


Abstract
Cascade regulatory circuits have been described that control numerous cell processes, and may provide models for the design of artificial circuits with novel properties. Here we describe the design of a transcriptional regulatory cascade to amplify the cell response to a given signal. We used the salicylate-responsive activators of Pseudomonas putida NahR of the naphthalene degradation plasmid NAH7 and XylS2, a mutant regulator of the TOL plasmid for catabolism of m-xylene and their respective cognate promoters Psal and Pm. Control of the expression of xylS2 with the nahR/Psal system permitted either their selective activation with specific effectors for each protein or the simultaneous activation of both of them with salicylate. When cells face the common effector of the two regulators, both the increase in XylS2 concentration and the stimulation of its activity act synergistically on the Pm promoter, amplifying the gene expression capacity by at least one order of magnitude with respect to the individual systems. By changing the hierarchy of regulators, we showed that the specific features of the downstream regulator were crucial for the amplification effect. Directed changes in the effector profile of the regulators allowed the extension of the amplifying system to other molecular signals.
INTRODUCTION
Transcriptional control circuits involving cascades of regulatory proteins are known to determine the functioning of numerous cellular processes both in prokaryotic and eukaryotic systems (1,2). Such cascades channel specific environmental signals into changes in gene expression, so that multiple simultaneous stimuli are processed and translated to bring about activation or repression of specific genes or groups of genes. The simplest regulatory cascades are found in prokaryotic systems and they typically involve at least two different transcriptional activators. These are arranged in such a fashion that a first regulatory component (upstream regulator) controls the expression of a second regulatory gene (downstream regulator) in response to specific signals. In turn, the downstream component acts directly on transcription of the structural genes of the system. Such an arrangement of regulatory genes may serve to control different cellular functions, i.e. to integrate different signals, for the sequential timing of different regulatory events or to amplify the gene expression capacity (ratio of induced:uninduced activity) in order to maximize cell responses. Although examples of transcriptional cascades can be found in prokaryotic systems (3,4), experimental analysis determinating the requisites for the amplifying properties of such a regulatory device remain scarce. The TOL operons for catabolism of toluene/xylenes in Pseudomonas putida via benzoate/toluate and catechol/methylcatechol intermediates (5), provide a suitable example of regulation in cascade. Transcription of the meta-operon of the TOL plasmid of P.putida, which determines the degradation of benzoate/toluates to tricarboxilic acid cycle intermediates, originates at the Pm promoter. Pm is activated by the XylS protein, which belongs to the AraC family of transcriptional regulators (6), in the presence of substrates of the meta pathway such as benzoate or m-toluate (7). Transcription of the xylS gene is in turn controlled by another regulator, XylR (8,9), in combination with the σ54-containing form of RNA polymerase (10) and inducers of the upper pathway. Within this scheme, XylR and XylS act as upstream and downstream regulators, respectively. One key feature of this circuit is that activation of Pm promoter by XylS can also be triggered by an excess of the regulator in the absence of inducers (11,12). This means that the presence of an upper pathway substrate such as benzyl alcohol alone causes Pm to be activated through XylS overproduction even without XylS effectors. Such an activation can then be further augmented in the presence of inducers like m-toluate (13). Furthermore, when the XylS level is artificially increased through different expression systems and gene copy numbers, Pm strength increases through an almost 1000-fold range maintaining its inducibility by benzoate (12). This wide window of intracellular activity indicates that the maximal potentiality of XylS-mediated regulation may require to concert intracellular XylS concentration to the presence of the aromatic inducer. The last work also showed that for a given configuration of the xylS/Pm expression system, the gene expression capacity could not be higher than 100-fold.
In contrast to the TOL system, the catabolic genes of plasmid NAH7 of P.putida for degradation of naphthalene are controlled by a single regulatory protein that is activated by salicylate, one of the intermediates of the catabolic pathway (14). In spite of the structural similarity between salicylate and benzoate, the transcriptional activator NahR of the NAH pathway, which mediates the response to salicylate, is unrelated to XylS since it belongs to the LysR family (15). Interestingly, it is possible to isolate XylS mutants that respond to salicylate (16) with similar ease as for the selection of NahR mutants responding to benzoate (17).
The availability of two different regulators (NahR and XylS2) which respond to the same inducer, and the existence of several effector mutants has allowed us to investigate the performance of the concerted action of the two activators and their cognate promoters in artificial cascade circuits. One of the possible arrangements merges the two systems into a regulatory cascade that amplifies synergistically the promoter output in response to salicylate, starting from a very low level of basal activity. Such an amplification relies on the properties of the downstream regulator–promoter pair (XylS and Pm) and permitted us to draw some general conclusions on the assembly of regulatory cascades designed for the amplification of signals.
MATERIALS AND METHODS
Strains, plasmids, media and general procedures
All plasmids and bacterial strains used in this work are listed in Table 1 and Figure 5. Cultures were grown in LB medium supplemented, where necessary, with ampicillin (150 µg/ml), kanamycin (50 µg/ml), streptomycin (25 µg/ml), tetracyclin (3–10 µg/ml) or piperacillin (40 µg/ml). All manipulations of DNA were made following standard protocols (18). Restriction enzymes were from New England Biolabs, while other modification enzymes (T4 DNA ligase, alkaline phosphatase) were from Boehringer Mannheim. Aromatic inducers were purchased from Aldrich Quimica. Promoter activity was measured by monitoring accumulation of β-galactosidase by cells grown under different conditions. To this end, overnight inoculations of the Escherichia coli strains under examination were diluted 1:100 and incubated for 2 h at 37°C (i.e., up to an OD600 of 0.1). The cultures were then added to the aromatic inducer at the final concentrations indicated in each case, and were further incubated for 5 h at 30°C. The same procedure was employed for the P.putida strains, except that the length of exposure to the inducers was limited to 3 h. In either case, β-galactosidase levels were measured according to Miller (19) in cells permeabilized with chloroform and SDS. Each enzymatic measurement was repeated at least twice in duplicate samples.
Table 1. Bacteria and plasmids.
| Strains and plasmids |
Description |
Reference/source |
| E.coli CC118 | phoA20 thi-1 rspE rpoB argE (Am) recA1 | (27) |
| CC118 (λpir) | CC118 lysogenized with λpir phage | (27) |
| S17-1 (λpir) | F–, recA, hsdR, RP4-2 (Tc::Mu) (Km::Tn7) lysogenized with λpir phage | (36) |
| CC118FH26 | CC118 with mini-Tn5 xylS2/Pm→lacZ inserted in the chromosome, KmR | (24) |
| CC118RSL9 | CC118 with mini-Tn5 nahR/Psal→lacZ inserted in the chromosome, KmR | This work |
| CC1184S2 | CC118 with mini-Tn5 nahR/Psal→xylS2 inserted in the chromosome, KmR | This work |
| CC1184S2PT32 | CC1184S2 with mini-Tn5 Pm→lacZ inserted in the chromosome, KmR, TcR | This work |
| CC118SAL1 | CC118 with mini-Tn5 nahG′::′lacZ inserted in the chromosome, Sp/SmR | This work |
| CC1182NRSL7 | CC118SAL1 with mini-Tn5 xylS2/Pm→nahR Psal→lacZ inserted in the chromosome, KmR, Sp/SmR | This work |
| P.putida KT2442 | Prototrophic, RifR derivative of KT2440 | (27) |
| Plasmids | ||
| pMS15 | ApR, nahR/Psal sequences clones as a 1.6 kb HindIII–PstI insert in pUC8 | (25) |
| pNM482 | ApR, ′lacZ promoter probe plasmid | (37) |
| pUJ9 | ApR, ′lacZ promoter probe plasmid | (38) |
| pUT/mini-Tn5 Sp/Sm | ApR Sp/SmR, R6KoriV, RP4oriT, delivery vector | (38) |
| pTSAL1 | ApR Sp/SmR, nahG′::′lacZ inserted as a NotI fragment in pUT/mini-Tn5 Sp/Sm | This work |
| pFH2 | ApR, similar to pBKT7-0 (23) but with NotI sites flanking the MCS | Lab stock |
| pFH28 | ApR KmR, pUC18Sfi-KmR-xylS-Pm→SfiI | (24) |
| pS2 | ApR, xylS2 with an engineered NcoI site in the first codon inserted as an EcoRI–HindIII fragment in pGC1 | This work |
| pNS2 | ApR, xylS2 cloned as a NcoI–HindIII fragment in pFH2 | This work |
| pNS | ApR, xylS cloned as a NcoI–HindIII fragment in pFH2 | This work |
| pNNR | ApR, nahR cloned as a NcoI–HindIII fragment in pFH2 | This work |
| pCNB2 | ApR KmR, pUT/mini-Tn5 xylS2/Pm→NotI | (24) |
| pCNB1-lacZ | ApR KmR, pUT/mini-Tn5 xylS/Pm→trp′::′lacZ | (24) |
| pCNB2-lacZ | ApR KmR, pCNB2 with trp′::′lacZ reporter downstream of Psal | (24) |
| pCNB2-NR | ApR KmR, pUT/mini-Tn5 xylS2/Pm→nahR | This work |
| pCNB4 | ApR KmR, pUT/mini-Tn5 nahR/Psal | (24) |
| pCNB4-lacZ | ApR KmR, pCNB4 with trp′::′lacZ reporter downstream of Psal | (24) |
| pCNB43-lacZ | ApR KmR, pUT/mini-Tn5 nahR3/Psal→trp′::′lacZ | (17) |
| pCNB44-lacZ | ApR KmR, pUT/mini-Tn5 nahR4/Psal→trp′::′lacZ | (17) |
| pCNB43-S2 | ApR KmR, pUT/mini-Tn5 nahR3/Psal→xyS2 | This work |
| pCNB44-S2 | ApR KmR, pUT/mini-Tn5 nahR4/Psal→xylS2 | This work |
| pCNB44-S | ApR KmR, pUT/mini-Tn5 nahR4/Psal→xylS | This work |
| pUT/Tc Pm-lacZ | ApR TcR, pUT/mini-Tn5 Pm→xylX′::′lacZ | This work |
| pTPmlac | ApR TcR, pUT/mini-Tn5 Pm→trp′::′lacZ | This work |
Figure 5.

Capacity for regulation of single and cascade circuits in P.putida. Strains in the chromosome bearing different minitransposons (first column) containing the regulatory system described in the text were assayed for their β-galactosidase accumulation in response to benzoate 2 mM. The fourth column displayed the regulators and sequential order in the corresponding strain. Data are mean values of three independent experiments. Basal values are indicated in MU.
Assembly of DNA segments bearing regulatory elements
A mobile DNA fragment bearing the xylS2 gene under the control of NahR and its cognate promoter Psal was constructed as follows. First, the xylS2 gene was excised from plasmid pERD2 (20) as a 1.5 kb BamHI fragment and cloned in vector pTZ18 to give rise to pVLT5, in which xylS2 transcription is codirectional with the Plac promoter of the vector. The insert was then recloned as an EcoRI–HindIII fragment in phagemid vector pCG1 (21) and further subjected to site-directed mutagenesis (22) to insert a new NcoI site overlapping the first ATG of its coding sequence. The resulting plasmid (pS2) was then digested to completion with HindIII and partially with NcoI to release a 1.2 kb NcoI–HindIII fragment spanning the entire xylS2 sequence but lacking any sequence upstream of the first structural codon. This fragment was then cloned in plasmid pFH2 (kindly provided by S. Fernández), which coupled the leading ATG of the xylS2 gene with the optimized translation initiation region of the ner gene of phage Mu and flanked the whole promoterless gene sequence with NotI sites (23). Such a DNA segment was excised from the resulting plasmid (pNS2), and inserted in the unique NotI site of pCNB4 (24), to yield plasmid pCNB4-S2. This construct placed expression of xylS2 under the control of the Psal promoter and its cognate salicylate-responsive regulator NahR, all assembled in a mini-Tn5 Km transposon vector.
Plasmids equivalent to pCNB4-S2, but with the wild-type nahR gene replaced by its variants nahR3 and nahR4 (encoding benzoate-responsive mutants; 17), were made by cloning the NotI insert of pNS2 (which carries the promoterless xylS2 as mentioned above) in the single NotI sites of plasmids pCNB43 and pCNB44, described previously. These derived plasmids, pCNB43-S2 and pCNB44-S2, are the delivery plasmids for mini-transposon mini-Tn5 nahR3Psal→xylS2 and mini-Tn5 nahR4Psal→xylS2, respectively. Finally, mini-transposon mini-Tn5 nahR4/Psal→xylS, equivalent to mini-Tn5 nahR4/Psal→xylS2 but expressing the wild-type xylS gene under the control of the Psal promoter, was produced by first exchanging NcoI inserts between pNS2 and pCNB1 (originating plasmid pNS) and then excising and cloning the resulting NotI segment into the NotI site of pCNB44. This gave rise to delivery plasmid pCNB44-S. Plasmid pCNB44-lacZ, bearing the mobile elements mini-Tn5 nahR4/Psal→lacZ, has been described previously (17).
A XylS-responsive and XylS2-responsive reporter segment bearing a transcriptional fusion Pm→lacZ was also constructed by exchanging the 2.3 kb XbaI fragment of plasmid pUT/TcPm→xylX′::′lacZ (12) for the 3.7 kb XbaI fragment of pCNB1-lacZ (24). This exchange gives rise to a delivery plasmid (pTPmlac) for a TcR mini-Tn5 element which bears the exact same Pm→lacZ transcriptional fusion to a reporter trp′::′lacZ gene as that of pCNB2-lacZ. As shown below, this allowed us to compare faithfully the activity of the Pm promoter controlled by either a simple regulator or by two regulators coupled in a cascade.
For the construction of a mobile DNA fragment bearing the nahR gene under the control of XylS2 and its cognate promoter Pm, the nahR gene was excised from plasmid pMS15 (25) as a 1.2 kb NcoI–HindIII fragment and cloned in pFH2. The resulting plasmid was digested with NotI to yield a fragment containing the promoterless nahR gene preceded by an optimized TIR. This fragment was cloned at the unique NotI site of pCNB2 (24), to yield plasmid pCNB2-NR. This construct placed expression of nahR under the control of the Pm promoter and its salicylate-responsive regulator XylS2, assembled in a mini-Tn5 Km transposon vector. The matching NahR-responsive element was produced by the mini-Tn5 Sm Psal→nahG′::′lacZ of pTSAL1.
Mobilization and transposition
Each of the DNA segments produced by the plasmids described above and assembled in mini-Tn5 transposon vectors were targeted to the chromosome of E.coli CC118 or P.putida KT2442 with the delivery system described in detail elsewhere (26), generally known as the pUT system. Donor plasmids (Table 1) were transformed into a λpir lysogen of the E.coli S17-1 strain, which contains a chromosomally-integrated RP4 derivative providing conjugal transfer functions (27). Biparental matings between E.coli S17-1λpir derivatives and the recipient strain were set on nitrocellulose filters as described elsewhere (26). To discriminate authentic transposition from cointegration, exconjugant colonies were replica-plated on media with ampicillin or piperacillin to screen for the loss of the bla gene present in the delivery plasmid (27). Strains bearing insertions arising from different transposition events did not differ significantly with regard to the inducibility or absolute values of β-galactosidase activity and, therefore, only one of them (Table 1) was selected in each case for studies on gene expression.
Protein techniques
Western blot assays to detect the XylS product were performed as described elsewhere (28). To this end, equal amounts of whole E.coli or Pseudomonas cells (typically 108) were lysed in a sample buffer with 2% SDS and 5% β-mercapthoethanol and run in denaturing 12% polyacrylamide gels. These were subsequently blotted and probed with a 1:1000 dilution of a pre-adsorbed rabbit serum raised against purified inclusion bodies of the XylS protein (kindly provided by G. Bertoni). The bands in the blots corresponding to XylS were developed with 1 µg/ml of Protein A coupled to alkaline phosphatase 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium (BCIP/NBT, Sigma).
RESULTS
Rationale for the design of a transcriptional cascade based on nahR and xylS2
We tested the possibility of amplifying the promoter/cell response to a specific compound by connecting the activity of two regulators, in response to the same signal, through the design of a cascade regulatory circuit. We employed two salicylate-responsive transcriptional regulators of P.putida to analyze experimentally the conditions under which two activators could be coupled to each other to multiply the separate response of each activator to a given inducer. These regulators, NahR and XylS2, belong to different families of prokaryotic regulators (LysR and AraC, respectively) and the mechanisms of transcriptional activation are different, yet the effector profile of each regulator overlaps by several compounds, including salicylate (16,17). One key feature of XylS and its effector-specificity variants (such as XylS2) is that its cognate promoter Pm can be triggered by either a low level of the protein in the presence of an inducer or by high levels of the regulator without inducer (11). For a fixed XylS concentration, the activity of Pm can be induced up to 100-fold in P.putida. However, the range of Pm in vivo activity seemed to be even broader, since varying the XylS intracellular concentration using different constructs could achieve a range of 1000-fold of Pm activity (12). We reasoned that maximal XylS/Pm transcriptional capacity could be obtained when xylS expression is inducible, as part of the secondary regulator in a transcriptional cascade. On this basis, we reasoned that coupling expression of xylS2 to the NahR-dependent Psal promoter should result in a cascade with a potential to multiply the activity of Pm in the presence of salicylate (Fig. 1). This is because the inducer should both lead to the overproduction and activation of XylS2, thus the two effects could cooperatively contribute to the activation of transcription of the same promoter. These notions were put to the test by placing the relevant regulatory elements of P.putida into the chromosome of E.coli hosts, where all the features of the transcriptional control can be faithfully reproduced.
Figure 1.

Schematic representation of the cascade regulatory circuit. An expression cassette containing the nahR gene and the Psal promoter controls the expression of xylS2. In the presence of a common inducer, NahR activates the expression of xylS2. Simultaneous high intracellular levels of XylS2 and activation of XylS2 intrinsic transcriptional activity could achieve amplified expression levels of a particular gene(s) under the control of the Pm promoter.
Amplified gene expression capacity resulting from a coupled NahR–XylS2 system inducible by salicylate
To ascertain whether NahR and XylS2 could coordinately function to expand the responsiveness of Pm to salicylate we produced a set of isogenic E.coli strains, differing only in the nature and arrangement of the regulatory elements controlling lacZ expression, and inserted them into the chromosome with mini-Tn5 transposon vectors. To this end, the accumulation of β-galactosidase in response to inducer addition was examined in single-inserted E.coli strains RSL9 (nahR/Psal→lacZ) and FH26 (xylS2/Pm→lacZ) as well as in the double-inserted strain E.coli 4S2PT32 (nahR/Psal→xylS2, Pm→lacZ). The results shown in Figure 2 demonstrate the multiplicative effect of coupling the two regulators. While β-galactosidase accumulation by the single-inserted strains ranged between 65 (average of the dynamic uninduced values) and 2100 U (induced) in strain RSL9 (nahR/Psal→lacZ) and between 150 (non-induced) and 2300 U (induced) in FH26 (xylS2/Pm→lacZ), the induced levels of the reporter enzyme increased by almost 20-fold in the double-inserted strain E.coli 4S2PT32 (nahR/Psal→xylS2, Pm→lacZ), while maintaining equivalent basal levels [~130 Miller Units (MU) average of dynamic values during the 24 h incubation]. This caused the overall induction ratio to be 180–240-fold. In fact, the accumulation over a few hours of about 40 000 β-galactosidase units raised from a single chromosomal promoter revealed the extraordinary strength of the Pm promoter when properly activated. Another finding from the experiment shown in Figure 2 was the instant coupling of the two regulators that showed the augmented effect at the earliest time of measurements, even 10 min after salicylate addition (data not shown). It thus became apparent that the gene expression capacity (ratio of induced level to basal level) of the Pm promoter in response to salicylate underwent a dramatic upshift when it was subjected to an activation cascade, but without any significant qualitative effect on the induction kinetics.
Figure 2.

Kinetics of β-galactosidase production by the three expression systems. Salicylate (2 mM) was added to the cultures (OD600 = 0.2), and β-galactosidase activity was monitored at different intervals. Open symbols, cultures without salicylate. Closed symbols, cultures with 2 mM salicylate. nahR/Psal::trp′::′lacZ (squares), xylS2/Pm:trp′::′lacZ (triangles) and nahR/Psal→xylS2/Pm→trp′::′lacZ (circles).
Dissection of the synergic coupling of NahR and XylS2
To analyze the specific contribution of each regulator to the steps involved in the cascade, we exploited the fact that both regulators can be induced independently by specific effectors to each regulator, with little response in the other (Fig. 3). When β-galactosidase production from each system was measured in the absence or presence of different compounds that were effectors of both regulators, the induction ratio obtained with the cascade system, with respect to basal level, was always superior to the simple circuits (Fig. 3A and B). Figure 3C and D illustrates the results of subjecting E.coli strains RSL9, FH26 and 4S2PT32 to induction experiments with the XylS2-only inducer benzoate as well as with the NahR-only effector 3,5-dichlorosalicylate. Although only the addition of a shared inducer (i.e., salicylate) leads to maximal augmentation effect on promoter output, the combined system still showed amplification when the upstream regulator (NahR) could be activated. As shown in Figure 3C, 500 µM of 3,5-dichlorosalicylate, which had no effect on XylS2 activity, could induce the NahR-mediated expression of Psal::lacZ 19-fold. The same concentration of inducer caused Pm activity to increase ~76-fold in the combined system of strain 4S2PT32. The same trend was observed when the coupled system was exposed to other NahR-exclusive inducers such as 4-chlorosalicylate, which at 2 mM increased Pm activity up to 190-fold, as compared with the 30-fold increase observed for nahR/Psal→lacZ (data not shown). These data reflected the ability of XylS2 to activate Pm in an inducer-independent manner. Such a feature seemed sufficient to have a substantial amplification effect (~4-fold), indicating that at certain XylS2 concentrations, a cooperative effect during the transcriptional activation of Pm may occur. In contrast, benzoate, which has no effect on NahR, promoted a 40-fold induction of Pm regardless of whether the strain used had the single xylS2/Pm→lacZ assembly or the coupled nahR/Psal→xylS2 and Pm→lacZ system. Immunoblots of the E.coli strain 4S2PT32 confirmed that XylS2 production was induced when using an effector molecule for NahR but not when using benzoate (Fig. 3E). These results suggest that the mere addition of an upstream regulator results in a greater amplification.
Figure 3.
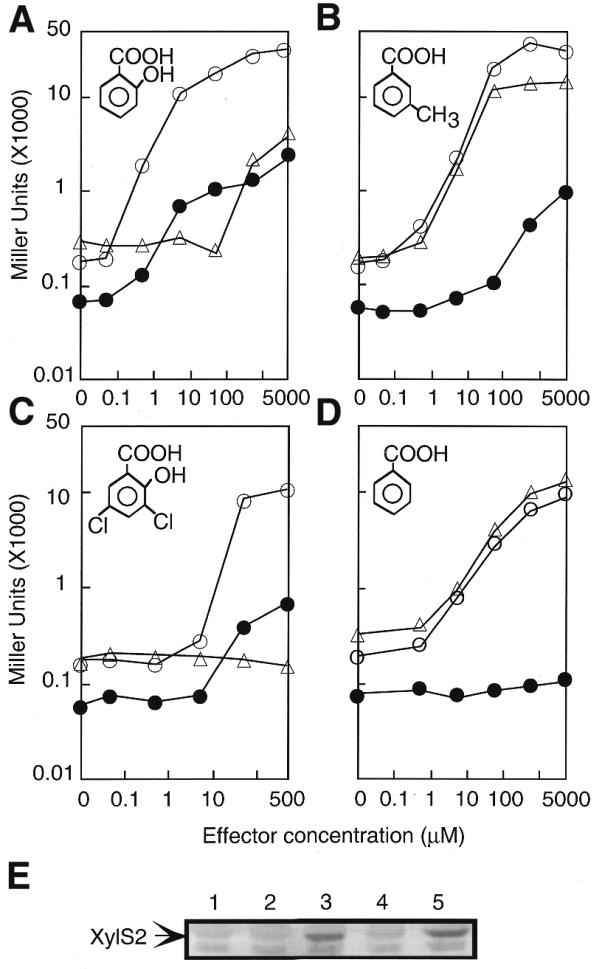
Gene expression analysis of the simple and cascade systems with different effectors. β-galactosidase activity from the three systems at different concentrations of salicylate (A), 3-methylbenzoate (B), 3,5-dichlorosalicylate (C) and benzoatSe (D). Closed circles, nahR/Psal→trp′::′lacZ; triangles, xylS2/Pm→trp′::′lacZ; open circles, nahR/Psal→xylS2/Pm→trp′::′lacZ. Note that the maximun concentration used for 3,5-dichlorosalicylate in the β-galactosidase assays was 0.5 mM because concentrations >2 mM resulted in growth inhibition. (E) Immunodetection of the XylS2 product in a control culture of CC118 (lane 1), and in the cascade-containing strain CC1184S2PT32 upon incubation without inducer (lane 2), or with 1 mM 3,5-dichlorosalicylate (lane 3), benzoate (lane 4) and salicylate (lane 5).
The performance of the XylS2/NahR cascade relies on a determined hierarchy of regulators
To ascertain the requirement of regulatory hierarchy in the cascade amplification, we studied the effect of swapping the upstream and downstream regulators in the XylS2/NahR-based cascade by constructing E.coli 2NRSL7. The chromosome of this strain bears the DNA elements xylS2/Pm→nahR and Psal→lacZ carried by specialized transposons (see above) and is thus equivalent to E.coli strain 4S2PT32, apart from the order of the regulators in the coupled system. As shown in Figure 4, the capacity of the reverse-coupled system responding to salicylate (24-fold induction) did not increase the capacity over the single nahR/Psal→lacZ element. Along the same lines, the reversed-coupled system of E.coli 2NRSL7 was completely insensitive to the XylS2-only effector benzoate. These observations indicated that overexpression of nahR did not result in a parallel increase in Psal activity, but in a non-productive excess of the second regulator because the same gene expression capacity from Psal could be achieved at a relatively low concentration of NahR. Studies on the mechanism of Psal activation indicated that the target site for this activator is occupied regardless of the induction conditions (29). Thus, the Psal promoter appears to depend exclusively on the presence or absence of salicylate and overexpression of nahR does not produce higher promoter activity. Nevertheless, the basal expression of nahR from the heterologous xylS2/Pm system seemed to limit the available NahR protein necessary to activate Psal promoter, since the xylS2/Pm→nahR/Psal→lacZ cascade could only be induced 4-fold after the NahR-unique inducer 3,5-dichlorosalicylate. In contrast, the same compound induced 20-fold lacZ expression from nahR/Psal.
Figure 4.

Capacity for regulation of simple circuits and with two cascades using a different hierarchy of upstream and downstream regulators. S, 2 mM salicylate; B, 2 mM benzoate; 3,5dClS, 2 mM 3,5-dichlorosalicylate. Basal values of β-galactosidase activity from each circuit established in E.coli were the following: nahR/Psal, 65 MU (black bars); xylS2/Pm, 192 MU (open bars); nahR/Psal→xylS2/Pm, 169 MU (gray bars); xylS2/Pm→nahR/Psal, 69 MU (hatched bars). Data are the mean values of three independent experiments. The corresponding standard deviations are indicated with error bars.
Taken together, these results indicate that the efficiency of the amplification of the regulatory cascade requires specific properties of the downstream regulator/promoter which include, at least, the activation of the final target promoter in a dose-dependent fashion for a broad range of regulator concentrations. This is true for XylS2 but not for NahR. Thus, the mere sensitivity of the two regulators to the same effector did not result in an augmentation effect unless the adequate hierarchy, based on the appropriate mechanisms of the second regulatory system, was present.
Redesigning a cascade circuit for benzoate hyper-response in P.putida
In view of the results above, and given the ease of obtaining both XylS and NahR mutants responsive to a large number of structural homologs of the natural effectors (16,17), it became possible to construct regulatory cascades, as required, to amplify some cell responses to predetermined inducers. In this context we pursued the design of a specialized P.putida strain bearing a benzoate cascade control circuit. Since the downstream regulator (XylS2) already responded to this inducer (Fig. 5), the design of a new cascade involved mainly the modification of the upstream regulatory system. To this end, we employed two nahR mutants encoding benzoate-responsive variants nahR3 and nahR4 (17). They were assembled in the coupled system nahR3/Psal→xylS2 and nahR4Psal→xylS2 along with the reporter segment Pm→lacZ and then inserted into the chromosome of P.putida strain KT2442 to yield P.putida 43S2PmL and P.putida 44S2PmL. The induction of these strains by benzoate was compared to that of strains bearing either the simple elements P.putida 43L (nahR3/Psal→lacZ), P.putida 44L (nahR4/Psal→lacZ) and P.putida S2PmL (xylS2/Pm→lacZ), or the cascade with the wild-type benzoate-insensitive nahR variant (Fig. 5). The cascade with the benzoate-responsive nahR mutants increased the gene expression capacity from 6- to 35-fold in response to benzoate, compared with the single expression systems with nahR3, nahR4 or xylS2. In contrast, the cascade circuit with the wild-type nahR in 4S2PmL (nahR/Psal→xylS2, Pm→lacZ) in Pseudomonas showed reduced induction capacity by benzoate (∼7-fold), due to the absence of response of the upstream regulator to benzoate. These values matched the predictions raised from the simultaneous induction of each regulator upon acquisition of the ability to respond to benzoate by NahR.
To eliminate the possibility that the amplification property was only a particular feature of the mutant form of XylS (XylS2) and not of the wild type, we constructed a regulatory circuit with the nahR4 mutant as a first regulator and xylS/Pm as a second regulator. We tested the inducibility of the circuits in response to the common effector benzoate and salicylate that is not recognized by the XylS. Augmentation of gene expression capacity was produced when the common effector was added but not when salicylate was present (Fig. 6). The regulatory scheme nahR4/Psal→xylS plus Pm→lacZ showed at least 4-fold more induction capacity in response to benzoate than the single circuits, nahR4/Psal→lacZ (Fig. 5) or xylS/Pm→lacZ (Fig. 6). Western blotting of P.putida nahR/Psal→xylS and Pm→lacZ cells exposed to each of the inducers revealed that the increase in XylS production brought about by salicylate was insufficient to match the amplification effect caused by benzoate even at lower intracellular concentrations of the regulator (Fig. 6). Therefore, the simultaneous response of the first and second regulators was also required in the natural host for the amplification effect using a wild-type xylS.
Figure 6.
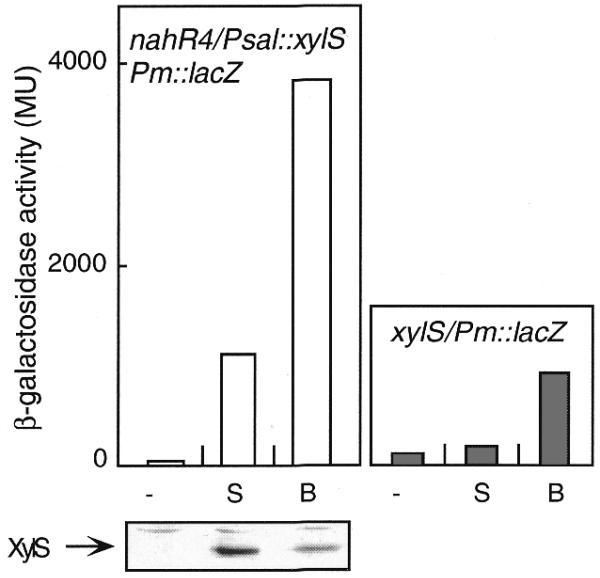
Analysis of a cascade circuit with nahR4/Psal leading to the expression of wild-type xylS in P.putida. β-galactosidase accumulation after addition of no effector (–), NahR4-specific effector (S, salicylate) or NahR4/XylS common effector (B, benzoate) at 2 mM in the cascade system (left) or a xylS/Pm→trp′::′lacZ simple circuit (right). A western blot for detection of XylS production in P.putida (nahR4/Psal→xylS/Pm→trp′::′lacZ) cultures after addition of the different effectors is shown under the corresponding graph.
DISCUSSION
In this work, we have shown how two regulators can amplify the response to a specific signal by combining the regulatory elements in a cascade circuit. Optimal coupling between the XylS and NahR regulators for the synergistic effect in gene expression required that (i) the downstream regulator activates the target promoter in a dose-dependent manner over a broad range of intracellular concentration, and (ii) both activators respond positively to the same inducer.
The effectiveness of the amplification by the analyzed cascade depends on a previous sensing of the signal molecule to lead the expression of the downstream regulator. The natural cascade regulatory circuit in which XylS is involved, implies the control of xylS expression through the XylR regulatory protein. This protein induces xylS expression and the upper TOL operon in response to m-xylene or toluene, which are the substrates of the upper pathway. The products of catabolism of the upper pathway (benzoate or m-toluate) are effectors of the XylS protein. This regulator induces the expression of the meta-operon, whose products degrade benzoate to three carboxylic acids. It is then expected that for the efficient regulation, the upstream regulator (XylR) senses first the signal that triggers all the cascade event through the downstream regulator, XylS. The XylR regulator may prepare the meta-operon for a high induction to the upper-pathway benzoate products by increasing the levels of the XylS regulator, and thus, increasing the response to the catabolytes that will be produced by the upper operon products. In fact, results of Marqués et al. (13) showed that mRNA produced from the Pm promoter, including the upper-pathway inducer-substrate, was 2-fold higher than with a meta-pathway substrate-effector.
Although maximal amplification of gene expression capacity occurred when the signal inducer was positive for both regulators, the inducer-independent activitation of Pm by XylS2 can also augment gene expression capacity from the upstream regulator in E.coli. Increment of XylS2 concentration due to the presence of a NahR-only inducer, multiply the gene expression capacity reached by nahR/Psal ~4-fold. An increment over the unit may indicate cooperative effect in the activation of Pm with the inducer independent XylS2 activity. However, in P.putida the constitutive transcriptional activity of XylS did not show such amplification of gene expression capacity since induction of β-galactosidase activity from the nahR/Psal→xylS/Pm→lacZ cascade by the NahR4-only inducer salicylate did not result in augmentation of gene expression capacity with respect to the single system. This might be explained by the superior constitutive activity of XylS2 with respect to XylS or because of different specific activity of these regulators in E.coli, with respect to P.putida, or both. It is noteworthy that the basal value of the Pm promoter is considerably lower in P.putida than in E.coli, indicating that there are differences in the specific activities of the regulatory elements in each background.
The comparison of 30 different prokaryotic regulatory circuits (30) revealed that the rhamnose-responsive system of E.coli presents the maximum gene expression capacity known in a natural system. Interestingly, this system works in a similar manner to the artificial cascade described in this work. Rhamnose-responsive promoters are induced by this sugar through the action of two activators of the AraC/XylS family called RhaR and RhaS. Each activator can separately stimulate transcription of their cognate promoters by 300-fold in the presence of the carbohydrate. Interestingly, RhaR controls the expression of the rhaS gene, whose product triggers rhamnose catabolism (31). The coupling of RhaR and RhaS allows a 30 000-fold induction of the rhamnose operon, which results from the cooperative stimulation with the same signal.
The efficiency of the amplification we observed in the NahR/XylS2 combination might not be that different to the fact that several naturally occurring regulatory cascades include an activator of the AraC family as a downstream regulator. For instance, a part of XylS itself in the TOL system, the VirF product of Vibrio cholerae, as well as the RhaS and SoxS regulators of E.coli, act as the second regulator in different circuits (32,33). The occurrence of XylS-related regulators as downstream regulatory factors described above may suggest the existence of a type of regulator whose expression is eventually controlled by a first or ‘master’ regulator and that might act as an amplifier in different control circuits. It is conceivable that at least part of the AraC/XylS family of regulators might share the signal amplifier potential shown for XylS and RhaR/S.
The degree of precision in the knowledge of the mechanisms and factors for gene expression control is allowing the design of artificial regulatory circuits to engineer new functions (34,35). The possibility of engineering the amplification of gene expression capacity in response to environmental signals by cascade circuits with the positive regulators shown might be used for the design of new expression systems, whole cell biosensors or control circuits for gene therapy requiring reduced basal levels and high induced levels.
Acknowledgments
ACKNOWLEDGEMENTS
The authors are indebted to G. Bertoni (University of Milan) for anti-XylS antibodies and gratefully acknowledge E. Santero, J. Pérez-Martín and S. Marqués for critical reading of the manuscript. This work was supported by contracts ENV4-CT95-0141 and BIO4-CT97-2040 from the EU and by grant BIO98-0808 from the Plan Nacional de Investigación (CICYT).
References
- 1.Stragier P. and Losick,R. (1990) Cascades of sigma factors revisited. Mol. Microbiol., 4, 1801–1806. [DOI] [PubMed] [Google Scholar]
- 2.Banuett F. (1998) Signalling in the yeasts: an informational cascade with links to the filamentous fungi. Microbiol. Mol. Biol. Rev., 62, 249–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fischer H.M. (1994) Genetic regulation of nitrogen fixation in rhizobia. Microbiol. Rev., 58, 352–386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dorman C.J. and Porter,M.E. (1998) The Shigella virulence gene regulatory cascade: a paradigm of bacterial gene control mechanisms. Mol. Microbiol., 29, 677–684. [DOI] [PubMed] [Google Scholar]
- 5.Harayama S. and Timmis,K.N. (1989) Catabolism of aromatic hydrocarbons by Pseudomonas. In Hopwood,D. and Chater,K. (eds), Genetics of Bacterial Diversity. Academic Press, London, UK, pp. 151–174.
- 6.Gallegos M.T., Schleif,R., Bairoch,A., Hofmann,K. and Ramos,J.L. (1997) AraC/XylS family of transcriptional regulators. Microbiol. Mol. Biol. Rev., 61, 393–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mermod N., Lehrbach,P.L., Reineke,W. and Timmis,K.N. (1984) Transcription of the TOL plasmid toluate catabolic pathway operon of Pseudomonas putida is determined by a pair of co-ordinately and positively regulated overlapping promoters. EMBO J., 3, 2461–2466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ramos J.L., Mermod,N. and Timmis,K.N. (1987) Regulatory circuits controlling transcription of TOL plasmid operon encoding meta-cleavage pathway for degradation of alkylbenzoates by Pseudomonas. Mol. Microbiol., 1, 293–300. [DOI] [PubMed] [Google Scholar]
- 9.Ramos J.L., Marqués,S. and Timmis,K.N. (1997) Transcriptional control of the Pseudomonas TOL plasmid catabolic operons is achieved through an interplay of host factors and plasmid-encoded regulators. Annu. Rev. Microbiol., 51, 341–373. [DOI] [PubMed] [Google Scholar]
- 10.Dixon R. (1986) The xylABC promoter from the Pseudomonas putida TOL plasmid is activated by nitrogen regulatory genes in Escherichia coli. Mol. Gen. Genet., 203, 129–136. [DOI] [PubMed] [Google Scholar]
- 11.Marqués S. and Ramos,J.L. (1993) Transcriptional control of the Pseudomonas putida TOL plasmid catabolic pathways. Mol. Microbiol., 9, 923–929. [DOI] [PubMed] [Google Scholar]
- 12.Kessler B., Herrero,M., Timmis,K.N. and de Lorenzo,V. (1994) Genetic evidence that the XylS regulator of the TOL meta-operon of Pseudomonas controls Pm promoter through weak DNA-protein interaction. J. Bacteriol ., 176, 3171–3176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Marqués S., Holtel,A., Timmis,K. and Ramos,J.L. (1994) Transcriptional induction kinetics from the promoters of the catabolic pathways of TOL plasmid pWW0 of Pseudomonas putida for metabolism of aromatics. J. Bacteriol ., 176, 2517–2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yen K.-M. and Serdar,C.M. (1988) Genetics of naphtalene metabolism in pseudomonads. Crit. Rev. Microbiol., 15, 247–268. [DOI] [PubMed] [Google Scholar]
- 15.Schell M.A. (1993) Molecular biology of the LysR family of transcriptional regulators. Annu. Rev. Microbiol., 47, 597–626. [DOI] [PubMed] [Google Scholar]
- 16.Ramos J.L., Stolz,A., Reineke,W. and Timmis,K.N. (1986) Altered effector specificities in regulators of gene expression: TOL plasmid xylS mutants and their use to engineer expansion of the range of aromatics degraded by bacteria. Proc. Natl Acad. Sci. USA, 83, 8467–8471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cebolla A., Sousa,C. and de Lorenzo,V. (1997) Effector specificity mutants of the transcriptional activator NahR of naphtalene degrading Pseudomonas define protein sites involved in binding of aromatic inducers. J. Biol. Chem., 272, 3986–3992. [DOI] [PubMed] [Google Scholar]
- 18.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 19.Miller J.H. (1972) Experiments in Molecular Genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 20.Ramos J.L., Carrero,M. and Timmis,K.N. (1988) Broad host range expression vectors containing manipulated meta-cleavage pathway regulatory elements of the TOL plasmid. FEBS Lett., 226, 241–246. [DOI] [PubMed] [Google Scholar]
- 21.Myers R.M., Lerman,L.S. and Maniatis,T. (1985) A general method for saturation mutagenesis of cloned DNA fragments. Science, 229, 242–247. [DOI] [PubMed] [Google Scholar]
- 22.Kunkel T.A., Roberts,J.D. and Zakour,R.A (1987) Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol., 154, 367–382. [DOI] [PubMed] [Google Scholar]
- 23.Kessler B., Timmis,K.N. and de Lorenzo,V. (1994) The organization of the Pm promoter of the TOL plasmid reflects the structure of its cognate activator protein XylS. Mol. Gen. Genet., 244, 596–605. [DOI] [PubMed] [Google Scholar]
- 24.de Lorenzo V., Fernández,S., Herrero,M., Jakubzik,U. and Timmis,K.N. (1993) Engineering of alkyl- and haloaromatic-responsive gene expression with mini-transposons containing regulated promoters of biodegradative pathways of Pseudomonas. Gene, 130, 41–46. [DOI] [PubMed] [Google Scholar]
- 25.Schell M. and Sukordhaman,M. (1989) Evidence that the transcriptional activator encoded by the Pseudomonas putida nahR gene is evolutionarily related to the transcriptional activators encoded by the Rhizobium meliloti nodD genes. J. Bacteriol ., 171, 1952–1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.de Lorenzo V. and Timmis,K.N. (1994) Analysis and construction of stable phenotypes in gram-negative bacteria with Tn5- and Tn10-derived minitransposons. Methods Enzymol., 235, 386–405. [DOI] [PubMed] [Google Scholar]
- 27.Herrero M., de Lorenzo,V. and Timmis,K.N. (1990) Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram negative bacteria. J. Bacteriol ., 172, 6557–6567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cebolla A., Guzman,C. and de Lorenzo,V. (1996) Non-disruptive detection of activity of catabolic promoters of Pseudomonas with an antigenic surface reporter system. Appl. Environ. Microbiol., 62, 214–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Huang J.Z. and Schell,M.A. (1991) In vivo interactions of the NahR transcriptional activator with its target sequences. Inducer-mediated changes resulting in transcription activation. J. Biol. Chem., 266, 10830–10838. [PubMed] [Google Scholar]
- 30.Hlavacek W.S. and Savageau,M.A. (1996) Rules for coupled expression of regulator and effector genes in inducible circuits. J. Mol. Biol., 255, 121–139. [DOI] [PubMed] [Google Scholar]
- 31.Egan S.M. and Schleif,R.F. (1993) A regulatory cascade in the induction of rhaBAD. J. Mol. Biol., 234, 87–98. [DOI] [PubMed] [Google Scholar]
- 32.DiRita V.J., Parsot,C., Jander,G. and Mekalanos,J.J. (1991) Regulatory cascade controls virulence in Vibrio cholerae. Proc. Natl Acad. Sci. USA, 88, 5403–5407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gallegos M.T., Michán,C. and Ramos,J.L. (1993) The XylS/AraC family of regulators. Nucleic Acids Res., 21, 807–810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Elowitz M.B. and Leibler,S. (2000) A synthetic oscillatory network of transcriptional regulators. Nature, 403, 335–338. [DOI] [PubMed] [Google Scholar]
- 35.Gardner T.S., Cantor,C.R. and Collins,J.J. (2000) Construction of a genetic toggle switch in Escherichia coli. Nature, 403, 339–342. [DOI] [PubMed] [Google Scholar]
- 36.de Lorenzo V., Eltis,L., Kessler,B. and Timmis,K.N. (1993) Analysis of the Pseudomonas gene products using lacIq/Ptrp-lac plasmids and transposons that confer conditional phenotypes. Gene, 123, 17–24. [DOI] [PubMed] [Google Scholar]
- 37.Minton N.P. (1984) Improved plasmid vectors for the isolation of translational lac gene fusions. Gene, 31, 269–273. [DOI] [PubMed] [Google Scholar]
- 38.de Lorenzo V., Herrero,M., Jakubzik,U. and Timmis,K.N. (1990) Mini-Tn5 transposon derivatives for insertion mutagenesis, promoter probing and chromosomal insertion of cloned DNA in gram negative eubacteria. J. Bacteriol ., 172, 6568–6572. [DOI] [PMC free article] [PubMed] [Google Scholar]