

Abstract
The mechanisms by which administration of interferon-α induces neuropsychiatric side effects, such as depressive symptoms and changes in cognitive function, are not clear as yet. Direct influence on serotonergic neurotransmission may contribute to these side effects. In addition, the enzyme indoleamine 2,3-dioxygenase (IDO), which converts tryptophan into kynurenine, may play an important role, first, because IDO activation leads to reduced levels of tryptophan, the precursor of serotonin (5-HT), and thus to reduced central 5-HT synthesis. Second, kynurenine metabolites such as 3-hydroxy-kynurenine (3-OH-KYN) and quinolinic acid (QUIN) have toxic effects on brain function. 3-OH-KYN is able to produce oxidative stress by increasing the production of reactive oxygen species (ROS), and QUIN may produce overstimulation of hippocampal N-methyl-D-aspartate (NMDA) receptors, which leads to apoptosis and hippocampal atrophy. Both ROS overproduction and hippocampal atrophy caused by NMDA overstimulation have been associated with depression.
Medical subject headings: depression, interferon-alpha, serotonin, tryptophan
Abstract
Les mécanismes par lesquels l'administration d'interféron alpha provoque des effets secondaires neuropsychiatriques comme des symptômes de dépression et des changements de la fonction cognitive ne sont pas encore clairs. L'influence directe sur la neurotransmission sérotoninergique peut contribuer à ces effets secondaires. La 2,3-dioxygénase (IDO), enzyme qui convertit le tryptophane en cynurénine, peut en outre jouer un rôle important tout d'abord parce que l'activation de l'IDO entraÎne une réduction des concentrations de tryptophane, précurseur de la sérotonine (5-HT) et, par conséquent, une baisse de la synthèse centrale de 5-HT. De plus, des métabolites de la cynurénine, comme la 3-hydroxy-cynurénine (3-OH-KYN) et l'acide quinolinique (QUIN), ont des effets toxiques sur la fonction cérébrale. La 3-OH-KYN peut produire un stress oxydatif en élevant la production d'espèces réagissant à l'oxygène (ERO) et la QUIN peut produire une surstimulation des récepteurs de la N-méthyl-D-aspartate (NMDA) de l'hippocampe, qui entraÎne l'apoptose cellulaire et une atrophie de l'hippocampe. La surproduction d'ERO et l'atrophie de l'hippocampe causée par la surstimulation de la NMDA ont toutes deux été associées à la dépression.
Introduction
The proinflammatory cytokine interferon-!#!alpha; (IFN-α) is commonly used in the treatment of patients with hepatitis C and cancer, but its administration induces neuropsychiatric side effects.1,2 Symptoms frequently associated with IFN-α treatment are fatigue, both increased sleepiness and difficulty sleeping, irritability, loss of appetite, weight loss and low mood. A full-blown depressive disorder is reported in up to 36% of cases.3 Cognitive changes involving verbal memory, cognitive speed and executive function are also reported.4,5,6
Administration of proinflammatory cytokines, such as interleukin-1β (IL-1β), IL-6, IFN-α and tumour necrosis factor-α (TNF-α), in rats or mice induces a behavioural pattern referred to as “sickness behaviour,” 7 which includes increased sleep,8 reduced locomotor activity,9,10 decreased sucrose consumption (as a measure for anhedonia),11 weight loss and decreased social exploration.12 These symptoms resemble the vegetative symptoms of depression in humans.2","13
The development of sickness behaviour14 and depression15","16 is linked to dysregulation of the neurotransmitter serotonin (5-HT). Several neurochemical changes in both the peripheral and central 5-HT system are seen in depressed patients, such as lower levels of peripheral tryptophan (TRP), the precursor of 5-HT;17 changes in central 5-HT transporter (5-HTT) function;18 and changes in 5-HT1A19 and 5-HT2A brain receptors.20 Therefore, one way in which IFN-α may induce depressive symptoms is by affecting the serotonergic system.
IFN-α and central 5-HT neurotransmission
IFN-α exerts direct influence on 5-HT brain neurotransmission. IFN-α and other proinflammatory cytokines, such as IFN-γ, TNF-α and IL-1, have been shown to upregulate 5-HTT, causing a decrease of extracellular 5-HT.21,22,23 IFN-αmay also indirectly influence 5-HTT activity by increasing the production of IFN-γ, TNF-α and IL-1. In addition, IFN-α modulates the 5-HT1A24 and 5-HT225 brain receptors.
IDO activity causes TRP depletion
A second pathway by which IFN-α modulates the 5-HT system is by induction of the enzyme indoleamine 2,3-dioxygenase (IDO). IDO is the rate-limiting enzyme in the L-TRP–kynurenine pathway that converts L-TRP, the precursor of 5-HT, to N-formylkynurenine, resulting in a diminished synthesis of central 5-HT. IDO is widely distributed in various tissues, including the brain, lung, heart, kidney and intestine.26
Cytokines, such as IFN-α, IFN-β, TNF-α and IFN-γ, have been shown to upregulate IDO expression.26,27,28,29,30,31 IFN-α has a weak direct effect on IDO induction and an indirect effect through a 15-kD protein, which is a product of IFN-α-stimulated monocytes and lymphocytes and stimulates IDO and IFN-γ production.32
IFN-α therapy in patients with hepatitis C causes a decrease in TRP (4–6 months after starting therapy) and an increase in kynurenine plasma levels seen at 2 weeks after starting therapy, with kynurenine plasma levels remaining the same as at 2 weeks when measured at weeks 4, 16 and 24, indicating higher IDO activity.33 In addition, IDO induction is also mediated by an IFN-γ-independent mechanism involving the proinflammatory cytokine TNF-α.26,6 Anti-inflammatory cytokines, such as IL-4 and IL-10, diminish TRP metabolism.34
Overstimulation of IDO leads to depletion of plasma concentrations of TRP and, therefore, to reduced synthesis of 5-HT in the brain,35 which may play a role in the development of depressive symptoms. In addition, not only TRP but also 5-hydroxytryptophan (5-HTP) and 5-HT itself can be substrates for IDO.36 Therefore, it can be postulated that in addition to lowering peripheral levels of TRP, 5-HT synthesis in the brain can also be reduced by central degradation of 5-HT by IDO. However, there is another possible way in which IFN-α-induced overstimulation of IDO may produce depressive symptoms. By upregulating IDO expression, IFN-α can initiate the kynurenine pathway leading to the production of a variety of neuroactive metabolites. These kynurenine metabolites themselves may play a causative role, because the ratio of kynurenine to TRP is positively associated with depression and anxiety scores.33,37
Neurotoxic metabolites of the kynurenine pathway
Several metabolites formed along the kynurenine pathway are found to have neurotoxic effects, such as 3-hydroxy-kynurenine (3-OH-KYN), a direct metabolite of kynurenine, and quinolinic acid (QUIN), which is formed later in the kynurenine pathway.38,39,40,41,42 Peripheral kynurenine is transported through the blood–brain barrier by a large neutral amino acid carrier and thus may easily reach the central nervous system. In the brain it is taken up by glia cells, by which it is further metabolized.43,44 In this way, neurotoxic metabolites are formed in the brain that can cause neurodegeneration.
The neurotoxicity of these kynurenine metabolites has been demonstrated in animals45,46,47,48,49,50 and in vitro,41 whereas data for humans show elevated levels in several degenerative disorders. Increased production of 3-OH-KYN or QUIN, or both, is found in certain neurodegenerative conditions in humans,38 such as in Huntington's disease,51,52 Parkinson's disease53 and in the AIDS–dementia complex.54,55 In the last condition, QUIN levels are increased in the cerebrospinal fluid up to 20-fold and are correlated with the severity of cognitive and motor dysfunctions. Increased production of 3-OH-KYN and QUIN may also contribute to neuronal damage in cognitive decline of aging,56 infections of the central nervous system,57 malaria,58 ischemia,59 hypoxia at birth,60 traumatic injury61 and epilepsy.62 In addition, they may play a role in the development of psychiatric diseases such as anxiety,63 depression64 and schizophrenia.65
The following mechanisms may account for the effect of 3-OH-KYN and QUIN on neurodegenerative diseases and depressive symptoms (Fig. 1). Even relatively low levels of 3-OH-KYN may cause neurotoxicity by inducing oxidative stress and neuronal apoptosis.40,41 3-OH-KYN is transferred into cells by neutral amino acid transporters. Only after interaction with cellular xanthine oxidase is 3-OH-KYN capable of producing sufficient amounts of reactive oxygen species (ROS), such as superoxide radical, hydrogen peroxide and hydroxyl radical, to induce internucleosomal DNA cleavage leading to apoptosis. Various antioxidative agents prevent cell death induced by 3-OH-KYN. The cortex and the striatum are most sensitive to 3-OH-KYN insults. Differences in vulnerability to 3-OH-KYN for different brain regions are likely to result from differences in their ability to take up large neutral amino acids.41 Overproduction of ROS has been associated with depression. An association between overproduction of ROS and increased monoamine oxidase (MAO) activity has been suggested.66 In addition, polyunsaturated fatty acids (PUFAs) are vulnerable to oxidation. Overproduction of ROS might result in destruction of phospholipids and reduce the viscosity of cell membranes.67 Alterations in membrane viscosity may influence receptor density or function of serotonergic or catecholaminergic receptors.68 Overproduction of proinflammatory cytokines, increased MAO activity and, thus, lower levels of catecholamines, disturbances in PUFA structures and ratios, and the decrement of serotonergic and catecholaminergic receptor densities and functioning are all associated with depression.2,69,70 Furthermore, selective serotonin reuptake inhibitors have antioxidant properties and reverse the overproduction of ROS.71
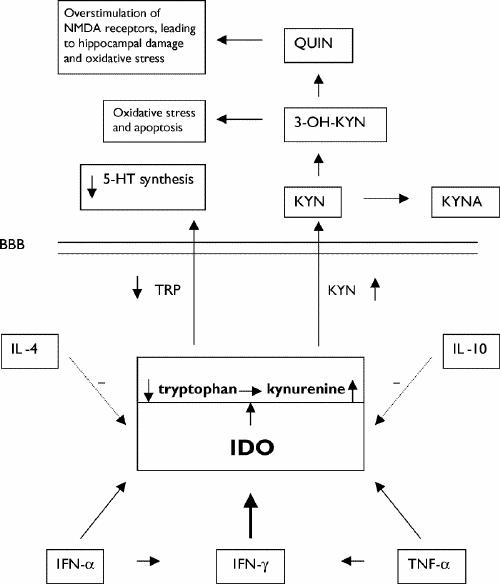
Fig. 1: The role of indoleamine 2,3-dioxygenase (IDO) in cytokine-induced lowered tryptophan (TRP) levels and production of neurotoxic metabolites. The dotted arrows indicate that IL-4 and IL-10 diminish IDO activity. The diagonal arrows leading from IFN-α and TNF-α to IDO indicate that these cytokines increase IDO activity and, thus, cause increased metabolism of TRP, resulting in decreased availability of TRP that crosses the BBB to synthesize 5-HT. The horizontal arrows between IFN-α and IFN-γ and between TNF-α and IFN-γ refer to the fact that IFN-α and TNF-α increase production of IFN-γ, which exerts a powerful stimulating effect (thick vertical arrow) on IDO activity. NMDA = N-methyl-D-aspartate, QUIN = quinolinic acid, 3-OH-KYN =3-hydroxy-kynurenine, 5-HT = serotonin, KYN = kynurenine, KYNA = kynurenate, BBB = blood–brain barrier, IL-4 = interleukin-4, IFN-α = interferon-α, TNF-α = tumour necrosis factor-α.
QUIN is a potent N-methyl-D-aspartate (NMDA) receptor agonist. Overstimulation of NMDA receptors increases calcium influx into the target neurons, which will lead to neuronal damage.47,72 In addition, QUIN can contribute to the formation of free radicals, first, because the influx of calcium ions into neurons after activation of glutamate receptors may lead to the generation of ROS.73,74 Furthermore, with the generation of free radicals, QUIN induces lipid peroxidation in membrane lipids and proteins, leading to changes in neuronal membrane fluidity, receptor function and ion permeability.75 Finally, QUIN and 3-OH-KYN act synergistically in the production of free radicals, so that even low doses of QUIN potentiate the excitotoxicity of 3-OH-KYN.76,77
Intrahippocampal injection of QUIN in rats causes substantial loss of hippocampal neurons.45,48,75 QUIN-induced neurotoxicity can be significantly reduced by pharmacologic blockade of kynurenine 3-hydroxylase, the enzyme responsible for the formation of 3-OH-KYN from L-kynurenine or by administration of kynurenate (KYNA), another metabolite of the kynurenine pathway, which inhibits NMDA receptor function and thus protects against excitotoxic insults.48
Hippocampal neurodegeneration and depression
Major depression is associated with hippocampal volume loss.78,79 Hypothalamic-pituitary-adrenal (HPA) axis inhibition appears to be mediated by negative feedback from circulating glucocorticoids.80 Destruction of the hippocampus attenuates the negative feedback via loss of glucocorticoid receptors, which will increase HPA activity. Loss of the normal glucocorticoid feedback has been reported in humans with depression and animals in chronic stress paradigms.81,82,83 Hippocampal atrophy results from an excess of excitatory amino acid neurotransmitters, such as glutamate that acts on the NMDA receptor, which can be produced by glucocorticoid overexposure.84,85,86 However, Magariños and McEwen87 found a dissociation between parameters indicating the glucocorticoid stress response and hippocampal atrophy in rats. Dendritic atrophy occurred in spite of the habituation of the glucocorticoid stress response after days of repeated stress. In addition, hypercortisolism occurs in about half of the patients with depression, whereas atrophy seems to be demonstrable in a far higher percentage of individuals.86 This suggests that, besides glucocorticoids, other factors contribute to neurodegeneration, for example, the neurotoxic metabolites of kynurenine, whose levels are raised in depression. Therefore, we assume as a hypothesis a second mechanism in which kynurenine metabolites, such as QUIN, cause NMDA receptor overstimulation, leading to hippocampal atrophy and subsequent interference with the normal negative feedback function of HPA axis activity.
Conclusion
Administration of IFN-α causes the development of depressive symptoms in a high percentage of patients. This may be caused by direct influence of IFN-α on central 5-HT transmission, or indirectly via induction of the enzyme IDO. The latter may play an important role in the pathophysiology of IFN-α-induced depression through its effect on brain TRP availability that is crucial to the formation of central 5-HT. Furthermore, IDO is the rate-limiting step in the brain kynurenine pathway that leads to the formation of neurotoxic substances, such as 3-OH-KYN and QUIN, which cause neurodegeneration that may contribute to the development of depression.
Footnotes
Competing interests: None declared.
Correspondence to: Ms. M.C. Wichers, Department of Psychiatry and Neuropsychology, Maastricht University, PO Box 616, 6200 MD Maastricht, the Netherlands; fax 31 43 3299708; m.wichers@sp.unimaas.nl
Submitted Oct. 29, 2002; Revised Mar. 6, 2003; Accepted Mar. 17, 2003
References
- 1.Bonaccorso S, Marino V, Biondi M, Grimaldi F, Ippoliti F, Maes M. Depression induced by treatment with interferon- alpha in patients affected by hepatitis C virus. J Affect Disord 2002; 72(3):237-41. [DOI] [PubMed]
- 2.Maes M. Evidence for an immune response in major depression: a review and hypothesis. Prog Neuropsychopharmacol Biol Psychiatry 1995;19(1):11-38. [DOI] [PubMed]
- 3.Collier J, Chapman R. Combination therapy with interferon- alpha and ribavirin for hepatitis C: practical treatment issues. BioDrugs 2001;15(4):225-38. [DOI] [PubMed]
- 4.Juengling FD, Ebert D, Gut O, Engelbrecht MA, Rasenack J, Nitzsche EU, et al. Prefrontal cortical hypometabolism during low-dose interferon alpha treatment. Psychopharmacology (Berl) 2000; 152(4):383-9. [DOI] [PubMed]
- 5.Pavol MA, Meyers CA, Rexer JL, Valentine AD, Mattis PJ, Talpaz M. Pattern of neurobehavioral deficits associated with interferon alfa therapy for leukemia. Neurology 1995;45(5):947-50. [DOI] [PubMed]
- 6.Adams F, Quesada JR, Gutterman JU. Neuropsychiatric manifestations of human leukocyte interferon therapy in patients with cancer. JAMA 1984;252(7):938-41. [PubMed]
- 7.Kent S, Bluthe RM, Kelley KW, Dantzer R. Sickness behavior as a new target for drug development. Trends Pharmacol Sci 1992; 13(1):24-8. [DOI] [PubMed]
- 8.Krueger JM, Takahashi S, Kapas L, Bredow S, Roky R, Fang J, et al. Cytokines in sleep regulation. Adv Neuroimmunol 1995; 5 (2): 171-88. [DOI] [PubMed]
- 9.Lacosta S, Merali Z, Anisman H. Influence of interleukin-1beta on exploratory behaviors, plasma ACTH, corticosterone, and central biogenic amines in mice. Psychopharmacology (Berl) 1998;137(4):351-61. [DOI] [PubMed]
- 10.Bianchi M, Sacerdote P, Ricciardi-Castagnoli P, Mantegazza P, Panerai AE. Central effects of tumor necrosis factor alpha and interleukin-1 alpha on nociceptive thresholds and spontaneous locomotor activity. Neurosci Lett 1992;148(1-2):76-80. [DOI] [PubMed]
- 11.Sammut S, Goodall G, Muscat R. Acute interferon-alpha administration modulates sucrose consumption in the rat. Psychoneuroendocrinology 2001;26(3):261-72. [DOI] [PubMed]
- 12.Bluthe RM, Pawlowski M, Suarez S, Parnet P, Pittman Q, Kelley KW, et al. Synergy between tumor necrosis factor alpha and interleukin-1 in the induction of sickness behavior in mice. Psychoneuroendocrinology 1994;19(2):197-207. [DOI] [PubMed]
- 13.Charlton BG. The malaise theory of depression: major depressive disorder is sickness behavior and antidepressants are analgesic. Med Hypotheses 2000;54(1):126-30. [DOI] [PubMed]
- 14.Linthorst AC, Reul JM. Brain neurotransmission during peripheral inflammation. Ann N Y Acad Sci 1998;840:139-52. [DOI] [PubMed]
- 15.Dursun SM, Blackburn JR, Kutcher SP. An exploratory approach to the serotonergic hypothesis of depression: bridging the synaptic gap. Med Hypotheses 2001;56(2):235-43. [DOI] [PubMed]
- 16.Graeff FG, Guimaraes FS, De Andrade TG, Deakin JF. Role of 5-HT in stress, anxiety, and depression. Pharmacol Biochem Behav 1996; 54(1):129-41. [DOI] [PubMed]
- 17.Maes M, Meltzer HY, Scharpe S, Bosmans E, Suy E, De Meester I, et al. Relationships between lower plasma L-tryptophan levels and immune-inflammatory variables in depression. Psychiatry Res 1993;49(2):151-65. [DOI] [PubMed]
- 18.Lesch KP, Mossner R. Genetically driven variation in serotonin uptake: Is there a link to affective spectrum, neurodevelopmental, and neurodegenerative disorders? Biol Psychiatry 1998; 44(3):179-92. [DOI] [PubMed]
- 19.Sargent PA, Kjaer KH, Bench CJ, Rabiner EA, Messa C, Meyer J, et al. Brain serotonin1A receptor binding measured by positron emission tomography with [11C]WAY-100635: effects of depression and antidepressant treatment. Arch Gen Psychiatry 2000; 57 (2):174-80. [DOI] [PubMed]
- 20.Yatham LN, Liddle PF, Shiah IS, Scarrow G, Lam RW, Adam MJ, et al. Brain serotonin2 receptors in major depression: a positron emission tomography study. Arch Gen Psychiatry 2000; 57(9):850-8. [DOI] [PubMed]
- 21.Ramamoorthy S, Ramamoorthy JD, Prasad PD, Bhat GK, Mahesh VB, Leibach FH, et al. Regulation of the human serotonin transporter by interleukin-1 beta. Biochem Biophys Res Commun 1995; 216(2):560-7. [DOI] [PubMed]
- 22.Mossner R, Heils A, Stober G, Okladnova O, Daniel S, Lesch KP. Enhancement of serotonin transporter function by tumor necrosis factor alpha but not by interleukin-6. Neurochem Int 1998; 33(3):251-4. [DOI] [PubMed]
- 23.Morikawa O, Sakai N, Obara H, Saito N. Effects of interferon-alpha, interferon-gamma and cAMP on the transcriptional regulation of the serotonin transporter. Eur J Pharmacol 1998; 349 (2-3): 317-24. [DOI] [PubMed]
- 24.Abe S, Hori T, Suzuki T, Baba A, Shiraishi H, Yamamoto T. Effects of chronic administration of interferon alpha A/D on serotonergic receptors in rat brain. Neurochem Res 1999;24 (3): 359-63. [DOI] [PubMed]
- 25.Kugaya A, Kagaya A, Uchitomi Y, Yokota N, Yamawaki S. Effect of interferon-alpha on DOI-induced wet-dog shakes in rats. J Neural Transm 1996;103(8-9):947-55. [DOI] [PubMed]
- 26.Fujigaki S, Saito K, Sekikawa K, Tone S, Takikowa O, Fujii H, et al. Lipopolysaccharide induction of indoleamine 2,3-dioxygenase is mediated dominantly by an IFN-gamma-independent mechanism. Eur J Immunol 2001;31(8):2313-8. [DOI] [PubMed]
- 27.Takikawa O, Kuroiwa T, Yamazaki F, Kido R. Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. J Biol Chem 1988; 263 (4): 2041-8. [PubMed]
- 28.Pemberton LA, Kerr SJ, Smythe G, Brew BJ. Quinolinic acid production by macrophages stimulated with IFN-gamma, TNF-alpha, and IFN-alpha. J Interferon Cytokine Res 1997;17 (10): 589-95. [DOI] [PubMed]
- 29.Jansen M, Reinhard JF Jr. Interferon response heterogeneity: activation of a pro-inflammatory response by interferon alpha and beta. A possible basis for diverse responses to interferon beta in MS. J Leukoc Biol 1999;65(4):439-43. [DOI] [PubMed]
- 30.Taylor MW, Feng GS. Relationship between interferon-gamma, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J 1991;5(11):2516-22. [PubMed]
- 31.Guillemin GJ, Kerr SJ, Pemberton LA, Smith DG, Smythe GA, Armati PJ, et al. IFN-beta1b induces kynurenine pathway metabolism in human macrophages: potential implications for multiple sclerosis treatment. J Interferon Cytokine Res 2001; 21 (12): 1097-101. [DOI] [PubMed]
- 32.Recht M, Borden EC, Knight E Jr. A human 15-kDa IFN- induced protein induces the secretion of IFN-gamma. J Immunol 1991; 147(8):2617-23. [PubMed]
- 33.Bonaccorso S, Marino V, Puzella A, Pasquini M, Biondi M, Artini M, et al. Increased depressive ratings in patients with hepatitis C receiving interferon-alpha-based immunotherapy are related to interferon-alpha-induced changes in the serotonergic system. J Clin Psychopharmacol 2002;22(1):86-90. [DOI] [PubMed]
- 34.Chiarugi A, Calvani M, Meli E, Traggiai E, Moroni F. Synthesis and release of neurotoxic kynurenine metabolites by human monocyte-derived macrophages. J Neuroimmunol 2001; 120 (1-2):190-8. [DOI] [PubMed]
- 35.Heyes MP, Saito K, Crowley JS, Davis LE, Demitrack MA, Der M, et al. Quinolinic acid and kynurenine pathway metabolism in inflammatory and non-inflammatory neurological disease. Brain 1992;115(Pt 5):1249-73. [DOI] [PubMed]
- 36.Stone TW, Darlington LG. Endogenous kynurenines as targets for drug discovery and development. Nat Rev Drug Discov 2002; 1 (8):609-20. [DOI] [PubMed]
- 37.Maes M, Verkerk R, Bonaccorso S, Ombelet W, Bosmans E, Scharpe S. Depressive and anxiety symptoms in the early puerperium are related to increased degradation of tryptophan into kynurenine, a phenomenon which is related to immune activation. Life Sci 2002;71(16):1837. [DOI] [PubMed]
- 38.Stone TW. Kynurenines in the CNS: from endogenous obscurity to therapeutic importance. Prog Neurobiol 2001;64(2):185-218. [DOI] [PubMed]
- 39.Dang Y, Dale WE, Brown OR. Comparative effects of oxygen on indoleamine 2,3-dioxygenase and tryptophan 2,3-dioxygenase of the kynurenine pathway. Free Radic Biol Med 2000;28 (4): 615-24. [DOI] [PubMed]
- 40.Moroni F. Tryptophan metabolism and brain function: focus on kynurenine and other indole metabolites. Eur J Pharmacol 1999; 375(1-3):87-100. [DOI] [PubMed]
- 41.Okuda S, Nishiyama N, Saito H, Katsuki H. 3-Hydroxykynurenine, an endogenous oxidative stress generator, causes neuronal cell death with apoptotic features and region selectivity. J Neurochem 1998;70(1):299-307. [DOI] [PubMed]
- 42.Guillemin GJ, Kerr SJ, Smythe GA, Smith DG, Kapoor V, Armati PJ, et al. Kynurenine pathway metabolism in human astrocytes: a paradox for neuronal protection. J Neurochem 2001; 78 (4):842-53. [DOI] [PubMed]
- 43.Speciale C, Hares K, Schwarcz R, Brookes N. High-affinity uptake of L-kynurenine by a Na+-independent transporter of neutral amino acids in astrocytes. J Neurosci 1989;9(6):2066-72. [DOI] [PMC free article] [PubMed]
- 44.Fukui S, Schwarcz R, Rapoport SI, Takada Y, Smith QR. Blood-brain barrier transport of kynurenines: implications for brain synthesis and metabolism. J Neurochem 1991;56(6):2007-17. [DOI] [PubMed]
- 45.Behan WM, McDonald M, Darlington LG, Stone TW. Oxidative stress as a mechanism for quinolinic acid-induced hippocampal damage: protection by melatonin and deprenyl. Br J Pharmacol 1999;128(8):1754-60. [DOI] [PMC free article] [PubMed]
- 46.Qin ZH, Wang Y, Chase TN. Stimulation of N-methyl-D- aspartate receptors induces apoptosis in rat brain. Brain Res 1996; 725(2):166-76. [DOI] [PubMed]
- 47.Schwarcz R, Whetsell WO Jr, Mangano RM. Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science 1983;219(4582):316-8. [DOI] [PubMed]
- 48.Wu HQ, Guidetti P, Goodman JH, Varasi M, Ceresoli-Borroni G, Speciale C, et al. Kynurenergic manipulations influence excitatory synaptic function and excitotoxic vulnerability in the rat hippocampus in vivo. Neuroscience 2000;97(2):243-51. [DOI] [PubMed]
- 49.Guidetti P, Schwarcz R. 3-Hydroxykynurenine potentiates quinolinate but not NMDA toxicity in the rat striatum. Eur J Neurosci 1999;11(11):3857-63. [DOI] [PubMed]
- 50.Heyes MP, Jordan EK, Lee K, Saito K, Frank JA, Snoy PJ, et al. Relationship of neurologic status in macaques infected with the simian immunodeficiency virus to cerebrospinal fluid quinolinic acid and kynurenic acid. Brain Res 1992;570(1-2):237-50. [DOI] [PubMed]
- 51.Reynolds GP, Pearson SJ. Increased brain 3-hydroxykynurenine in Huntington's disease. Lancet 1989;2(8669):979-80. [DOI] [PubMed]
- 52.Reynolds GP, Pearson SJ, Halket J, Sandler M. Brain quinolinic acid in Huntington's disease. J Neurochem 1988;50(6):1959-60. [DOI] [PubMed]
- 53.Ogawa T, Matson WR, Beal MF, Myers RH, Bird ED, Milbury P, et al. Kynurenine pathway abnormalities in Parkinson's disease. Neurology 1992;42(9):1702-6. [DOI] [PubMed]
- 54.Heyes MP, Saito K, Lackner A, Wiley CA, Achim CL, Markey SP. Sources of the neurotoxin quinolinic acid in the brain of HIV-1-infected patients and retrovirus-infected macaques. FASEB J 1998;12(10):881-96. [DOI] [PubMed]
- 55.Chao CC, Hu S, Gekker G, Lokensgard JR, Heyes MP, Peterson PK. U50,488 protection against HIV-1-related neurotoxicity: involvement of quinolinic acid suppression. Neuropharmacology 2000;39(1):150-60. [DOI] [PubMed]
- 56.Moroni F, Lombardi G, Moneti G, Aldinio C. The excitotoxin quinolinic acid is present in the brain of several mammals and its cortical content increases during the aging process. Neurosci Lett 1984;47(1):51-5. [DOI] [PubMed]
- 57.Heyes MP, Saito K, Milstien S, Schiff SJ. Quinolinic acid in tumors, hemorrhage and bacterial infections of the central nervous system in children. J Neurol Sci 1995;133(1-2):112-8. [DOI] [PubMed]
- 58.Sanni LA, Thomas SR, Tattam BN, Moore DE, Chaudhri G, Stocker R, et al. Dramatic changes in oxidative tryptophan metabolism along the kynurenine pathway in experimental cerebral and noncerebral malaria. Am J Pathol 1998;152(2):611-9. [PMC free article] [PubMed]
- 59.Saito K, Nowak TS Jr, Markey SP, Heyes MP. Mechanism of delayed increases in kynurenine pathway metabolism in damaged brain regions following transient cerebral ischemia. J Neurochem 1993;60(1):180-92. [DOI] [PubMed]
- 60.Kazda H, Taylor N, Healy D, Walker D. Maternal, umbilical, and amniotic fluid concentrations of tryptophan and kynurenine after labor or cesarean section. Pediatr Res 1998;44(3):368-73. [DOI] [PubMed]
- 61.Sinz EH, Kochanek PM, Heyes MP, Wisniewski SR, Bell MJ, Clark RS, et al. Quinolinic acid is increased in CSF and associated with mortality after traumatic brain injury in humans. J Cereb Blood Flow Metab 1998;18(6):610-5. [DOI] [PubMed]
- 62.Heyes MP, Wyler AR, Devinsky O, Yergey JA, Markey SP, Nadi NS. Quinolinic acid concentrations in brain and cerebrospinal fluid of patients with intractable complex partial seizures. Epilepsia 1990;31(2):172-7. [DOI] [PubMed]
- 63.Orlikov AB, Prakhye IB, Ryzov IV. Kynurenine in blood plasma and DST in patients with endogenous anxiety and endogenous depression. Biol Psychiatry 1994;36(2):97-102. [DOI] [PubMed]
- 64.Mangoni A. The “kynurenine shunt” and depression. Adv Biochem Psychopharmacol 1974;11:293-8. [PubMed]
- 65.Issa F, Kirch DG, Gerhardt GA, Bartko JJ, Suddath RL, Freedman R, et al. A multidimensional approach to analysis of cerebrospinal fluid biogenic amines in schizophrenia: II. Correlations with psychopathology. Psychiatry Res 1994;52(3):251-8. [DOI] [PubMed]
- 66.Gutteridge JM. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin Chem 1995;41(12 Pt 2):1819-28. [PubMed]
- 67.McIntyre N. Familial LCAT deficiency and fish-eye disease. J Inherit Metab Dis 1988;11(Suppl 1):45-56. [DOI] [PubMed]
- 68.Van der Vliet A, Bast A. Effect of oxidative stress on receptors and signal transmission. Chem Biol Interact 1992;85(2-3):95-116. [DOI] [PubMed]
- 69.Maes M, Meltzer H. The serotonin hypothesis of major depression. In: Bloom F, Kupfer D, editors. Psychopharmacology. New York: Raven Press; 1995: p. 933-44.
- 70.Maes M, Smith R, Christophe A, Cosyns P, Desnyder R, Meltzer H. Fatty acid composition in major depression: decreased omega 3 fractions in cholesteryl esters and increased C20: 4 omega 6/C20:5 omega 3 ratio in cholesteryl esters and phospholipids. J Affect Disord 1996;38(1):35-46. [DOI] [PubMed]
- 71.Bilici M, Efe H, Koroglu MA, Uydu HA, Bekaroglu M, Deger O. Antioxidative enzyme activities and lipid peroxidation in major depression: alterations by antidepressant treatments. J Affect Disord 2001;64(1):43-51. [DOI] [PubMed]
- 72.Perkins MN, Stone TW. An iontophoretic investigation of the actions of convulsant kynurenines and their interaction with the endogenous excitant quinolinic acid. Brain Res 1982;247(1):184-7. [DOI] [PubMed]
- 73.Hartley DM, Kurth MC, Bjerkness L, Weiss JH, Choi DW. Glutamate receptor-induced 45Ca2+ accumulation in cortical cell culture correlates with subsequent neuronal degeneration. J Neurosci 1993;13(5):1993-2000. [DOI] [PMC free article] [PubMed]
- 74.Atlante A, Gagliardi S, Minervini GM, Ciotti MT, Marra E, Calissano P. Glutamate neurotoxicity in rat cerebellar granule cells: a major role for xanthine oxidase in oxygen radical formation. J Neurochem 1997;68(5):2038-45. [DOI] [PubMed]
- 75.Santamaria A, Galvan-Arzate S, Lisy V, Ali SF, Duhart HM, Osorio-Rico L, et al. Quinolinic acid induces oxidative stress in rat brain synaptosomes. Neuroreport 2001;12(4):871-4. [DOI] [PubMed]
- 76.Guillemin GJ, Brew BJ. Implications of the kynurenine pathway and quinolinic acid in Alzheimer's disease. Redox Rep 2002; 7 (4):199-206. [DOI] [PubMed]
- 77.Nakagami Y, Saito H, Katsuki H. 3-Hydroxykynurenine toxicity on the rat striatum in vivo. Jpn J Pharmacol 1996;71(2):183-6. [DOI] [PubMed]
- 78.Sheline YI, Sanghavi M, Mintun MA, Gado MH. Depression duration but not age predicts hippocampal volume loss in medically healthy women with recurrent major depression. J Neurosci 1999;19(12):5034-43. [DOI] [PMC free article] [PubMed]
- 79.Bremner JD, Narayan M, Anderson ER, Staib LH, Miller HL, Charney DS. Hippocampal volume reduction in major depression. Am J Psychiatry 2000;157(1):115-8. [DOI] [PubMed]
- 80.Sapolsky RM, Meaney MJ, McEwen BS. The development of the glucocorticoid receptor system in the rat limbic brain. III. Negative-feedback regulation. Brain Res 1985;350(1-2):169-73. [DOI] [PubMed]
- 81.Sapolsky RM, Krey LC, McEwen BS. The adrenocortical axis in the aged rat: impaired sensitivity to both fast and delayed feedback inhibition. Neurobiol Aging 1986;7(5):331-5. [DOI] [PubMed]
- 82.Liberzon I, Krstov M, Young EA. Stress-restress: effects on ACTH and fast feedback. Psychoneuroendocrinology 1997;22(6): 443-53. [DOI] [PubMed]
- 83.Young EA, Haskett RF, Murphy-Weinberg V, Watson SJ, Akil H. Loss of glucocorticoid fast feedback in depression. Arch Gen Psychiatry 1991;48(8):693-9. [DOI] [PubMed]
- 84.McEwen BS. Effects of adverse experiences for brain structure and function. Biol Psychiatry 2000;48(8):721-31. [DOI] [PubMed]
- 85.McEwen BS. Possible mechanisms for atrophy of the human hippocampus. Mol Psychiatry 1997;2(3):255-62. [DOI] [PubMed]
- 86.Sapolsky RM. Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch Gen Psychiatry 2000;57:925-35. [DOI] [PubMed]
- 87.Magariños AM, McEwen BS. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: comparison of stressors. Neuroscience 1995;69(1):83-8. [DOI] [PubMed]