

Abstract
Here we introduce a new technique that probes voltage-dependent charge displacements of excitable membrane-bound proteins using extracellularly applied radio frequency (RF, 500 kHz) electric fields. Xenopus oocytes were used as a model cell for these experiments, and were injected with cRNA encoding Shaker B-IR (ShB-IR) K+ ion channels to express large densities of this protein in the oocyte membranes. Two-electrode voltage clamp (TEVC) was applied to command whole-cell membrane potential and to measure channel-dependent membrane currents. Simultaneously, RF electric fields were applied to perturb the membrane potential about the TEVC level and to measure voltage-dependent RF displacement currents. ShB-IR expressing oocytes showed significantly larger changes in RF displacement currents upon membrane depolarization than control oocytes. Voltage-dependent changes in RF displacement currents further increased in ShB-IR expressing oocytes after ∼120 µM Cu2+ addition to the external bath. Cu2+ is known to bind to the ShB-IR ion channel and inhibit Shaker K+ conductance, indicating that changes in the RF displacement current reported here were associated with RF vibration of the Cu2+-linked mobile domain of the ShB-IR protein. Results demonstrate the use of extracellular RF electrodes to interrogate voltage-dependent movement of charged mobile protein domains — capabilities that might enable detection of small changes in charge distribution associated with integral membrane protein conformation and/or drug–protein interactions.
Introduction
Techniques to monitor displacement currents in the protein-rich cell membrane have been used extensively in previous studies to examine ion-channel voltage-sensor movement [1], [2], [3], [4], [5], [6] and the piezoelectric-like behavior of transmembrane proteins [7], [8], [9]. These and similar studies record the magnitude and timing of charge displacement in the membrane to reveal the nature of voltage-dependent protein function and excitability in living cells. One technical challenge has been the electrical capacitance of the membrane lipid bilayer itself, which is in parallel with integral membrane proteins and obscures observations of protein-dependent electrical charge movements. This is compounded by the passive capacitance of the patch-clamp glass/quartz pipette that often limits the bandwidth of displacement current recording. In the present report, we introduce a new technique to monitor protein-dependent charge displacements by superimposing an extracellularly applied RF interrogation signal on top of a traditional voltage-clamp commanded membrane potential. This RF interrogation technique has its basis in electric impedance spectroscopy, which has been applied previously to probe membrane dielectric properties of isolated cells by measuring the electrical impedance between pairs or groups of extracellular electrodes [10], [11], [12], [13], [14], [15]. Here, we extend such RF dielectric measurements to study electrical charge displacement arising from electrically excited voltage-sensitive membrane-bound proteins.
Xenopus oocytes were used as a model cell for these experiments because they express a large amount of exogenous protein in their membrane; in these experiments, oocytes were transfected with the Shaker gene encoding ShB-IR [15], [16], [17], [18], [19], [20], a well-characterized voltage-sensitive K+ ion channel with fast inactivation removed. A custom recording chamber was devised to allow for simultaneous two-electrode voltage clamp (TEVC) and RF impedance interrogation (Fig. 1A). Oocytes were placed individually into the center of an annular electrode that formed a tight conducting extracellular belt around the meridian of an oocyte (Fig. 1A). RF signals were passed from the annular belt (black electrode in Fig. 1A) to a single ground-wire electrode located above the oocyte. Quadrature lock-in amplification was used to measure the frequency-domain RF component of the voltage drop (VRF), current (IRF), and impedance (ZRF = VRF/IRF) across the recording chamber. In this configuration, the RF component of the total current IRF was the sum of the conductive shunt current around the cell and the displacement current in the plasma membrane complex (Fig. 1B). Since the extracellular shunt conductance was much greater than the membrane conductance (Fig. 1B, 1 /Rs ≫Real(1/Zm)), RF impedance was not sensitive to voltage-dependent changes in membrane conductance and instead reflected voltage-dependent changes in membrane displacement currents [21]. Fig. 1C illustrates that a change in membrane displacement current would be detectable only over a limited frequency range due to the fact that shunt resistance (Rs) would dominate at low frequencies and membrane capacitance would effectively short at high frequencies. In fact, previous work has shown extracellular shunt path estimation (frequencies <100 kHz) can be used to map cell shape in a recording chamber, and that frequencies above 3 MHz may be used to interrogate intracellular organelle distribution [21]. Examination of the impedance spectra for the specific system used in the present study revealed 500 kHz to be near the center of the sensitive range to maximize detection of membrane-related RF charge displacements [22].
Figure 1. Set-up and circuit model.

A) Changes in RF membrane impedance (|ΔZRF|) during TEVC were measured by passing RF current from an electrode surrounding the meridian of the cell (black) to a ground electrode (media, above the cell). Contour lines and colors of the saggital cross-section of a cell in the recording chamber, shown here, illustrate the general spatial distribution of the RF electric potential expected based on the Maxwell equations for a passive cell under axisymmetric conditions (π/4 phase shown). B) A circuit model of the chamber including the shunt resistance (Rs,), membrane impedance (Zm), intracellular resistance (Ri), and electrode double layer (Zdl). C) Using the circuit model, the frequency-dependent RF impedance would change (Δ|ZRF|) with an increase or decrease in membrane capacitance — a change that would be most easily detectable at frequency ω* where the maxima of the |ΔZRF| occurs. The present study reports changes in RF impedance |ΔZRF| evoked by TEVC step changes in membrane potential.
Here we report changes in 500 kHz RF charge displacement evoked by step changes in membrane potential. TEVC was used to step the membrane potential (Vm*) in the time domain, and to measure whole-cell currents (Im*, Fig. 1A). RF interrogating signals (VRF, IRF) were superimposed on top of the low-frequency TEVC commands. TEVC Vm* commands were low-pass filtered at 30 kHz so the TEVC signal contributed negligible power at the 500 kHz RF interrogation frequency. Similarly, TEVC Im* signals were low-pass filtered to avoid the RF component. Lock-in amplification was used to extract electrical signals at the RF interrogation frequency. Data for ShB-IR expressing oocytes were compared to control oocytes to examine the contributions of the expressed ion channel relative to endogenous proteins and lipids constituting the native oocyte membrane (control). In a subset of experiments, Cu2+ was applied to shift the voltage-dependence of ShB-IR to more depolarized levels [23], [24] and to bind charge to the protein. Results show significant differences between voltage-dependent RF charge displacements in ShB-IR expressing oocytes vs. controls. Differences are significant near the half-activation potential for the ShB-IR channels, suggesting that RF data might correspond to ion-channel activity. Results further show that Cu2+ increased the magnitude of RF charge displacement while simultaneously shifting Vm* sensitivity to more depolarized levels on ShB-IR expressing oocytes. This response was not as noticeable in control oocytes, indicating that the RF signal measured reflects RF vibration of the channel-protein–Cu2+ interaction. These results suggest that the RF-based technique introduced here could potentially supplement conventional membrane-biophysics studies, by monitoring RF mobility associated with membrane-bound charge distribution and charged molecule–protein (potentially drug–protein) interactions during voltage-clamp.
Results
Time-resolved changes in RF Impedance
Fig. 2A compares time-resolved changes in RF impedance |ΔZRF| in control ooyctes expressing endogenous channels (avg. n = 10, left) to oocytes expressing ShB-IR channels (avg. n = 9, right). Voltage-dependent RF charge displacement occurred in both cell expression systems, but differed in magnitude and temporal waveform between the control and ShB-IR expressing oocytes. The left axis reports the magnitude of the change in impedance, |ΔZRF| = |ZRF- Z0|, where the reference, Z0, for each record was the average RF impedance 5–40 ms prior to the voltage step. Although signals were noisy in our apparatus, milliohm changes in |ΔZRF| during depolarization were readily discernable from noise. In all cases, |ΔZRF| consisted of an initial onset response when the rate of change of TEVC membrane potential was large (o, |ΔZRF|o, dVm*/dt>0) and a steady-state response when TEVC membrane potential was approximately constant (s, |ΔZRF|s, dVm*/dt ≅ 0). The RF onset response (o) occurred in the first millisecond after a voltage step was applied to the cell, and was similar in both the control and ShB-IR expressing oocytes. Interestingly, the RF onset response was rectified and did not occur at the end of the voltage command step. Hence, the fast RF onset response was not causally related to the standard TEVC capacitive transient (current spikes in Fig. 2B). Steady state RF changes (s) 5–35 ms after the voltage step were also observed in both control and ShB-IR oocytes, but were significantly larger in the ShB-IR expressing oocytes at membrane potentials above −20 mV. This indicated a difference in RF response attributable to Shaker activation. Simultaneous TEVC recordings shown in the lower panels (Fig. 2B, current; Fig. 2C, voltage) were used to confirm expression and test whole-cell currents. Darker lines indicate increased levels of depolarization during the voltage step, commanded from a holding potential of −90 mV (Fig. 2C). TEVC whole-cell currents in the control condition (Fig. 2B, left) were small relative to the large currents in ShB-IR expressing cells (Fig. 2B, right). Voltage sensitivity and sustained ionic currents reported here are typical for ShB-IR cells with fast inactivation removed, and indicate successful protein expression and activity.
Figure 2. Temporally Resolved RF Measurements.

A) RF impedance changes (|ΔZRF|) measured during TEVC relative to the impedance at holding potential (−90 mV) in control oocytes, expressing endogenous proteins only (n = 10, left column), and ShB-IR expressing oocytes (n = 9, right column). ShB-IR expressing oocytes elicited a membrane-potential-dependent (Vm *) RF response different than control oocytes. RF impedance changes were analyzed in two regions; the RF response during the onset of voltage-step (o, average |ΔZRF|o 0–1 ms after voltage step, dVm*/dt > 0) and the RF response after membrane potential achieved its command (steady-state) level (s, average |ΔZRF|s 5–35 ms after voltage step, dVm*/dt ≅ 0). B) TEVC current measurements were used to verify ion-channel expression and responses (leak current subtracted, capacitive transient unsubtracted) to C) whole-cell voltage-clamp.
Steady-state RF response for ShB-IR and Control Cells
To further examine differences between steady-state RF impedance in control (n = 10, “Endo”, orange) vs. ShB-IR expressing oocytes (n = 9, “ShB-IR + Endo”, brown), we averaged |ΔZRF|s 5–35 ms (see Methods) after the onset of voltage step (Methods), and plotted the result against the average membrane potential Vm* (Fig. 3A). ShB-IR expressing oocytes include the exogenously expressed ShB-IR proteins and endogenous membrane proteins and both contribute to the RF data. Averaged control data were subtracted from the ShB-IR expressing oocyte data, to estimate the RF response associated with the ShB-IR protein only (Fig. 3A, “ShB-IR only”, blue line). Error bars denote standard errors of the mean (+/−, SEM), and are shown as a function of Vm* for ShB-IR expressing oocytes and control oocytes in Fig. 3B. To demonstrate successful transfection, steady-state ionic currents are shown in Fig. 3C, and ShB-IR channel conductance is shown in the inset. Like the averaged RF data, the SEM associated with the ShB-IR protein only (Fig. 3B, “ShB-IR only”, blue line) was estimated by subtracting the SEM associated with the ShB-IR expressing oocytes at each membrane potential from the control oocytes SEM(s) (see Methods). Significant differences in |ΔZRF|s were found between Shaker-expressing oocytes and control oocytes at and above −10 mV (Fig. 3A), and differences are easily observable above the half-activation voltage for ShB-IR channels (x p<0.10, *p<0.05, Fig. 3A). Interestingly, the RF ShB-IR-only data (Fig. 3A, blue line) and the corresponding ShB-IR conductance data (shown in Fig. 3C) have a linear correlation coefficient of .94 (and a correlation coefficient of .99 in the voltage range of −45 mV to 2 mV, when the ShB-IR data increase from 5–95% of their final value), indicating a positive association between channel activity and the measured RF response. Furthermore, unlike the control oocytes that showed increased SEM for large Vm*, the ShB-IR-only SEM (Fig. 3B, blue line) showed the largest value near the half-activation voltage, and standard errors for the ShB-IR expressing oocytes were comparable to baseline values when the ensemble of channels in the membrane were predominantly closed or open. These results are consistent with voltage-dependent SEM arising from probabilistic conformational state of the Shaker ion-channels expressed in the membrane.
Figure 3. Steady-state RF response for ShB-IR and Control Cells.

A) Significant (x p = .1, *p = .05) voltage-dependent differences in |ΔZRF| s were observed between control oocytes, expressing endogenous proteins only (“Endo”, orange), and ShB-IR expressing oocytes (express both endogenous and ShB-IR proteins, “ShB-IR + Endo”, brown). The “Endo” response was subtracted from the “ShB-IR + Endo” response to estimate the isolated RF response from the ShB-IR channels only (“ShB-IR only”, blue). Error bars denote +/− standard errors of the mean (SEM). B) The SEM for the isolated ShB-IR proteins (“ShB-IR only”, blue) was also estimated by subtracting the SEM from the control oocytes (“Endo”, orange) from the SEM associated with the ShB-IR expressing oocytes (“ShB-IR + Endo”, brown). The SEM for isolated ShB-IR expressing oocytes was largest near the half-activation potential for these ion channels. C) ShB-IR channel expression and voltage-dependent whole-cell current was verified using TEVC, and this data was used to estimate ShB-IR conductance (G/Gmax, inset).
Copper treatment and steady-state ShB-IR RF response
A subset of ShB-IR expressing oocytes (n = 4) were treated with Cu2+, a K+ ion-channel modulator, and normalized |ΔZRF|s were determined for these oocytes before and after Cu2+ treatment (exp2). Results are shown in Fig. 4A as a function of steady-state membrane potential (Vm*). Data from each cell were normalized to its RF impedance at +30 mV for the nontreated ShB-IR expressing oocytes, to permit comparisons between cells before/after copper treatment (see Methods). As expected, Cu2+ application greatly reduced the ShB-IR current (Im*) in the voltage-range where ShB-IR channels activate (Fig. 4B). ShB-IR channel conductance before and after copper addition are shown in the inset (Fig. 4B). The change in RF impedance after copper treatment was larger than the untreated condition (Fig. 4A). To examine statistical significance for this relatively small population, data above −60 mV (see Methods) were pooled together. Pooled data showed a statistically significant difference between nontreated vs. Cu2+-treated ShB-IR expressing oocytes (p = .04, U = 1007, Total Points = 80, normalized median-values of the nontreated/treated ShB-IR expressing oocytes are .36 and .90, respectively; see Methods). Results from control cells (n = 2, Fig. 4A) are scaled by the ratio of the control cell to ShB-IR cell data at +30 mV (shown in Fig. 3) to enable inter-cellular comparison, after normalizing the data from each control cell to its nontreated |ΔZRF|s measured at +30 mV (see Methods). A change in the normalized |ΔZRF|s is noticeable in the control cells after Cu2+ application (above −60 mV), indicating that Cu2+ might non-specifically interact with the endogeneous oocyte membrane in addition to the known effect of binding the ShB-IR channels. While non-specific Cu2+ likely contributed to the change in RF response before/after Cu2+ addition, the effect in ShB-IR expressing oocytes was much larger, suggesting that the RF also detected Cu2+ interaction specifically with the Shaker channels. As depolarization level increased, the effect of the bound Cu2+ increased. At 30 mV, the Cu2+-treated ShB-IR expressing oocytes exhibited a response approximately 1.5 times that of the non-treated ShB-IR expressing oocytes, even though the ionic current (TEVC) was approximately one fifth of the non-treated cells.
Figure 4. Copper treatment and steady-state ShB-IR RF response.

A) Voltage-dependent differences in |ΔZRF|s were observed between Shaker expressing oocytes (“ShB-IR + Endo”, green line) and the same cells exposed to ∼120 µM Cu2+ (purple line). A similar effect was apparent, albeit to a lesser extent, for control cells before (“Endo”, green markers)/after Cu2+ treatment (purple markers). Error bars denote +/− standard errors of the mean (SEM). B) Even though RF charge displacements increased in Cu2+-exposed ShB-IR expressing oocytes, TEVC whole-cell current decreased showing that Cu2+ successfully blocked the channels (channel conductance shown as inset).
RF Onset Response in ShB-IR Expressing Oocytes
RF onset responses, occurring during the rise of Vm* (when dVm*/dt was maximum), exhibited trends different than those seen in steady state. Fig. 5 plots the onset response |ΔZRF|o, averaged 0–1 ms after voltage command was applied (see Methods) vs. the rate of change of membrane potential (dVm*/dt) and membrane potential (parenthetically noted below the values of dVm*/dt). Cell data have been normalized using the same values in Fig. 4A, to allow for comparisons between transient and steady-state RF results (see Methods). The fast onset recorded in ShB-IR expressing oocytes and controls both increased with the magnitude of the membrane potential change, but there were no statistically significant differences due to expression of K+ channels (p>.1, data not shown). Onset RF charge displacements in control oocytes treated with Cu2+ slightly decreased as voltage-step increased (Fig. 5, filled circles), and in contrast to ShB-IR expressing oocytes, the average onset response of controls treated with Cu2+ was smaller than the non-treated cells. Cu2+-treated ShB-IR expressing oocytes pooled for all voltage commands > −60 mV exhibited significantly larger onset responses than non-treated ShB-IR expressing oocytes (p = .005, U = 1087, Total Points = 80, median values untreated/treated ShB-IR expressing oocytes of 1.0/1.4, respectively). Interestingly, increases in the average onset |ΔZRF|o for Cu2+ treated ShB-IR expressing oocytes were about 1.5 times greater than those for Cu2+ treated ShB-IR oocytes in the steady state (|ΔZRF|s) for membrane potentials >−60 mV. One hypothesis that might explain the Cu2+-dependent onset response is that Cu2+ might have stabilized a transitional state in the ion channel during rapid membrane depolarization that momentarily affected the charge distribution on the protein, thereby enhancing the momentary RF charge displacement. The onset response was not detectable for -dVm*/dt at the end of the command pulse (see Fig. 2A), showing a rectification in |ΔZRF| quite distinct from dVm*/dt driven linear capacitive transients (present in Fig. 2B).
Figure 5. Onset RF Response in ShB-IR Expressing Oocytes.

Changes in RF impedance during the onset of voltage-clamp (|ΔZRF|o, 0–1 ms after whole-cell depolarization) were slightly depressed in control oocytes with the addition of Cu2+ (Cu2+-free - green markers, Cu2+ addition - purple markers), but were significantly greater in ShB-IR expressing oocytes (Cu2+-free - green line, Cu2+ addition - purple line).
Discussion
Here we introduce a new technique to monitor voltage-dependent charge displacement associated with ShB-IR K+ ion channels using RF electric fields during TEVC. The RF electric field provided a vibrational force that resulted in measurable RF charge displacement on a cell membrane complex caused by vibrating membrane-bound charges. The 500 kHz RF was superimposed on top of low-frequency TEVC (<30 kHz) allowing the RF to continuously interrogate changes in charge displacement mobility during whole-cell depolarization. Changes in RF impedance, |ΔZRF|, were recorded every 20 µs, and provided a nearly instantaneous view of the effective dielectric behavior of the protein–membrane–ion complex. Voltage-dependent changes in |ΔZRF| were averaged into millisecond bins (Fig. 2) and showed time-resolved differences for ShB-IR expressing oocytes relative to controls, demonstrating the sensitivity of this approach to detect RF vibrational motion associated with ion-channel-specific charge displacements.
Analysis of RF charge displacements revealed significant differences in |ΔZRF|s between ShB-IR expressing oocytes and controls at depolarized membrane potentials (Fig. 3A). Differences were largest above the half-activation potential for the Shaker channels used here. The ShB-IR channels have fast inactivation removed, and as such, significant changes in steady-state RF vibration demonstrate that the open conformation of ShB-IR channels have measurably different effective dielectric behavior than the closed conformation. It is important to note that this difference was present at steady-state and did not reflect the classical gating-charge movement that typically occurs for these channels ∼0–2 ms after voltage command [4], [25]. Instead, voltage-dependent changes in RF charge displacement |ΔZRF|s appear to reflect changes in mobile charge displacement magnitude within the transmembrane RF electric field and/or stiffness associated with the open vs. closed state of the Shaker protein (measured during steady-state). Consistent with this, the steady-state RF impedance associated with the ShB-IR proteins had a positive correlation with channel conductance (cf. blue curve in Fig. 3A and 3C inset). This hypothesis is further supported by the fact that the SEM for the ShB-IR proteins (ShB-IR expressing oocytes – control oocytes) was greatest at the channel's half-activation potential, and smaller when the protein was completely hyperpolarized (all channels closed) or depolarized (all channels open). This same trend in SEM was not observed in control oocytes, and can only be explained if the observed changes in effective dielectric were due to charge vibrations specific to the Shaker protein.
Cu2+ addition to the bath significantly changed steady-state RF impedance |ΔZRF|s in ShB-IR expressing oocytes relative to controls. In particular, data indicated that while a portion of the steady-state RF response may be caused by non-specific binding of Cu2+ to the cell membrane, Cu2+ also binds specifically to the ShB-IR protein and increases RF –evoked charge displacement during membrane depolarization. Analysis of the ShB-IR expressing oocyte data showed that differences between Cu2+-treated and non-treated cells were present at membrane potentials as low as −50 mV and increased with TEVC depolarization (Fig. 4A). The increase in RF response caused by Cu2+–Shaker interaction occured even as ion-channel conduction current decreased (Fig. 4B). One possibility is that Cu2+ binding to the ion channel enhanced |ΔZRF|s by increasing the effective charge moving within the transmembrane electric field and this thereby enhanced the signal arising from the RF-evoked protein vibration.
In addition to steady-state changes in RF charge displacement, rapid onset responses |ΔZRF|o were detected during positive depolarizing steps (dVm/dt>0) in both ShB-IR expressing and control oocytes (see Fig. 2). Interestingly, the onset response was large and easily observable in both cell expression systems (control and ShB-IR) only for positive dVm/dt. Hence, the onset RF data are not directly related to the traditional capacitive transient current evoked during voltage clamp. The present data were not adequate to analyze the decay time constant of the RF onset response in the ShB-IR expressing cells due to the superimposition of the large RF response associated with the ShB-IR channel activation. The magnitude of the transient response in both control and ShB-IR expressing oocytes was comparable, indicating the importance of endogenous mechanisms contributing to the RF onset response (data not shown here). This shows that the RF onset displacement currents measured here are not caused by the gating charge movement of Shaker channels. This was not surprising, due to the slow voltage clamp rise time in the present experiments. A faster voltage-clamp and RF recording system might allow future investigation of voltage-sensor displacement via RF interrogation. Even with the slow voltage-clamp used here, the onset response |ΔZRF|o increased significantly in ShB-IR expressing oocytes treated with Cu2+ (Fig. 5). Data for Shaker-expressing cells in Fig. 5 were normalized to the same values as the steady-state figure, and show that |ΔZRF|o in the Cu2+-treated cells was 1.5 times as high as the values shown for the same copper-treated cells in the steady-state. One possible explanation for the large value of |ΔZRF|o could be that the membrane-ion complex undergoes a non-equilibrium polarization when dVm/dt is positive, compelling a brief period of enhanced dipole mobility. If true, this effect may be amplified or augmented by Cu2+–membrane–ShB-IR interactions.
Although interpretation of the RF results can be challenging on the surface, this type of data cannot be obtained by traditional electrophysiological techniques and may prove useful in understanding membrane-bound protein dynamics. Present results show that the method can be used to track time- and voltage-dependent changes in RF charge displacement associated with membrane-bound ion-channel state/conformation and electrostatic binding of charged compounds. As such, the technique supplements conventional electrophysiological techniques and is suitable to examine voltage-dependent membrane-bound protein conformations and potentially also pharmacological interactions.
Methods
Oocyte Injection
Xenopus oocytes were isolated at the University of Utah's Cardiovascular Research and Training Institute. cRNA encoding ShB-IR was injected into oocytes 1–2 days before experimentation, to allow ample time for channel expression. Oocytes were stored in Superbarths oocyte media (88 mM NaCl, 1 mM KCl, 0.41 mM CaCl2, 0.33 mM Ca(NO3)2, 1 mM MgSO4, 2.4 mM NaHCO3, 10 mM HEPES, 1 mM pyruvate, and 50 µg/ml gentamicin, titrated using NaOH to a pH of 7.4) at 17° Celsius and ionic currents measured using two-electrode voltage clamp (TEVC) were used to verify channel expression.
Experimental Set-Up
An oocyte (∼1.2 mm diameter) was positioned in a 1.2 mm recording chamber filled with Super Barth's Oocyte media surrounded by a 18 µm thick, gold-plated copper, effectively annular electrode (ORFLO, Woodinville, WA). Oocyte positioning and electrode stabilization were facilitated by a polycarbonate interface; the electrode was stabilized by two separate plates of the polycarbonate interface, and an oocyte was loaded into the recording chamber through a bowl-shaped well [22]. Two glass micropipettes, used for two-electrode voltage clamp and filled with 3M KCl, were carefully placed in the oocyte using a 2X magnification lens on an upright microscope (AxioTech, Zeiss, Thornwood, NY). Each micropipette had a 0.5–2 MΩ access resistance, and was driven by a two-electrode amplifier (AxoClamp2B, Molecular Devices, Sunnyvale, CA) that controlled voltage-measuring (HS-2Ax1LU, Molecular Devices, Sunnyvale, CA) and current-injecting headstages (HS-2Ax10MGU, Molecular Devices, Sunnyvale, CA). An Ag/AgCl ground electrode was placed approximately 5 mm above the oocyte and immersed in media near the top of the recording chamber. This ground was common for both the RF recordings and TEVC. Photographs taken of the oocyte before and after experimentation were used to confirm minimal oocyte movement during recording (Q-Color3, Olympus, Center Valley, PA).
A∼120 µM Cu2+ solution (Copper ICP/DCP standard solution 10,000 µg/mL Cu2+in 2% HNO3, Sigma Aldrich, St. Louis, MO) was applied as an ion channel modulator to a small subset of the oocytes involved in experimentation. The Cu2+ solution was manually pipetted directly above the cell. The concentration of Cu2+ added was approximate, as it depended on the volume of Superbarths oocyte media in the recording chamber (∼1.25 mL).
Recording
A 500 kHz radio frequency was passed from the gold-plated annular electrode to the ground electrode [22]. Simultaneously, TEVC was used to control membrane potential (Vm*, single-pole low pass filter, 30 kHz). Oocytes were held at −90 mV (holding potential) for 360 ms, and a voltage step was then applied for 40 ms. Nine different voltage steps were applied to the oocyte (−120 mV, −60 mV, −40 mV, −30 mV, −20 mV, −10 mV, 0 mV, 10 mV, 40 mV), and are collectively referred to as a voltage train. Voltage trains were applied to each cell 50 times. Voltage-clamp was automated using a software package designed for this purpose (Patchmaster, HEKA Inst., Bellmore, NY; Igor Pro, Wave Metrics, OR).
RF voltage drop across the recording chamber (VRF) and RF current (IRF) were monitored using an onboard reference impedance to track excitable membrane impedance changes (ZRF = VRF/IRF). Head-stage amplification (OPA356, Texas Instruments, Dallas, TX), a 100 kHz high pass filter (48 dB/Oct Bessel HPF; SIM965, Stanford Research Systems, Sunnyvale, CA) and lock-in amplification (SR844, Stanford Research Systems, Sunnyvale, CA) were used to extract the RF component applied during TEVC. Voltage-clamp current and commands were low-pass filtered at <30 kHz to further ensure minimal cross-talk between the TEVC and RF recording systems. The quadrature lock-in signals, as well as TEVC ionic current, membrane potential and applied command potential were recorded through a 16-bit A/D converter (ITC-1600, HEKA Inst. Bellmore, NY). Patch-clamp software (Patchmaster, HEKA Inst., Bellmore, NY) sampled and saved data from each A/D channel at a rate of 20 µs. This voltage-clamp software in conjunction with the signal averaging time on the lock-in were the rate limiting factors for temporal resolution during RF interrogation. The technique, however, given the Nyquist frequency of the RF, has the potential to resolve events down to 4 µs.
Data Analysis
Data from individual oocytes were averaged over 50 command presentations (each consisting of 9 voltage levels, as described above). TEVC currents were leak compensated for each cell by assuming that currents measured during hyperpolarization were predominately caused by leak, and that these currents changed linearly with voltage. Leak-subtracted TEVC data for each oocyte were binned in 1 ms intervals in the time domain, and used to estimate ShB-IR channel conductances (Ohm's law: ITEVC
* = GSH
B-IR (Vm
*-Erev)). Capacitive transients were not subtracted. TEVC currents were plotted as a function of membrane potential, and current data (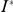 ) collected at Vm
* > −10 mV were linearly interpolated to 0 to estimate Erev. Error between I and (V- Erev) was then minimized by providing the best fits to a sigmoidal conductance (
) collected at Vm
* > −10 mV were linearly interpolated to 0 to estimate Erev. Error between I and (V- Erev) was then minimized by providing the best fits to a sigmoidal conductance (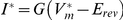 , where
, where  ;
; 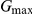 is the maximum whole-cell conductance of ShB-IR expressing oocytes; VHA is the half activation potential; λ is the rate of conductance rise). Channel conductance values were normalized to the maximum conductance value (G/Gmax), and are shown as insets in Fig. 3 and 4.
is the maximum whole-cell conductance of ShB-IR expressing oocytes; VHA is the half activation potential; λ is the rate of conductance rise). Channel conductance values were normalized to the maximum conductance value (G/Gmax), and are shown as insets in Fig. 3 and 4.
Magnitude change in RF impedance (|ΔZRF|, 500 kHz interrogation frequency) for each cell was calculated relative to the average RF impedance 5–35 ms before voltage command (holding potential ∼ = −90 mV). This was done in one millisecond bins. Temporally based comparisons of control vs. ShB-IR cells and Cu2+-treated vs. non-treated oocytes (like in Fig. 2) were made by averaging data (TEVC current, TEVC voltage, |ΔZRF|) across control oocytes (n = 10), ShB-IR expressing oocytes (consist of both endogenous and ShB-IR protein, n = 9), ShB-IR expressing oocytes before/after Cu2+ treatment (n = 4) and control oocytes before/after Cu2+ treatment (n = 2). Voltage data were used to align voltage steps from individual cells in time to minimize jitter during averaging.
To facilitate comparisons across cells, the average value of |ΔZRF| was calculated for each cell at every voltage-step 1) 5–35 ms after the voltage step had been applied (|ΔZRF|s, dVm*/dt (voltage clamp) ≅ 0, “steady state response”) and 2) 0–1 ms after the voltage step had been applied (|ΔZRF|o, dVm*/dt (voltage clamp) ≠ 0, “onset response”). To compensate for slight differences in TEVC membrane potential between individual cells, normalized RF data were linearly interpolated between voltages for each cell before averaging. Steady-state data were interpolated at the following membrane potential levels {−110 mV, −100 mV, −90 mV, −80 mV, −70 mV, −60 mV, −50 mV, −40 mV, −30 mV, −20 mV, −10 mV, 0 mV, 10 mV, 20 mV, 30 mV}. Onset response was plotted as a function of dVm */dt, and results were interpolated to the following values {−12 mV/ms, −8 mV/ms, −4 mV/ms, 0 mV/ms, 4 mV/ms, 8 mV/ms, 12 mV/ms, 16 mV/ms, 20 mV/ms, 24 mV/ms, 28 mV/ms, 32 mV/ms, 36 mV/ms, 40 mV/ms, 44 mV/ms}.
The sample sizes used in the ShB-IR cell (n = 9) vs. control cell (n = 10) experiment (exp1) allowed for direct (non-normalized) comparison between both data sets, as it was assumed that the approximate shunt path associated with the ShB-IR expressing oocytes and control oocytes was approximately equal across the populations (Fig. 3). Standard errors of the mean were calculated for both control and ShB-IR expressing oocytes at specified membrane potentials (−110 mV, −100 mV, −90 mV, −80 mV, −70 mV, −60 mV, −50 mV, −40 mV, −30 mV, −20 mV, −10 mV, 0 mV, 10 mV, 20 mV, 30 mV). The ShB-IR expressing oocytes consisted of RF responses from both endogenous proteins on the oocyte membrane and the exogenously expressed ShB-IR protein (referred to as “ShB-IR + Endo” in Fig. 3). The ShB-IR and endogenous protein responses were assumed to be approximately independent, and the averaged control data (endogenous protein only, “Endo”) was subtracted from the averaged ShB-IR expressing oocyte data (“ShB-IR + Endo”, Fig. 3) to estimate the RF response associated with the ShB-IR-protein only (“ShB-IR only”, blue line, Fig. 3). Similarly, the standard error of the mean associated with the ShB-IR protein-only was calculated by subtracting the SEM associated with control oocytes from the SEM associated with the ShB-IR expressing oocytes (blue line, Fig. 3b).
The effects of Cu2+ application on both steady-state and transient RF impedance were compared for ShB-IR expressing oocytes (n = 4) and control oocytes (n = 2, exp2). The small sample size prevented voltage-dependent population statistics for these controls, but we were able to make comparisons within cells. Data for each cell before/after copper treatment were normalized to the steady-state RF impedance value of that specific cell before Cu2+ treatment at +30 mV. After normalization, data for each group (Control-untreated/treated, ShB-IR-untreated/treated) were averaged together, and the effects of Cu2+ before/after treatment were compared within each cell expression system. Control data (both before/after copper treatment) were scaled by the ratio between the control cells and ShB-IR cells at +30 mV from the previous experiment (larger sample sizes, Fig. 3). This scaling enabled comparisons between the control cell data and the ShB-IR cell data before/after copper treatment. Standard errors of the mean were calculated for ShB-IR expressing oocytes at specified membrane potentials (−110 mV, −100 mV, −90 mV, −80 mV, −70 mV, −60 mV, −50 mV, −40 mV, −30 mV, −20 mV, −10 mV, 0 mV, 10 mV, 20 mV, 30 mV). SEM was not calculated for control cells, due to the small (<3) sample size.
In exp1, data at each voltage for both the control and ShB-IR expressing cells were not normally distributed, due to differences in shunt path between cells and differences in exogenous protein expression level on the ShB-IR expressing oocytes. As such, a Mann-Whitney nonparametric statistical test was used to compare control vs. ShB-IR expressing oocyte data (exp1) at each of the sampled voltage levels (the median of the sample values were slight lower than the averages shown in Fig. 3A, although the trends were similar). The null hypothesis (H0) for this comparison was that the RF data for the ShB-IR cells before are the same as the RF data for control oocytes at a specified voltage (Vm *, membrane potential) level. The linear correlation coefficient between ShB-IR conductance and the subtracted ShB-IR-only RF response was calculated both between -110 mV and 30 mV and between −45 and 2 mV (the regime where the RF ShB-IR-only response increased from 5–95% of its final value).
In exp2, the sample size of the ShB-IR expressing oocytes (n = 4) before/after Cu2+ treatment was too small to apply the Mann-Whitney test at every voltage level (requires a total sample size of 10). Instead, all the normalized data sampled from cells above −60 mV (where the channel begins to activate) were pooled, and these pooled samples were compared before/after copper treatment using the Mann-Whitney test (the pooled voltage levels for each group of cells was the same). This allowed for a significance test between the two treatment groups of cells, with the null hypothesis (H0) that the RF data for the ShB-IR cells before copper treatment are the same as the RF data for the ShB-IR expressing oocytes after copper treatment.
Acknowledgments
M. Sanguinetti (CVRTI, Univ. Utah) provided Xenopus oocytes. C. King and H.E. Ayliffe (ORFLO Technologies) assisted in the fabrication of the electrodes used in RF recording.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This work was supported by National Institutes of Health grant numbers R01 DC004928 and R01 DC006685, and National Science Foundation grant number IGERT DGE-9987616. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Armstrong C, Bezanilla F. Inactivation of the sodium channel II: gating current experiments. The Journal of General Physiology. 1977;70:567–590. doi: 10.1085/jgp.70.5.567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cha A, Bezanilla F. Characterizing voltage-dependent conformational changes in the Shaker K+ channel with fluorescence. Neuron. 1997;19:1127–1140. doi: 10.1016/s0896-6273(00)80403-1. [DOI] [PubMed] [Google Scholar]
- 3.Glauner K, Mannuzzu L, Gandhi C, Isacoff E. Spectroscopic mapping of voltage sensor movement in the Shaker potassium channel. Nature. 1999;402:813–817. doi: 10.1038/45561. [DOI] [PubMed] [Google Scholar]
- 4.Perozo E, MacKinnon R, Bezanilla F, Stefani E. Gating currents from a nonconducting mutant reveal open-closed conformations in Shaker K+ channels. Neuron. 1993;11:353–358. doi: 10.1016/0896-6273(93)90190-3. [DOI] [PubMed] [Google Scholar]
- 5.Sigg D, Bezanilla F. Total charge movement per channel: the relation between gating charge displacement and the voltage sensitivity of activation. The Journal of General Physiology. 1997;109:27–39. doi: 10.1085/jgp.109.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sigg D, Stefani E, Bezanilla F. Gating current noise produced by elementry transitions in Shaker potassium channels. Science. 1994;264:578–582. doi: 10.1126/science.8160016. [DOI] [PubMed] [Google Scholar]
- 7.Santos-Sacchi J, Navarette E. Voltage-dependant changes in specific membrane capacitance caused by prestin, the outer hair cell lateral membrane motor. Pflugers Archiv – European Journal of Physiology. 2002;444:99–106. doi: 10.1007/s00424-002-0804-2. [DOI] [PubMed] [Google Scholar]
- 8.Rabbitt R, Ayliffe H, Christensen D, Pamarthy K, Durney C, et al. Evidence of piezoelectric resonance in isolated outer hair cells. Biophysical Journal. 2005;88:2257–2265. doi: 10.1529/biophysj.104.050872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zheng J, Shen W, He D, Long K, Madison L, et al. Prestin is the motor protein of cochlear outer hair cells. Nature. 2000;405:149–155. doi: 10.1038/35012009. [DOI] [PubMed] [Google Scholar]
- 10.Ayliffe H, Frazier B, Rabbitt R. Electric impedance spectroscopy using microchannels with integrated metal electrodes. IEEE Journal of Microelectromechanical Systems. 1999;8:50–57. [Google Scholar]
- 11.Cheung K, Gawad S, Renaud P. Impedance spectroscopy flow cytometry: on-chip label-free cell differentiation. Cytometry Part A. 2005;65A:124–132. doi: 10.1002/cyto.a.20141. [DOI] [PubMed] [Google Scholar]
- 12.Gawad S, Schild L, Renaud P. Micromachined impedance spectroscopy flow cytometer for cell analysis and particle sizing. Lab on a Chip. 2001;1:76–82. doi: 10.1039/b103933b. [DOI] [PubMed] [Google Scholar]
- 13.Gritsch S, Nollert P, Jahnig F, Sackmann E. Impedance spectroscopy of porin and gramicidin pores reconstituted into supported lipid bilayers on indium-tin-oxide electrodes. Langmuir. 1998;14:3118–3125. [Google Scholar]
- 14.Han A, Frazier AB. Ion channel characterization using single cell impedance spectroscopy. Lab on a Chip. 2006;6:1412–1414. doi: 10.1039/b608930e. [DOI] [PubMed] [Google Scholar]
- 15.Werdich A, Lima E, Ivanov B, Ges I, Anderson M, et al. A microfluidic device to confine a single cardiac myocyte in a sub-nanoliter volume on planar microelectrodes for extracellular potential recordings. Lab on a Chip. 2004;4:357–362. doi: 10.1039/b315648f. [DOI] [PubMed] [Google Scholar]
- 16.Dascal N. The use of Xenopus oocytes for the study of ion channels. CRC Critical Reviews in Biochemistry. 1987;22:317–387. doi: 10.3109/10409238709086960. [DOI] [PubMed] [Google Scholar]
- 17.Hoshi T, Zagotta W, Aldrich R. Biophysical and molecular mechanisms of Shaker potassium channel inactivation. Science. 1990;250:533–538. doi: 10.1126/science.2122519. [DOI] [PubMed] [Google Scholar]
- 18.Hoshi T, Zagotta W, Aldrich R. Two types of inactivation in Shaker K+ channels: effects of alterations in the carboxy-terminal region. Neuron. 1991;7:547–556. doi: 10.1016/0896-6273(91)90367-9. [DOI] [PubMed] [Google Scholar]
- 19.Iverson L, Rudy B. The role of the divergent amino and carboxyl domains on the inactivation properties of potassium channels derived from the Shaker gene of Drosophila. The Journal of Neuroscience. 1990;10:2903–2916. doi: 10.1523/JNEUROSCI.10-09-02903.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stuhmer W. Electrophysiological recording from Xenopus oocytes. Methods in Enzymology. 1992;207:319–339. doi: 10.1016/0076-6879(92)07021-f. [DOI] [PubMed] [Google Scholar]
- 21.Dharia S, Ayliffe HE, Rabbitt RD. Single cell electric impedance topography: Mapping membrane capacitance. Lab on a Chip. 2009;9:3370–3377. doi: 10.1039/b912881f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dharia S, Richard RD. Monitoring voltage-sensitive membrane impedance change using radio frequency interrogation. 2010:889–894. doi: 10.1109/IEMBS.2010.5627860. In Proceedings of the Annual International Conference of the IEEE-EMBS, Buenos Aires, Argentina, [DOI] [PubMed] [Google Scholar]
- 23.Elinder F, Arhem P. Metal ion effects on ion channel gating. Quarterly Reviews of Biophysics. 2004;36:373–427. doi: 10.1017/s0033583504003932. [DOI] [PubMed] [Google Scholar]
- 24.Ma Z, Wong KY, Horrigan FT. An extracellular Cu2+ binding site in the voltage sensor of BK and Shaker potassium channels. The Journal of General Physiology. 2008;131:483–502. doi: 10.1085/jgp.200809980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hille B. Sunderland, Massachusetts, USA: Sinauer Associates, Inc; 2001. Ion Channels of Excitable Membranes, Third Edition. [Google Scholar]