

Abstract
Purpose
Physically active women have a reduced risk of breast cancer, but the dose of activity necessary and the role of energy balance and other potential mechanisms have not been fully explored in animal models. We examined treadmill and wheel running effects on mammary tumorigenesis and biomarkers in p53-deficient (p53+/−): MMTV-Wnt-1 transgenic mice.
Methods
Female mice (9 wks old) were randomly assigned to the following groups in Experiment 1: treadmill exercise 5 d/wk, 45 min/d, 5% grade at 20 m/min, ~0.90 km/d (TREX1, n=20); at 24 m/min, ~1.08 km/d (TREX2, n=21); or a non-exercise control (CON-TREX, n=22). In Experiment 2, mice were randomly assigned to voluntary wheel-running (WHL, n=21, 2.46 ± 1.11 km/d (mean ± SD)) or a non-exercise control (CON-WHL, n=22). Body composition was measured at ~9 weeks and serum insulin-like growth factor-1 (IGF-1) at 2–3 monthly time points beginning at ~9 weeks on study. Mice were sacrificed when tumors reached 1.5 cm, mice became moribund, or there was only one mouse per treatment group remaining.
Results
TREX1 (24 wks) and TREX2 (21 wks) had shorter survival median survival times than CON-TREX (34 wks; p<0.01); WHL and CON-WHL survival was similar (23 vs. 24 wks; p=0.32). TREX2 had increased multiplicity of mammary gland carcinomas compared to CON-TREX; WHL had a higher tumor incidence than CON-WHL. All exercising animals were lighter than their respective controls, and WHL had lower body fat than CON-WHL (p<0.01). There was no difference in IGF-1 between groups (p>0.05).
Conclusion
Despite beneficial or no effects on body weight, body fat, or IGF-1, exercise had detrimental effects on tumorigenesis in this p53-deficient mouse model of spontaneous mammary cancer.
Keywords: physical activity, mammary, IGF-1, body composition
Epidemiological evidence suggests that physically active women have a 20–30% reduced risk of breast cancer compared to physically inactive women (20). Despite the reasonable consistency of the evidence, details as to the types and amount of activity necessary to reduce risk are lacking since not all studies have assessed physical activity in detail. Although numerous mechanisms have been proposed to explain the risk reduction, there is little evidence to substantiate any of them (35). Furthermore, key exercise-gene interactions that may impact the breast cancer preventive effects of exercise have not been described. Animal models that reflect important aspects of breast tumorigenesis in humans would be useful to help fill these gaps in our understanding of the exercise-breast cancer relationship.
To date, the primary animal model used to examine exercise in breast cancer has been chemically induced mammary carcinogenesis in rats or mice. Earlier studies were remarkably consistent in finding beneficial effects of exercise on mammary tumor development, as reviewed by Shephard (29); however, these studies often used particularly long or strenuous exercise protocols. Recent studies have used more moderate treadmill running protocols and generally found protection against either tumor incidence, multiplicity, or growth (19;32;33;37;38) however, some have found negative effects of exercise on the carcinogenic process (30;31). It has been shown that there may be an intensity-dependent effect of treadmill running, with low-intensity running having no effect on carcinogenesis, but with intensities beyond an oxygen consumption of ~35% of maximum capacity resulting in decreased incidence and multiplicity in a dose-response fashion (32;33). Voluntary wheel running has been shown to decrease tumor multiplicity and increase tumor latency in rat models, although there are relatively few studies of the effects of this type of exercise(5;7). In one prior study which examined both treadmill and wheel running exercise, neither the incidence or multiplicity of lung tumor metastases following injection of the MMT 66 mouse mammary tumor cell line was effected by the exercise (16).
Taken together, exercise-training studies have shown variability in the relationship between changes in body weight and/or body fat and changes in carcinogenesis. For example, in studies in which exercise reduced tumor outcome/development, body weights have been higher (5;6), lower (19), or no different (7;32;37) from the non-exercise control groups. Similarly, body fat has been either unchanged (7) or lower (5;6) with exercise regimens; conversely, in one study in which exercise resulted in lower body fat, carcinogenesis was not inhibited (15). These disparate results suggest that the mechanism through which exercise impacts tumorigenesis may be independent of energy balance, although the link between exercise/physical activity in energy balance and breast cancer reduction is still unclear (34). Animal models in which body weight and activity can be objectively and accurately monitored in the context of carcinogenesis will provide more insight into these potential relationships.
Newer models of carcinogenesis use mice with known mutations in specific genes. One such model of breast cancer is the MMTV-Wnt-1 transgenic mouse model of breast cancer (12;21). Female MMTV-Wnt-1 mice spontaneously develop mammary tumors with 100% penetrance, and the latency for tumor development is greatly reduced by p53 deficiency. We previously found that calorie restriction (CR) dramatically delayed tumorigenesis in heterozygous p53-deficient (p53+/−):MMTV-Wnt-1 transgenic mice (18). We then sought to determine how altering energy expenditure, the other side of the energy balance equation, would affect mammary tumorigenesis in this model.
Other than changes in body weight or composition, few studies have examined potential mechanisms of the exercise-induced protection against mammary tumors. In those that have examined other markers, neither estrogen or progesterone, nor differential expression of estrogen or IGF-1 receptors in the tumors have been related to the exercise treatment (7;36;38;39). In the current study, we have examined the effects of voluntary wheel running and involuntary treadmill exercise on mammary tumorigenesis, insulin-like growth factor-1 and body composition in a mouse model of cancer.
METHODS
Animals
All experimentation was approved by the National Cancer Institute’s Animal Care and Use Committee. NCI-Frederick is accredited by AAALAC International and follows the Public Health Service Policy for the Care and Use of Laboratory Animals. Animal care was provided in accordance with the procedures outlined in the Guide for Care and Use of Laboratory Animals. (National Research Council; 1996; National Academy Press; Washington, D.C.) Female heterozygous p53-deficient (p53+/−): MMTV-Wnt-1 transgenic mice were obtained from a breeding colony at University of Texas MD Anderson Cancer Center, Smithville, TX. Mice were received in five separate shipments of 16–26 mice and were 9.3 ± 1.7 wks of age (mean ± SD). Within each shipment mice were randomized to treatment groups. All mice were housed individually within 7 days of arrival, given water and AIN-76A diet ad libitum, and kept on a 12-h light/dark cycle. All mice spent the first ~8 wks in quarantine, after which they were confirmed to be pathogen free and transferred to the main animal facility.
Experimental Design
There were two primary experiments: one examining the effects of non-voluntary treadmill running and the other voluntary wheel running. Both examined survival and specific biomarkers as endpoints. In Experiment 1, mice were randomized to one of three groups: a non-exercised control group (CON-TREX, n=22), a moderate (TREX1, n=20) intensity, or a vigorous (TREX2, n=21) intensity treadmill running group. Treadmill running was done 5 days/wk, 45 min/day on a 5% grade, with TREX1 running at 20m/min and TREX2 at 24 m/min. Mice were motivated to run using gentle hand prodding. We have previously found exercise of this intensity to increase citrate synthase in the gastrocnemius/soleus complex in mice.(8) CON-TREX mice were housed in the same room as the treadmill and were placed briefly on the stationary treadmill each day that the TREX1 and TREX2 mice ran. In Experiment 2, mice were randomized into either a non-wheel control group (CON-WHL, n=21) or a voluntary wheel-running condition (WHL, n=20) in which they had 24-hr access to a running wheel in their cage. Mice randomized to any of the exercise groups (TREX1, TREX2, WHL) were given access to running wheels while in quarantine. Wheel running activity was monitored during the study using magnetic switches in conjunction with VitalView software (MiniMitter, Bend, OR). Time on study was considered to begin when the mice were randomized.
Mouse body weights and food consumption were measured weekly. Mammary glands were palpated twice per week for the detection of tumors, and dates, locations, and sizes of tumors were recorded. Volume of the tumors (mm3) was calculated as 1/6*π*length*width*height and is calculated for the initial detection of the tumor and twice weekly measurements until sacrifice. Retroorbital eye bleeds were drawn once per month beginning at ~9 wks on study, and the serum was stored at −80°C.
When a palpable mammary tumor reached 1.5 cm in diameter, or when mice became moribund, they were sacrificed. Additionally, when only one mouse per group was left in a given treatment group in an experiment, they were sacrificed. Complete necropsies were performed and gross lesions were recorded. Mammary gland (MG) masses were measured, and diagramed using mouse MG designation 1 though 10. MG 7&8, or other non-tumor bearing glands were whole mounted. A portion of the largest mammary tumor in each animal was collected in OCT for cryotomy, and a portion was snap frozen. Remaining mammary glands were individually identified and fixed in 10% neutral buffered formalin. Gastrocnemius muscle, heart, and a terminal serum sample were collected, frozen, and stored. Mammary glands, lungs, and gross lesions designated by the pathologist, were taken to hematoxylin and eosin stained slides. Sections were evaluated by a board-certified veterinary pathologist to determine the histopathological classification of mammary tumors and other lesions (author DCH).
Body composition
Fat mass, lean mass, and bone mineral density (BMD) values were determined on all mice, (with the exception of mice in the last shipment due to an oversight), using dual-energy x-ray absorptiometry (DXA) (GE Lunar Piximus II, Madison, WI). Each mouse was lightly anesthetized with isoflurane and scanned three successive times, and the mean of those three scans was used for analysis. Animals in the treadmill experiment were scanned at ~10 wks on study and those in the wheel running experiment were scanned at ~8 wks on study. We have previously validated the use of DXA on mice through comparison to gravimetric and chemical (soxhlet) extraction methods (1).
IGF-1
Serum IGF-1 was measured using radioimmunoassay in a sample of mice at 2 or 3 monthly time points, beginning with the first blood draw taken outside quarantine (Diagnostic Systems Laboratories, Inc., Webster, TX). These early timepoints were selected with the thought that potential IGF-1 changes would occur earlier on in the training program as fitness was developing. Only mice that had no evidence of tumors at the time of the blood draw were assayed at each time point to isolate the effects of exercise from any effects of tumors themselves. Mice from each group within an experiment were assayed at the same time. Sample size varied at each time from a minimum of 4 to a maximum of 11. The average coefficient of variation (cv) for all samples (performed in duplicate) was 7.2%.
p53
Tumor loss of heterozygosity (LOH) for p53 was measured by Southern blotting as described previously (13). Briefly, 5–10 ug of tumor DNA was digested with BamHI restriction endonuclease, subjected to electrophoresis on a 0.7% agarose gel and then Southern blotted. The blots were hybridized with a mouse p53 cDNA exon 2–6 probe radiolabelled with alpha 32P-dCTP. Three bands were generally observed: a 5.0 kb wild-type band, a 6.5 kb mutant band, and a 10.0 kb pseudogene band. Radioactivity was quantitated on a GE Storm 860 phosphorimager. LOH was characterized by comparison of wild-type to mutant bands, with bands that were 70–100+%, 30–70%, or 0–30% the intensity of the mutant alleles considered “no LOH,” “partial LOH,” and “LOH,” respectively. Tumors from 10–19 mice/group were analyzed.
Wnt-1
Mammary gland Wnt-1 transgene expression was measured in non-tumor bearing mammary glands in 8–10 mice/group, harvested at sacrifice. Briefly, RNA was extracted from frozen glands using Stratagene’s Absolutely RNA® Miniprep kit, according to the manufacturer’s protocol. RNA (2.5 μg of total RNA) was reverse transcribed with MMLV reverse transcriptase and random primers (Promega). Real-time quantitative PCR was performed using the ABI Prizm® 7900HT Sequence Detection System (Applied Biosystems). The pre-designed TaqMan® gene expression assay for mouse Wnt-1 was purchased from Applied Biosystems. TATA box binding protein (TBP) was used as a reference gene. TBP primers and probe were designed with Primer Express® software (Applied Biosystems) according to manufacturer’s instructions, and cross-evaluated with the Oligo 4.0® Primer Design software. The TBP primers and probe used were: forward primer (5′-GGTGGCAGCATGAAGTGACA-3′), reverse primer (5′-GCACAGAGCAAGCAACTCACA-3′) and TaqMan® probe (5′-FAM-CCTCTGCACTGAAATCACCTGCAGCA-TAMRA-3′). A calibrator, i.e. relative standard curve, was generated from cDNA from an untreated Wnt-1 transgenic mouse mammary gland. Assays were run in duplicate and repeated on two different days, and the values averaged. The average threshold cycle (Ct) of each sample was plotted against the log of the nanograms cDNA added to the wells of each standard curve point, and the reverse log of that number was normalized to its corresponding TBP value. The result is a unit-less number that reflects the amount of target gene in different samples relative to the calibrator.
Statistical Analyses
Mice that did not run for more than 50% of their time on the treadmill portion of the study were censored on the day they stopped running for both the survival and body weight analysis, and, additionally, were not used in the tumor growth calculation in order to examine the effects of the exercise specifically. A total of 10 out of the 41 mice (24%) run on the treadmill were censored for poor running. Additionally, only mice with mammary gland carcinomas were used in the tumor growth analysis.
All statistical tests were performed using SAS (Cary, NC). Log-rank tests were used for the nonparametric comparison of survival across groups. Differences in tumor growth and Wnt expression were analyzed using log-transformed data due to non-normal distributions. ANOVA with Tukey’s post-hoc test was used for the three group comparisons in the treadmill experiment and unpaired t-tests for the wheel experiment. Geometric means are presented in the tables when log-transformed data were used. Fisher’s Exact Test was used to compare the frequency of multiple tumors, metastatic tumors, and LOH of p53 in tumors across treatment groups. Pearson correlations were calculated for the survival time and Wnt-1 expression in tumors (log-transformed data).
For the analysis of body weight, food consumption, and IGF-1 levels over time, a two-factor ANOVA with repeated measures on time was conducted using Proc Mixed. When interaction terms were significant, contrast statements were used to determine specific differences of interest. The body weight analysis was truncated at 30 weeks on study as this is the point at which there were still at least 4 mice/group. ANOVA was used to examine differences in body composition and body weights at sacrifice by treatment group in the treadmill experiment, and unpaired t-tests were used in the wheel experiment. For BMD, heart, and gastrocnemius weights, ANCOVA was used to adjust for body weight.
RESULTS
During their quarantine period, when all exercising mice had access to running wheels, mice in the TREX1 group ran 1.60 km/d (1.24 – 3.32 km/d ) (median, interquartile range) on the running wheels, while TREX2 mice ran 2.59 km/d (2.07 – 3.26 km/d). Mice in the WHL group had their running monitored during their entire stay in quarantine, and during most of the rest of their time on study (on average, 74% of their days on study). The WHL group ran 2.82 km/d (2.20 – 3.05 km/d).
There was no difference in tumor p53 LOH across treatment groups in either experiment. Of the 53 tumors tested, 60% had complete LOH, 8% partial LOH, and 32% no LOH. A representative Southern blot is shown in Figure 1. There was no significant difference in mammary Wnt-1 mRNA expression by exercise treatment group in normal mammary glands taken at sacrifice (p>0.05). Additionally, Wnt-1 expression was not correlated with survival time (r= − 0.17, p=0.27).
Figure 1.
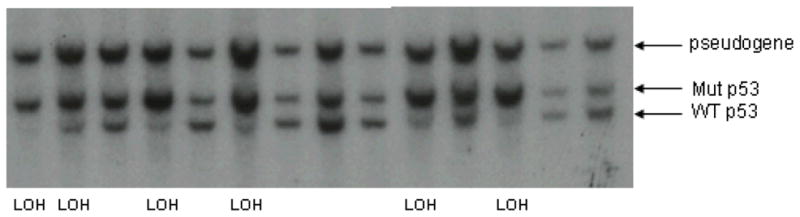
Southern blot hybridization analysis of p53+/− Wnt-1 TG mammary tumor DNA demonstrates loss of the wildtype p53 allele in a subset of tumors. Genomic DNA was prepared from p53+/− Wnt-1 TG mammary tumors and restricted with Bam HI prior to agarose gel electrophoresis and Southern blot hybridization with a labeled mouse p53 cDNA probe (from p53 exons 2–6)(13). Fourteen representative tumor DNAs are shown. Six of these 14 tumors show definitive loss of the wildtype p53 allele (LOH) (fastest migrating band). Residual presence of a wild type (WT) allele in these LOH tumors is ascribed to stromal and normal cell contamination of the tumor samples.
The mammary neoplasm morphology was similar across the groups: glandular carcinomas. Incidence and multiplicity (more than one gland with carcinoma), however, tended to be higher in the exercise groups (Table 1). The increase in multiplicity was statistically significant in the TREX2 group, and tended to be higher in TREX1 and WHL than in their respective controls. Incidence of mammary carcinoma metastasis to the lung was similar across the treadmill groups, but was elevated in WHL compared to CON-WHL. When the poor runners from the TREX1 and TREX2 groups were excluded, the results were unchanged. We also measured the time from the initial detection of a tumor until it reached 1.5 cm as a proxy measure of tumor growth. As seen in Table 1, tumor growth was faster in TREX1 vs. CON-TREX (p=0.01) with no differences between TREX2 and CON-TREX. Tumor growth tended to be slower in WHL vs. CON-WHL but was not significantly different (p=0.10). Figure 2 depicts the growth of tumors in the mice uncensored from the survival analysis. In cases where mice had more than one tumor, only the tumor that caused their sacrifice is plotted.
Table 1.
Tumor incidence, multiplicity, metastasis, and growth by experiment and treatment group.
| Experiment 1 |
p | Experiment 2 |
|||||
|---|---|---|---|---|---|---|---|
| CON-TREX (n=22) | TREX1 (n=20) | TREX2 (n=21) | CONWHL (n=21) | WHL (n=20) | p | ||
| No. mice with mammary carcinoma*,† | 19/22 (86%) | 19/20 (95%) | 21/21 (100%) | 0.26 | 16/21 (76%) | 20/20 (100%) | 0.05 |
| No. mice with mammary carcinoma in >1 gland | 5/19 (26%) | 11/19 (58%) | 14/21 (67%) ‡ | 0.03 | 8/16 (50%) | 16/20 (80%) | 0.08 |
| No. mice with mammary carcinoma metastasis in lung | 6/19 (27%) | 4/19 (21%) | 5/21 (24%) | 0.81 | 1/16 (6%) | 6/20 (30%) | 0.10 |
| TUMOR GROWTH § | CON-TREX (n=19) | TREX1 (n=13) | TREX2 (n=17) | p | CONWHL (n=16) | WHL (n=20) | p |
| Time to 1.5 cm tumor (days; mean ± SE) | 24.8 ± 1.1 | 13.8 ± 1.2‡ | 19.5 ± 1.1 | 0.02 | 17.2 ± 1.1 | 22.5 ± 1.1 | 0.10 |
Other causes of sacrifice/death included: CON-TREX-adrenal pheochromocytoma, lipoma, end of study; TREX1-end of study; CON-WHL-T-cell lymphoma, sarcoma (n=3), end of study.
Additional, non-mammary gland neoplasms: CON-TREX-sarcoma, clitoral gland carcinoma, rhabdomyosarcoma; TREX1-lymphoma.
p<0.05 vs. CON-TREX
Includes only mice not censored in the survival analysis. Log transformed data used in the statistical analysis due to non-normal distribution; geometric means presented in table.
Figure 2.

Tumor growth by treatment group. Individual tumors from mice from each treatment group are plotted. Volume of the tumors was calculated as 1/6*π*length*width*height and was calculated for the initial detection of the tumor and twice weekly measurements until sacrifice.
Mice that did not develop mammary epithelial tumors were censored for the survival analysis (specific reasons indicated in Table 1). Survival times were significantly different in Experiment 1 (p<0.001) (Figure 3). The median survival times for TREX1 (24 wks; p<0.01) and TREX2 (21 wks; p<0.001) were significantly different from that of CON-TREX (34 wks). Survival time was not significantly different in Experiment 2, in which median survival of WHL was 21.5 wks vs. 24.0 wks in CON-WHL (p=0.39).
Figure 3.

Survival by treatment group. Mice were censored for non-mammary tumor endpoints including discontinuation of running (for TREX1 or TREX2) and death due to other causes. a) CON-TREX vs. TREX1, p<0.01; CON-TREX vs. TREX2. p<0.001. b) CON-WHL vs. WHL, p=0.39).
Body weights over the course of the first 30 wks of study are presented in Figure 4. In both experiments, there were significant time by group interactions. TREX2 resulted in lower body weights vs. CON-TREX during weeks 1–20 (p<0.05), while TREX1 had lower body weights vs. CON-TREX from wks 11–20 (p=0.03). WHL body weight was significantly lower than CON-WHL from wks 11–30 (p<0.001). Due to attrition in survival over the course of the study, variability in the measurements increased with time.
Figure 4.

Body weight (mean ± SE) by time and group in each experiment. a) There was a significant time-by-group interaction (p<0.01) in the treadmill experiment. Body weight was significantly lower in TREX2 vs. CON-TREX during wks 1–10 (p=0.02) but not in TREX1 vs. CON-TREX during the same time (p=0.08). During wks 11–20, both TREX1 and TREX2 were significantly lower than CON-TREX (p<0.05): in wks 21–30, when survival in the TREX1 and TREX2 groups had fallen to less than 50%, there were no significant differences between groups (p>0.05). b) There was a significant time-by-group interaction (p<0.001) in the wheel experiment. WHL was significantly lower than CON-WHL during weeks 11–30 (p<0.001), but not during wks 1–10 (p=0.20).
There was a significant time-by-group interaction in food consumption (p<0.01) for the treadmill experiment, but the main group effect was not significant (p=0.88) (data not shown). Follow-up contrasts showed no significant difference between the CON-TREX group and either TREX1 or TREX2 during weeks 1–10, 11–20, or 21–30 (p>0.05). There was a significant time-by-group interaction in food consumption for the wheel (p<0.001) experiments. In Experiment 2, WHL mice had lower food intakes than CON-WHL during weeks 11–30 (p=0.001) but not weeks 1–10 (p=0.91).
In neither of the experiments were there any significant time-by-group interactions, nor were there significant main effects of either group or time, on IGF-1 levels (data not shown). Table 2 presents data on body composition at week ~8–10 on study, as well as body, heart, and gastrocnemius weights at sacrifice. After ~8–10 wks on study, TREX2 weighed less than CON-TREX (p<0.01), and WHL weighed less than CON-WHL (p<0.05). Although percent body fat tended to be lower in exercised mice, the only significant difference was a lower percent body fat in WHL vs. CON-WHL. Mice in all exercising groups had higher BMDs than did their respective non-exercised controls. WHL mice had significantly greater lean mass than CON-WHL at this time point. Among mice not censored from the survival study, body weights at sacrifice were significantly lower in all exercising mice compared to their respective controls. Additionally, WHL mice had greater heart and gastrocnemius weights than CON-WHL, after adjustment for body weight.
Table 2.
Body composition at ~8–10 weeks on study and body and muscle weights at sacrifice by experiment and treatment group.*
| Group | DXA data at ~8–10 wks on study | Weights at sacrifice | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | N | Body Weight (g) | Fat (%) | BMD (mg/cm2) † | Lean mass(g) | N | Body weight (g) | Heart (g) † | Gastrocnemius (g) † |
| CON-TREX | 22 | 27.4 ± 0.9 | 32.4 ± 2.0 | 48.5 ± 0.7 | 18.4 ± 0.7 | 19 | 36.5 ± 2.0 | 0.15 ± 0.01 | 0.10 ± 0.01 |
| TREX1 | 20 | 24.6 ± 0.9 | 30.3 ± 2.1 | 51.6 ± 0.7§ | 17.0 ± 0.7 | 13 | 28.8 ± 2.2‡ | 0.16 ± 0.01 | 0.11 ± 0.01 |
| TREX2 | 21 | 23.2 ± 0.9§ | 26.7 ± 2.1 | 52.0 ± 0.7§ | 16.8 ± 0.7 | 16 | 30.1 ± 1.5‡ | 0.17 ± 0.01 | 0.10 ± 0.01 |
| Experiment 2 | |||||||||
| CON-WHL | 18 | 24.9 ± 0.9 | 40.9 ± 1.9 | 48.9 ± 0.8 | 14.5 ± 0.2 | 16 | 35.3 ± 2.4 | 0.14 ± 0.01 | 0.08 ± 0.01 |
| WHL | 18 | 22.2 ± 0.6‡ | 29.3 ± 1.7§ | 51.5 ± 0.8‡ | 15.5 ± 0.3§ | 20 | 24.7 ± 0.8§ | 0.19 ± 0.01§ | 0.12 ± 0.01§ |
Only mice not censored from the survival analysis are included in the measurements of weight at sacrifice.
adjusted for total body mass
p<0.05 vs. respective control group (using Tukey-Kramer adjustment for multiple comparisons in Treadmill experiment)
p<0.01 vs. respective control group (using Tukey -Kramer adjustment for multiple comparisons in Treadmill experiment)
DISCUSSION
In p53 +/−:MMTV-Wnt-1 transgenic mice, we found that exercise produced an increase in spontaneous mammary tumorigensis, including decreased latency and shortened survival time. Previously, our group had seen beneficial effects of calorie restriction (CR) on tumorigenesis in this same mouse model (27). Mice on moderate levels of CR, as we had used previously, still grow and gain weight, but at a much slower rate compared to mice fed ad libitum (17). We hypothesized that the same beneficial effects on tumorigenesis might be attained by increasing energy expenditure rather than decreasing energy intake. With the ad libitum feeding regimen used here, all mice had a positive energy balance throughout the study based on the body weight trajectories. While treadmill exercise produced only a modestly lower body weight, the voluntary wheel running condition, consistent with the longer average distance run per day, resulted in a statistically significant lower body weight over the course of the study, as well as a reduction in body fat compared to the control mice. Despite this beneficial change in body composition, survival was not improved by the physical activity and a higher proportion of exercised mice had multiple mammary carcinomas. These data suggest that increasing energy expenditure does not have the same effect as decreasing energy intake on mammary tumor development, at least in p53 +/−:MMTV-Wnt-1 transgenic mice.
In contrast, we have previously observed beneficial exercise effects in the ApcMin model of intestinal polyp development, in which a negative energy balance produced via wheel running lowered polyp number, though still not of the same magnitude as with CR, while a milder degree of negative energy balance produced through treadmill exercise had no effect on polyps (9;10;23). This idea of the necessity for a negative energy balance with exercise has also recently been supported by a study comparing skin gene expression in mice after CR, ad libitum-fed exercise, or pair-fed exercise (22). Gene expression changes in pair-fed exercised mice were more similar to CR mice, while changes in gene expression in ad libitum-fed exercised mice were more similar to those seen in ad libitum-fed non-exercised mice.
One interesting finding across our two experiments was that the median survival for the two groups of control mice was quite different, at 34 wks for the treadmill control (CON-TREX) vs. 24 wks for wheel control (CON-WHL) mice. This occurred despite no difference in the ages of the mice at randomization, and we do not have an explanation for the difference, other than the inherent differences between different litters of mice. We did randomize mice from each shipment into the treatment groups within a given experiment, so valid comparisons can only be made to the respective control group. This finding speaks to the need to always use concurrent control groups when using these animals.
We saw no effect of the exercise treatments on serum IGF-1 levels in these mice over time. A change in IGF-1 as a result of regular physical activity has been proposed as a potential mechanism to explain the apparent reduction in cancer risk in active individuals compared to their more sedentary peers (24); however, there is little evidence to support this (26). In particular, prior exercise studies in various rodent models have found that exercise does not lower circulating IGF-1 levels (4;9;10;25) unless the exercise is paired with calorie restriction(40). Consequently, if IGF-1 is important for mammary tumorigenesis, our treatment may have been insufficient. It is also possible that it is not the IGF-1 level alone that is important, and that accounting for levels of relevant binding proteins, IGFBP-3 in particular, might give a different result. To date though, evidence suggests that the ratio of IGF-1/IGFBP-3 is also not related to chronic physical activity (26).
To our knowledge, this is the first study to examine exercise effects in a transgenic model of mammary cancer. Clearly, exercise accelerated mammary tumor development in this mouse model. One possibility is that the exercise treatments may have increased expression of the Wnt-1 transgene, but our results indicate that this was not the case. We cannot rule out, however, that Wnt-1 expression was increased earlier on in the study or at some other time point that we did not measure. Alternatively, there are several important pathologic and physiologic differences between these mouse tumors and typical sporadic ER-positive mammary tumors arising in women (2).
One example of such physiologic differences suggests that there could be interactions between p53 status, responses to endocrine therapy, and exercise. For example, transgenic p53+/−:MMTV-Wnt-1 mice develop estrogen receptor (ER)-positive mammary adenocarcinomas, but these tumors are somewhat resistant to modulation of the estrogen pathway: female MMTV-Wnt-1 transgenic mice with ER-alpha deleted still develop mammary tumors, although these tumors arise much later in life than in MMTV-Wnt-1 transgenic mice with wild-type ER(3). Nevertheless, we have recently seen that mammary tumor development in MMTV-Wnt-1 transgenic mice with wild type p53 is significantly delayed by tamoxifen treatment, while p53+/−:MMTV-Wnt-1 transgenic mice are completely resistant to tamoxifen treatment (unpublished data). This is consistent with a hypothesis that we and others are currently testing, i.e., that p53 gene dosage impacts the response to endocrine therapy.
Unfortunately, we did not compare exercise effects in p53+/− and wild-type p53 MMTV Wnt-1 transgenic mice in the present study, but our data from this study conclusively show that physical activity enhances mammary tumor development in p53+/−:MMTV-Wnt-1 transgenic mice. These surprising findings suggest that there may be an interaction between p53 gene dosage and exercise that has not been previously reported, and if confirmed in future studies, could have important implications for women at high risk for breast cancer. Indeed, among epidemiologic studies there has been considerable heterogeneity in the physical activity and breast cancer association, with some, but not all studies finding that the association is limited to those without a family history of breast cancer(14). Thus, it is certainly plausible that individuals with certain genetic susceptibilities may not benefit, or may even be adversely affected, by increased physical activity of higher intensities.
Although we don’t know the specific intensity of the exercise in our study, we estimate both the wheel and treadmill running to be at least of a moderate intensity based on prior published reports(28). As can be seen by the attrition rate, it would be difficult to sustain a treadmill intervention of higher intensity over the course of such a long study. Recently, it has been demonstrated that mice voluntarily run on wheels at near maximal intensity (11), although the pattern is quite different from a single bout of treadmill running. In terms of distance run, the wheel runners voluntarily ran as much as four times farther than the treadmill runners; however, these p53+/−: MMTV-Wnt-1 transgenic mice ran considerably less than what has been reported previously in other models with different mouse strains (10;16). It is therefore possible that these mice did not have a sufficient volume of exercise to induce effects. Our work is limited by the fact that we did not measure markers of fitness in our mice. It is possible that the doses of exercise we chose were not sufficient to alter cardiorespiratory fitness, that the two different treadmill exercise treatments did not substantially differ, or that these transgenic mice did not respond to exercise as we would suspect. Our bone mineral density and muscle weight data do indicate some physiological response to the exercise, but oxygen consumption or muscle enzymes should be measured in future studies. Another limitation is that we do not have complete data on the wheel running distances throughout the study. There is also some inherent error in the recording system in that wheel revolutions are recorded when the wheel turns even if the mouse is not running (e.g., the mouse jumps off). Therefore, there is undoubtedly some error in the average running distances we report.
In this study of exercise in a p53-deficient transgenic model of mammary tumorigenesis, treadmill running exercise and voluntary wheel running were detrimental to overall survival, tumor burden, or both. Treadmill running increased the rate of tumor development, decreased survival time, and increased the proportion of mice with multiple mammary carcinomas. Although wheel running did not affect survival or tumor growth, it did result in increased incidence and multiplicity of mammary carcinomas. These effects occurred despite favorable changes in body weight and fatness and no difference in IGF-1 levels. They also contrast with data from concurrent studies showing that a reduction in caloric intake (30% CR, 14% CR or a one-day per week fast) significantly delays mammary tumor development in p53+/−:MMTV-Wnt-1 transgenic mice irregardless of p53 gene dosage(18). These findings suggest, for the first time, the possibility of an important interaction between p53 gene dosage and physical activity that may influence breast cancer preventive effects of exercise. In addition, they suggest that increased energy expenditure may not have the same effect as decreased energy intake on mammary tumor development in this model.
Acknowledgments
The U.S. Army Medical Research Material Command under DAMD 17-01-1-0338 supported this work (to LHC). This project has been funded in part with federal funds from the National Cancer Institute, National Institutes of Health, under contract N01-CO-12400. SDH was supported in part by the Breast Cancer Research Foundation and a DOD Concept Award (06-1-0636). The content of this publication does not necessarily reflect the views of policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government. The results of this study do not constitute endorsement by ACSM. We acknowledge the excellent technical support of Lisa Riffle and dedicate this paper to the memory of Heather Hill, our good friend and talented technician who lost her battle with ovarian cancer before this work was completed.
References
- 1.Berrigan D, Lavigne JA, Perkins SN, Nagy TR, Barrett JC, Hursting SD. Phenotypic effects of calorie restriction and insulin-like growth factor-1 treatment on body composition and bone mineral density of C57BL/6 mice: implications for cancer prevention. In Vivo. 2005;19:667–74. [PubMed] [Google Scholar]
- 2.Blackshear PE. Genetically engineered rodent models of mammary gland carcinogenesis: an overview. Toxicol Pathol. 2001;29:105–16. doi: 10.1080/019262301301418919. [DOI] [PubMed] [Google Scholar]
- 3.Bocchinfuso WP, Korach KS. Mammary gland development and tumorigenesis in estrogen receptor knockout mice. J Mammary Gland Biol Neoplasia. 1997;2:323–34. doi: 10.1023/a:1026339111278. [DOI] [PubMed] [Google Scholar]
- 4.Bravenboer N, Engelbregt MJT, Visser NA, Popp-Snijders C, Lips P. The effect of exercise on systemic and bone concentrations of growth factors in rats. J Orthopaed Res. 2001;19:945–9. doi: 10.1016/S0736-0266(01)00026-2. [DOI] [PubMed] [Google Scholar]
- 5.Cohen LA, Choi K, Backlund JY, Harris R, Wang CX. Modulation of N-nitrosomethylurea induced mammary tumorigenesis by dietary fat and voluntary exercise. In Vivo. 1991;5:333–44. [PubMed] [Google Scholar]
- 6.Cohen LA, Choi KW, Wang C. Influence of dietary fat, caloric restriction, and voluntary exercise on n-nitrosomethylurea-induced mammary tumorigenesis in rats. Cancer Research. 1988;48:4276–83. [PubMed] [Google Scholar]
- 7.Cohen LA, Kendall ME, Meschter C, Epstein MA, Reinhardt J, Zang E. Inhibition of rat mammary tumorigenesis by voluntary exercise. In Vivo. 1993;7:151–8. [PubMed] [Google Scholar]
- 8.Colbert LH, Davis JM, Essig DA, Ghaffar A, Mayer EP. Exercise and tumor development in a mouse predisposed to multiple intestinal adenomas. Med Sci Sports Exerc. 2000;32:1704–8. doi: 10.1097/00005768-200010000-00007. [DOI] [PubMed] [Google Scholar]
- 9.Colbert LH, Mai V, Perkins SN, Berrigan D, Lavigne JA, Wimbrow HH, et al. Exercise and intestinal polyp development in APCMin mice. Med Sci Sports Exerc. 2003;35:1662–9. doi: 10.1249/01.MSS.0000089349.54813.41. [DOI] [PubMed] [Google Scholar]
- 10.Colbert LH, Mai V, Tooze JA, Perkins SN, Berrigan D, Hursting SD. Negative energy balance induced by voluntary wheel running inhibits polyp development in APCMin mice. Carcinogenesis. 2006;27:2103–7. doi: 10.1093/carcin/bgl056. [DOI] [PubMed] [Google Scholar]
- 11.De Bono JP, Adlam D, Paterson DJ, Channon KM. Novel quantitative phenotypes of exercise training in mouse models. Am J Physiol Regul Integr Comp Physiol. 2006;290:R926–R934. doi: 10.1152/ajpregu.00694.2005. [DOI] [PubMed] [Google Scholar]
- 12.Donehower LA, Godley LA, Aldaz CM, Pyle R, Shi Y-P, Pinkel D, et al. Deficiency of p53 accelerates mammary tumorigenesis in Wnt-1 transgenic mice and promotes chromosomal instability. Genes & Development. 1995;9:882–95. doi: 10.1101/gad.9.7.882. [DOI] [PubMed] [Google Scholar]
- 13.Donehower LA, Harvey M, Slagle BL, McArthur MJ, Montgomery CA, Jr, Butel JS, et al. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumors. Nature. 1992;356:215–21. doi: 10.1038/356215a0. [DOI] [PubMed] [Google Scholar]
- 14.Friedenreich CM, Cust AE. Physical activity and breast cancer risk: impact of timing, type and dose of activity and population subgroup effects. Br J Sports Med. 2008;42:636–47. doi: 10.1136/bjsm.2006.029132. [DOI] [PubMed] [Google Scholar]
- 15.Gillette CA, Zhu Z, Westerlind KC, Melby CL, Wolfe P, Thompson HJ. Energy availability and mammary carcinogenesis: effects of calorie restriction and exercise. Carcinogenesis. 1997;18:1183–8. doi: 10.1093/carcin/18.6.1183. [DOI] [PubMed] [Google Scholar]
- 16.Hoffman-Goetz L, May KM, Arumugam Y. Exercise training and mouse mammary tumor metastasis. Anticancer Res. 1994;14:2627–32. [PubMed] [Google Scholar]
- 17.Hursting SD, Lavigne JA, Berrigan D, Perkins SN, Barrett JC. Calorie restriction, aging, and cancer prevention: mechanisms of action and applicability to humans. Annu Rev Med. 2003;54:131–52. doi: 10.1146/annurev.med.54.101601.152156. [DOI] [PubMed] [Google Scholar]
- 18.Hursting SD, Perkins SN, Colbert LH, Berrigan DA, Davis BJ, Fuchs-Young R, et al. Effects of energy balance modulation on spontaneous mammary tumorigenesis in p53-deficient, Wnt-1 transgenic mice. Cancer Epidemiol Biomarkers Prev. 2002;11:1154s. [Google Scholar]
- 19.Lane HW, Teer P, Keith RE, White MT, Strahan S. Reduced energy intake and moderate exercise reduce mammary tumor incidence in virgin female BALB/c mice treated with 7,12-dimethylbenz(a)anthracene. J Nutr. 1991;121:1883–8. doi: 10.1093/jn/121.11.1883. [DOI] [PubMed] [Google Scholar]
- 20.Lee I-M. Physical activity and cancer prevention - data from epidemiologic studies. Med Sci Sports Exerc. 2003;35:1823–7. doi: 10.1249/01.MSS.0000093620.27893.23. [DOI] [PubMed] [Google Scholar]
- 21.Li Y, Hively WP, Varmus HE. Use of MMTV-Wnt-1 transgenic mice for studying the genetic basis of breast cancer. Oncogene. 2000;19:1002–9. doi: 10.1038/sj.onc.1203273. [DOI] [PubMed] [Google Scholar]
- 22.Lu J, Xie L, Sylvester J, Wang J, Bai J, Baybutt R, et al. Different gene expression of skin tissues between mice with weight controlled by either calorie restriction or physical exercise. Exp Biol Med. 2007;232:473–80. [PubMed] [Google Scholar]
- 23.Mai V, Colbert LH, Berrigan D, Perkins SN, Pfeiffer R, Lavigne JA, et al. Calorie restriction and diet composition modulate spontaneous intestinal tumorigenesis in APCMin mice through different mechanisms. Cancer Res. 2003;63:1752–5. [PubMed] [Google Scholar]
- 24.McTiernan A, Ulrich C, Slate S, Potter J. Physical activity and cancer etiology: associations and mechanisms. Cancer Causes Control. 1998;9:487–509. doi: 10.1023/a:1008853601471. [DOI] [PubMed] [Google Scholar]
- 25.Mehl K, Davis J, Clements J, Berger F, Pena M, Carson J. Decreased intestinal polyp multiplicity is related to exercise mode and gender in APCMin/+ mice. J Appl Physiol. 2005;98:2219–25. doi: 10.1152/japplphysiol.00975.2004. [DOI] [PubMed] [Google Scholar]
- 26.Orenstein MR, Friedenreich CM. Review of physical activity and the IGF family. J Physical Act Aging. 2004;1:291–320. [Google Scholar]
- 27.Patel AC, Nunez NP, Perkins SN, Barrett JC, Hursting SD. Effects of energy balance on cancer in genetically altered mice. J Nutr. 2004;134:3394S–8S. doi: 10.1093/jn/134.12.3394S. [DOI] [PubMed] [Google Scholar]
- 28.Schefer V, Talan MI. Oxygen consumption in adult and aged C57BL/6J mice during acute treadmill exercise of different intensity. Experimental Gerontology. 1996;31:387–92. doi: 10.1016/0531-5565(95)02032-2. [DOI] [PubMed] [Google Scholar]
- 29.Shephard RJ. Exercise in the prevention and treatment of cancer. Sports Medicine. 1993;15:258–80. doi: 10.2165/00007256-199315040-00004. [DOI] [PubMed] [Google Scholar]
- 30.Thompson HJ, Ronan AM, Ritacco KA, Tagliaferro AR. Effect of type and amount of dietary fat on the enhancement of rat mammary tumorigenesis by exercise. Cancer Research. 1989;49:1904–8. [PubMed] [Google Scholar]
- 31.Thompson HJ, Ronan AM, Ritacco KA, Tagliferro AR, Meeker LD. Effect of exercise on the induction of mammary carcinogenesis. Cancer Research. 1988;48:2720–3. [PubMed] [Google Scholar]
- 32.Thompson HJ, Westerlind KC, Snedden J, Briggs S, Singh M. Exercise intensity dependent inhibition of 1-methyl-1-nitrosurea induced mammary carcinogenesis in female F-344 rats. Carcinogenesis. 1995;16:1783–6. doi: 10.1093/carcin/16.8.1783. [DOI] [PubMed] [Google Scholar]
- 33.Thompson HJ, Westerlind KC, Snedden JR, Briggs S, Singh M. Inhibition of mammary carcinogenesis by treadmill exercise. J Natl Cancer Inst. 1995;87:453–5. doi: 10.1093/jnci/87.6.453. [DOI] [PubMed] [Google Scholar]
- 34.Thompson HJ, Zhu Z, Jiang W. Weight control and breast cancer prevention: are the effects of reduced energy intake equivalent to those of increased energy expenditure? J Nutr. 2004;134:3407S–11S. doi: 10.1093/jn/134.12.3407S. [DOI] [PubMed] [Google Scholar]
- 35.Westerlind KC. Physical activity and cancer prevention - mechanisms. Med Sci Sports Exerc. 2003;35:1834–40. doi: 10.1249/01.MSS.0000093619.37805.B7. [DOI] [PubMed] [Google Scholar]
- 36.Westerlind KC, McCarty HL, Gibson KJ, Strange R. Effect of exercise on the rat mammary gland: implications for carcinogenesis. Acta Physiol Scand. 2002;175:147–56. doi: 10.1046/j.1365-201X.2002.00980.x. [DOI] [PubMed] [Google Scholar]
- 37.Westerlind KC, McCarty HL, Schultheiss PC, Story R, Reed AH, Baier ML, et al. Moderate exercise training slows mammary tumour growth in adolescent rats. Eur J Cancer Prev. 2003;12:281–7. doi: 10.1097/00008469-200308000-00007. [DOI] [PubMed] [Google Scholar]
- 38.Whittal KS, Parkhouse WS. Exercise during adolescence and its effects on mammary gland development, proliferation, and nitrosomethylurea (NMU) induced tumorigenesis in rats. Breast Cancer Res Treat. 1996;37:21–7. doi: 10.1007/BF01806628. [DOI] [PubMed] [Google Scholar]
- 39.Whittal-Strange KS, Chadau S, Parkhouse WS. Exercise during puberty and NMU induced mammary tumorigenesis in rats. Breast Cancer Res Treat. 1998;47:1–8. doi: 10.1023/a:1005838721890. [DOI] [PubMed] [Google Scholar]
- 40.Xie L, Jian Y, Ouyang P, Chen J, Doan H, Herndon B, et al. Effects of dietary calorie restriction or exercise on the PI3K and ras signalling pathways in the skin of mice. J Biol Chem. 2007;282:28025–35. doi: 10.1074/jbc.M604857200. [DOI] [PubMed] [Google Scholar]