

Abstract
Traditional investigations of the evolution of human social and political institutions trace their ancestry back to nineteenth century social scientists such as Herbert Spencer, and have concentrated on the increase in socio-political complexity over time. More recent studies of cultural evolution have been explicitly informed by Darwinian evolutionary theory and focus on the transmission of cultural traits between individuals. These two approaches to investigating cultural change are often seen as incompatible. However, we argue that many of the defining features and assumptions of ‘Spencerian’ cultural evolutionary theory represent testable hypotheses that can and should be tackled within a broader ‘Darwinian’ framework. In this paper we apply phylogenetic comparative techniques to data from Austronesian-speaking societies of Island South-East Asia and the Pacific to test hypotheses about the mode and tempo of human socio-political evolution. We find support for three ideas often associated with Spencerian cultural evolutionary theory: (i) political organization has evolved through a regular sequence of forms, (ii) increases in hierarchical political complexity have been more common than decreases, and (iii) political organization has co-evolved with the wider presence of hereditary social stratification.
Keywords: social evolution, phylogenetic comparative methods, cultural evolution, cultural phylogenetics, evolutionism, evolutionary trend
1. Introduction
The application of evolutionary theory to investigate human cultural diversity goes back to at least the founding of Anthropology in the latter half of the nineteenth century [1]. However, there has been considerable debate as to the most appropriate intellectual framework for understanding how human societies and cultures change over time [1–5].1 In this paper we draw a distinction between two research traditions (which we label ‘Spencerian’ and ‘Darwinian’), which show differences in their method and focus of investigations, and their concept of evolution. While these two approaches are often seen as incompatible, we argue that many of the defining features and assumptions of Spencerian evolutionary theories represent testable hypotheses that can be reconciled with an explicitly Darwinian view of cultural evolution. In this paper we demonstrate how these ideas can be tested using phylogenetic comparative analyses.
The archaeological record indicates that since the end of the last ice age there has been an overall increase in the scale and complexity of human groups. This trend has been described as ‘history's broadest pattern’ [6, p. 267], and ‘the most salient feature in human history’ [1, p. 288]. The idea of directionality is central to the concept and definition of evolution traditionally employed in Anthropology, particularly in studies of social and political organization: ‘…by evolved we mean—as I believe most anthropologists do—advanced along a trajectory of increasing complexity.’ [1, p. 276]. Other defining features of traditional social evolutionary hypotheses have been that human societies can be classified as falling into a few discrete forms of organization, and that societies pass through these different forms (or ‘stages’) in the same order across cultures [1,2,7–10]. For example, in Service's influential scheme of Band, Tribe, Chiefdom and State, small family groups of foragers (‘Bands’) evolve into larger, yet still essentially, egalitarian kinship-based, agricultural groups (‘Tribes’). Tribes evolve into societies where political leadership over a collection of local groups is centralized in a hereditary office of paramount chief. Chiefdoms finally evolve into States, characterized by a centralized political bureaucracy that contains specialized offices for decision-making and control functions [7,9]. To distinguish them from the research into cultural evolution described below, we label such studies Spencerian, acknowledging the influence of Herbert Spencer in the development of these ideas [1,11,12].
More recent evolutionary theories of cultural and social change draw their inspiration from the neo-Darwinian synthesis in Biology and apply the term ‘evolution’ in the same way that biologists do: to mean ‘descent with modification’ [3,4,12–16]. Studies within this framework are thus part of a larger movement in the social sciences to apply theories and methods developed originally in Evolutionary Biology to understand human behaviour and evolution [4,15]. Darwinian cultural evolutionary theory, sometimes labelled dual-inheritance theory, takes as its starting point the fact that humans inherit not only genetic information from their parents but also cultural information from a variety of sources, both of which affect the phenotype of an organism [4,13]. A key feature of such investigations is the use of techniques from population genetics to model the transmission of traits to help make sense of the complex interactions between genes and culture, and to examine how processes such as natural selection (working on both individuals and groups), cognitive biases and non-selective processes such as drift affect the maintenance and spread of cultural or genetic variants [3,15]. Thus far, this approach has been predominantly theoretical with the focus on the construction of mathematical models, which demonstrate how cultural evolutionary process can work [4,13,17]. Recently, methods from cultural psychology have also been applied to examine cultural transmission processes in experimental situations [18].
Spencerian evolutionary hypotheses have been criticized for trying to pigeon-hole societies into evolutionary types. These critics argue that the forms of social and political organization, and contingent historical pathways, are too numerous to fit easily within the proposed evolutionary sequences [19–22]. Furthermore, there has been a tendency to focus on changes towards increased socio-political complexity, yet there are many examples of societies that have decreased in complexity [22,23]. It also argued that these theories are deeply ethnocentric and that the perceived move towards more ‘civilized’ societies was seen as being universal, and indicative of progress in human affairs, i.e. ‘things were getting better’ [12,24]. Particularly in the earliest writings, ‘Civilized’ societies (as typified by the kind of Western nation states in which the theorists lived) were deemed to be superior to other kinds of societies. In the most extreme arguments, differences in ‘cultural attainment’ were put down to biological differences between societies and used as a justification for domination and colonization of traditional societies by European powers [24].
Darwinian researchers have often sought to distance themselves from these Spencerian hypotheses. For example, Richerson and Boyd state ‘The progressive evolutionary theories debated by generations of anthropologists have almost nothing in common with …[the] Darwinian notion of evolution’ [3, p. 59], and Mesoudi et al. contrast Darwinian theories of cultural evolution with the ‘progressive, unilinear theories … in which human societies were seen as progressing through a fixed set of stages, from ‘savagery’ through ‘barbarism’ to ‘civilisation’, which they describe as ‘erroneous’ and ‘flawed’ [15, p. 331]. While the Darwinian perspective offers a powerful way of investigating cultural evolution, this has not always been appreciated by Spencerians (e.g. [1,11]). This has partly been due to some researchers under the Darwinian banner wishing to draw too close an analogy between the mechanisms of genetic and cultural evolution. However, the true defining characteristic of Darwinian evolution is the notion of descent with modification, i.e. that heritable traits change over time. It does not matter whether innovations are accidental or purposefully acquired, or whether cultural traits are discrete, replicating entities, analogous to genes: the important point is that these traits are passed on with sufficient fidelity [25]. Furthermore, many interesting aspects of cultural evolution are the results of differences between biological and cultural systems such as modes of transmission other than parent-to-offspring [13], and the possibility of the widespread adoption of behaviours that are maladaptive from a biological perspective [4].
In many respects, the focus of research of these two approaches has been very different. Darwinian cultural evolutionists tend to examine the micro-evolutionary processes affecting the frequencies of cultural traits within populations of individuals [15]. Unlike the Spencerians, these researchers have generally not tackled questions relating to long-term macro-evolutionary patterns and processes in the evolution of social and political organization since the last ice age. Also, it is important to separate out the normative critiques that Spencerian evolutionary theory is ethnocentric, racist or progressive, from the many empirical questions relating to these topics which remain unresolved and could benefit from being addressed within an explicitly Darwinian framework. We argue that many of the assumptions and defining features of Spencerian theories represent testable hypotheses about the mode and tempo of social and cultural evolution, and that, thus far, there has been a distinct lack of quantitative tests of many of these ideas.
While archaeological information has an important role to play in addressing these questions [10], data may not always be available in sufficient detail for a large enough number of sites to allow a rigorous test of competing hypotheses. This is a common problem in investigations of prehistory, and in such cases it is important to supplement the insights from archaeology with those of other disciplines [26]. Recently, researchers interested in cultural evolution have begun to apply phylogenetic methods, originally developed in Evolutionary Biology, to empirically investigate questions relating to the pattern and process of human cultural evolution within a Darwinian framework [5]. These techniques have been used to address questions about general evolutionary processes such as whether rates of linguistic change are linked to such factors as the frequency with which words are used [27], as well more specific questions relating to the longue durée of human history such as the dispersal and diversification of widespread language families [28], the ancestral forms of residence practices in South-East Asia and the Pacific [26], and the adoption of cattle and changes in inheritance systems in sub-Saharan Africa [29]. In this paper, we show how a particular type of phylogenetic analysis, Phylogenetic Comparative Methods (PCMs) [5], can be used to test hypotheses relating to three aspects of socio-political evolution that have been key features of Spencerian hypotheses, but which have often been questioned: (i) the sequence of evolution of human groups (do changes in political organization follow a regular sequence?), (ii) the direction of evolution (have societies increased in the scale and complexity of their organization over time, and if so, have increases tended to be more common than decreases?), and (iii) the co-evolution of social traits (do different aspects of social organization tend to change together?).
2. Analyses
A phylogenetic tree (or sample of trees) represents a hypothesis about the historical relationships between the units of analysis. PCMs make statistical inferences about evolutionary processes by mapping traits onto the tips of these trees. These techniques are based on the logical proposition that given data about the present distribution of traits across taxa and knowledge about the historical relationships between these taxa, it is possible to infer what the traits were like in the past and how they have changed to give rise to their present distribution [30]. These methods therefore offer a principled way of investigating the long-term patterns and processes of cultural evolution. Here we combine ethnographic data from Austronesian-speaking societies of Island South-East Asia and the Pacific with a sample of phylogenetic trees, which have previously been constructed using linguistic data (figure 1) to perform phylogenetic comparative analyses using Maximum Likelihood and Stochastic Character Mapping techniques in the software packages BayesTraits (http://www.evolution.rdg.ac.uk/BayesTraits.html) and SIMMAP (http://www.simmap.com/) (see electronic supplementary material).
Figure 1.

Geographical distribution of the 84 Austronesian-speaking societies analysed in this study and their associated forms of political organization (top right). Data on political organization mapped on to a phylogeny of these societies (centre). This figure shows a single phylogenetic tree from the sample of 1000 most likely trees. Analyses in §2 were conducted using all 1000 trees. Colours along the branches indicate the inferred forms of organization from a single stochastic character mapping (SCM). Pie chart at the root of the tree represents the proportional probabilities of different forms of political organization in the ancestral Austronesian society inferred using maximum likelihood. Distributions of number of changes between states inferred using SCM (bottom left). Unfilled circle, Acephalous; blue circle, Simple Chiefdom; red circle, Complex Chiefdom; black circle, State.
(a). Evolutionary sequences
A key feature of Spencerian hypotheses is that changes in human socio-political organization follow evolutionary sequences (e.g. Band, Tribe, Chiefdom and State discussed earlier). Such classificatory schemes contain two logically distinct elements: (i) societies are grouped together based on observed similarities in the way they are organized, (ii) the categories are arranged on a scale of complexity with societies hypothesized to pass through adjacent stages of organization in the direction of increasing socio-political complexity. Therefore, it is perfectly possible to classify societies according to some criteria without this classification representing an evolutionary sequence, and societies could evolve without having to pass through the same stages in the same order. For our present purposes we can classify political organization into four categories of increasing complexity based on the number of hierarchical decision-making levels in a society. Acephalous societies are organized politically only at the level of the local community (e.g. the village). Simple Chiefdoms have one permanent level of leadership uniting several villages, while Complex Chiefdoms have two levels. Societies with more than two hierarchical decision-making levels above the local community are labelled ‘States’ (see electronic supplementary material). Whether this classification does in fact represent an evolutionary sequence is an empirical question on which suitable lines of evidence must be brought to bear.
PCMs can be used to examine the support for evolutionary sequences, and are commonly used to address questions about evolutionary pathways in biological evolution (e.g. the evolution of Carnivore social systems [31]). We have previously evaluated six different models of the evolution of political organization [32] (figure 2), which are derived from discussions in the literature. Three of these models reflect the Spencerian hypothesis that change in political organization has been sequential in the direction of increasing hierarchical complexity (i.e. the transitions Acephalous society to Simple Chiefdom, Simple Chiefdom to Complex Chiefdom, Complex Chiefdom to State have occurred, but the larger, direct increases Acephalous society to Complex Chiefdom, Acephalous society to State and Simple Chiefdom to State have not). The RECTILINEAR model reflects the idea that only sequential increases in complexity can occur. This view is often attributed to the classical evolutionists such as Spencer and Morgan [1]. We also specified two models in which increases are sequential but decreases are also possible. In the UNILINEAR model, decreases occur only to adjacent levels of complexity, while in the RELAXED UNILINEAR model, decreases can occur to any lower level. In contrast, in the other three models increasing political complexity does not follow a regular sequence. We specified two models based on the idea that different forms or organization have developed along separate evolutionary pathways having evolved from an acephalous form of organization [22]: ALTERNATIVE TRAJECTORIES (only increases possible) and ALTERNATIVE TRAJECTORIES (REVERSIBLE) (decreases also possible). Finally, in the FULL model any change is possible, representing the idea that political organization has been completely unconstrained.
Figure 2.
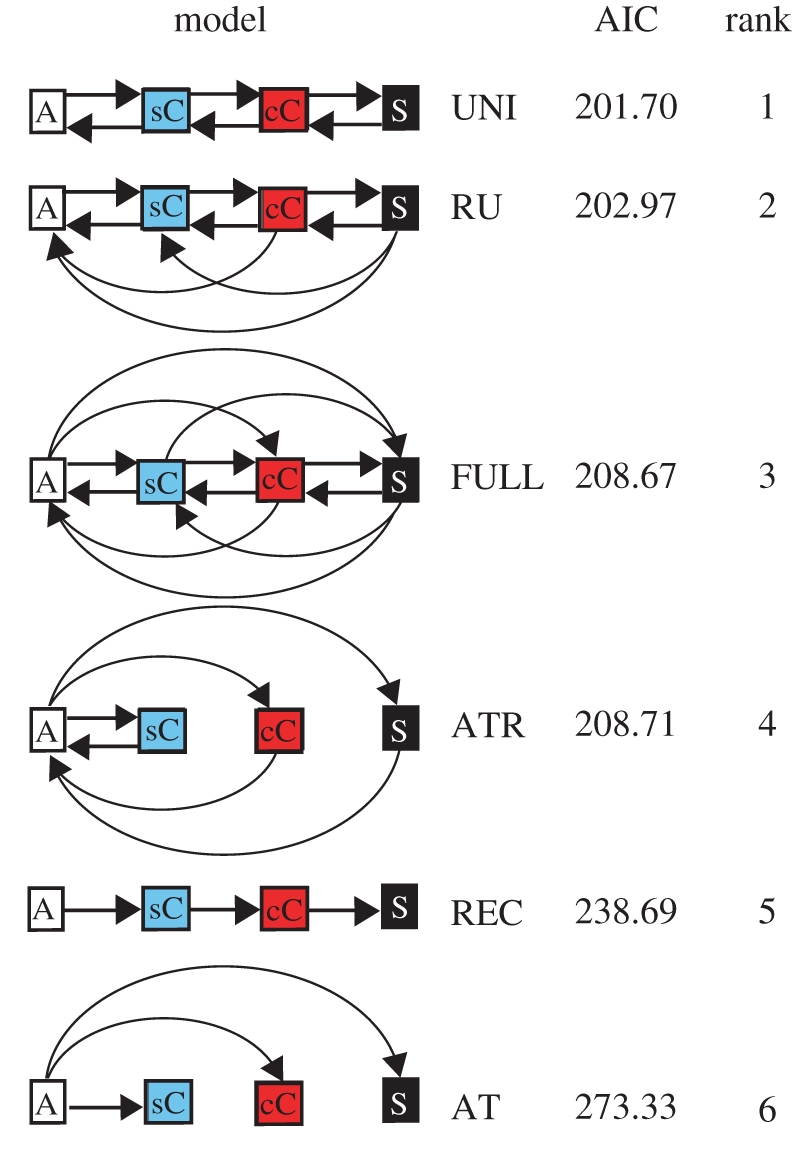
Support for different models of political evolution based on the Akaike Information Criterion (AIC) (REC, Rectilinear; UNI, Unilinear; RU, Relaxed Unilinear; FULL, Full model; AT(R), Alternative Trajectories (Reversible)) (adapted from Currie et al. [32]).
Analyses show the UNILINEAR model to be the best supported, closely followed by the RELAXED UNILINEAR model (table 1). The FULL model and the reversible version of the ALTERNATIVE TRAJECTORIES model are less well supported, while those models that do not allow declines in political complexity (i.e. RECTILINEAR and non-reversible ALTERNATIVE TRAJECTORIES model) are even poorer fits to the data. Overall, the analyses provide strong support for the type of sequences of political evolution that have formed a core feature of the Spencerian hypotheses of cultural evolution. Importantly, they highlight that change has not always been in the direction of increasing complexity. Whether increases in complexity have been more common than decreases is dealt with in the next section.
Table 1.
Percentage of Stochastic Character Mappings in which increases or decreases in complexity are more common, and comparisons of inferred number of changes between different forms of organization using paired sample t-tests (d.f. = 99 998). In all comparisons, the number of changes towards the more complex form of organization is significantly higher than the number of changes towards less complex forms.
| comparison | % increases | % decreases | % equal | mean increases | mean decreases | mean difference | s.d. | t | p |
|---|---|---|---|---|---|---|---|---|---|
| A → sC versus sC → A | 60.3 | 27.0 | 12.7 | 11.1 | 10.6 | 0.6 | 3.15 | 55.98 | <0.001 |
| sC → cC versus cC → sC | 99.9 | <0.1 | <0.1 | 17.9 | 2.0 | 15.9 | 3.23 | 1561.99 | <0.001 |
| cC → S versus S → cC | 100 | 0 | 0 | 5.1 | 0.1 | 5.0 | 0.05 | 34 107.00 | <0.001 |
| sC → cC versus sC → A | 98.2 | 0.9 | 0.8 | 17.9 | 10.6 | 7.4 | 2.81 | 828.38 | <0.001 |
| cC → S versus cC → sC | 93.0 | 3.7 | 3.3 | 5.1 | 2.0 | 3.1 | 1.72 | 569.15 | <0.001 |
(b). Direction of evolution
Another defining feature of the Spencerian hypotheses is that there is a direction to cultural evolution, i.e. the complexity of socio-political organization increases over time. That the archaeological record indicates an overall increase in complexity since the end of the last ice age is not in dispute [33]. Yet, the archaeological and historical records also indicate periods when societies have decreased in complexity [23], which is supported by the findings described in §2a, and it is unclear if increases have generally been more common than decreases. Additionally, it is important to understand how such a macro-evolutionary trend can arise if cultural evolution, like biological evolution, is not goal directed.
Although Spencerian cultural evolutionary theories have been characterized as assuming that increases in complexity have dominated, there has, in fact, been a lack of consensus on this issue [1]. For example, Tylor proposed that human history ‘is not the history of a course of degeneration, or even of equal oscillations to and fro, but of a movement which, in spite of frequent stops and relapses, has on the whole been forward’ (Tylor 1870, p. 193 cited in [1, p. 28]), while Spencer argued that ‘the theory of progression, in its ordinary form, seems to me to be untenable…It is possible, and, I believe, probable, that retrogression has been as frequent as progression’ (Spencer 1890, p. 93, cited in [1, p. 27]). More recently, Diamond states that increasing complexity is ‘no more than an average long-term trend, with innumerable shifts in either direction: 1000 amalgamations for 999 reversals’ [6, p. 281]. Richerson & Boyd [33], while acknowledging that decreases in complexity can and have occurred, clearly see increases in complexity as more common, arguing that complex social organization is compulsory in the long run, owing to the competitive advantage that societies hold in competition between groups.
Despite the fact that biological evolution is not goal-directed, there are a number of large-scale trends that can be witnessed over evolutionary time. A trend here is defined as a directional shift in a measurement value of some attribute over time (e.g. the extreme of a distribution, or a measure of its central tendency) [34]. Some trends are present only in certain clades and over certain time scales, while others seem to hold over the entire history of life on earth (e.g. increases in the maximum degree of biological complexity, and body size—see ref. [35] for a summary of the proposed macro-evolutionary trends). In our own lineage, the trend of increasing brain size in homonins is well-established even if there is much discussion as to the reasons for it [36].
One explanation for such macro-evolutionary trends is that selection has favoured a consistent directional shift in the trait in question. For example, it has been argued that selection acts as a driving force favouring increased body size owing to the potential advantages that are gained from being large (e.g. more efficient metabolism and homeostasis, advantage in competition over resources, greater mobility, and decreased predation) [37]. However, it also possible for trends to occur even in the absence of such a driving force if the ‘phase space’ in which a trait is evolving is constrained and the trait originally arises near one of the constraints (a so-called ‘passive’ trend) [38]. For example, the trend of increasing biological complexity could be the result of the earliest single-celled organisms arising near a ‘left-wall’ of minimum complexity, i.e. it was almost impossible for them to have been any less complex [34]. Initially any changes in complexity must be in the direction of increased complexity. Subsequently, increases and decreases are possible, yet over time the maximum degree of complexity will increase. Similar mechanisms could explain the trend towards increasing political complexity (figure 3).
Figure 3.

Alternative trend mechanisms underlying the increase in political complexity over time. (b) In a driven trend, change is biased with increases in complexity more likely than decreases (this represents an extreme example in which only increases have occurred). (a) In a passive trend, increases and decreases are equally likely. An increase in the maximum degree of complexity over time occurs because the ancestral lineage arose near a ‘left wall’ of minimum complexity, meaning evolution in the direction of decreasing complexity was initially limited. Phylogenetic comparative methods can be used to test between these scenarios.
Biologists have employed PCMs to investigate trends in biological evolution [39]. Here we use a PCM to directly estimate the number of changes between forms of political organization to assess whether increases in complexity have actually been more common than decreases. As a first step we inferred the form of political organization in the ancestral Austronesian society under the best-fitting model of evolution from the previous analysis, the UNILINEAR model. The results suggest that the ancestral Austronesian society was politically acephalous (proportional support for different forms of organization: Acephalous = 0.76, Simple Chiefdom = 0.22, Complex Chiefdom = 0.01, State = 0.00) (figure 1), which confirms that the maximum degree of hierarchical political organization in Austronesian societies has indeed increased over time. We then used a PCM to infer the number of increases and decreases between levels of complexity under the UNILINEAR model of trait evolution. Figure 1 shows the distributions of the estimated numbers of changes from these analyses. We can see that increases in complexity have occurred more frequently than decreases (table 1) (although in the comparison between changes A → sC and sC → A, the mean difference was less than one). As the more complex forms of political organization evolved later, the significant differences in the first three comparisons could be owing to the fact that more time has been spent in the form of lower complexity, therefore allowing for more opportunities for increases to occur. The final two comparisons are situations where there were equal opportunities for increases or decreases and in these comparisons increases are again significantly greater than decreases. These results suggest that increases in political complexity in Austronesian-speaking societies have generally been more common than decreases.
(c). Co-evolution of social and political traits
Another aspect of social evolution that has been the subject of considerable debate is the idea that different aspects of social organization are correlated with one another, i.e. classificatory schemes such as Band, Tribe, Chiefdom and State are based on regular hypothesized differences between these categories in such things as subsistence practices, degree of social differentiation, inherited inequalities and permanent offices of leadership [1,7,9]. An associated idea is that change from one category to another involves the relatively rapid restructuring of these different aspects of social organization, i.e. socio-political evolutionary change is punctuational [2]. However, it has been argued that different aspects of social organization do not co-evolve this closely and that societies exhibit too much variation to fit easily into categories such Band, Tribe, Chiefdom and State [19].
There has been a lack of quantitative comparative analyses attempting to address this question. We examined whether political organization co-evolves with hereditary social stratification (i.e. some individuals or groups of individuals within a society are afforded higher social status and have greater influence owing to who their ancestors were). In traditional social evolutionary theories, Chiefdoms and States are thought to be socially stratified along these lines, while such hereditary ranking is thought to be absent in societies organized politically only at the level of the local community (i.e. Bands and Tribes) [6,7,9]. Table 2 shows the co-occurrence of these two aspects of socio-political organization in our sample, which suggests that Acephalous societies generally lack hereditary forms of social stratification, while it is generally present in Chiefdoms and States. However, as societies are hierarchically related they may have several features in common, not because they are functionally linked, but because they have all been inherited from a common ancestral society (e.g. most of the Polynesian societies in the sample are organized as chiefdoms and have hereditary social stratification (see electronic supplementary material, figure S2), which potentially could be owing to either process). Therefore, societies cannot be treated as independent data points in a cross-cultural analysis. Phylogenetic comparative analyses can overcome these problems by identifying whether the traits under investigation are co-evolving while controlling for the historical relatedness between societies [5].
Table 2.
Contingency table showing the occurrence of different forms of political organization and the presence or absence of hereditary social stratification in our sample.
| political organization |
||
|---|---|---|
| hereditary social stratification | Acephalous | Chiefdoms/States |
| absence | 31 | 3 |
| presence | 12 | 37 |
To formally test whether political organization has co-evolved with the wider presence of hereditary social stratification, we use a PCM to compare two alternative models of trait evolution: (i) a dependent model where the rate of change of one trait is different depending on the state of the other, and (ii) an independent model where the rate of trait change does not vary according to the state of the other (see electronic supplementary material, figure S1). For our sample of Austronesian-speaking societies, the dependent model of evolution fits the data much better than the independent model (figure 4 and electronic supplementary material). These results support the hypothesis that political organization, as represented by the number of hierarchical jurisdictional levels, has indeed co-evolved with the wider presence of hereditary social stratification in Austronesian societies.
Figure 4.

Flow diagram showing the estimated rates of change between different combinations of the binary variables of Class Stratification and Jurisdictional Hierarchy under the dependent model of evolution. This dependent model has a much better fit to the data (AIC: 176.2) than the independent model of evolution (AIC: 215.6) (see electronic supplementary material). Relative thickness of arrows represents relative estimated rate parameters, with the dotted line representing an estimated rate of zero.
Figure 4 shows the estimated rates of change between the different combinations of these variables, and thus shows the most likely pathways of evolution of these traits. It appears that Acephalous societies lacking hereditary class distinctions can develop either hereditary class distinctions or a chiefdom form of political organization first. However, the rate of change from Acephalous society to Chiefdom is greater in the presence of hereditary class distinctions, while the particular form of political organization does affect the rate at which hereditary class distinctions evolve. Interestingly, the rates of change away from the intermediate states (i.e. Bands or Tribes exhibiting hereditary class distinctions and vice versa) are generally higher than those going towards these intermediate states. This suggests that these intermediate forms of organization are unstable, which is consistent with the idea that different features of social organization may alter relatively rapidly once other elements have changed.
3. Discussion
In this paper we have demonstrated how PCMs can be used to test hypotheses about the macro-evolutionary patterns and processes of socio-political evolution. Despite suggestions that Darwinian and Spencerian evolutionary approaches are incompatible, we actually found support for three defining features of Spencerian evolutionary theories. First, political organization has evolved through a regular sequence in the direction of increasing levels of hierarchal complexity. Secondly, the overall direction of political evolution has been from less hierarchically complex to more hierarchically complex, and the number of increases in complexity appears to have been greater than the number of decreases. Finally, hierarchical political organization appears to have co-evolved with hereditary forms of social stratification with politically Acephalous societies lacking hereditary distinctions, while they are generally present in societies organized politically as Chiefdoms and States. Interestingly, the intermediate combinations of these traits were relatively less stable, which is consistent with a punctuational process of socio-political change. These studies illustrate the benefits a Darwinian evolutionary framework can have in addressing important questions relating to these aspects of social and cultural change, while avoiding many of the problems associated with the traditional Spencerian studies.
Despite the logical possibility that political evolution could involve larger jumps in hierarchical complexity, our results indicate that in fact only sequential increases have occurred. This may reflect constraints imposed by a number of micro-evolutionary forces, such as a social psychology, that evolved in the context of life in small-scale groups requiring the evolution of cultural ‘work-arounds’ [40], the difficulty in reorganizing existing institutions that rely on the coordination of large numbers of individuals, or the requirement for certain pre-existing institutions or cultural practices [33]. More complex forms of political organization appear to result from the welding together of smaller pre-existing units [6], a process which requires the cultural evolution of institutions that allow these large-scale groups to be stable. For example, it has been argued that a stable form of state organization uniting previously independent Chiefdoms in Hawaii was made possible only by the presence of administrative institutions such as local land managers (konohiki) under the control of district chiefs [2]. Just as biologists seek to understand what constrains biological evolution to take small steps in ‘design space’ rather than large, saltational jumps [41], future work could examine what factors have been most important in preventing larger increases in political organization.
The results suggest that while decreases in political complexity have occurred, increases have been more common. This is consistent with the idea that increases in complexity over time have been the result of a driven trend mechanism [38]. This may be owing to selection having favoured more complex forms of political organization because they improve the coordination, cooperation and division of labour between individuals, thus providing an advantage in direct or indirect competition between groups [6,33,42]. It should be stressed that these potential advantages to more complex forms of political organization may not always be realized and may be contingent on the presence of other factors. For example, environmental instability in the Pleistocene appears to have created a ‘right wall’ of maximum complexity that could not be breached until the climatic amelioration of the Holocene allowed the evolution of agriculture [3]. Furthermore, the rate of political evolution has not been even across the world, a fact that has had important consequences for the present day [6,33,42]. Future work should aim to empirically investigate rates of political evolution and which factors have been most important in determining these rates.
We also found support for the idea that different aspects of socio-political organization evolve together. Furthermore, the estimated rates of change were consistent with a punctuational process of socio-political evolution. So, as forms of socio-political organization follow a pattern of sequential increases in complexity, the change from one form to another may involve the relatively rapid change of several different aspects of social structure. This highlights the fact that a Darwinian view of evolution does not require an adherence to the idea that evolutionary change is always a continuous, gradual process. Indeed, investigating the rate at which evolution proceeds, and how it can vary over time or across different traits, is important for both biological and cultural phenomena [27,28]. The co-evolutionary techniques we have outlined will be applied to test the extent to which other social and cultural features (such as those described in §2c) evolve in a correlated fashion. This will allow us to build up a network of the dependencies between these traits in a manner similar to the way in which PCMs have been used to investigate networks of functional linkages between genes [43]. Additionally, many different factors have been proposed to be important in leading to more complex forms of political organization including such things as intensification of subsistence practices, warfare and trade [1,9]. With their ability to make inferences about the order in which traits change, these methods raise the possibility of being able to provide more rigorous tests of such causal hypotheses. For example, Holden and Mace [29] show that in Bantu-speaking societies of sub-Saharan Africa, the adoption of cattle-herding led to the loss of matrilineal forms of inheritance.
It is important to point out that the idea that socio-political organization can be classified as falling into a limited number of forms does not imply that societies belonging to these categories will be exactly the same. Instead, it is proposed that there is a ‘limited array of basic designs underlying the apparently wide range of variability’ [2]. For example, in Hawaiian society, both secular and religious power were concentrated in the paramount Ali'i ai moku chiefs, while in Tonga, a dual paramountship was in operation with the Tu'i Tonga as a spiritual figurehead and the Hau chief wielding political authority [44]. Yet both societies can be classified as Complex Chiefdoms in terms of the number of hierarchical decision-making levels. Furthermore, the question of whether or not several features of social and political organization co-evolve does not mean that any functional associations between traits must follow the previously proposed schemes such as Band, Tribe, Chiefdom and State. Other forms of patterned variation not captured by these schemes may be observable. For example, Blanton and colleagues [45] argue that the Mesoamerican archaeological sites they investigate are best described as representing either ‘network’ polities, characterized by self-aggrandizing rulers, or ‘corporate’ polities, in which leaders had minimized personal identities and visibility. We feel the extent to which societies can be usefully classified based on different features is an empirical question that is best answered using the kind of quantitative analyses we have advocated in this paper.
We have argued that the focus of Spencerian and Darwinian evolutionary approaches has often been very different, with Spencerians tending to focus on broad-scale macro-evolutionary phenomena. An important goal of research in Evolutionary Biology is to understand the relationship between micro-evolution and macro-evolution [46]. A Darwinian view of cultural evolution is also well suited to addressing such questions with respect to cultural and social evolution [33]. Recent empirical research has begun to explore how cultural transmission between individuals relates to patterns of cultural diversity at the population level [16]. Similarly, attention needs to be paid to how particular forms of social and political organization emerge and are maintained from the perspective of the interactions between individuals and the transmission of institutions, customs, practices and rules of behaviour within and between generations. Mathematical models and computer simulations of such micro-evolutionary processes promise to shed light on the evolution of socio-political organization (e.g. [17]) and provide an important complement to the phylogenetic techniques we have employed here.
The use of PCMs avoids some of the problems attributed to the Spencerian approach. For example, Shennan [14] argues that often archaeological investigations within this framework have done little more than propose that the remains of a society found at an archaeological site are indicative of a particular stage of evolution, without understanding the processes by which societies and culture change. Spencerian evolutionary theory has also been criticized for taking a progressive, archaic, ladder-like view of evolution [22]. The use of PCMs makes the fact explicit that the societies under investigation are contemporaries and enables us to make inferences about the sequence in which the traits of these societies, such as their form of political organization, have evolved. Furthermore, they allow us not only to infer the pattern of trait changes over time, but also what processes have occurred to give rise to the diversity of forms of organization we witness in the ethnographic record.
Our goal in this paper has been to highlight the huge potential that the use of PCMs within a Darwinian evolutionary framework hold for addressing questions relating to the pattern and process of socio-political evolution. Although Darwinian and Spencerian evolutionists have often had fundamental disagreements about the concept of ‘evolution’, and despite suggestions that the two approaches are incompatible, we have demonstrated how some of these differences can be resolved. In biology, the concept of evolution provides a unifying principle around which a large body of information can be organized and synthesized [47]. Given the great success of Darwinism in helping us understand the natural world, it is hoped that the theory of evolution as ‘descent with modification’ can be the same kind of organizing principle for anthropology that it is in biology [15]. The use of PCMs to investigate socio-political evolution is a key step forward in this endeavour, allowing us to go beyond the purely verbal arguments which have hitherto dominated the debates on this topic, and to provide quantitative tests of opposing hypotheses.
Acknowledgements
T.E.C. was supported by an ESRC/NERC Interdisciplinary Research Studentship and a Japan Society for the Promotion of Science Postdoctoral Fellowship. R.M. was supported by a European Research Council grant. We thank Russell Gray and Simon Greenhill for providing the Austronesian phylogenetic trees, and who, along with Robert Foley, Michael Dunn and Fiona Jordan, provided many valuable comments at various stages during this research. We also thank Andrew Whiten, and two anonymous reviewers for their comments on the manuscript.
Endnote
In addition to the references listed below an extended bibliography is available as electronic supplementary material.
One contribution of 26 to a Discussion Meeting Issue ‘Culture evolves’.
References
- 1.Carneiro R. L. 2003. Evolutionism in cultural anthropology. Boulder, CO: Westview Press [Google Scholar]
- 2.Spencer C. S. 1990. On the tempo and mode of state formation: neoevolutionism reconsidered. J. Anth. Arch. 9, 1–30 10.1016/0278-4165(90)90004-W (doi:10.1016/0278-4165(90)90004-W) [DOI] [Google Scholar]
- 3.Richerson P. J., Boyd R. 2005. Not by genes alone. Chicago, IL: University of Chicago Press [Google Scholar]
- 4.Laland K. N., Brown G. R. 2002. Sense and nonsense. Oxford, UK: Oxford University Press [Google Scholar]
- 5.Mace R., Holden C. J. 2005. A phylogenetic approach to cultural evolution. Trends Ecol. Evol. 20, 116–121 10.1016/j.tree.2004.12.002 (10.1016/j.tree.2004.12.002) [DOI] [PubMed] [Google Scholar]
- 6.Diamond J. 1997. Guns, germs and steel. London, UK: Vintage [Google Scholar]
- 7.Service E. R. 1962. Primitive social organization. New York, NY: Random House [Google Scholar]
- 8.Earle T. 1994. Political domination and social evolution. In Companion encyclopedia of anthropology (ed. Ingold T.), pp. 940–961 London, UK: Routledge [Google Scholar]
- 9.Flannery K. V. 1972. The cultural evolution of civilizations. Ann. Rev. Ecol. Evol. Syst. 3, 399–426 10.1146/annurev.es.03.110172.002151 (doi:10.1146/annurev.es.03.110172.002151) [DOI] [Google Scholar]
- 10.Marcus J. 2008. The archaeological evidence for social evolution. Ann. Rev. Anth. 37, 251–266 10.1146/annurev.anthro.37.081407.085246 (doi:10.1146/annurev.anthro.37.081407.085246) [DOI] [Google Scholar]
- 11.Graber R. B. 2007. Bye-Bye BABY! A cultural evolutionist's response to evolutionary culture theorists' complaints. Social Evol. Hist. 6, 3–28 [Google Scholar]
- 12.Dunnell R. C. 1980. Evolutionary theory and archaeology. Adv. Archaeol. Method Theory 3, 35–99 [Google Scholar]
- 13.Boyd R., Richerson P. J. 1985. Culture and the evolutionary process. Chicago, IL: Chicago University Press [Google Scholar]
- 14.Shennan S. 2002. Genes, memes, and human history. London, UK: Thames & Hudson [Google Scholar]
- 15.Mesoudi A., Whiten A., Laland K. N. 2006. Towards a unified science of cultural evolution. Behav. Brain Sci. 29, 329–383 [DOI] [PubMed] [Google Scholar]
- 16.Shennan S. 2011. Descent with modification and the archaeological record. Phil. Trans. R. Soc. B 366, 1070–1079 10.1089/rstb.2010.0380 (doi:10.1089/rstb.2010.0380). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Henrich J., Boyd R. 2008. Division of labor, economic specialization, and the evolution of social stratification. Curr. Anth. 49, 715–724 10.1086/587889 (doi:10.1086/587889) [DOI] [Google Scholar]
- 18.Mesoudi A., O'Brien M. J. 2008. The cultural transmission of Great Basin projectile-point technology. I. an experimental simulation. Am. Antiquity 73, 3–28 [Google Scholar]
- 19.Feinman G. M. 2008. Variability in states: comparative frameworks. Social Evol. Hist. 7, 54–66 [Google Scholar]
- 20.Campbell R. B. 2009. Toward a networks and boundaries approach to early complex polities: the Late Shang case. Curr. Anth. 50, 821–848 10.1086/648398 (doi:10.1086/648398) [DOI] [Google Scholar]
- 21.Bondarenko D. M., Grinin L. E., Korotayev A. V. 2002. Alternative pathways of social evolution. Social Evol. Hist. 1, 54–79 [Google Scholar]
- 22.Yoffee N. 1993. Too many chiefs? (or, safe texts for the '90s). In Archaeological theory: who sets the agenda? (eds Yoffee N., Sherratt A.), pp. 60–78 Cambridge, UK: Cambridge University Press [Google Scholar]
- 23.Tainter J. A. 2006. Archaeology of overshoot and collapse. Ann. Rev. Anth. 35, 59–74 10.1146/annurev.anthro.35.081705.123136 (doi:10.1146/annurev.anthro.35.081705.123136) [DOI] [Google Scholar]
- 24.Eriksen T. H., Nielsen F. S. 2001. A history of anthropology. London, UK: Pluto Press [Google Scholar]
- 25.Henrich J., Boyd R., Richerson P. J. 2008. Five misunderstandings about cultural evolution. Hum. Nat. 19, 119–137 10.1007/s12110-008-9037-1 (doi:10.1007/s12110-008-9037-1) [DOI] [PubMed] [Google Scholar]
- 26.Jordan F. M., Gray R. D., Greenhill S. J., Mace R. 2009. Matrilocal residence is ancestral in Austronesian societies. Proc. R. Soc. B 276, 1957–1964 10.1098/rspb.2009.0088 (doi:10.1098/rspb.2009.0088) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Calude A. S., Pagel M. 2011. How do we use language? Shared patterns in the frequency of word use across 17 world languages. Phil. Trans. R. Soc. B 366, 1101–1107 10.1089/rstb.2010.0315 (doi:10.1089/rstb.2010.0315). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gray R. D., Atkinson Q. D., Greenhill S. J. 2011. Language evolution and human history: what a difference a date makes. Phil. Trans. R. Soc. B 366, 1090–1100 10.1089/rstb.2010.0378 (doi:10.1089/rstb.2010.0378). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Holden C. J., Mace R. 2003. Spread of cattle led to the loss of matrilineal descent in Africa: a coevolutionary analysis. Proc. R. Soc. Lond. B 270, 2425–2433 10.1098/rspb.2003.2535 (doi:10.1098/rspb.2003.2535) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pagel M. 1999. Inferring the historical patterns of biological evolution. Nature 401, 877–884 10.1038/44766 (doi:10.1038/44766) [DOI] [PubMed] [Google Scholar]
- 31.Dalerum F. 2007. Phylogenetic reconstruction of carnivore social organizations. J Zool 273, 90–97 10.1111/j.1469-7998.2007.00303.x (doi:10.1111/j.1469-7998.2007.00303.x) [DOI] [Google Scholar]
- 32.Currie T. E., Greenhill S. J., Gray R. D., Hasegawa T., Mace R. 2010. Rise and fall of political complexity in island South-East Asia and the Pacific. Nature 467, 801–804 10.1038/nature09461 (doi:10.1038/nature09461) [DOI] [PubMed] [Google Scholar]
- 33.Richerson P. J., Boyd R. 2001. Institutional evolution in the Holocene: the rise of complex societies. In The origin of human social institutions (ed. Runciman W. G.), pp. 197–234 Oxford, UK: Oxford University Press [Google Scholar]
- 34.Gould S. J. 1997. Life's grandeur: the spread of excellence from Plato to Darwin. London, UK: Vintage [Google Scholar]
- 35.McShea D. W. 1998. Possible largest-scale trends in organismal evolution: eight ‘live hypotheses’. Ann. Rev. Ecol. Syst. 29, 293–318 10.1146/annurev.ecolsys.29.1.293 (doi:10.1146/annurev.ecolsys.29.1.293) [DOI] [Google Scholar]
- 36.Lee S. H., Wolpoff M. H. 2003. The pattern of evolution in Pleistocene human brain size. Paleobiology 29, 186–196 (doi:10.1666/0094-8373(2003)029<0186:TPOEIP>2.0.CO;2) [DOI] [Google Scholar]
- 37.Kingsolver J. G., Pfennig D. W. 2004. Individual-level selection as a cause of Cope's rule of phyletic size increase. Evolution 58, 1608–1612 10.1111/j.0014-3820.2004.tb01740.x (doi:10.1111/j.0014-3820.2004.tb01740.x) [DOI] [PubMed] [Google Scholar]
- 38.McShea D. W. 1994. Mechanisms of large-scale evolutionary trends. Evolution 48, 1747–1763 10.2307/2410505 (doi:10.2307/2410505) [DOI] [PubMed] [Google Scholar]
- 39.Marcot J. D., McShea D. W. 2007. Increasing hierarchical complexity throughout the history of life: phylogenetic tests of trend mechanisms. Paleobiology 33, 182–200 10.1666/06028.1 (doi:10.1666/06028.1) [DOI] [Google Scholar]
- 40.Richerson P. J., Boyd R. 1999. Complex societies: the evolutionary origins of a crude superorganism. Hum. Nat. 10, 253–289 10.1007/s12110-999-1004-y (doi:10.1007/s12110-999-1004-y) [DOI] [PubMed] [Google Scholar]
- 41.Dennett D. C. 1996. Darwin's dangerous idea: evolution and the meanings of life. New York, NY: Simon & Schuster [Google Scholar]
- 42.Currie T. E., Mace R. 2009. Political complexity predicts the spread of ethnolinguistic groups. Proc. Natl Acad. Sci. USA 106, 7339–7344 10.1073/pnas.0804698106 (doi:10.1073/pnas.0804698106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Barker D., Pagel M. 2005. Predicting functional gene links from phylogenetic-statistical analyses of whole genomes. PLoS Comput. Biol. 1, e3. 10.1371/journal.pcbi.0010003 (doi:10.1371/journal.pcbi.0010003) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kirch P. V. 1984. The evolution of the Polynesian chiefdoms. Cambridge, UK: Cambridge University Press [Google Scholar]
- 45.Blanton R. E., Feinman G. M., Kowalewski S. A., Peregrine P. N. 1996. A dual-processual theory for the evolution of Mesoamerican civilization. Curr. Anth. 37, 1–14 10.1086/204471 (doi:10.1086/204471) [DOI] [Google Scholar]
- 46.Penny D., Phillips M. J. 2004. The rise of birds and mammals: are microevolutionary processes sufficient for macroevolution. Trends Ecol. Evol. 19, 516–522 10.1016/j.tree.2004.07.015 (doi:10.1016/j.tree.2004.07.015) [DOI] [PubMed] [Google Scholar]
- 47.Dobzhansky T. 1973. Nothing in biology makes sense except in light of evolution. Am. Biol. Teacher 35, 125–129 [Google Scholar]