

Abstract
A recent evaluation of upper first molar (M1) crown size and cusp proportions in the genus Homo (Quam et al. 2009) describes Homo antecessor as maintaining a primitive pattern of cusp proportions, similar to that identified in australopithecines and the earliest members of the genus Homo. These results contrast with those of Gómez-Robles et al. (2007), who described the crown shape in these molars as derived and similar to Neanderthals and European Homo heidelbergensis. The reassessment of these measurements following the same methodology described by Quam et al. (2009) in all the M1s that are currently part of the hypodigm of H. antecessor demonstrates that the fossils from TD6 not only have the same cusp proportions identified in later Homo species, but also a strongly reduced metacone and a large hypocone shared with Middle and Upper Pleistocene members of the Neanderthal lineage. The evolutionary significance of these features should be evaluated in light of the results provided by recently discovered dental, cranial, mandibular, and postcranial H. antecessor fossils.
Keywords: Atapuerca, cusp base areas, dental anthropology, dental morphometrics, TD6
Introduction
Several recent works have assessed the morphological differences in upper first molar morphology among different hominin species (Bailey, 2004; Gómez-Robles et al. 2007; Quam et al. 2009). Among these, Bailey (2004) found a Neanderthal-specific morphology based on the relative proportion of cusp areas and the relative size of the occlusal polygon. Gómez-Robles et al. (2007) identified similar morphologies in European Lower and Middle Pleistocene specimens by using geometric morphometric methods, and Quam et al. (2009) proposed a derived pattern of relative cusp proportions shared by most species of the genus Homo. Following Quam et al. (2009)H. antecessor maintains a primitive molar morphology, shared with the earliest members of the genus, consisting of a relative metacone area larger than the relative paracone area. However, in later Homo species (including Homo ergaster, Homo erectus, Homo heidelbergensis, Homo neanderthalensis and Homo sapiens) ‘the relative area relationships have shifted so that the relative area of the paracone is now larger than that of the metacone’. These results contrast with those of Gómez-Robles et al. (2007), who described H. antecessor morphology as derived and similar to that of Neanderthals and European H. heidelbergensis. However, Quam et al. (2009) did not directly measure the relative areas they used to infer the primitive/derived condition of the upper first molars of H. antecessor, but rather, relied on the mean values published in Bermúdez de Castro et al. (1999). In light of the contradiction between the results of Quam et al. (2009) and Gómez-Robles et al. (2007), a detailed quantification of the absolute and relative cups areas of all the upper first molars that are part of the current hypodigm of the species H. antecessor is provided.
Materials and methods
Five upper first molars of the species H. antecessor were measured by one of us (A.G.-R.). This sample includes both antimeres of the maxilla ATD6-69 (individual H3 in Bermúdez de Castro et al. 1999), both antimeres of the individual H1 (ATD6-10 and ATD6-11), and a right upper first molar (ATD6-103) not published in detail so far (but included in Gómez-Robles et al. 2007) and currently attributed to individual H5 (Bermúdez de Castro et al. 2010a; see also Bermúdez de Castro et al. 1999, 2006).
Relative and absolute cusp base areas of the molars were measured using standardized images of the occlusal surface of the original molars. As in similar works based on image analysis of the occlusal surface of teeth, photographs were taken with a Nikon D1H digital camera fitted with an AF Micro-Nikkor 105 mm, f/2.8D. The camera was attached to a Kaiser Copy Stand kit RS-1 with grid baseboard, column, and adjustable camera arm. For maximum depth of field, an aperture of f/32 was used. The magnification ratio was adjusted to 1 : 1, and a scale was placed parallel to and at the same distance from the lens as the occlusal plane.
ImageJ software was used to measure the areas from the occlusal photographs using the general methodology described elsewhere (Wood & Engleman, 1988; Bailey, 2004; Quam et al. 2009). Individual cusp base areas were measured by following the main fissures of the occlusal surface. The part of the main fissures eroded by wear were estimated by extrapolating the course of the preserved part of the fissures, but no correction was applied to interproximal wear (the TD6 molars are only minimally worn on their mesial and distal faces). However, some TD6 molars do present a crista obliqua connecting the metacone with the protocone that makes it difficult to separate these two cusps. In molars with this crest, the protocone and the metacone have been differentiated by continuing the course of the central groove between these two cusps (see Figs 1 and 2). The M1s of individual H3 (maxilla ATD6-69) present a c5 or metaconule (Fig. 2). Following Bailey (2004) and Quam et al. (2009), the area of this accessory cusp was measured and divided between the adjacent cusps (metacone and hypocone). The relative occlusal polygon area (area defined by the lines connecting the cusp tips divided by the total crown base area; Bailey, 2004) and cusp angles were measured in the only molar that preserves the four cusp apices due to its minimal degree of wear (ATD6-103). In general terms, the levels of interobserver measurement error in cusp base areas have been described as similar to the levels of intraobserver error and, in any case, lower than 4.5%, thus making the measurements taken by different authors comparable when they are adjusted to the same standards (Bailey et al. 2004).
Fig. 1.
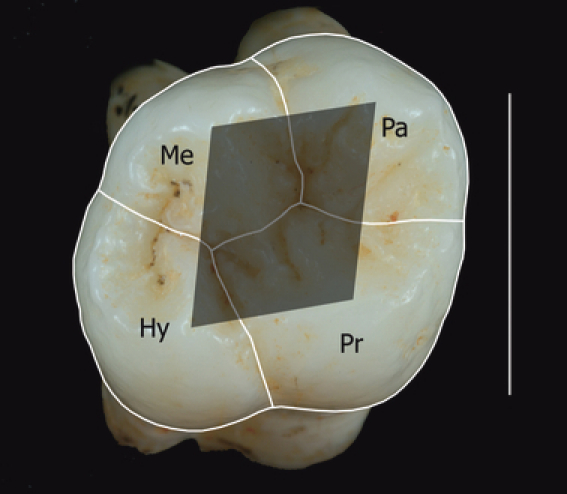
Right upper first molar ATD6-103 showing the separation of the different cusp base areas (Pr: protocone; Pa: paracone; Me: metacone; Hy: hypocone), the occlusal polygon (shaded polygon) and the angles located at each cusp (angles of the occlusal polygon). Scale bar: 1 cm. Buccal: top part of the image; lingual: bottom; mesial: right; distal: left.
Fig. 2.
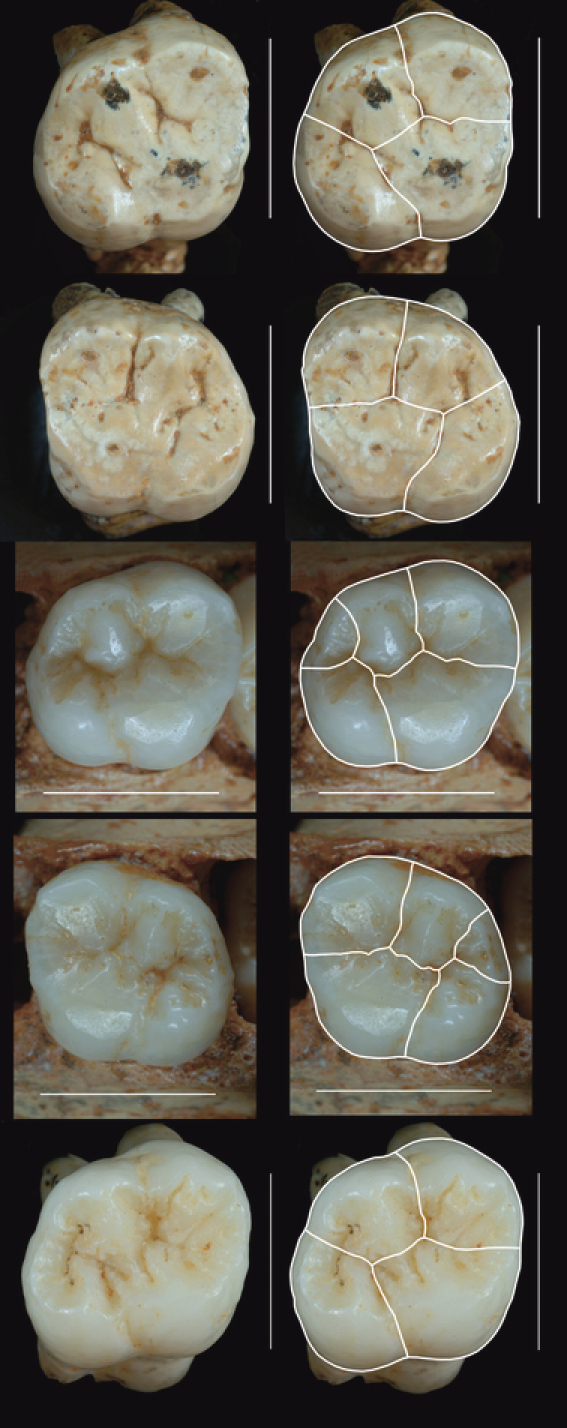
Homo antecessor upper first molars. Right half of the figure: photographs of the molars showing the cusp division used to obtain the measurements in Table 1. From top to bottom: ATD6-10: right antimere of individual H1 (incorrectly referred to as ATD6-18 in Table 1 of Gómez-Robles et al. 2007); ATD6-11: left antimere of individual H1; ATD6-69: right antimere of individual H3; ATD6-69: left antimere of individual H3; ATD6-103: right antimere (the only preserved one) of individual H5. Scale bars: 1 cm. Buccal: top; lingual: bottom. Right antimeres: mesial – right, distal – left. Left antimeres: mesial – left, distal – right. The small cusp located between the metacone and the hypocone in the molars of ATD6-69 is the metaconule, or c5.
Results
Absolute and relative cusp base areas of the five H. antecessor molars are provided in Table 1. Individual H5 (ATD6-103) has the largest total crown base area (TCBA), and individual H3 (ATD6-69) the smallest. The protocone is the largest cusp in the five measured molars, with an approximate size of 30% of the TCBA, but the size sequence of the other three cusps is not constant: the pattern of relative cusp size is protocone > hypocone > paracone > metacone in individuals H1 and H5, but protocone > paracone > hypocone > metacone in individual H3. In the five analysed molars, however, the metacone is the smallest cusp, with an approximate value of 21% of the TCBA, clearly smaller than the paracone.
Table 1.
Absolute and relative cusp base areas of the five Homo antecessor upper first molars.
| TCBA | Proto Abs | Para Abs | Meta Abs | Hypo Abs | Proto Rel (%) | Para Rel (%) | Meta Rel (%) | Hypo Rel (%) | |
|---|---|---|---|---|---|---|---|---|---|
| ATD6-10 (R) H1 | 127.6 | 37.3 | 29.5 | 28.1 | 32.7 | 29.2 | 23.1 | 22.0 | 25.6 |
| ATD6-11 (L) H1 | 125.8 | 38.2 | 29.3 | 26.7 | 31.6 | 30.4 | 23.3 | 21.2 | 25.1 |
| ATD6-69 (R) H3 | 117.4 | 36.8 | 28.9 | 23.7 | 28.0 | 31.3 | 24.6 | 20.2 | 23.9 |
| ATD6-69 (L) H3 | 112.6 | 34.1 | 27.6 | 24.1 | 26.8 | 30.3 | 24.5 | 21.4 | 23.8 |
| ATD6-103 (R) H5 | 135.6 | 41.2 | 31.5 | 29.0 | 33.9 | 30.4 | 23.2 | 21.4 | 25.0 |
| Total mean | 123.8 | 37.5 | 29.4 | 26.3 | 30.6 | 30.3 | 23.8 | 21.2 | 24.7 |
| Mean 1 antimere | 124.7 | 37.8 | 29.5 | 26.6 | 30.8 | 30.4 | 23.7 | 21.3 | 24.6 |
TCBA, total crown base area; Proto, protocone; Para, paracone; Meta, metacone; Hypo, hypocone; Abs, absolute area; Rel, relative area; R, right; L, left; Total mean: mean of the five molars; Mean 1 antimere: mean based on left antimeres of individuals H1 and H3 and on the only preserved antimere (right) of individual H5. Absolute areas are measured in mm2. Relative areas are represented as the percentage of the TCBA.
Cusp angles and the relative occlusal polygon area (OPA) of ATD6-103 are shown in Table 2 (see also Fig. 1). Angles at the protocone and at the metacone have very similar values, approximately 106–107°, whereas angles at the paracone and metacone are also roughly coincident (≈ 70°). The relative occlusal polygon area accounts for 25.3% of the TCBA in ATD6-103.
Table 2.
Cusp angles and relative occlusal polygon area measured in the ATD6-103 M1.
| ATD6-103 | |
|---|---|
| Angle protocone | 107.8° |
| Angle paracone | 74.5° |
| Angle metacone | 106.0° |
| Angle hypocone | 71.3° |
| Occlusal polygon area (OPA) | 34.3 mm2 |
| Total crown base area (TCBA) | 135.6 mm2 |
| Relative occlusal polygon area | 25.3% |
Relative occlusal polygon area = (OPA/TCBA)*100.
Discussion
The total crown base area of H. antecessor molars shows an intermediate value (TCBA = 123.8 mm2) between early Homo molars (TCBA = 134.9 mm2; Quam et al. 2009) and later Homo fossil molars (TCBA = 109.0 mm2; Quam et al. 2009). Nonetheless, the molars from TD6 show the cusp proportions described by Quam et al. (2009) as typical of later Homo taxa, consisting of a reduced metacone with respect to the paracone. These cusp proportions have been identified in later Homo species, including H. ergaster, H. erectus, H. heidelbergensis, H. neanderthalensis, and H. sapiens.
Furthermore, not only are the TD6 M1s characterized by the derived pattern of relative cusp proportions for the genus Homo, they also share some characters with modern humans and, especially, with Neanderthals (see Gómez-Robles et al. 2007). Like those from other European species, H. antecessor upper first molars are markedly skewed (Gómez-Robles et al. 2007) and have a large hypocone and a reduced metacone, the latter accounting for 21.2% of the TCBA (relative metacone size: H. ergaster: 26.0%; H. erectus: 22.9%; H. heidelbergensis: 20.1%; H. neanderthalensis: 20.6%; H. sapiens:≈ 22%; Quam et al. 2009). The pattern of relative cusp sizes typical of Neanderthals (protocone > paracone ≥ hypocone > metacone; Bailey, 2004) has been identified in one of the three H. antecessor individuals (H3) and in the mean value of the three individuals (protocone: 30.3% > paracone: 23.8%≥ hypocone: 24.7% > metacone: 21.2%; note that paracone and hypocone relative sizes are considered to be equivalent because ‘as a general rule, individual cusps that differ in their relative size by < 1% should be considered equal in size’; Quam et al. 2009). The two remaining individuals (H1 and H5) are characterized by a very large hypocone, even larger than the paracone, which gives rise to a different pattern of cusp sizes (protocone > hypocone > paracone > metacone) that has not been identified in any species included in Quam et al. (2009). These large and bulging hypocones (larger than those from any hominin species, including H. neanderthalensis and H. heidelbergensis), together with their reduced metacone, are responsible for the skewed outline and, hence, for the location in the morphospace (in the area occupied only by Neanderthals and pre-Neanderthals) of H. antecessor molars described in Gómez-Robles et al. (2007).
However, cusp angles in ATD6-103 are similar to those measured in different H. sapiens samples (approximately 70° at the paracone and hypocone, and 105° at the protocone and metacone). The discordant patterns between relative cusp size and cusp angles in H. antecessor are not surprising because these two factors have moderate or low correlations and ‘cusp size cannot be used to accurately predict cusp angle’ (Bailey, 2004). Finally, a marked reduction of the relative occlusal polygon area has been identified in ATD6-103 (the only upper first molar of H. antecessor where the morphology of the occlusal polygon has not been eroded by wear), which has a relative OPA of 25.3% (relative OPA: H. erectus: 30.8–35.0%; Early anatomically modern humans: 29.6–36.6%; Neanderthals: 24.5–30.5%; Upper Paleolithic modern humans: 31.8–36.8%; Contemporary modern humans: 27.0–50.4%; Bailey, 2004).
These results demonstrate that the morphology of H. antecessor upper first molars from TD6 does not represent an exception to the rule observed in the majority of Homo species. Rather, these molars show the derived cusp proportions found in all later taxa within the genus Homo, as well as some features identified in classic Neanderthals and European H. heidelbergensis. The evolutionary significance of this morphology should be carefully evaluated by considering together all the traits that H. antecessor shares with H. sapiens, with H. neanderthalensis and with both species, taking into account that isolated characters are unlikely to provide a complete scenario of the evolution of these species. For this reason, forthcoming papers will undertake the task of re-evaluating H. antecessor dental and skeletal characters with the goal of shedding light on the question of its phylogenetic position, which is still debated because of its complex mixture of primitive and derived characters (e.g. Bermúdez de Castro et al. 1997, 2003; Carbonell et al. 2005). New findings of Lower Pleistocene fossils coming from the different sites located at the Sierra de Atapuerca (Bermúdez de Castro et al. 2008, 2010a,b; Carbonell et al. 2008) will help to clarify some unresolved questions about this species.
Acknowledgments
The present manuscript has benefited from fruitful discussions with R. Quam and S. Bailey. The authors are deeply grateful to all the members of the Atapuerca Research Team, especially those who excavate or have excavated the TD6 level of the Gran Dolina site. Special thanks are due to Pilar Fernández and Elena Lacasa, from the Area of Conservation and Restoration of the CENIEH, for their assistance photographing the Gran Dolina fossils. This research was sponsored by the Ministerio de Ciencia e Innovación (Grant DGI CGL2006-1352-C03), Junta de Castilla y León (Grupo de Excelencia Project and field season), and Fundación Atapuerca (field season, staff support, and grants to A.G.-R. and L.P.-S.).
References
- Bailey SE. A morphometric analysis of maxillary molar crowns of middle-late Pleistocene hominins. J Hum Evol. 2004;47:183–198. doi: 10.1016/j.jhevol.2004.07.001. [DOI] [PubMed] [Google Scholar]
- Bailey SE, Pilbrow VC, Wood BA. Interobserver error involved in independent attempts to measure cusp base areas of Pan M1s. J Anat. 2004;205:323–331. doi: 10.1111/j.0021-8782.2004.00334.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermúdez de Castro JM, Arsuaga JL, Carbonell E, et al. A hominid from the lower pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans. Science. 1997;276:1392–1395. doi: 10.1126/science.276.5317.1392. [DOI] [PubMed] [Google Scholar]
- Bermúdez de Castro JM, Rosas A, Nicolás ME. Dental remains from Atapuerca-TD6 (Gran Dolina site, Burgos, Spain) J Hum Evol. 1999;37:523–566. doi: 10.1006/jhev.1999.0323. [DOI] [PubMed] [Google Scholar]
- Bermúdez de Castro JM, Martinón-Torres M, Sarmiento S, et al. Gran Dolina-TD6 versus Sima de los Huesos dental samples from Atapuerca: evidence of discontinuity in the European Pleistocene population? J Archaeol Sci. 2003;30:1421–1428. [Google Scholar]
- Bermúdez de Castro JM, Carbonell E, Gómez A, et al. Paleodemografía del hipodigma de fósiles de homininos del nivel TD6 de Gran Dolina (Sierra de Atapuerca, Burgos): estudio preliminar. Estud Geol. 2006;62:145. [Google Scholar]
- Bermúdez de Castro JM, Pérez-González A, Martinón-Torres M, et al. A new early Pleistocene hominin mandible from Atapuerca-TD6, Spain. J Hum Evol. 2008;55:729. doi: 10.1016/j.jhevol.2008.03.006. [DOI] [PubMed] [Google Scholar]
- Bermúdez de Castro JM, Martinón-Torres M, Prado L, et al. New immature hominin fossil from European Lower Pleistocene shows the earliest evidence of a modern human dental development pattern. Proc Natl Acad Sci U S A. 2010a;107:11739–11744. doi: 10.1073/pnas.1006772107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermúdez de Castro JM, Martinón-Torres M, Gómez-Robles A, et al. New human evidence of the Early Pleistocene settlement of Europe, from Sima del Elefante site (Sierra de Atapuerca, Burgos, Spain) Quat Inter. 2010b;223–224:431–433. [Google Scholar]
- Carbonell E, Bermúdez de Castro JM, Arsuaga JL, et al. An early Pleistocene hominin mandible from Atapuerca-TD6, Spain. Proc Natl Acad Sci U S A. 2005;102:5674–5678. doi: 10.1073/pnas.0501841102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carbonell E, Bermúdez de Castro JM, Parés JM, et al. The first hominin of Europe. Nature. 2008;452:465. doi: 10.1038/nature06815. [DOI] [PubMed] [Google Scholar]
- Gómez-Robles A, Martinón-Torres M, Bermúdez de Castro JM, et al. A geometric morphometric analysis of hominin upper first molar shape. J Hum Evol. 2007;55:627–638. doi: 10.1016/j.jhevol.2007.02.002. [DOI] [PubMed] [Google Scholar]
- Quam R, Bailey SE, Wood BA. Evolution of M1 crown size and cusp proportions in the genus Homo. J Anat. 2009;214:655. doi: 10.1111/j.1469-7580.2009.01064.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood BA, Engleman CA. Analysis of the dental morphology of Plio-Pleistocene hominids: V. Maxillary postcanine tooth morphology. J Anat. 1988;161:1. [PMC free article] [PubMed] [Google Scholar]