]
- Shim J. H., Kim S. E., Woo D. H., Kim S. K., Oh C. H., McKay R., Kim J. H. (2007). Directed differentiation of human embryonic stem cells towards a pancreatic cell fate. Diabetologia 50, 1228-1238 [DOI] [PubMed] [Google Scholar]
- Stafford D., Prince V. E. (2002). Retinoic acid signaling is required for a critical early step in zebrafish pancreatic development. Curr. Biol. 12, 1215-1220 [DOI] [PubMed] [Google Scholar]
- Tateishi K., He J., Taranova O., Liang G., D'Alessio A. C., Zhang Y. (2008). Generation of insulin-secreting islet-like clusters from human skin fibroblasts. J. Biol. Chem. 283, 31601-31607 [DOI] [PubMed] [Google Scholar]
- Teitelman G., Alpert S., Polak J. M., Martinez A., Hanahan D. (1993). Precursor cells of mouse endocrine pancreas coexpress insulin, glucagon and the neuronal proteins tyrosine hydroxylase and neuropeptide Y, but not pancreatic polypeptide. Development 118, 1031-1039 [DOI] [PubMed] [Google Scholar]
- Vecchi A., Garlanda C., Lampugnani M. G., Resnati M., Matteucci C., Stoppacciaro A., Schnurch H., Risau W., Ruco L., Mantovani A., et al. (1994). Monoclonal antibodies specific for endothelial cells of mouse blood vessels. Their application in the identification of adult and embryonic endothelium. Eur. J. Cell Biol. 63, 247-254 [PubMed] [Google Scholar]
- Wandzioch E., Zaret K. S. (2009). Dynamic signaling network for the specification of embryonic pancreas and liver progenitors. Science 324, 1707-1710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wells J. M., Melton D. A. (1999). Vertebrate endoderm development. Annu. Rev. Cell Dev. Biol. 15, 393-410 [DOI] [PubMed] [Google Scholar]
- Wood H. B., Episkopou V. (1999). Comparative expression of the mouse Sox1, Sox2 and Sox3 genes from pre-gastrulation to early somite stages. Mech. Dev. 86, 197-201 [DOI] [PubMed] [Google Scholar]
- Yu P. B., Hong C. C., Sachidanandan C., Babitt J. L., Deng D. Y., Hoyng S. A., Lin H. Y., Bloch K. D., Peterson R. T. (2008). Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism. Nat. Chem. Biol. 4, 33-41 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang D., Jiang W., Liu M., Sui X., Yin X., Chen S., Shi Y., Deng H. (2009). Highly efficient differentiation of human ES cells and iPS cells into mature pancreatic insulin-producing cells. Cell Res. 19, 429-438 [DOI] [PubMed] [Google Scholar]
- Zhou X., Sasaki H., Lowe L., Hogan B. L., Kuehn M. R. (1993). Nodal is a novel TGF-beta-like gene expressed in the mouse node during gastrulation. Nature 361, 543-547 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}