


Abstract
The C-terminal domain (CTD) of the large subunit of RNA polymerase II plays a role in transcription and RNA processing. Yeast ESS1, a peptidyl-prolyl cis/trans isomerase, is involved in RNA processing and can associate with the CTD. Using several types of assays we could not find any evidence of an effect of Pin1, the human homolog of ESS1, on transcription by RNA polymerase II in vitro or on the expression of a reporter gene in vivo. However, an inhibitor of Pin1, 5-hydroxy-1,4-naphthoquinone (juglone), blocked transcription by RNA polymerase II. Unlike N-ethylmaleimide, which inhibited all phases of transcription by RNA polymerase II, juglone disrupted the formation of functional preinitiation complexes by modifying sulfhydryl groups but did not have any significant effect on either initiation or elongation. Both RNA polymerases I and III, but not T7 RNA polymerase, were inhibited by juglone. The primary target of juglone has not been unambiguously identified, although a site on the polymerase itself is suggested by inhibition of RNA polymerase II during factor-independent transcription of single-stranded DNA. Because of its unique inhibitory properties juglone should prove useful in studying transcription in vitro.
INTRODUCTION
Peptidyl-prolyl cis/trans isomerases (PPIases) are able to accelerate the cis/trans isomerization of peptide bonds preceding prolyl residues and have diverse functions (1,2). Recently ESS1, an essential yeast PPIase, was isolated from a genetic screen designed to identify trans-acting factors involved in mRNA 3′-end processing (3). ESS1 belongs to a family of PPIases that includes Escherichia coli parvulin (4), Drosophila dodo (5), Arabidopsis thaliana Pin1 (6) and human Pin1 (7). All eukaryotic parvulin family members have in common the presence of a WW domain that is involved in the recognition and association of the conserved motifs, phosphoserine-proline and phosphothreonine-proline (8), and a PPIase domain that contains its enzymatic activity. The C-terminal domain (CTD) of the large subunit of RNA polymerase II contains from 26 to 52 copies of the sequence YSPTSPS (9). This essential domain becomes hyperphosphorylated during the transition from initiation to elongation (9) and acts as a scaffold for a number of RNA processing enzymes (10). Consistent with its role in RNA processing, ESS1 associated with the hyperphosphorylated CTD of the large subunit of RNA polymerase II (11). However, the mechanism by which ESS1 exerts its effect on RNA processing is still not clear.
Juglone, 5-hydroxy-1,4-naphthoquinone, inhibits the members of the parvulin PPIase family and functions by modifying sulfhydryl groups (12). Juglone inactivates the E.coli PPIase, parvulin, by covalent modification of two cysteine residues in a slow process (12). The activity of members of the other PPIase families is not affected by juglone at all (12). While the inhibition of parvulin family members by juglone is somewhat specific, inhibition of other enzymes (pyruvate decarboxylase and glutathione S-tranferase) has been reported (13).
Because phosphorylation of the CTD is involved in controlling transcription and Pin1 associates with the phospho-CTD, we decided to explore the potential role of Pin 1 in transcription. Phosphorylation of the CTD occurs after initiation and is associated with the transition into productive elongation mediated by the CTD kinase activity of P-TEFb (14). This elongation control process requires the CTD as well as a number of negative factors that limit the elongation potential of RNA polymerase II (15,16). It is likely that there is an exchange of factors functionally associating with the CTD during a single round of transcription from those required for initiation to those required for controlling elongation and, finally, to those involved in RNA processing. We reasoned that conformational changes in the CTD resulting from the action of the PPIase activity of Pin1 might facilitate or control this factor exchange. To determine if Pin1 played a role in transcription we first examined the effect of the Pin1 inhibitor in vitro. Juglone clearly inhibited transcription, but the results indicated that the effects seen were not mediated by inhibition of Pin1. Our results indicate that juglone directly inhibits transcription of RNA polymerase I, II and III. Unlike N-ethylmaleimide (NEM), which non-specifically blocked all phases of transcription, we show that inhibition of RNA polymerase II by juglone is accomplished by disrupting the preinitiation step of transcription with only a slight effect on initiation or elongation. Because of this unique property juglone should be a useful tool in studying transcription.
MATERIALS AND METHODS
Materials
[α-32P]CTP (3000 Ci/mmol) was from ICN. Ribonucleoside triphosphates were from Pharmacia Biotech Inc. Juglone (Aldrich) and NEM (Sigma) were dissolved in ethanol to 10 mM and 1 M, respectively, and both were stored at –80°C. All other chemicals were reagent grade.
Generation and purification of the recombinant Pin1 proteins
The coding region (492 nt) of human Pin1 was amplified by reverse transcription (RT)–PCR using two primers (5′-GGAGGAGCATATGGCGGACGAGGAGAAG and 5′-CCAGTCTCGAGTCACTCAGTGCGGAGGAT) and total RNA of HeLa cells. The amplified fragment was digested with NdeI and XhoI and cloned into a pET21a vector (Novagen) to generate pET21a-Pin1. A His-tagged Pin1 construct was then created by PCR amplification of the Pin1 coding region from pET21a-Pin1 using two primers (5′-GGAGGAGCATATGGCGGACGAGGAGAAG and 5′-CCAGTCTCGAGCTCAGTGCGGAGGATGA). The resulting fragment was digested with NdeI and XhoI and cloned into pET21a to generate a plasmid (pET21a-Pin1-His). A similar approach was performed to generate a plasmid (pET21a-Pin1WW-His) for expression of the His-tagged Pin1WW domain (amino acids 1–44) using pET21a-Pin1 as the template and two primers (5′-GGAGGAGCATATGGCGGACGAGGAGAAG and 5′-AAACTCGAGACCACTGCTGCTGTTGCCGCT).
To generate the construct expressing the knockout Pin1 protein (pET21a-Pin1H59A-His) (17), which contains a substitution of alanine for histidine at position 59, site-directed mutagenesis was performed according the Stratagene manual (QuikChange Site-Directed Mutagenesis Kit) using two mutagenic primers (5′-GCCAGGGTCCGCTGCTCGGCCCTGCTGGTGAAGCACAGC and 5′-GCTGTGCTTCACCAGCAGGGCCGAGCAGCGGACCCTGGC). The mutation was confirmed by sequencing.
Three plasmids, pET21a-Pin1-His, pET21a-Pin1WW-His and pET21a-Pin1H59A-His, were used to transform E.coli BL21-CodonPlus(DE3)-RIL competent cells (Stratagene). Ni2+-NTA–agarose column (Qiagen) was utilized to purify the expressed Pin1 proteins as described in the manufacturer’s manual. The purified proteins were analyzed by SDS–PAGE and visualized by silver staining.
Generation of eukaryotic Pin1 expression constructs
The Pin1/Pin1H59A (492 nt) coding regions were amplified using pET21a-Pin1/pET21a-Pin1H59A as the templates and two primers (5′-AAATCTAGAATGGCGGACGAGGAGAAGCTG and 5′-AAAGAATTCCTCACTCAGTGCGGAGGATGAT). The amplified 0.5 kb fragment was digested with XbaI and EcoRI and cloned into pCDNA3-HA [containing a cytomegalovirus (CMV) promoter and HA-tag on the N-terminus] to generate the plasmids (pCDNA3-HA-Pin1 and pCDNA3-HA-Pin1H59A). A similar approach was used to generate pCNDA3-HA-Pin1WW using pET21a-Pin1 as the template and two primers (5′-GGAGGAGCATATGGCGGACGAGGAGAAG and 5′-AAACTCGAGACCACTGCTGCTGTTGCCGCT).
RNA polymerase II in vitro transcription assay
Pulse–chase transcription assays were carried out as described (18). The CMV template was linearized with EcoRI to give a 660 nt run-off transcript. Transcription reactions (20 µl) contained 1.5 µl HeLa cell nuclear extract, 0.75 µl recombinant P-TEFb (comprised of Cdk9 and cyclin T1), 20 mM HEPES, pH 7.6, 7 mM MgCl2, 60 mM KCl, 600 µM each of ATP, GTP and UTP, and 5 µCi of [α-32P]CTP and were incubated for 15–20 min for preincubation, 30 s for pulse, 7 min for chase at 30°C. Conditions during the chase were identical to those during the pulse except that the CTP was raised to 1.2 mM. The reactions were stopped after the pulse or chase as indicated. Pin1, Pin1H59A and Pin1WW proteins, NEM and juglone were included as indicated. Labeled transcripts were analyzed on 6 or 9% denaturing gel. The extent of phosphorylation was quantitated from dried gels using a Packard InstantImager. The IC50 values were calculated by fitting the data to a logistic dose–response curve using the program TableCurve (Jandel Scientific).
RNA polymerase I and III in vitro transcription assays
RNA polymerase I transcription reactions were performed as previously described with few modifications (19). A plasmid with a ribosomal RNA gene promoter (pHr160) was linearized with EcoRI giving a 681 nt run-off transcript. Transcription reactions (16 µl) contained 1.6 µl HeLa cell nuclear extract, 20 mM HEPES, pH 7.6, 7 mM MgCl2, 50 mM KCl, 200 µM each of ATP, GTP and UTP, 10 µM CTP and 5 µCi of [α-32P]CTP, and were incubated for 1 h at 30°C. RNA polymerase III transcription reactions were performed as described (20). A basal Alu construct, Alu-T, was used as the template. Transcription reactions (16 µl) contained 1.5 µl HeLa cell nuclear extract, 20 mM HEPES, pH 7.6, 7 mM MgCl2, 100 mM KCl, 600 µM each of ATP, GTP and UTP, 40 µM CTP and 10 µCi of [α-32P]CTP, and were incubated for 1 h at 30°C. Labeled transcripts were analyzed on 6% denaturing gel. Data were quantitated as described in the previous section.
Transient cotransfection assay
HeLa cells were grown to 50–80% confluence in the 24-well plates and transfected with LipofectAmine Plus Reagent as described in the manufacturer’s manual (Gibco BRL). To investigate the role of Pin1 during transcription, HeLa cells were transiently transfected with 0.1 µg CMV-luciferase (CMV-luc) and different amounts of a Pin1 expression vector (0.1, 0.2 or 0.4 µg of pCDNA3-HA-Pin1 or pCDNA3-HA-Pin1H59A or pCDNA3-HAPin1WW). Luciferase activity was measured 24 h after transfection with the Promega luciferase assay system and the activity was detected with a Zylux FB12 luminometer. The IC50 was determined as described above.
Restriction enzyme digestion
Plasmid pCDNA3-HA (Promega) was digested by BamHI, EcoRI, KpnI, XbaI, HindIII, ScaI and SalI with or without the addition of juglone (50 µM) at 37°C for 1 h, and analyzed on 1% agarose gel. Except SalII, which cut pCDNA3-HA twice, the other enzymes cut the plasmid only once.
T7 RNA polymerase assay
pET21a was linearized with BamHI resulting in a template giving a 112 nt run-off transcript. Each reaction (25 µl) contained 20 U of T7 RNA polymerase (New England Biolabs), 1× RNA polymerase buffer (New England Biolabs), 85 ng linearized DNA template, 2 µg BSA, 200 µM GTP, UTP, ATP, 10 µM CTP, 5 µCi of [α-32P]CTP, and was incubated at 37°C for 15 min. Labeled transcripts were analyzed on 6% denaturing gel.
RNA polymerase II assay
The general RNA polymerase assay was performed as described previously except that 2.5 µCi [α-32P]CTP was used as label (21). Purified Drosophila RNA polymerase II (14) was preincubated with juglone for 2 min. Denatured sheared salmon sperm DNA was used as the template.
RESULTS
Juglone inhibits RNA polymerase II transcription in vitro
We started our investigation of the involvement of Pin1 in RNA polymerase II transcription by examining the effect of the Pin1 inhibitor juglone. An in vitro assay using a template containing the CMV promoter, HeLa nuclear extract and recombinant P-TEFb (comprised of Cdk9 and cyclin T1) was performed. The inclusion of increasing concentrations of juglone in the reaction resulted in the inhibition of the 660 nt run-off transcript (Fig. 1A). The radioactivity in run-off transcripts was quantitated and the IC50 for the inhibitory effect was determined to be 7 µM (Fig. 1B). The data indicate that juglone is an effective inhibitor of RNA polymerase II transcription and suggest that Pin1 may be involved. To determine what effect juglone might have on transcription in vivo we incubated cells with increasing concentrations of the drug for 24 h. Consistent with a general effect on transcription, 30 or 100 µM juglone caused significant abnormalities in cell morphology and cell death (data not shown). To circumvent this problem of toxicity we attempted a nuclear run-on assay on cells treated for only 1 h with 50 µM juglone. Unfortunately, juglone treatment caused the cells to be refractory to lysis during homogenization (data not shown).
Figure 1.
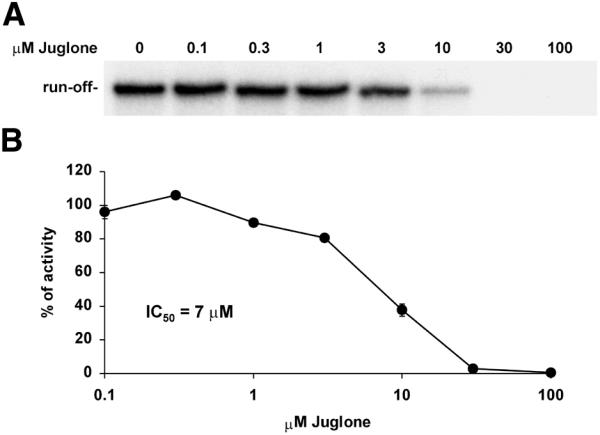
Inhibition of RNA polymerase II transcription by juglone in vitro. (A) Pulse–chase transcription assay using a linearized CMV template, HeLa nuclear extract with recombinant P-TEFb added to enhance productive elongation. Juglone was added during preincubation at the indicated concentrations. Run-off transcripts generated were analyzed in a 6% denaturing gel. (B) Quantitation of the run-off transcripts. The IC50 for the inhibitory effect of juglone on RNA polymerase II transcription was determined as described in Materials and Methods.
Pin1 has no detectable effect on RNA polymerase II transcription in vitro and in vivo
To clarify the role of Pin1 in RNA polymerase II transcription, we generated expression constructs, produced recombinant Pin1 proteins and determined their effects on transcription in vitro. Three His-tagged proteins, wild-type Pin1, a Pin1 mutant (Pin1H59A) without isomerase activity (17) and the WW domain of Pin1 (Pin1WW) were expressed in E.coli and purified (Fig. 2A). We reasoned that since Pin1H59A and Pin1WW should be able to associate with the CTD but do not contain enzymatic activity, both proteins might act as dominant-negative regulators of wild-type Pin1. An in vitro transcription assay was performed with the individual addition of purified Pin1, Pin1H59A and Pin1WW proteins. Unexpectedly, none of the proteins had any significant effect on the appearance of run-off transcripts (Fig. 2B). This lack of effect is probably not due to improper folding of the recombinant proteins because similar recombinant yeast proteins were shown to have CTD binding activity (11). Furthermore, we found the wild-type Pin1, but not Pin1H59A or Pin1WW, stimulated autophosphorylation of P-TEFb and this stimulation was inhibited by juglone (data not shown). To confirm this result in vivo, eukaryotic expression constructs of HA-tagged Pin1, Pin1H59A and Pin1WW were generated. HeLa cells were transiently cotransfected with these constructs and a CMV promoter-driven luciferase reporter gene. A western blot of transfected cell lysates probed with anti-HA antibodies indicated that Pin1 and Pin1H59A were both expressed well and in similar amounts (data not shown). However, no detectable levels of Pin1WW were observed, probably due to the instability of the Pin1WW protein (data not shown). As found in vitro, no effect on transcription of the reporter gene was found with either overexpression of Pin1 or the potentially dominant-negative Pin1H59A protein (Fig. 2C). This suggests that Pin1 may not be involved in the regulation of RNA polymerase II transcription. Since only one promoter and reporter gene construct was tested using a transient transfection assay, it is possible that Pin1 may play a role in RNA processing that is not detected by this assay. Since juglone did inhibit transcription in vitro it is likely that its effect is through another mechanism besides inhibition of Pin1.
Figure 2.
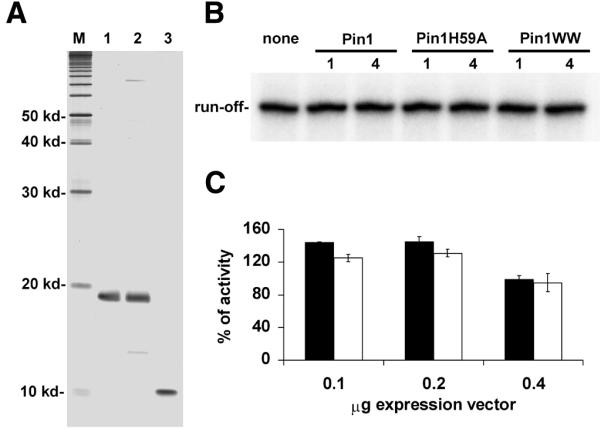
RNA polymerase II transcription is not affected by Pin1 in vitro or in vivo. (A) SDS–PAGE of the indicated three purified recombinant Pin1 proteins. M, 10 kDa ladder as a marker; lane 1, Pin1 (2 µl); lane 2, Pin1H59A (5 µl); lane 3, Pin1WW (0.4 µl). (B) Pulse–chase transcription assay with the indicated amounts (1 or 4 µl) of Pin1 proteins. Labeled transcripts were analyzed on 6% denaturing gel. (C) Co-transfection assay with a reporter (CMV-luc) and a Pin1 expression construct (pCDNA3-HA-Pin1 or pCNDA3-HA-Pin1H59A). The amounts of Pin1 constructs used are indicated.
Juglone quickly and specifically blocks a preinitiation step in RNA polymerase II transcription by attacking sulfhydryl groups
To determine what phase of transcription was inhibited by juglone, a pulse–chase assay was carried out to separate preinitiation complex formation, initiation and elongation. Juglone was added into the reactions during the formation of preinitiation complexes, initiation or elongation. Transcription was inhibited only when juglone was included in the reaction during the formation of preinitiation complexes as indicated by the disappearance of short transcripts during the subsequent pulse and the elimination of run-off transcripts during the subsequent chase (Fig. 3A, preinc.). When added after preinitiation complex formation, but before initiation, or when added at the beginning of the chase, juglone had only a small effect (Fig. 3A, pulse and chase). We conclude that juglone blocks RNA polymerase II transcription by interfering with the formation of a functional preinitiation complex and does not significantly inhibit the initiation of preformed preinitiation complexes or subsequent elongation.
Figure 3.
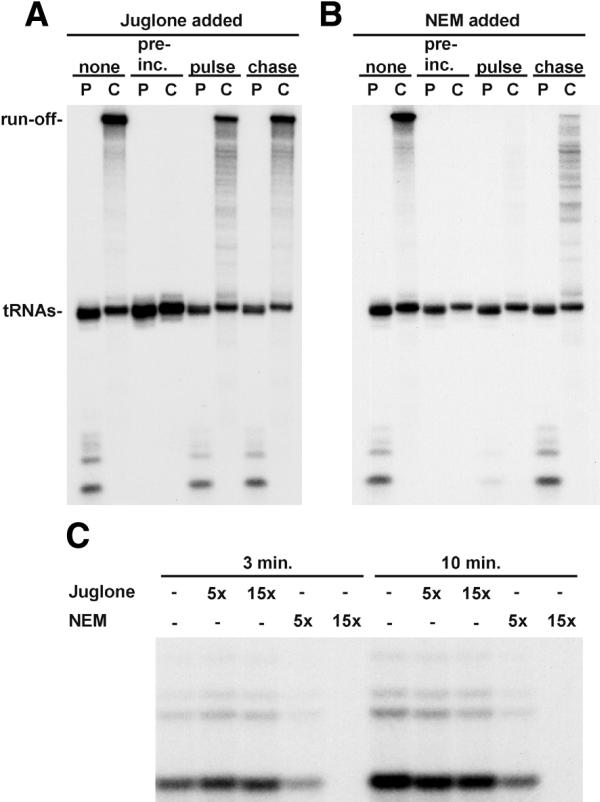
Specific disruption of the RNA polymerase II preinitiation complexes by juglone. (A) Juglone (50 µM) was added at different stages of a pulse–chase reaction as indicated: none (no drug added), preinc. (preincubation), pulse (chase). Reactions were stopped after the pulse (P) or after the chase (C). (B) Identical experiment with NEM (1 mM) instead of juglone. (C) Juglone (5× 35 µM; 15× 105 µM) and NEM (5× 580 µM; 15× 1740 µM) were added after 15–20 min preincubation and incubated for an additional 3 or 10 min. The reactions were stopped after the pulse. Labeled transcripts were analyzed on 6 (A and B) or 9% (C) denaturing gels.
NEM, a general sulfhydryl reagent, has been reported to inhibit transcription by RNA polymerase II (22). We first determined the IC50 of NEM on transcription by RNA polymerase II to be 116 µM (data not shown). Then, the identical pulse–chase experiment as described above was performed by adding 1 mM NEM (8.6 × IC50), instead of 50 µM juglone (7.1 × IC50), during different steps. As shown in Figure 3B, NEM disrupted all stages of transcription. Furthermore, the inhibitory actions of juglone and NEM during initiation were examined in detail. For comparison purposes the concentrations of these two reagents were chosen to be either five or 15 times their IC50 values. After 15–20 min of preincubation, the reactions were incubated for 3 or 10 min with the addition of juglone or NEM and then stopped after the pulse. Juglone had only a small effect on initiation indicated by the appearance of short transcripts, which were not significantly different from those of the control (Fig. 3C). However, the initiation of preinitiation complexes was blocked by NEM as indicated by the significant decrease in the appearance of short transcripts (Fig. 3C). These data suggest that juglone is a more potent and specific inhibitor of RNA polymerase II. It is possible that juglone and NEM exert their inhibitory effects on transcription by modifying different sulfhydryl groups.
The kinetics of the inhibition of the formation of preinitiation complexes by juglone was examined next. It has been shown that juglone inactivates Pin1 by forming a covalent bond through the sulfhydryl groups of cysteines in a slow and time-dependent fashion (12). A pulse–chase experiment was performed as described above, except that the reactions were stopped after the pulse to quantify initiation. The time of preincubation of extract and template was varied from 1 to 16 min in the absence or presence of 50 µM juglone. As expected, subsequent initiation was increased at longer preincubation times due to an increase in the number of preinitiation complexes that formed (Fig. 4). However, at all preincubation times juglone inhibited the production of short transcripts (Fig. 4). This indicates that, unlike inhibition of Pin1, inhibition of transcription by juglone is rapid.
Figure 4.
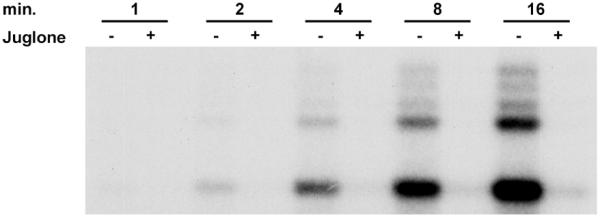
Immediate inhibition of the RNA polymerase II preinitiation complexes by juglone. Juglone (50 µM) was added at the preincubation stage of a pulse–chase reaction and reactions incubated for the indicated time. The reactions without juglone served as a control. Reactions were stopped after the pulse and labeled transcripts were analyzed on 9% denaturing gel.
To prove that juglone exerted its inhibitory effect by modifying sulfhydryl groups, another pulse-only experiment was carried out with the addition of increasing amounts of dithiothreitol (DTT). DTT at concentrations up to 10 mM had no effect on the formation of preinitiation complexes as indicated by the lack of inhibition of the production of short transcripts during the subsequent pulse (Fig. 5). However DTT blocked the inhibitory effect of juglone as indicated by the reappearance of short transcripts at higher concentrations of DTT (Fig. 5). Reversal of the inhibition of juglone by DTT strongly suggests that the mechanism of inhibition is through modification of sulfhydryl groups. The inhibitory effect of juglone was not reversed when DTT was added after a 10 min incubation with juglone (data not shown) suggesting that DTT can block but not reverse the juglone modification.
Figure 5.
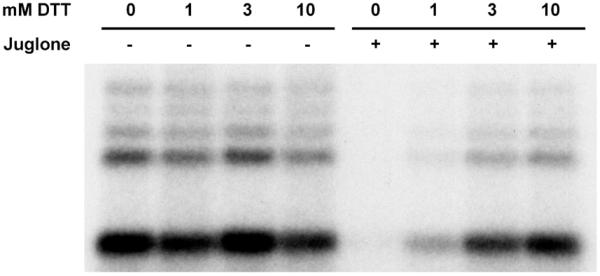
Involvement of sulfhydryl groups in the inhibitory effect of juglone. Juglone (50 µM) and the indicated amounts of DTT were added at the preincubation stage of a pulse–chase reaction. Reactions were stopped after the pulse and labeled transcripts were analyzed on 9% denaturing gel.
Transcription by RNA polymerase I and III is also inhibited by juglone
To determine if juglone was a specific inhibitor of RNA polymerase II transcription we examined the effect of juglone on RNA polymerase I and III transcription. An in vitro RNA polymerase I transcription assay with a template containing the ribosomal RNA gene promoter and HeLa nuclear extract was performed. The inclusion of increasing concentrations of juglone in the reaction resulted in the inhibition of the 681 nt run-off transcripts (Fig. 6A). The radioactivity in run-off transcripts was quantitated and the IC50 was determined to be 7 µM (Fig. 6B). A similar in vitro transcription assay was carried out to examine RNA polymerase III transcription using an Alu template that generates a 300 nt run-off transcript. Inhibition of the Alu transcripts was observed as the concentrations of juglone increased (Fig. 6C) and the IC50 was determined to be 2 µM (Fig. 6D). The results indicate that juglone inhibits RNA polymerase I, II and III transcriptions equally efficiently with similar IC50 values (Figs 1 and 6).
Figure 6.
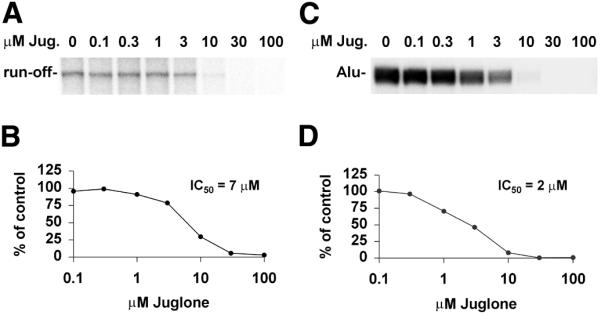
Inhibition of RNA polymerase I and III transcriptions by juglone in vitro. (A) RNA polymerase I transcription assay using linearized pHr160 as the template. (B) Quantitative data of the polymerase I transcripts. (C) RNA polymerase III transcription assay using an Alu-T template. (D) Quantitative data of Alu transcripts. The IC50 values were determined as described in Materials and Methods.
Juglone has no effect on the digestion of restriction enzymes or transcription by T7 RNA polymerase
To further investigate the specificity of juglone inhibition we determined if the compound would inhibit a variety of restriction enzymes. A 5.5 kb plasmid was digested by seven different restriction enzymes. All enzymes had only one site in the plasmid except SalI, which cuts the plasmid twice to generate the 2.2 and 3.3 kb fragments. Five of the seven restriction enzymes (BamHI, EcoRI, KpnI, XbaI and SalI) contain at least one cysteine residue. As shown in Figure 7A, 50 µM juglone had no effect on the activity of any of the restriction enzymes. We further examined the effects of juglone on transcription by T7 RNA polymerase that contains 12 cysteine residues. No inhibition was observed until the concentration of juglone reached 100 µM (∼20% inhibition) (Fig. 7B). These results indicate that juglone does not inhibit all enzymes that act on DNA including enzymes that contain cysteine.
Figure 7.
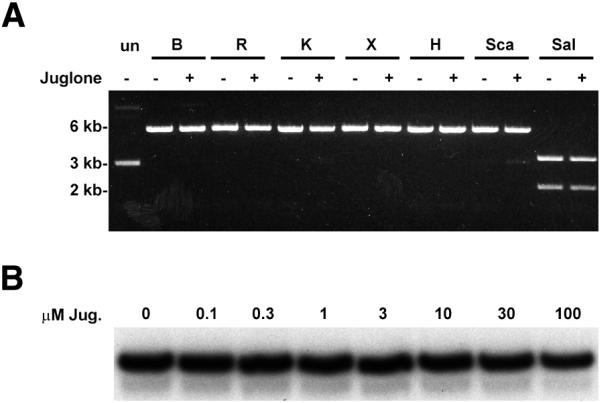
The substrate specificity of juglone. (A) The effect of juglone on restriction enzyme digestions. Plasmid pCDNA3-HA was digested by BamHI (B), EcoRI (R), KpnI (K), XbaI (X), HindIII (H), ScaI (Sca) and SalI (Sal) with (+) or without (–) the addition of juglone (50 µM). The uncut plasmid (un) served as a control. (B) The effect of juglone on the transcription of T7 RNA polymerase. Reactions were as described in Materials and Methods.
RNA polymerase II is a potential target of juglone
It has been reported that purified RNA polymerase II can be inhibited by strong sulfhydryl reagents (22,23). To find out if RNA polymerase II is the target of juglone, a standard polymerase assay (21) was performed. Purified Drosophila RNA polymerase II was preincubated with juglone for 2 min and then a single-stranded DNA template and nucleoside triphosphates were added. The mixture was incubated at 37°C for 15 min. Incorporation of label into RNA was inhibited at the higher concentrations of juglone and an IC50 of 26 µM was determined (Fig. 8). This result suggests that RNA polymerase II might be the target of juglone. However, the IC50 was higher than that determined in run-off transcription assays that utilize linearized double-stranded DNA template and HeLa nuclear extract (Fig. 1). It is not clear if the target of juglone is RNA polymerase II and the difference in IC50 values obtained just reflects differences in the assays used, or if there is an additional highly sensitive target other than RNA polymerase II seen in run-off transcription assays.
Figure 8.

RNA polymerase II as a possible target of juglone. A standard polymerase assay (21) was carried out using the purified Drosophila RNA polymerase II and ssDNA. The polymerase was preincubated with the indicated concentrations of juglone for 2 min. The IC50 was calculated as described in Materials and Methods.
DISCUSSION
We have found that juglone, a previously identified inhibitor of the peptidyl-prolyl isomerase Pin1, blocks specific initiation by RNA polymerase II. Through a Pin1-independent mechanism, juglone quickly disrupts the preinitiation process, but has little effect on initiation or elongation. Compared with NEM, juglone is a more specific and potent inhibitor of RNA polymerase II. Furthermore, promoter-initiated transcription by RNA polymerase I and III was also blocked by juglone with similar IC50 values to that of RNA polymerase II (between 2 and 7 µM). The inhibitory specificity of juglone was demonstrated by the fact that it had no significant effect on restriction enzyme digestion or promoter-initiated transcription by T7 RNA polymerase. The exact target of the compound was not determined, however the results indicated that modification of cysteine residues, potentially in the polymerase itself, were most likely involved.
It has been reported that NEM can block initiation, but not elongation, of RNA polymerase II transcription (24). Using a more precise in vitro transcription system (i.e. pulse–chase assay), we clearly demonstrated that all the processes of transcription, including preinitiation, initiation and elongation, were affected by the action of NEM. Although juglone also exerts its inhibitory effect by modifying the sulfhydryl groups, our data indicated that juglone was not just more potent, but also more specific than NEM in inhibition of RNA polymerase II. It is likely that different sulfhydryl groups are modified by juglone and NEM. Identification of the different cysteine residues modified by juglone and NEM will provide insight into understanding the regulatory mechanism of the preinitiation process of RNA polymerase II transcription.
In addition to NEM, several compounds have been discovered that block transcription by RNA polymerase II. The mushroom toxin, α-amanitin, is the most commonly used RNA polymerase II inhibitor because it blocks elongation by the polymerase and at low concentrations is specific for RNA polymerase II (25). Novobiocin, a coumarin class antibiotic, has been shown to inhibit a step in transcription by RNA polymerase I, II and III (26–29). The mechanism of inhibition by novobiocin is not known, however it has been suggested that it blocks the binding of factors to the DNA (29). Tagetitoxin, a bacterial phytotoxin, has been reported as an inhibitor of eukaryotic RNA polymerase III, bacterial RNA polymerase and chloroplast RNA polymerase, but again its mechanism of action is unclear (30–32). The inhibitory properties of heparin and Sarkosyl have been used to separate specific stages of transcription (33). However, both reagents exert their effects by blocking many protein–protein and protein–nucleic acid interactions. A significant difference between these two agents and juglone is that heparin and Sarkosyl inhibit the action of both negative and positive transcription elongation factors (34) and juglone does not. Our data demonstrate that juglone is unique in that it inhibits transcription by all three nuclear eukaryotic polymerases by modifying suflhydryl groups.
There are at least two possibilities for proteins targeted by juglone. RNA polymerase II is a strong candidate and is supported by our results showing the inhibition in a non-specific assay with pure polymerase and a single-stranded DNA template. Unfortunately, due to the nature of the non-specific assay it cannot be determined if initiation or elongation is inhibited by juglone. Because RNA polymerase I, II and III are all evolutionarily related, it is possible that one or more conserved cysteine residues are covalently modified by juglone and that this modification disrupts the activity of all three polymerases. If this is the case, modification of the site must be blocked by a conformational change that occurs during elongation. Possibly, the modification site is masked due to the binding of the polymerase to the template. In this way, modification of a specific site on the polymerase would interfere with the association of the polymerase and the template. The second possible target is the TATA box binding protein (TBP). TBP is the only known transcription factor required for the transcription by all three nuclear RNA polymerases (35). Examination of the structure of TBP bound to DNA indicates that there are several cysteines that if modified by juglone might block the binding of the factor to DNA and others that might interfere with the ability of the TBP to recruit RNA polymerase to the DNA bound TBP.
We were unable to uncover a role for Pin1 in transcription by RNA polymerase II either in vivo or in vitro. The yeast homolog of Pin1 is clearly involved in mRNA 3′-end processing and can physically associate with the phosphorylated CTD. Since the CTD plays an essential role in transcription we thought it was possible that Pin1 might affect transcription, especially the elongation control step regulated by the CTD kinase, P-TEFb (14–16,). It is not clear why we did not see any effect of Pin1 on transcription by RNA polymerase II. One possibility is that Pin1 plays a different role in mammalian cells compared to its essential role in yeast. Recently, Pin1 knockout mice have been generated and these mice exhibit no developmental abnormalities (36). Of course, the lack of a phenotype for the Pin1 knockout could be due to the presence of other family members that take over its function. Our results cannot rule out a role for Pin1 or other PPIases in RNA processing in mammalian cells. Possibly, phosphorylation of the CTD by P-TEFb allows Pin1 to associate and facilitate mRNA processing without affecting transcription. The CMV reporter construct that was unaffected by overexpression of the Pin1 knockout did contain an intron upstream of the polyadenylation site, but processing of this intron was not required for production of full-length luciferase protein. Further understanding of the function of Pin1 may be facilitated by in vitro systems that have efficiently coupled transcription and RNA processing or by a close examination of the expression of a number of genes in their native chromatin context.
Our results indicate that the unique properties of juglone could be exploited in a variety of transcriptional studies. Juglone is the only compound known that just blocks the formation of functional RNA polymerase II preinitiation complexes. Juglone would be useful in vitro when it is desirable to block re-initiation without affecting initiation of polymerases already in preinitiation complexes or polymerases that had already initiated. The unique properties of juglone complement other inhibitors such as α-amanitin that blocks both initiation and elongation of RNA polymerase II and DRB, or flavopiridol (37) that blocks only the transition into productive elongation without affecting initiation or subsequent elongation.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Jude McDonough for performing the initial RT–PCR that resulted in full-length Pin1 cDNA. This work was supported by NIH grant GM35500.
References
- 1.Gothel S.F. and Marahiel,M.A. (1999) Peptidyl-prolyl cis-trans isomerases, a superfamily of ubiquitous folding catalysts. Cell. Mol. Life Sci., 55, 423–436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lu K.P. (2000) Phosphorylation-dependent prolyl isomerization: a novel cell cycle regulatory mechanism. Prog. Cell Cycle Res., 4, 83–96. [DOI] [PubMed] [Google Scholar]
- 3.Hani J., Schelbert,B., Bernhardt,A., Domdey,H., Fischer,G., Wiebauer,K. and Rahfeld,J.U. (1999) Mutations in a peptidylprolyl-cis/trans-isomerase gene lead to a defect in 3′-end formation of a pre-mRNA in Saccharomyces cerevisiae. J. Biol. Chem., 274, 108–116. [DOI] [PubMed] [Google Scholar]
- 4.Rahfeld J.U., Rucknagel,K.P., Schelbert,B., Ludwig,B., Hacker,J., Mann,K. and Fischer,G. (1994) Confirmation of the existence of a third family among peptidyl-prolyl cis/trans isomerases. Amino acid sequence and recombinant production of parvulin. FEBS Lett., 352, 180–184. [DOI] [PubMed] [Google Scholar]
- 5.Maleszka R., Lupas,A., Hanes,S.D. and Miklos,G.L. (1997) The dodo gene family encodes a novel protein involved in signal transduction and protein folding. Gene, 203, 89–93. [DOI] [PubMed] [Google Scholar]
- 6.Landrieu I., De Veylder,L., Fruchart,J.S., Odaert,B., Casteels,P., Portetelle,D., Van Montagu,M., Inze,D. and Lippens,G. (2000) The Arabidopsis thaliana PIN1At gene encodes a single-domain phosphorylation-dependent peptidyl prolyl cis/trans isomerase. J. Biol. Chem., 275, 10577–10581. [DOI] [PubMed] [Google Scholar]
- 7.Lu K.P., Hanes,S.D. and Hunter,T. (1996) A human peptidyl-prolyl isomerase essential for regulation of mitosis. Nature, 380, 544–547. [DOI] [PubMed] [Google Scholar]
- 8.Lu P.J., Zhou,X.Z., Shen,M. and Lu,K.P. (1999) Function of WW domains as phosphoserine- or phosphothreonine-binding modules. Science, 283, 1325–1328. [DOI] [PubMed] [Google Scholar]
- 9.Dahmus M.E. (1996) Phosphorylation of mammalian RNA polymerase II. Methods Enzymol., 273, 185–193. [DOI] [PubMed] [Google Scholar]
- 10.Bentley D. (1999) Coupling RNA polymerase II transcription with pre-mRNA processing. Curr. Opin. Cell Biol., 11, 347–351. [DOI] [PubMed] [Google Scholar]
- 11.Morris D.P., Phatnani,H.P. and Greenleaf,A.L. (1999) Phospho-carboxyl-terminal domain binding and the role of a prolyl isomerase in pre-mRNA 3′-end formation. J. Biol. Chem., 274, 31583–31587. [DOI] [PubMed] [Google Scholar]
- 12.Hennig L., Christner,C., Kipping,M., Schelbert,B., Rucknagel,K.P., Grabley,S., Kullertz,G. and Fischer,G. (1998) Selective inactivation of parvulin-like peptidyl-prolyl cis/trans isomerases by juglone. Biochemistry, 37, 5953–5960. [DOI] [PubMed] [Google Scholar]
- 13.Webb J.L. (1966) Enzyme and Metabolic Inhibitors. Academic Press, New York.
- 14.Marshall N.F., Peng,J., Xie,Z. and Price,D.H. (1996) Control of RNA polymerase II elongation potential by a novel carboxyl-terminal domain kinase. J. Biol. Chem., 271, 27176–27183. [DOI] [PubMed] [Google Scholar]
- 15.Yamaguchi Y., Wada,T. and Handa,H. (1998) Interplay between positive and negative elongation factors: drawing a new view of DRB. Genes Cells, 3, 9–15. [DOI] [PubMed] [Google Scholar]
- 16.Price D.H. (2000) P-TEFb, a cyclin-dependent kinase controlling elongation by RNA polymerase II. Mol. Cell. Biol., 20, 2629–2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shen M., Stukenberg,P.T., Kirschner,M.W. and Lu,K.P. (1998) The essential mitotic peptidyl-prolyl isomerase Pin1 binds and regulates mitosis-specific phosphoproteins. Genes Dev., 12, 706–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marshall N.F. and Price,D.H. (1992) Control of formation of two distinct classes of RNA polymerase II elongation complexes. Mol. Cell. Biol., 12, 2078–2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hara R., Selby,C.P., Liu,M., Price,D.H. and Sancar,A. (1999) Human transcription release factor 2 dissociates RNA polymerases I and II stalled at a cyclobutane thymine dimer. J. Biol. Chem., 274, 24779–24786. [DOI] [PubMed] [Google Scholar]
- 20.Chu W.M., Wang,Z., Roeder,R.G. and Schmid,C.W. (1997) RNA polymerase III transcription repressed by Rb through its interactions with TFIIIB and TFIIIC2. J. Biol. Chem., 272, 14755–14761. [DOI] [PubMed] [Google Scholar]
- 21.Price D.H., Sluder,A.E. and Greenleaf,A.L. (1987) Fractionation of transcription factors for RNA polymerase II from Drosophila Kc cell nuclear extracts. J. Biol. Chem., 262, 3244–3255. [PubMed] [Google Scholar]
- 22.Gissinger F., Kedinger,C. and Chambon,P. (1974) Animal DNA-dependent RNA polymerases. 10. General enzymatic properties of purified calf thymus RNA polymerases AI and B. Biochimie, 56, 319–333. [DOI] [PubMed] [Google Scholar]
- 23.Chambon P. (1974) In Boyer,P. (ed.), The Enzymes, Vol. X. Academic Press, San Diego, CA, pp. 261–331. [Google Scholar]
- 24.Krueger R.C. (1990) RNA elongation by RNA polymerase II is not inhibited by N-ethylmaleimide or iodoacetamide. Arch. Biochem. Biophys ., 278, 475–477. [DOI] [PubMed] [Google Scholar]
- 25.Cochet-Meilhac M. and Chambon,P. (1974) Animal DNA-dependent RNA polymerases. 11. Mechanism of the inhibition of RNA polymerases B by amatoxins. Biochim. Biophys. Acta, 353, 160–184. [DOI] [PubMed] [Google Scholar]
- 26.Webb M.L., Maguire,K.A. and Jacob,S.T. (1987) Novobiocin inhibits initiation of RNA polymerase II-directed transcription of the mouse metallothionein-I gene independent of its effect on DNA topoisomerase II. Nucleic Acids Res., 15, 8547–8560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Felts S.J., Weil,P.A. and Chalkley,R. (1987) Novobiocin inhibits interactions required for yeast TFIIIB sequestration during stable transcription complex formation in vitro. Nucleic Acids Res., 15, 1493–1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Webb M.L. and Jacob,S.T. (1988) Inhibition of RNA polymerase I-directed transcription by novobiocin. Potential use of novobiocin as a general inhibitor of eukaryotic transcription initiation. J. Biol. Chem., 263, 4745–4748. [PubMed] [Google Scholar]
- 29.Van Dyke M.W. and Roeder,R.G. (1987) Novobiocin interferes with the binding of transcription factors TFIIIA and TFIIIC to the promoters of class III genes. Nucleic Acids Res., 15, 4365–4374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Steinberg T.H., Mathews,D.E., Durbin,R.D. and Burgess,R.R. (1990) Tagetitoxin: a new inhibitor of eukaryotic transcription by RNA polymerase III. J. Biol. Chem., 265, 499–505. [PubMed] [Google Scholar]
- 31.Mathews D.E. and Durbin,R.D. (1990) Tagetitoxin inhibits RNA synthesis directed by RNA polymerases from chloroplasts and Escherichia coli. J. Biol. Chem., 265, 493–498. [PubMed] [Google Scholar]
- 32.Mathews D.E. and Durbin,R.D. (1994) Mechanistic aspects of tagetitoxin inhibition of RNA polymerase from Escherichia coli. Biochemistry, 33, 11987–11992. [DOI] [PubMed] [Google Scholar]
- 33.Hawley D.K. and Roeder,R.G. (1985) Separation and partial characterization of three functional steps in transcription initiation by human RNA polymerase II. J. Biol. Chem., 260, 8163–8172. [PubMed] [Google Scholar]
- 34.Kephart D.D., Marshall,N.F. and Price,D.H. (1992) Stability of Drosophila RNA polymerase II elongation complexes in vitro. Mol. Cell. Biol., 12, 2067–2077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lee T.I. and Young,R.A. (1998) Regulation of gene expression by TBP-associated proteins. Genes Dev., 12, 1398–1408. [DOI] [PubMed] [Google Scholar]
- 36.Fujimori F., Takahashi,K., Uchida,C. and Uchida,T. (1999) Mice lacking Pin1 develop normally, but are defective in entering cell cycle from G(0) arrest. Biochem. Biophys. Res. Commun., 265, 658–663. [DOI] [PubMed] [Google Scholar]
- 37.Chao S.H., Fujinaga,K., Marion,J.E., Taube,R., Sausville,E.A., Senderowicz,A.M., Peterlin,B.M. and Price,D.H. (2000) Flavopiridol inhibits P-TEFb and blocks HIV-1 replication. J. Biol. Chem., 275, 28345–28348. [DOI] [PubMed] [Google Scholar]