


Abstract
Ribosomal protein S7 from Escherichia coli binds to the lower half of the 3′ major domain of 16S rRNA and initiates its folding. It also binds to its own mRNA, the str mRNA, and represses its translation. Using filter binding assays, we show in this study that the same mutations that interfere with S7 binding to 16S rRNA also weaken its affinity for its mRNA. This suggests that the same protein regions are responsible for mRNA and rRNA binding affinities, and that S7 recognizes identical sequence elements within the two RNA targets, although they have dissimilar secondary structures. Overexpression of S7 is known to inhibit bacterial growth. This phenotypic growth defect was relieved in cells overexpressing S7 mutants that bind poorly the str mRNA, confirming that growth impairment is controlled by the binding of S7 to its mRNA. Interestingly, a mutant with a short deletion at the C-terminus of S7 was more detrimental to cell growth than wild-type S7. This suggests that the C-terminal portion of S7 plays an important role in ribosome function, which is perturbed by the deletion.
INTRODUCTION
Ribosomal protein S7 from Escherichia coli is a primary binding protein that organizes the folding of the 3′ major domain of 16S rRNA and enables the subsequent binding of other ribosomal proteins, so as to form the head of the 30S subunit. It is located at the subunit interface, in proximity to the decoding center and its structure, which has been solved by X-ray crystallography for thermophilic bacteria, consists of a six α-helix bundle with a β-hairpin between helices 3 and 4 (1,2, reviewed in 3). Mutagenesis studies have been used to define which amino acid residues in S7 are involved in its interaction with rRNA (4,5) and it was found that several regions play an important role in S7 binding, including its N-terminal region, the β-hairpin, α-helix 4 and loops 2 and 5, connecting α-helices 1 and 2, and 4 and 5, respectively. In E.coli, the expression levels of most ribosomal proteins are regulated autogenously by primary binding proteins, which exert a feedback control at the translational level by binding to a specific site of their operon mRNA (reviewed in 6,7). S7 is one of these translational repressors (8). It is encoded by the str operon, which also codes for ribosomal protein S12, and elongation factors G and Tu. Overexpression of S7 interferes with bacterial growth (9), an effect which is assumed to result from repression of translation of the str operon mRNA upon S7 binding, causing an imbalance between rRNA transcription and ribosomal protein and elongation factor translation. The binding site of S7 in the lower half of the 3′ major domain of 16S rRNA was delineated to a short fragment comprising two multibranch loops (10), whereas its binding site on its mRNA is contained in an irregular hairpin structure located in the intercistronic region between the first cistron coding for S12 and the second cistron coding for S7 (11). Although these binding sites appear to have a very different secondary structure, they contain two identical sequence elements (Fig. 1). In 16S rRNA, one of these elements is located in loop A, at the junction between helices 30 and 41, and the other element is located in loop B, proximal to helix 43. Protection by S7 against hydroxyl radical attack, crosslinking studies and mutational and chemical probing of the S7 binding site (13–15, reviewed in 16) previously suggested that these two elements correspond to recognition features for S7 on 16S rRNA. The recent crystal structures of the bacterial 30S subunit at 3.0 and 3.3 Å resolution (17,18) confirmed this suggestion, in showing that these two elements contact specific amino acid residues in protein S7. The element in loop A of 16S rRNA interacts with residues in loop 2 and loop 5 of protein S7, whereas the element in loop B of 16S rRNA interacts with residues in the N-terminal region, in loop 2, in the β-hairpin and in α-helix 4 of protein S7. The binding of S7 to its mRNA is much less characterized than its binding to 16S rRNA, but it could be hypothesized that the two elements that are common to 16S rRNA and the str mRNA also constitute the recognition signals for S7 on its mRNA.
Figure 1.

Secondary structure of mRNA and 16S rRNA fragments that bind S7. (A) S12–S7 intercistronic region of the str mRNA that contains the S7 binding site. The stop codon for S12 (UAA) and the initiation codon for S7 (AUG) are underlined. The Shine–Dalgarno sequence for the S7 cistron is indicated by a dashed line. (B) Lower half of the 3′ major domain of 16S rRNA, with the circled sequence containing the minimal binding site for S7 (10). Helices are numbered according to Brimacombe (12). The sequences that are boxed are identical in the mRNA and the rRNA. The arrows point to bases in 16S rRNA that were crosslinked to S7 (15).
In this study we have compared the effect of various deletion and point mutations in E.coli S7 on its binding to its own mRNA and to 16S rRNA. We also investigated the effect of mutations in S7 on its capacity to interfere with cell growth, allowing a direct comparison between the binding affinity of the repressor for its mRNA and its in vivo effectiveness. Our results suggest that S7 uses the same features to recognize its binding site on its mRNA and on 16S rRNA, and demonstrate a direct relationship between the capacity of the protein to bind its mRNA and its ability to interfere with cell growth.
MATERIALS AND METHODS
Construction of plasmids and expression of S7 and its mutant derivatives
Plasmid pET-21a(+)-S7, which codes for E.coli K12 S7 under control of a T7 promoter, and its derivatives coding for various S7 deletion and point mutants were previously described (5). These plasmids were used to transform E.coli BL21(DE3)/pLysS (Novagen), the recombinant proteins were expressed after induction with isopropyl-d-galactopyranoside (IPTG) and purified by affinity chromatography as described (5). Plasmid pcDNA3.1/Hygro(+) (Invitrogen), which allows in vitro transcription of a cloned DNA sequence with the T7 RNA polymerase, was used for the cloning of the intercistronic S12–S7 region of the str operon, extending on the S12 and S7 coding sequences. Genomic DNA was obtained from E.coli K12A19 following a standard procedure (19), and we PCR amplified the sequence corresponding to bases –160 to +71 (the first base of S7 initiation codon on the str mRNA corresponds to position 1). The forward primer (#1) was 5′-GCGCGGCTAGCCTCCGGCGTTAAAGACCGTAAGC-3′ and the reverse primer (#2) 5′-GCGCGTTCGAACCAGCAGTTCTGATCCGAACTTCG-3′ (bold letters in the sequences correspond to NheI and HindIII restriction sites, respectively). These sites were used for the cloning of the PCR fragment containing the S12–S7 intercistronic region into pcDNA3.1/Hygro(+), generating pcDNA3.1/Hygro(+)-S12ICRS7. PCR was carried out using the Vent DNA polymerase (New England Biolabs) in a RobocyclerTM 40 (Stratagene) under the following conditions: 4 min of denaturation at 94°C, 28 cycles of 1 min at 94°C, 1 min at 58°C and 1 min at 72°C, followed by a final extension step of 5 min at 72°C.
RNA preparation and nitrocellulose filter binding assays
The 32P-labeled fragment of the str operon used for binding studies was obtained by run-off transcription in the presence of [32P]UTP of plasmid pcDNA3.1/Hygro(+)-S12ICRS7 linearized with HindIII. The RNA fragment of the str mRNA contains 252 nt, of which 230 nt encompass the end of the S12 sequence, the intercistronic region and the beginning of the S7 sequence, and 22 nt originate from the vector. A 32P-labeled fragment of 218 nt corresponding to the lower half of the 3′ major domain of 16S rRNA and containing the S7 binding site was generated as described (5). Binding assays were done as described previously (5), in 50 µl of binding buffer [50 mM HEPES–KOH, pH 7.8, 20 mM Mg(OAc)2, 350 mM KOAc, 5 mM β-mercaptoethanol] with increasing concentrations of protein.
Growth assays
Twenty milliliters of LB medium were inoculated with a single colony of E.coli BL21(DE3)/pLysS or a recA derivative, BLR(DE3)/pLysS, transformed with either pET-21a(+)-S7 or its various derivative plasmids coding for the S7 mutants. When the optical density at 600 nm was 0.05, IPTG was added to the cell culture to a final concentration of 0.75 mM and cell growth was monitored at 37°C by measuring the optical density at 600 nm every 20 min. Cell doubling time was calculated using GraphPad Prism version 3.00 for Windows.
RESULTS
To identify amino acid residues critical for the binding of S7 to 16S rRNA, a set of four deletion and 12 single amino acid substitution mutants had been previously generated (5; Fig. 2), and circular dichroism spectra had indicated that the mutant proteins did not suffer major structural perturbations (5). The same mutants were investigated for their capacity to interact with the str mRNA fragment encompassing the S7 binding site in a high-ionic-strength buffer [20 mM Mg(OAc)2, 350 mM KOAc]. With this buffer, the maximal retention efficiency was ~35% for the complex between the str mRNA fragment and wild-type S7, but was <20% in a buffer containing chloride instead of acetate anions. We therefore selected the acetate buffer for studying the interaction between S7 mutants and the str mRNA fragment. The apparent association constant (K′a) of wild-type S7 for its mRNA was determined from the amount of protein required to half-saturate the RNA. Its value (6.8 ± 0.7 µM–1) was identical to that measured for the association constant between S7 and the 16S rRNA fragment (7.1 ± 1.4 µM–1), a value comparable to that of 5.3 ± 0.5 µM–1 found previously in a slightly different high-ionic-strength buffer containing chloride and not acetate anions (5).
Figure 2.
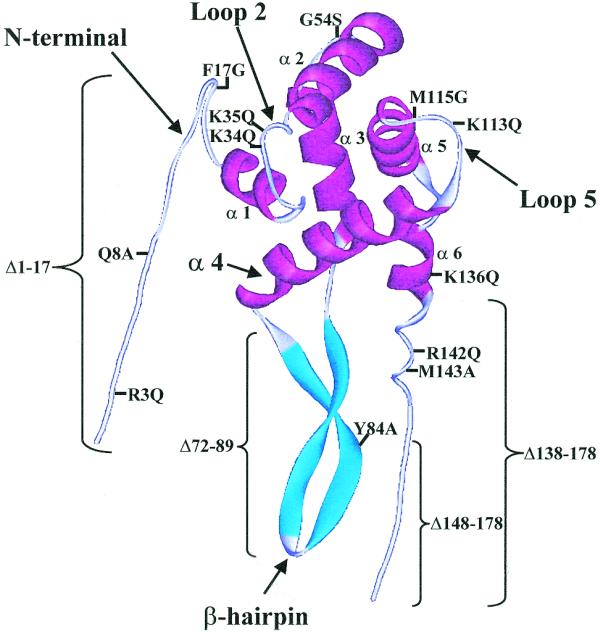
Location of the S7 mutations in the S7 structure. Crystallographic structure of S7 adapted from Hosaka et al. (1), showing the deletion and point mutations that were investigated in this study. In the case of the β-hairpin deletion, the 72–89 sequence was replaced with a short flexible loop, RRGGGGS, so as not to perturb the structure of the protein (5). The regions of S7 involved in binding to 16S rRNA, as determined by mutagenesis studies (4,5) and analysis of the crystal structures of the 30S subunit (17,18), are indicated by arrows.
Representative binding curves for different mutants are shown in Figure 3. The apparent association constants of the mutant derivatives of S7 for the str mRNA fragment are presented in Table 1, and the effect of the mutations on S7 binding to the str mRNA is compared to their effect on S7 binding to 16S rRNA. Several deletion and point mutations located throughout the protein sequence significantly decreased the affinity of S7 for its mRNA. They include an N-terminal deletion (Δ1–17), deletion of the β-hairpin (Δ72–89), point mutations in the N-terminal region (Q8A, F17G), mutation K35Q in loop 2 and mutation M115G in loop 5. Interestingly, the same mutations had been found to significantly interfere with S7 binding to the rRNA (5). The other mutations modestly or hardly affected S7 binding to the mRNA, comparable to their effect on rRNA binding. The fact that the effects of mutations are similar for 16S rRNA and mRNA binding strongly suggests that S7 uses the same features to bind the two RNA targets.
Figure 3.

Binding curves for the interaction of wild-type S7 and its mutant derivatives with str mRNA. The curves show binding isotherms measured by a nitrocellulose filter binding assay. They correspond to the wild-type protein and to representative examples of mutations that affect binding modestly, strongly or dramatically. The portion of the str mRNA containing the S7 binding site was synthesized in vitro and incubated with increasing amounts of protein. Apparent association constants (K′a) between the str mRNA and S7 or its mutant derivatives were calculated from the binding isotherms and are presented in Table 1.
Table 1. Affinity of wild-type and mutated S7 for its mRNA.
| Mutation |
K′a (µM–1) |
Relative affinity for str mRNA |
Relative affinity for 16S rRNAa |
| None |
6.8 ± 0.9 |
1.00 |
1.00 |
| Δ1–17 |
0.6 ± 0.1 |
0.09 |
n.d. |
| Δ72–89 |
1.9 ± 0.3 |
0.28 |
0.34 |
| Δ138–178 |
3.7 ± 0.4 |
0.54 |
0.58 |
| Δ148–178 |
6.6 ± 1.2 |
0.97 |
1.06 |
| R3Q |
2.9 ± 1.1 |
0.43 |
0.57 |
| Q8A |
1.8 ± 0.3 |
0.26 |
0.21 |
| F17G |
1.3 ± 0.2 |
0.19 |
0.17 |
| K34Q |
6.7 ± 0.9 |
0.99 |
1.04 |
| K35Q |
1.9 ± 0.3 |
0.28 |
0.45 |
| G54S |
3.3 ± 0.8 |
0.49 |
0.40 |
| Y84A |
4.7 ± 0.6 |
0.69 |
0.64 |
| K113Q |
4.0 ± 0.7 |
0.59 |
0.42 |
| M115G |
0.7 ± 0.1 |
0.10 |
0.26 |
| K136Q |
6.0 ± 0.8 |
0.88 |
0.98 |
| R142Q |
5.6 ± 0.9 |
0.82 |
0.83 |
| M143A | 4.4 ± 0.8 | 0.65 | 0.68 |
A fragment of the str operon mRNA encompassing the intercistronic region between S12 and S7 coding sequences was synthesized in vitro and incubated with increasing amounts of protein. The binding affinity of wild-type and mutated S7 for the str mRNA fragment was measured by a nitrocellulose filter binding assay. Binding assays were performed in a high-ionic-strength buffer [20 mM Mg(OAc)2, 350 mM KOAc]. K′a values are means ± standard deviation of at least three independent experiments; n.d., not detectable.
aThe relative affinity of the S7 mutants for a fragment corresponding to the lower half of the 3′ major domain of 16S rRNA was determined by Robert et al. (5) in a high-ionic-strength buffer comparable to that used in this study (20 mM MgCl2, 300 mM KCl).
The effect of overexpression of the S7 mutants on cell growth was assessed by measuring the cell doubling time, under conditions where wild-type chromosome-encoded S7 was also expressed. A direct correlation could be readily observed between the capacity of the mutants to bind the str mRNA and their ability to interfere with cell growth (Table 2). Mutants that bind poorly the str mRNA had only a weak effect on cell growth. This was the case for mutants Δ1–17, Δ72–89, Q8A, F17G, K35Q and M115G. On the other hand, mutants that bind efficiently to the mRNA such as K34Q, K136Q and R142Q strongly interfered with bacterial growth as does wild-type S7. One of the mutants, Δ148–178, with a short C-terminal deletion, was even more detrimental than wild-type S7.
Table 2. Effect of overexpression of S7 and its mutant derivatives on bacterial growth.
| Plasmid |
Doubling time (min) |
| pET-21a (+) |
62 ± 9 |
| pET-21a (+)-S7 |
174 ± 18 |
| pET-21a (+)-S7Δ1–17 |
92 ± 13 |
| pET-21a (+)-S7Δ72–89 |
87 ± 9 |
| pET-21a (+)-S7Δ138–178 |
144 ± 11 |
| pET-21a (+)-S7Δ148–178 |
No growth |
| pET-21a (+)-S7R3Q |
145 ± 8 |
| pET-21a (+)-S7Q8A |
95 ± 9 |
| pET-21a (+)-S7F17G |
97 ± 13 |
| pET-21a (+)-S7K34Q |
172 ± 10 |
| pET-21a (+)-S7K35Q |
96 ± 9 |
| pET-21a (+)-S7G54S |
122 ± 15 |
| PET-21a (+)-S7Y84A |
123 ± 7 |
| pET-21a (+)-S7K113Q |
117 ± 8 |
| pET-21a (+)-S7M115G |
97 ± 8 |
| pET-21a (+)-S7K136Q |
161 ± 13 |
| pET-21a (+)-S7R142Q |
167 ± 16 |
| pET-21a (+)-S7M143A | 131 ±12 |
BL21(DE3)/pLysS cells were transformed with plasmid pET-21a(+)-S7, containing the gene coding for wild-type S7 under control of a T7 promoter, or with derivatives of this plasmid coding for S7 mutants, and the growth rate of the bacterial cultures was monitored in the LB medium at 37°C. Doubling time values are means ± standard deviation of at least three independent experiments. Identical results were obtained with BLR(DE3)/pLysS cells (data not shown).
DISCUSSION
Our results show that E.coli S7 binds its mRNA and 16S rRNA with the same affinity. S8, a regulatory ribosomal protein, binds its mRNA with an affinity which is ~5-fold less than its affinity for the rRNA (20), whereas S4, another regulatory ribosomal protein, has the same affinity for its mRNA and for the rRNA (21). One would have expected regulatory ribosomal proteins to bind preferentially the rRNA, so that they repress translation of their mRNA only when there is a shortage of rRNA transcript. Deckman and Draper (21) concluded from their studies with S4 that the preferential incorporation of regulatory ribosomal proteins into ribosomes results from the high cooperativity of ribosome assembly. Our observation that S7 binds its mRNA and the 16S rRNA with the same affinity fully supports this conclusion.
As indicated in the Introduction, the binding sites of S7 on 16S rRNA and on its mRNA, although apparently dissimilar at the secondary structure level, present two identical sequence elements, which were identified as recognition elements for S7 binding in rRNA. Our results with a variety of S7 mutants clearly suggest that S7 uses the same determinants to bind the str mRNA and 16S rRNA since the same mutations which decreased S7 binding to rRNA decreased its affinity for the mRNA. Thus, it is very likely that S7 recognizes these two identical elements on the mRNA and 16S rRNA. Among ribosomal proteins acting as translational repressors, the binding sites for S8 and L1 on their mRNA are strikingly similar to their binding sites on the rRNA, from which it can readily be inferred that they recognize identical elements on the two targets (reviewed in 22). In contrast, like for S7, the mRNA binding sites of the translational repressors S4 and S15 seem very different from their binding sites on the rRNA (reviewed in 23). However, S4 appears to use the same determinants to bind rRNA and its mRNA since S4 deletions that decreased rRNA recognition had parallel effects on mRNA binding (24). As to S15, its interaction with 16S rRNA has been characterized in detail from the crystallographic structure of a complex between S15 from a thermophilic bacterium and a 16S rRNA fragment encompassing its binding site (25), as well as from footprinting and interference studies (14,26,27). Footprinting and mutagenesis studies with its mRNA binding site (28,29) strongly suggest that S15 recognizes similar elements on both rRNA and its mRNA. Therefore, it appears to be a common characteristic of ribosomal proteins acting as translational repressors that they recognize the same elements in their mRNA and in their rRNA binding site, whether these sites appear similar or dissimilar. Translation control involving the binding of a regulatory ribosomal protein generally causes translation repression by inducing a conformational change in mRNA that hinders ribosome binding or, alternatively, does not prevent ribosome binding but traps the bound ribosome in an inactive state (6,7). In the case of S7 that binds between the first and second cistrons of the str mRNA, the S7 cistron is translated by ribosomes, which, for the vast majority, have translated the upstream S12 cistron, scanned the intercistronic S12–S7 region and resumed translation when reaching the S7 translation start site (8). It has been proposed that the binding of S7 in the intercistronic S12–S7 region induces a conformational change in this region that prevents its scanning by the ribosomes (11). Our results, suggesting that S7 recognizes two specific sequence elements within the S12–S7 intercistronic region, agree with this hypothesis.
We observed that overexpression of wild-type S7 interferes with cell growth, which, as mentioned above, can be related to binding of its own polycistronic mRNA, the str mRNA, and repressing its translation (8,9). Our results show that there is a direct correlation between the capacity of each S7 mutant to bind the str mRNA in vitro and the effect of its overexpression on bacterial growth. Indeed, mutants that bind well the str RNA efficiently interfered with cell growth whereas overexpression of the mutants that bind poorly the str mRNA only weakly affected cell growth. Alternatively, these mutants that weakly interfere with cell growth could be rapidly degraded in vivo, thus accounting for their lack of effect. However, examination of the amount of the overexpressed S7 proteins at different times of bacterial growth does not support this possibility (data not shown). Interestingly, Fredrick et al. (9), who also investigated the effect of various mutations in S7 on cell growth, found a close correlation between the capacity of different S7 mutants to interfere with cell growth and their proficiency in 30S subunit assembly and suggested that S7 residues involved in mRNA binding are also involved in rRNA binding. This suggestion is directly confirmed by the results of the present study.
In our in vivo assays, the chromosome-encoded wild-type S7 is coexpressed with the plasmid-encoded S7 mutants. As suggested by the results from Fredrick et al. (9), it is likely that the 30S subunits contain almost exclusively wild-type S7 when the coexpressed S7 mutants bind poorly the 16S rRNA, whereas S7 mutants that bind efficiently 16S rRNA can outcompete wild-type S7 for the formation of 30S subunits. Therefore, with these mutants that conserve a high affinity for 16S rRNA as well as for the str mRNA, effects on growth could result not only from S7 binding to the str mRNA and repressing its translation but also from a decreased activity of the ribosomes having incorporated the mutated protein. Mutant Δ148–178, with a deletion of 31 amino acids at the C-terminal end, likely corresponds to such a case. This mutant binds well to 16S rRNA and to the str mRNA, and its overexpression is much more detrimental than that of wild-type S7. One could argue here that mutant Δ138–178, with a larger deletion at the C-terminus, is not as detrimental as Δ148–178, although it also lacks the sequence deleted in Δ148–178. However, it is likely that mutant Δ138–178 is less efficiently incorporated into the 30S subunit than mutant Δ148–178, due to its weaker binding affinity for the rRNA. Greuer et al. (30) recently showed that a region in the C-terminus of E.coli S7 (positions 149–155) could be crosslinked to a mRNA analog, in the spacer between the Shine–Dalgarno sequence and the initiator codon. An attractive hypothesis is that the C-terminal region of S7 participates in the formation of a functional 30S initiation complex and that 30S subunits having incorporated mutant Δ148–178 would be impaired in translation initiation. Experiments are in progress in our laboratory to investigate this hypothesis. Interestingly, the portion of the C-terminal region of S7 that is crosslinked to mRNAs is extremely well-conserved in bacteria (1,2), but not in the eukaryotic homolog of S7 (31,32). This could be related to the known difference in the way prokaryotic and eukaryotic ribosomes initiate protein synthesis (33).
Acknowledgments
ACKNOWLEDGEMENTS
We thank François Dragon and Stephen Michnick for stimulating discussions and for comments on this manuscript, and Serguei Chteinberg and Katia Zagriadskaïa for their help in the analysis of the sites of interaction between S7 and 16S rRNA in the crystal structures of the 30S subunit. This study was supported by a grant from the Medical Research Council of Canada.
References
- 1.Hosaka H., Nakagawa,A., Tanaka,I., Harada,N., Sano,K., Kimura,M., Yao,M. and Wakatsuki,S. (1997) Ribosomal protein S7: a new RNA-binding motif with structural similarities to a DNA architectural factor. Structure, 5, 1199––1208.. [DOI] [PubMed] [Google Scholar]
- 2.Wimberly B.T., White,S.W. and Ramakrishnan,V. (1997) The structure of ribosomal protein S7 at 1.9 Å resolution reveals a β-hairpin motif that binds double-stranded nucleic acids. Structure, 5, 1187––1198.. [DOI] [PubMed] [Google Scholar]
- 3.White S.W., Clemons,W.M.,Jr, Davies,C., Ramakrishnan,V. and Wimberly,B.T. (2000) Structures of bacterial ribosomal proteins: high-resolution probes of the architecture and mechanism of the ribosome. In Garrett,R.A., Douthwaite,S.R., Liljas,A., Matheson,A.T., Moore,P.B. and Noller,H.F. (eds), The Ribosome: Structure, Function,Antibiotics,and Cellular Interactions. ASM Press, Washington, DC, pp. 73–83.
- 4.Miyamoto A., Usui,M., Yamasaki,N., Yamada,N., Kuwano,E., Tanaka,I. and Kimura,M. (1999) Role of the N-terminal region of ribosomal protein S7 in its interaction with 16S rRNA which binds to the concavity formed by the β-ribbon arm and the α-helix. Eur. J. Biochem., 266, 591––598.. [DOI] [PubMed] [Google Scholar]
- 5.Robert F., Gagnon,M., Sans,D., Michnick,S. and Brakier-Gingras,L. (2000) Mapping of the RNA recognition site of Escherichia coli ribosomal protein S7. RNA, 6, 1649––1659.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Keener J. and Nomura,M. (1996) Regulation of ribosome synthesis. In Neidhardt,F.C., Ingraham,J.L., Low,K.B., Magasanik,B., Schaechter,M. and Umbarger,H.E. (eds), Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd Edn. ASM Press, Washington, DC, Vol. 1, pp. 1417–1431.
- 7.Zengel J.M. and Lindahl,L. (1994) Diverse mechanisms for regulating ribosomal protein synthesis in Escherichia coli. Prog. Nucleic Acids Res. Mol. Biol., 47, 331––370.. [DOI] [PubMed] [Google Scholar]
- 8.Saito K., Mattheakis,L.C. and Nomura,M. (1994) Post-transcriptional regulation of the str operon in Escherichia coli. Ribosomal protein S7 inhibits coupled translation of S7 but not its independent translation. J. Mol. Biol., 235, 111––124.. [DOI] [PubMed] [Google Scholar]
- 9.Fredrick K., Dunny,G.M. and Noller,H.F. (2000) Tagging ribosomal protein S7 allows rapid identification of mutants defective in assembly and function of 30S subunits. J. Mol. Biol., 298, 379––394.. [DOI] [PubMed] [Google Scholar]
- 10.Dragon F. and Brakier-Gingras,L. (1993) Interaction of Escherichia coli ribosomal protein S7 with 16S rRNA. Nucleic Acids Res., 21, 1199––1203.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Saito K. and Nomura,M. (1994) Post-transcriptional regulation of the str operon in Escherichia coli. Structural and mutational analysis of the target site for translational repressor S7. J. Mol. Biol., 235, 125––139.. [DOI] [PubMed] [Google Scholar]
- 12.Brimacombe R. (1991) RNA-protein interactions in the Escherichia coli ribosome. Biochimie, 73, 927––936.. [DOI] [PubMed] [Google Scholar]
- 13.Dragon F., Payant,C. and Brakier-Gingras,L. (1994) Mutational and structural analysis of the RNA binding site for Escherichia coli ribosomal protein S7. J. Mol. Biol., 244, 74––85.. [DOI] [PubMed] [Google Scholar]
- 14.Powers T. and Noller,H.F. (1995) Hydroxyl radical footprinting of ribosomal proteins on 16S rRNA. RNA, 1, 194––209.. [PMC free article] [PubMed] [Google Scholar]
- 15.Urlaub H., Thiede,B., Müller,E.C., Brimacombe,R. and Wittmann-Liebold,B. (1997) Identification and sequence analysis of contact sites between ribosomal proteins and rRNA in Escherichia coli 30S subunits by a new approach using matrix-assisted laser desorption/ionization-mass spectrometry combined with N-terminal microsequencing. J. Biol. Chem., 272, 14547––14555.. [DOI] [PubMed] [Google Scholar]
- 16.Mueller F. and Brimacombe,R. (1997) A new model for the three-dimensional folding of Escherichia coli 16S ribosomal RNA. II. The RNA-protein interaction data. J. Mol. Biol., 271, 545––565.. [DOI] [PubMed] [Google Scholar]
- 17.Schluenzen F., Tocilj,A., Zarivach,R., Harms,J., Gluehmann,M., Janell,D., Bashan,A., Bartels,H., Agmon,I., Franceschi,F. and Yonath,A. (2000) Structure of functionally activated small ribosomal subunit at 3.3 Å resolution. Cell, 102, 615––623.. [DOI] [PubMed] [Google Scholar]
- 18.Wimberly B.T., Brodersen,D.E., Clemons,W.M., Morgan-Warren,R.J., Carter,A.P., Vonrhein,C., Hartsch,T. and Ramakrishnan,V. (2000) Structure of the 30S ribosomal subunit. Nature, 407, 327––339.. [DOI] [PubMed] [Google Scholar]
- 19.Marmur J. (1961) A procedure for the isolation of deoxyribonucleic acid from microorganisms. J. Mol. Biol., 3, 208––218.. [Google Scholar]
- 20.Gregory R.J., Cahill,P.B.F., Thurlow,D.L. and Zimmermann,R.A. (1998) Interaction of Escherichia coli ribosomal protein S8 with its binding sites in ribosomal RNA and messenger RNA. J. Mol. Biol., 204, 295––307.. [DOI] [PubMed] [Google Scholar]
- 21.Deckman I.C. and Draper,D.E. (1985) Specific interaction between ribosomal protein S4 and the α operon messenger RNA. Biochemistry, 24, 7860––7865.. [DOI] [PubMed] [Google Scholar]
- 22.Zimmermann R.A., Alimov,A., Uma,K., Wu,H., Wower,I., Nikonowicz,E.P., Drygin,D., Dong,P. and Jiang,L. (2000) How ribosomal proteins and rRNA recognize one another. In Garrett,R.A., Douthwaite,S.R., Liljas,A., Matheson,A.T., Moore,P.B. and Noller,H.F. (eds), The Ribosome: Structure, Function, Antibiotics, and Cellular Interactions. ASM Press, Washington, DC, pp. 93–104.
- 23.Draper D.E. (1996) Ribosomal protein-RNA interactions. In Zimmermann,R.A. and Dahlberg,A.E. (eds), Ribosomal RNA: Structure, Evolution, Processing and Function in Protein Synthesis. CRC Press, Boca Raton, FL, pp. 171–197.
- 24.Baker A.-M. and Draper,D.E. (1995) Messenger RNA recognition by fragments of ribosomal protein S4. J. Biol. Chem., 270, 22939––22945.. [DOI] [PubMed] [Google Scholar]
- 25.Nikulin A., Serganov,A., Ennifar,E., Tishchenko,S., Nevskaya,N., Shepard,W., Portier,C., Garber,M., Ehresmann,B., Ehresmann,C., Nikonov,S. and Dumas,P. (2000) Crystal structure of the S15-rRNA complex. Nature Struct. Biol., 7, 273––277.. [DOI] [PubMed] [Google Scholar]
- 26.Batey R.T. and Williamson,J.R. (1996) Interaction of the Bacillus stearothermophilus ribosomal protein S15 with 16 S rRNA: II. Specificity determinants of RNA-protein recognition. J. Mol. Biol., 261, 550––567.. [DOI] [PubMed] [Google Scholar]
- 27.Serganov A.A., Masquida,B., Westhof,E., Chachia,C., Portier,C., Garber,M., Ehresmann,B. and Ehresmann,C. (1996) The 16S rRNA binding site of Thermus thermophilus ribosomal protein S15: comparison with Escherichia coli S15, minimum site and structure. RNA, 2, 1124––1138.. [PMC free article] [PubMed] [Google Scholar]
- 28.Bénard L., Mathy,N., Grunberg-Manago,M., Ehresmann,B., Ehresmann,C. and Portier,C. (1998) Identification in a pseudoknot of a U.G motif essential for the regulation of the expression of ribosomal protein S15. Proc. Natl Acad. Sci. USA, 95, 2564––2567.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Phillippe C., Bénard,L., Portier,C., Westhof,E., Ehresmann,B. and Ehresmann,C. (1995) Molecular dissection of the pseudoknot governing the translational regulation of Escherichia coli ribosomal protein S15. Nucleic Acids Res., 23, 18––28.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Greuer B., Thiede,B. and Brimacombe,R. (1999) The cross-link from the upstream region of mRNA to ribosomal protein S7 is located in the C-terminal peptide: experimental verification of a prediction from modelling studies. RNA, 5, 1521––1525.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kuwano Y., Olvera,J. and Wool,I.G. (1992) The primary structure of rat ribosomal protein S5. A ribosomal protein present in the rat genome in a single copy. J. Biol. Chem., 267, 25304––25308.. [PubMed] [Google Scholar]
- 32.Vladimirov S.N., Ivanov,A.V., Karpova,G.G., Musolyamov,A.K., Egorov,T.A., Thiebe,B., Wittmann-Liebold,B. and Otto,A. (1996) Characterization of the human small-ribosomal-subunit proteins by N-terminal and internal sequencing, and mass spectrometry. Eur. J. Biochem., 239, 144––149.. [DOI] [PubMed] [Google Scholar]
- 33.Jackson R.J. (1996) A comparative view of initiation site selection mechanisms. In Hershey,J.W.B., Mathews,M.B. and Sonenberg,N. (eds), Translational Control. Cold Spring Harbor Laboratory Press, New York, NY, pp. 71–112.