

Abstract
Alterations in brain serotonin (5-hydroxytryptamine [5-HT]) function are thought to contribute to diverse aspects of eating disorders, including binge eating, perfectionism, impulsivity and mood-regulation problems. In addition, 5-HT anomalies in individuals with eating disorders are believed to have multiple determinants associated with secondary (state-related) effects of their nutritional status, hereditary effects (related to such trait variations as impulsivity or perfectionism) and, possibly, long-term neurobiologic sequelae of developmental stressors (such as childhood abuse). On the strength of the available neurobiologic and genetic data, this paper presents the idea that 5-HT variations in those with eating disorders represent (1) a structured coaggregation of biologic, psychologic and social influences and (2) converging state, trait and developmental effects. Data are taken to support a multidimensional model of 5-HT function in eating disorders that, it is argued, can serve as a prototype for etiologic modelling, diagnostic classification and clinical decision-making bearing not only upon eating disorders but also upon other psychiatric disturbances.
Medical subject headings: anorexia nervosa, bulimia, eating disorders, serotonin
Abstract
On croit que des altérations de la fonction sérotoninergique (5-hydroxytryptamine [5-HT]) dans le cerveau contribuent à divers aspects des troubles de l'alimentation, y compris les épisodes de frénésie alimentaire, le perfectionnisme, l'impulsivité et des problèmes de régulation de l'humeur. On croit en outre que des anomalies de la 5-HT chez des personnes ayant des troubles de l'alimentation comportent de multiples déterminants associés aux effets secondaires (reliés à l'état) de leur état nutritif, à des effets héréditaires (reliés à des variations de caractéristiques comme l'impulsivité ou le perfectionnisme) et, peut-être, à des séquelles neurobiologiques de longue date découlant de facteurs de stress du développement (comme la violence au cours de l'enfance). En se fondant sur la solidité des données neurobiologiques et génétiques disponibles, l'auteur de cette communication énonce l'idée selon laquelle les variations de la 5-HT chez des personnes souffrant de troubles de l'alimentation représentent (1) une coaggréation structurée d'influences biologiques, psychologiques et sociales et (2) une convergence d'effets liés à l'état, aux caractéristiques et au développement. On a réuni des données pour appuyer un modèle multidimensionnel de la fonction de la 5-HT dans les cas de troubles de l'alimentation. On soutient que ce modèle peut servir de prototype de modélisation étiologique, de classification diagnostique et de prise de décisions cliniques portant non seulement sur des troubles de l'alimentation, mais aussi sur d'autres troubles psychiatriques.
Introduction
The eating disorders (EDs) anorexia nervosa and bulimia nervosa are symptomatically complex syndromes, defined by maladaptive attitudes and behaviours around eating, weight and body image, but invariably including “nonspecific” disturbances (of self-image, mood, impulse regulation and interpersonal functioning). Befitting such phenomenologic complexity, contemporary etiologic models of EDs generally invoke a multidimensional (biopsychosocial) causality,1,2,3,4 postulating a causal “collision" among biologic factors (e.g., heritable influences on appetite, mood, temperament and impulse controls), social pressures (promoting body consciousness or generalized self-definition problems), psychologic tendencies (autonomy disturbances, perfectionism, preference for order and control) and developmental processes (conducive to self-image or adjustment problems). Although such formulations are comprehensive, to be fully successful they need to trace the specific convergences of biologic, psychologic and social forces that produce clinical ED syndromes.
In this paper, I apply findings from serotonin (5- hydroxytryptamine [5-HT]) studies as a heuristic toward the development of a multidimensional model of ED pathology. My thinking is based on the likelihood that the momentary 5-HT status in a given individual (especially when she has an active ED) represents diverse contributing processes: transient effects of dietary factors (states), stable effects related to behavioural traits (like impulsivity or perfectionism) and sequelae of adverse developmental experiences (such as childhood traumas). (Not to disregard the existence of EDs in males, their disproportionate representation among women and girls has caused me to opt for female personal pronouns.) Before proceeding, however, I think that some readers may want some general background on the EDs.
ED classification
The current Diagnostic and Statistical Manual of Mental Disorders, fourth edition (DSM-IV),5 nosology recognizes 2 official ED forms: anorexia nervosa and bulimia nervosa. The principal symptom of anorexia nervosa is self-imposed starvation, typically arising from a morbid fear of becoming fat. “Phobic” avoidance of weight gain motivates the patient to avoid eating, with the consequence that she gradually (and often quite unwittingly) renders herself too thin. It is important to highlight, however, that despite the meaning of its name in Latin, anorexia nervosa implies no true “loss of appetite.” On the contrary, people with anorexia nervosa typically become fully preoccupied with “things appetite related.” This suggests that anorexia nervosa is best construed as an anxious avoidance of weight gain, obsession with weight and shape, and compulsion surrounding control of eating and weight, namely, as more a disorder of “anxiety” and “impulse regulation” than of “feeding.” This point has conceptual implications that will soon become obvious.
Symptomatology in bulimia nervosa is characterized by recurrent binge eating and frantic compensatory behaviours (such as vomiting, laxative misuse or compulsive exercise). The dietary dyscontrol that characterizes bulimia nervosa appears, superficially, to be antithetical to the overcontrol found in anorexia nervosa. However, as in anorexia nervosa, the “engine” that drives bulimia nervosa is often compulsive dieting, with prolonged cognitive dietary restraint being understood to potentiate eventual counter-regulation, or disinhibition, of eating behaviour.6 The progression to binge eating in only some individuals with EDs has motivated some theorists to postulate a distinction between “restricters” (patients with anorexia nervosa who indefinitely restrict food intake without bingeing and purging) and “bingers/purgers” (patients with an ED, sometimes anorexic, sometimes bulimic and sometimes obese, who binge or purge).7,8 Other theorists have preferred a parallel classification, based not on eating behaviours but on concurrent behavioural-trait characteristics, thus proposing a distinction between those patients with an ED (often but not exclusively those with binge-eating symptoms) who are behaviourally and affectively dysregulated and those (often, but not exclusively, “restricters”) who are behaviourally and emotionally constricted.3,9,10,11,12 The present paper concerns itself with the interface at which these distinctions (“restricting” versus “bingeing” and “dysregulation” versus “inhibition”) meet. To clarify this point, it will be useful to survey comorbid psychopathology in the EDs.
Psychiatric comorbidity
EDs frequently co-occur with other psychiatric syndromes, such as mood, anxiety and personality disorders.3,12,13,14 However, there are intriguing preferential affinities of certain comorbid syndromes with bulimic rather than restrictive ED variants, including alcohol and chemical dependencies,15 seasonal affective disorder,16 and “cluster B” or “dramatic–erratic” personality disorders.3,17,18 In other words, bulimic ED variants co-aggregate more strongly than do restrictive ones with syndromes characterized by behavioural or affective dysregulation. This tendency is repeated in first-degree relatives of patients with EDs, in that we see preferential propensity in relatives of bulimic probands toward substance-use, panic and impulse-control disorders (e.g., borderline personalty disorder) compared with propensities toward obsessive–compulsive personality traits in relatives of “restricters.”3,13,19 In other words, as a generalization, dysregulation, impulsivity and undercontrol coincide with bulimic eating syndromes, whereas compulsivity or overcontrol coincide with restrictive ones.
Although the preceding idea holds in general, potentially important heterogeneities emerge within the “binger” subgroup.3,17,18 Here, roughly one-third of cases are found to be repeatedly impulsive, reckless or self-destructive, but another third are found to display the same perfectionistic, compulsive or harm-avoiding tendencies that have come to be typically associated with restricting-type anorexia nervosa. Such variations have led some theorists to postulate the existence of distinct “binge-eater” subtypes, with distinct etiologic paths,3,10,12,14 namely, in some people with bulimic eating patterns, binge eating is driven by a pervasive “dysregulatory pathology” (affecting mood and impulse regulation), whereas in others binge eating has a more circumscribed pathogenesis, related to sequelae of chronic dietary restraint. This view is consistent, on several levels, with predictions from a “serotonin hypothesis” of the EDs, which I will now introduce.
5-HT and eating behaviour
5-HT has a well-confirmed role in the regulation of eating behaviour. In animals and in humans, manipulations that increase 5-HT neurotransmission lead to reduced eating behaviour, whereas those that reduce 5-HT activity precipitate compulsive or binge eating.20 These tendencies lead to the expectation that anorexia nervosa, in its restrictive form, should coincide with increased 5-HT tone, whereas syndromes characterized by binge eating (such as anorexia nervosa binge–purge type or bulimia nervosa) should correspond with reduced 5-HT activity. Studies of the 5-HT system in patients with clinical EDs are only partly consistent with these expectations.
At first blush, findings in women with active anorexia nervosa appear to contradict the expectation articulated here, because they indicate reduced 5-HT tone: decreased platelet binding of serotonin uptake inhibitors,21 blunted prolactin (PRL) and cortisol responses to 5-HT agonists and partial agonists,22 reduced cerebrospinal fluid (CSF) levels of 5-HT metabolite23 and decreased platelet monoamine oxidase (MAO) activity.24 However, Kaye et al23 have documented that CSF 5-hydroxyindoleacetic acid (5-HIAA) in forms of anorexia nervosa characterized by binge eating is lower than that in restrictive forms of the disorder, a finding that associates bulimic symptomatology with an expected reduction in 5-HT tone. Furthermore, findings in people who have recovered fully from anorexia nervosa reveal that data from actively ill individuals may be deceptive: following restoration of full weight, former patients with anorexia nervosa reportedly display normal PRL responses to the 5-HT-releasing agent, d-fenfluramine,25 as might indicate improvement of a state-related 5-HT abnormality. Furthermore, Kaye et al26 and Kaye and Frank27 have noted that fully recovered former patients with anorexia nervosa display elevated CSF 5-HIAA and elevated 5-HT1A receptor binding, measured by positron emission tomography (PET). These findings suggest that anorexia nervosa may actually correspond to a primary state of increased 5-HT tone, which is then masked during active illness by malnutrition-induced reductions in 5-HT activity. Indeed, Kaye et al26,27 have proposed that findings in the active stages of illness distort the existence of an underlying hyperserotonergic trait in the pathophysiology of anorexia nervosa. The same group has recently reported findings from PET studies in fully weight-recovered women who formerly had anorexia nervosa that may also be compatible with a general “5-HT overactivity hypothesis.” These findings suggest reduced 5-HT2A receptor binding in mesial temporal regions and cingulate and sensorimotor cortical regions28 and could reveal a compensation resulting from exposure to increased extracellular levels of 5-HT.
Findings in patients with active bulimia nervosa are compatible with a “low 5-HT tone hypothesis” in the sense that they associate the syndrome with abnormally low 5-HT turnover and neurotransmission and establish the 5-HT system as a potential site of action for the antecedent effects of dieting in binge episodes. Patients with active bulimia nervosa display decreased CSF 5-HIAA (at least when they binge at high frequency29), reduced platelet binding of 5-HT uptake inhibitors,30,31 reduced central transporter availability32 and blunted neuroendocrine responses to 5-HT precursors and 5-HT agonists/partial agonists, such as meta-chlorophenylpiperazine (m-CPP).33,34,35 Furthermore, tryptophan- depletion paradigms that use dietary manipulations to lower brain tryptophan and 5-HT synthesis have been shown to exacerbate bulimic symptoms in patients with active bulimia nervosa36 and to lead to the transient reappearance of such symptoms in fully recovered individuals.37
Findings in individuals who have recovered fully from bulimia nervosa, however, suggest a more complicated picture: in 2 studies, recovery from bulimia nervosa is reported to coincide with normal endocrine responses after 5-HT agonists.38,39 In a third study, based on PET techniques, findings suggest persistent reductions in postsynaptic 5-HT2A receptor activity following recovery from bulimia nervosa.40 A fourth set of results, from 23 women who had recovered fully from bulimia nervosa, suggest abnormally high CSF 5-HIAA (i.e., increased 5-HT metabolism) compared with a group of women with normal eating habits.38 These last findings parallel those obtained in women who had recovered from anorexia nervosa and support the concept that, setting disorder sequelae aside, anorexia nervosa and bulimia nervosa may both involve elevated 5-HT activity. This is possible, but we caution that the findings of Kaye et al38 indicating heightened 5-HT metabolism following recovery from bulimia nervosa may be confounded by weight discrepancies between a heavier clinical group and a lighter nonclinical group, which could partly explain the higher 5-HT metabolism observed in the group with bulimia nervosa.
At best, the available 5-HT findings lend themselves to ambiguous interpretation, with the results probably reflecting differences related to measurement, stages of illness, brain regions and other factors. Supportable generalizations, nonetheless, appear to include the following: anorexia nervosa and bulimia nervosa implicate general alterations in brain 5-HT function; there is evidence of serotonergic dysregulation, with no sweeping unidirectional tendency (toward elevation or reduction) emerging in either disorder; and compared with findings in restrictive ED variants, which imply some propensity toward increased 5-HT tone, serotonergic tendencies in binge-eating syndromes appear to be inconsistently skewed toward reduction of 5-HT activity.
Influences of malnutrition or dieting on 5-HT status
Even modest dieting is known to produce reductions in tryptophan availability and in the ratio of tryptophan to total large neutral amino acids (LNAA).41 Thus, dieting can result in reduced brain 5-HT synthesis. Likewise, animal studies show that long-term food restriction reduces central 5-HT reuptake (transporter) activity.42 Intriguingly, studies by Cowen et al43 demonstrate that dieting is associated with more pronounced alterations in postsynaptic 5-HT activity in women than in men. This finding not only links dieting to 5-HT alterations that could be causative for EDs, it also suggests that the disproportionate prevalence of EDs in females may be partly attributable to gender-linked serotonergic sensitivities! Binge eating might also affect 5-HT status: Kaye and Weltzin44 have argued that exaggerated carbohydrate consumption during binges could result in an insulin-mediated drop in total plasma LNAA, reducing competition with tryptophan across the blood–brain barrier, increasing tryptophan availability in the brain, accelerating brain 5-HT release and, ultimately, mobilizing compensatory downregulation of postsynaptic 5-HT receptors.
Possible trait-linked effects
Although the preceding supports the old adage “you are what you eat,” it is likely (as far as 5-HT function is concerned) that “you are even before you eat.” Independent of dietary factors, 5-HT and personality-trait variations are shown to correspond. Impulsivity, for example, has been consistently linked to decreased 5-HT activity: studies in rodents and primates associate reduced 5-HT neurotransmission with impulsive aggression and increased central 5-HT function with reduced aggression.45,46,47 In humans, suicide, fire setting, violence and borderline personality disorder have all been linked with decreased 5-HT metabolism (as indicated by reduced CSF 5-HIAA).48,49 Likewise, impulsive suicidality and aggression have been linked to low platelet 5-HT contents and reduced PRL response to 5-HT agonists.48,49
On the other side of the coin, findings in samples of individuals who do not have EDs have inconsistently associated anxiety, or compulsivity with increased 5-HT tone. For example, it has been reported that patients with obsessive–compulsive disorder (OCD) display elevated CSF 5-HIAA50 and increased PRL response after administration of the 5-HT agonist fenfluramine51 in some studies, but not in others.52,53 Likewise, the partial 5-HT agonist m-CPP has been observed to increase obsessionality in patients with obsessive–compulsive disorder in one study,54 but not in another.55 The same agent (m-CPP) has been shown to increase anxiety in patients with generalized anxiety disorder56,57 and, likewise, elevated 5-HT activity has been associated with anxiety in generalized anxiety disorder58 and in anorexia nervosa.59 Findings have been consistent enough to encourage some theorists to postulate that “impulsive” and "compulsive” traits occupy opposite poles of a continuum of 5-HT underactivity to overactivity,60,61 but this notion remains controversial.62 Nonetheless, it is tempting to contemplate the possibility that 5-HT findings in restrictive versus bulimic ED variants may partly reflect variations associated with differential loadings of compulsive or impulsive traits in these ED subgroups. To restate a point introduced earlier on, it may be that 5-HT variations linked to “anxiety” or “impulse regulation,” and not to “feeding behaviour,” explain serotonergic variations observed in populations with EDs.
Correspondence of trait and 5-HT variations in the EDs
Recent studies have indicated that individuals with an ED display trait-based variations on 5-HT indices. Waller et al63 observed that self-reportedly hostile patients with bulimia nervosa, compared with less hostile ones, showed smaller neuroendocrine responses following buspirone (presumed to be a 5-HT1A agonist). Likewise, Carrasco et al64 observed systematically lower platelet MAO concentrations (taken as a proxy for reduced 5-HT activity) in patients with bulimia nervosa with impulsive or “borderline” traits. Several of our studies provide comparable findings: in one, we measured PRL responses after m-CPP in women with bulimia nervosa who reported, or who denied, a history of self-mutilating or suicidal impulsivity.35 (These 2 groups of women were quite comparable on indices of body mass and in binge and purge frequency, suggesting that any differences between them were not attributable to differences in nutritional state or body weight.) The women with bulimia nervosa who reported self-harming tendencies showed blunted neuroendocrine responses following m-CPP, not only when compared with women with normal eating habits, but also when compared with the women with bulimia nervosa who denied self-harming tendencies (Fig. 1). In other words, we observed an association between blunting of the m-CPP-stimulated PRL response and self-destructiveness in women with bulimia nervosa comparable to associations with impulsivity and self-destructiveness obtained in other populations.65 Our finding suggests that hypoactivity of the 5-HT system in bulimia nervosa may, indeed, be more strongly linked to self-aggressive impulsivity than it is to binge–purge symptoms per se. Another study by our group examined platelet [3H]-paroxetine binding in healthy women and in women with bulimia nervosa and assessed the effects of “nonplanning impulsivity” (i.e., the tendency to act without considering consequences).34 Women with bulimia nervosa tended to display reduced density of paroxetine-binding sites. However, among these women, reduction in binding-site density was inversely correlated with “nonplanning.” In other words, reduced peripheral 5-HT reuptake corresponded to increased impulsivity. This effect parallels inverse relations noted between platelet 5-HT binding and aggressive impulsivity66 or self-mutilation67 in subjects with a personality disorder, raising the notion that in some individuals with bulimia nervosa we could be observing a constitutional (trait-linked) susceptibility to underactivity of the 5-HT system. Furthermore, when exacerbated by effects of dieting, such susceptibilities could cause certain people to become especially impulsive or prone to binge eating, or both.
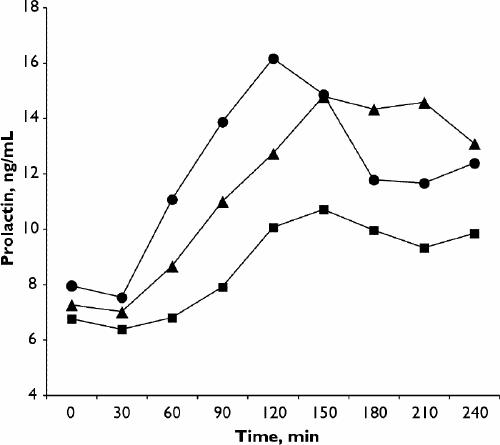
Fig. 1: Mean values obtained in serial prolactin samplings following oral administration of meta-chlorophenylpiperazine (m-CPP) in self-harming patients with bulimia (n = 29: squares), non-self-harming patients with bulimia (n = 11: circles) and controls with normal eating habits (n = 21: triangles). Reprinted with permission from Elsevier Science (Psychiatry Research 2001;103:15-26).
One more piece of evidence from our laboratory is relevant to this “trait-centred” concept of 5-HT activity. In a very recent study, we compared psychologic traits across 2 groups of women with bulimia nervosa (total n = 73), created by entering receptor density and affinity indices (from paroxetine-binding tests) into a cluster analysis.68 The analysis indicated 2 clusters of patients: one with low paroxetine binding and a second, smaller group with elevated binding. These groups showed generally similar eating symptoms and body mass, suggesting (once again) that any group differences were not likely to be attributable to differences in nutritional variables. However, the groups differed substantially as to comorbid psychopathologic characteristics. The “low 5-HT reuptake” group evinced more symptoms of borderline personality disorder, whereas the “high 5-HT” group showed more perfectionism and compulsivity. Providing a potential parallel to this finding, in long-term recovered individuals who had anorexia nervosa levels of serotonin metabolites are reported to be positively correlated with traits such as behavioural constraint, perfectionism, anxiety and obsessionality.26 Together, such results seem to be compatible with the notion (discussed earlier) of competing, trait-linked serotonergic processes that are responsible for associations between behavioural under-regulation and hyposerotonergic status, on the one hand, and over-regulation and hyperserotonergic status, on the other.
Genetic effects
If 5-HT anomalies underlie primary trait disturbances and risk of ED development, then it should be possible to demonstrate an association of particular candidate 5-HT gene variants with particular personality traits in patients with EDs and the relevance of the same gene variants to ED behaviours. Interest in 5-HT activity in the EDs has, to date, led to a catalogue of studies of 5-HT system genes: controlling the activity of 5-HT receptors and 5-HT transporter (reuptake) mechanisms.69 For anorexia nervosa, the results suggest a possible association with genes for some 5-HT receptors (e.g., 5-HT2A) and for the 5-HT transporter. However, the results are inconsistent, and none of the available studies address the possible mediating effects of concurrent traits.
For bulimia nervosa, there have been various association studies, some of which examine “trait” effects: studies of 5-HT2C polymorphisms in bulimia nervosa have detected no syndrome-linked associations.69 Similarly, 3 of 4 available studies of the 5-HT2A receptor gene indicate an absence of association with bulimia nervosa.70,71,72 However, a fourth (in a heterogeneous anorexic–bulimic sample) associates the 5-HT2A “G” allele with propensity to bulimic symptoms, borderline personality disorder and generalized impulsivity.73 Yet another recent study, the first to examine the 5-HT transporter gene (promoter region 5-HTTLPR) in bulimia nervosa, indicates that a short-allele variation confers a 7-fold risk of bulimia nervosa.74 The short (s) allele of 5-HTTLPR has been linked to reduced transcription of the 5-HT transporter protein, decreased 5-HT reuptake in lymphoblasts75 and traits such as suicidality,76 neuroticism and impulsivity.77 Preliminary findings from our laboratory provide a second indication of the relevance of the 5-HT transporter (5-HTT) gene to binge eating.78 In 48 women with binge-eating syndromes, we studied the association of 5-HTTLPR variations with psychopathologic traits (e.g., impulsivity) and platelet binding of [3H]-paroxetine. The results showed that individuals who carried the s allele (either s/s or s/l genotypes) had more tendencies compatible with impulsivity and lower density of paroxetine- binding sites than did long (l) allele homozygotes. These results, if confirmed, would have 2 significances. First, they would crossvalidate (at a genetic level) a link between impulse-control problems and hyposerotonergic status, indicating convergence among impulsive traits, low 5-HT transporter activity and the s allele. Second, they would suggest particular reduction in paroxetine binding in individuals who binge and who carry the s allele. Because prolonged restriction of food intake reduces 5-HT uptake,42 and patients with bulimia nervosa are at least intermittently restrictive in their food intake, it is possible that the s allele might confer special susceptibilities to 5-HT dysregulation in vulnerable (s-allele) individuals who challenge their 5-HT systems with chronic dieting. The hypothetical links to which I refer need to be confirmed but would be quite compatible with the notion, implicit in this paper, of “state–trait” interactions affecting 5-HT status, in this case, an interaction between a genetic propensity toward “low-ebb” 5-HT function and dietary behaviours conducive to a lowering of 5-HT availability.
Developmental correlates of 5-HT function
Beyond dietary and trait-related effects, an accumulating body of evidence links traumatic stress to reduced 5-HT function. In animals, prolonged stress alters 5-HT synthesis rates and consequent sensitivities of various 5-HT receptors.79 In humans, traumatic stress has also been linked to altered 5-HT activity. In survivors of childhood abuse, rape, holocaust or combat, post-traumatic symptoms coincide with reduced density or affinity of paroxetine-binding sites.80,81 Likewise, in women with borderline personality disorder, the strength of the neuroendocrine response after m-CPP corresponds inversely to the severity of childhood sexual or physical abuse.65
Our group has explored the implications of childhood traumata (severe physical and sexual abuse) for serotonin activity in women with and those without bulimia nervosa. The results of one of our studies associated “being bulimic” or “having been abused” with comparable degrees of blunting of neuroendocrine (PRL) responses following administration of m-CPP.82 In other words, the results were consistent with the idea that childhood abuse could be associated with a hyposerotonergic status also seen in many women with bulimia nervosa. Another of our studies found women with bulimia nervosa with reduced platelet paroxetine binding to be more likely than those with higher binding to report childhood sexual abuse.68 Together, these findings indicate a coincidence of traumatic experiences (in childhood) with hyposerotonergic tendencies in adult women with EDs. Causal interpretation of such effects is complicated (a point to which I return in the Discussion). However, it is possible that childhood abuse contributes to lasting sensitivities of the 5-HT system and, if so, such effects might heighten susceptibility, in sensitized individuals, to serotonergic dysregulation following dieting (or other factors, such a stress or depression, that may influence this system).
Discussion
In this paper, I have argued that the momentary 5-HT status in any individual, and specifically in an individual with an ED is likely to reflect a principled convergence of state, trait and developmental influences. Not surprisingly, given that brain 5-HT is derived exclusively from dietary tryptophan, one likely source of variation in 5-HT status relates to the individual's nutritional state. Among various observations that link dietary factors to 5-HT functioning are findings indicating a reduced ratio of tryptophan to LNAA after dieting,43 reduced 5-HT transporter activity following food restriction (in animals)42 and exacerbation of mood and eating symptoms in individuals with EDs following manipulations that reduce the availability of dietary serotonin precursors.36
In addition, a growing body of neurobiologic and genetic data suggest that, in individuals with EDs, personality-trait variations coincide meaningfully with variations in 5-HT system function. “Dysregulatory” phenomena (notably those associated with cognitive and behavioural forms of impulsivity) coincide, according to various indices, with reduced 5-HT neurotransmission and tone,34,35,63,64 and there is recent evidence linking trait impulsivity in patients with EDs with genetic variations that code for reduced 5-HT activity.73,78 Less consistently, some evidence in both individuals with and without bulimia nervosa points to an “opposite” tendency, linking compulsive or perfectionistic personality traits with elevated 5-HT activity.26,68 (Such variations are, I note, quite independent of eating-symptom severities and, hence, seem to reflect processes that are independent of dietary sequelae.) All of the preceding create a case for the idea that individuals' 5-HT functioning probably varies as a function of constitutionally determined (latent or manifest) personality-trait tendencies. In turn, constitutional traits associated with hypoactivity of the 5-HT system (e.g., impulsivity) may predispose toward binge eating, and traits associated with elevated 5-HT tone (e.g., compulsivity or harm avoidance) may predispose toward dietary restriction.
Evidence indicates that traumatic experiences may also affect 5-HT functioning, or predict variations in it, in both the general population80,81 and in the population with EDs.82 Such findings suggest that childhood-abuse experiences coincide with serotonergic effects measurable in full adulthood. Interpretation of such effects is not obvious, because the concurrence of 5-HT anomalies, elevated symptoms (such as impulsivity) and childhood abuse could represent several causal connections: the presence of a trait in the “high-risk” child (e.g., impulsivity or sensation seeking) that increases vulnerability to abuse; transgenerational transmission of a trait that underlies simultaneous abusiveness of certain parents and impulsiveness of certain children; chance co-occurrence of risk agents, some developmental and some constitutional/genetic, in a maladjusted population (e.g., individuals with EDs); and activation of an underlying constitutional vulnerability (e.g., tendency to reduced 5-HT activity) by traumatic stressors (e.g., childhood sexual abuse). The available data do not conclusively support a preference for any one of these alternative explanations.
Viewed together, it is likely that we need to consider (and understand) the interacting processes that have an impact upon 5-HT system function. For example, environmental stressors (e.g., childhood abuse) might act upon genetic predispositions, whereas sequelae of dieting might activate genetic (or developmental) predispositions.
Theoretical and clinical implications
Despite areas of ambiguity, current findings do suggest that traits are an important organizing construct in modelling eating and related pathologies. Indeed, the available evidence suggests that traits may be more central in characterizing patterns of variation on developmental and biologic indices within the population of individuals with EDs than are basic diagnostic distinctions (i.e., anorexia nervosa or bulimia nervosa) themselves. As a corollary, differential loadings of childhood abuse or neurobiologic abnormalities observed across individuals with anorexia nervosa and bulimia nervosa may need to be regarded as more an artifact of preferential loadings of certain traits (e.g., impulsivity) in certain ED subgroups (e.g., those who binge) than of eating-symptom (or subtype) distinctions themselves.
The multifactorial concept of serotonin functioning described in this paper has various practical implications. First, it tends to erase a strict distinction between states and traits: someone who is genetically predisposed toward low 5-HT tone, or exposed to marked developmental adversity, might be likely to display long-standing hyposerotonergic status. We might find her to be consistently prone to impulsive responses, susceptible to dietary dysregulation and any of a number of other 5-HT mediated symptoms, and we might label her as carrying a “trait disturbance.” In contrast, someone who was more favourably disposed constitutionally, if she dieted severely, might eventually place herself (and her 5-HT system) in a precarious state and, in the process, render herself susceptible to impulsive responses, dietary dysregulation and any of a number of other 5-HT mediated symptoms. We might describe this second person as having a state disturbance. From a perspective in which the momentary serotonin status is understood to represent multiple sources (some hereditary, some experiential and some diet-induced), the state–trait distinction just described loses some of its meaning. Traits, to most intents and purposes, become simply “long-lasting states.”
The concept I have just raised, if carried to its limit, calls into question assumptions that underlie DSM categorical distinctions and distinctions between Axis I (major psychiatric) and Axis II (personality) disorders. A classification by Axis I diagnosis, if valid, should organize individuals into etiologically and clinically distinct subgroups, that is, individuals with anorexia nervosa should be truly distinct from those with bulimia nervosa. Reality has it instead, however, that people cross unsettlingly from one category to the other (patients with anorexia nervosa often becoming bulimic), within-class variations are uncomfortably high (bulimia nervosa including equal proportions of highly constricted and highly disinhibited individuals) and between-class variations are sometimes unremarkable (individuals with anorexia nervosa who binge or purge being comparable, on various behavioural indices, with individuals of normal weight with bulimia nervosa). The importance of this issue is nicely illustrated in an important study by Westen and Harnden-Fischer,11 who applied Q-sort (cluster-analytic) techniques to descriptions of patients with ED provided by 103 experienced clinicians. Three broad clusters, which generally cut across DSM diagnostic designations, were identified: “high functioning and perfectionistic” (over-represented by patients with bulimia nervosa); “constricted and overcontrolled” (over-represented by patients with anorexia nervosa who are restricters); and “emotionally dysregulated, undercontrolled and impulsive” (over-represented by “bingers,” but about equally often by people with “anorexic” or “normal” weight). Such findings make a case for important heterogeneities within ED subtypes and suggest that it may be important to stress traits over “overt eating behaviours” as a basis for ED subtype distinctions. An implicit point is that personality-trait variations may be quite essential to diagnostic determinations and not as separable from syndromic distinctions as DSM would have it.
According to an expanding repertoire of findings, personality-trait variations (and especially impulsivity) prove to be relevant to outcome after ED treatment.17,83,84 Such findings imply that the trait concept (because of its implications for serotonergic involvement or because it may reflect probability of exposure to developmental abuse”) may facilitate development of more individualized (and, it is hoped, more effective) treatment efforts. For example, a “high-functioning” (less serotonergically compromised) group of patients with bulimia nervosa might have relatively focal treatment needs. Nutritionally oriented therapies might prove adequate in treating disturbances that result more from prolonged caloric restraint than from a fundamental defect of the 5-HT system. Alternatively, pathology in a more “dysregulated” subgroup of patients with an ED probably requires a different type and intensity of intervention. If EDs are influenced by pervasive serotonergic problems (related to marked constitutional defects or consequences of severe developmental stressors), nutritionally focused therapies might prove inadequate. Pharmacologic support, or more elaborate psychotherapeutic interventions aimed at pervasive impulse-control problems or post-traumatic sequelae, might be much more appropriate.
Conclusions
Personality-trait variations provide important signposts for modelling in the EDs (and elsewhere). Trait variables provide clues as to probable neurobiologic tendencies (e.g., increased or decreased 5-HT status), developmental typologies (related to likelihood of exposure to childhood abuse) and genetic substrates and, therefore, constitute an important organizing principle for our thinking about eating and more generalized psychopathology. Of course, we need further studies to characterize relevant neurobiologic/genetic variations and the phenomenologic and clinical implications of such variations. Toward this end, larger samples from multicentre studies will be needed to help characterize the complexities of state–trait (or gene–environment) interactions. Despite the limitations of the available data, however, a trait-centred (dimensional) notion of psychopathology appears to hold promise as a heuristic process toward the development of a model of psychopathology that is validated by genetic, neurobiologic, developmental and clinical findings.
Footnotes
2002 CCNP Heinz Lehmann Award Paper
Competing interests: None declared.
Correspondence to: Dr. Howard Steiger, Eating Disorders Program, Douglas Hospital, 6875 LaSalle Blvd., Montréal QC H4H 1R3; fax 514 888-4085; stehow@douglas.mcgill.ca
Submitted Jan. 29, 2003; Revised May 8, 2003; Accepted May 13, 2003
References
- 1.Garfinkel P, Garner D. Anorexia nervosa: a multidimensional perspective. New York: Brunner/Mazel; 1982.
- 2.Johnson C, Connors M. The etiology and treatment of bulimia nervosa: a multidimensional perspective. New York: Basic Books; 1987.
- 3.Steiger H, Bruce KR, Israel M. Eating disorders. In: Widiger TA, Stricker G, editors. Handbook of psychology. vol 8. Clinical psychology. New York: Wiley; 2003. p. 173-94.
- 4.Striegel-Moore RH, Silberstein LR, Rodin J. Toward an understanding of risk factors for bulimia. Am Psychol 1986;41:246-8. [DOI] [PubMed]
- 5.American Psychiatric Association. Diagnostic and statistical manual of mental disorders, fourth edition (DSM-IV). Washington: American Psychiatric Association; 1994.
- 6.Polivy J, Herman CP. Etiology of binge eating: psychological mechanisms. In: Fairburn CH, Wilson GT, editors. Binge eating: nature, assessment and treatment. New York: Guilford; 1993. p. 173-205.
- 7.DaCosta M, Halmi KA. Classifications of anorexia nervosa: question of subtypes. Int J Eat Disord 1992;11:305-13.
- 8.Garner DM, Garfinkel PE, O'Shaughnessy M. The validity of the distinction between bulimia with and without anorexia nervosa. Am J Psychiatry 1985;142:581-7. [DOI] [PubMed]
- 9.Fichter MM, Quadflieg N, Rief W. Course of multi-impulsive bulimia. Psychol Med 1994;24:591-632. [DOI] [PubMed]
- 10.Lacey JH, Evans CD. The impulsivist: a multi-impulsive personality disorder. Br J Addict 1986;81:641-9. [DOI] [PubMed]
- 11.Westen D, Harnden-Fischer J. Personality profiles in eating disorders: rethinking the distinction between axis I and axis II. Am J Psychiatry 2001;158(4):547-62. [DOI] [PubMed]
- 12.Wonderlich SA, Mitchell JE. The role of personality in the onset of eating disorders and treatment implications. Psychiatr Clin North Am 2001;24:249-58. [DOI] [PubMed]
- 13.Lilenfeld LR, Kaye WH, Greeno CG, Merikangas KR, Plotnicov K, Pollice C, et al. A controlled family study of anorexia and bulimia nervosa: psychiatric disorders in first-degree relatives and effects of proband comorbidity. Arch Gen Psychiatry 1998; 55:603-10. [DOI] [PubMed]
- 14.Vitousek K, Manke F. Personality variables and disorders in anorexia nervosa and bulimia nervosa. J Abnorm Psychol 1994; 103: 137-47. [DOI] [PubMed]
- 15.Holderness C, Brooks-Gunn J, Warren M. Co-morbidity of eating disorders and substance abuse: review of the litertaure. Int J Eat Disord 1994;16:1-34. [DOI] [PubMed]
- 16.Ghadirian AM, Marini N, Jabalpurwala S, Steiger H. Seasonal mood patterns in eating disorders. Gen Hosp Psychiatry 1999; 21: 354-9. [DOI] [PubMed]
- 17.Grilo CM. Recent research of relationships among eating disorders and personality disorders. Curr Psychiatry Rep 2002; 4(1): 18-24. [DOI] [PubMed]
- 18.Rosenvinge JH, Martinussen M, Ostensen E. The comorbidity of eating disorders and personality disorders: a meta-analytic review of studies published between 1983 and 1998. Eat Weight Disord 2000;5:52-61. [DOI] [PubMed]
- 19.Klump K, Kaye WH, Strober M. The evolving genetic foundations of eating disorders. Psychiatr Clin North Am 2001;24: 215-25. [DOI] [PubMed]
- 20.Blundell JE. Serotonin manipulations and the structure of feeding behavior. Appetite 1986;7(Suppl):39-56. [DOI] [PubMed]
- 21.Weizman R, Carmi M, Tyano S, Apter A, Rehavi M. High affinity [3H]imipramine binding and serotonin uptake to platelets of adolescent females suffering from anorexia nervosa. Life Sci 1986;38:1235-42. [DOI] [PubMed]
- 22.Monteleone P, Brambilla F, Bortolototti F, LaRocca A, Maj M. Prolactin response to d-fenfluramine is blunted in people with anorexia nervosa. Br J Psychiatry 1998;172:438-42. [DOI] [PubMed]
- 23.Kaye WH, Ebert MH, Gwirtsman HE, Weiss SR. Differences in brain serotonergic metabolism between bulimic and nonbulimic patients with anorexia nervosa. Am J Psychiatry 1984; 141: 1598-601. [DOI] [PubMed]
- 24.Diaz-Marsa M, Carrasco JL, Saiz J. A study of temperament and personality in anorexia and bulimia nervosa. J Personal Disord 2000;14(4):352-9. [DOI] [PubMed]
- 25.Ward A, Brown N, Lightman S, Campbell IC, Treasure J. Neuroendocrine, appetitive and behavioral responses to d-fenfluramine in women recovered from anorexia nervosa. Br J Psychiatry 1998;172:351-8. [DOI] [PubMed]
- 26.Kaye WH, Gwirtsman HE, George DT, Ebert MH. Altered serotonin activity in anorexia nervosa after long-term weight restoration. Does elevated cerebrospinal fluid 5-hydroxyindoleacetic acid level correlate with rigid and obsessive behavior? Arch Gen Psychiatry 1991;48:556-62. [DOI] [PubMed]
- 27.Kaye W, Frank G. Gene-environment interactions: brain and behavior in anorexia nervosa [lecture]. Annual meeting of the Eating Disorders Research Society; 2002 Nov 20-23; Charleston, South Carolina.
- 28.Frank GK, Kaye WH, Meltzer CC, Price JC, Greer P, McConaha C, et al. Reduced 5-HT2A receptor binding after recovery from anorexia nervosa. Biol Psychiatry 2002;52(9):896-906. [DOI] [PubMed]
- 29.Jimerson DC, Lesem MD, Kaye WH, Brewerton TD. Low serotonin and dopamine metabolite concentrations in cerebrospinal fluid from bulimic patients with frequent binge episodes. Arch Gen Psychiatry 1992;49:132-8. [DOI] [PubMed]
- 30.Marazziti D, Macchi E, Rotondo A, Placidi GF, Cassano GB. Involvement of the serotonin system in bulimia. Life Sci 1988; 43:2123-6. [DOI] [PubMed]
- 31.Steiger H, Leonard S, Ng Ying Kin NM, Ladouceur C, Ramdoyal D, Young SN. Childhood abuse and platelet tritiated paroxetine binding in bulimia nervosa: implications of borderline personality disorder. J Clin Psychiatry 2000;61:428-35. [DOI] [PubMed]
- 32.Tauscher J, Pirker W, Willeit M, de Zwaan M, Bailer U, Neumeister A, et al. Beta-CIT and single photon emission computer tomography reveal reduced brain serotonin transporter availability in bulimia nervosa. Biol Psychiatry 2001;49: 326-32. [DOI] [PubMed]
- 33.Levitan RD, Kaplan AS, Joffe RT, Levitt AJ, Brown GM. Hormonal and subjective responses to intravenous meta-chlorophenylpiperazine in bulimia nervosa. Arch Gen Psychiatry 1997;54:521-7. [DOI] [PubMed]
- 34.Steiger H, Young SN, Kin NM, Koerner N, Israel M, Lageix P, et al. Implications of impulsive and affective symptoms for serotonin function in bulimia nervosa. Psychol Med 2001;31:85-95. [DOI] [PubMed]
- 35.Steiger H, Koerner NM, Engleberg M, Israël M, Ng Ying Kin NM, Young SN. Self-destructiveness and serotonin function in bulimia nervosa. Psychiatry Res 2001;103(1):15-26. [DOI] [PubMed]
- 36.Kaye WH, Gendall KA, Fernstrom MH, Fernstrom JD, McConaha CW, Weltzin TE. Effects of acute tryptophan depletion on mood in bulimia nervosa. Biol Psychiatry 2000;47:151-7. [DOI] [PubMed]
- 37.Smith KA, Fairburn CG, Cowen PJ. Symptomatic relapse in bulimia nervosa following acute tryptophan depletion. Arch Gen Psychiatry 1999;56:171-6. [DOI] [PubMed]
- 38.Kaye WH, Greeno CG, Moss H, Fernstrom JD, Fernstrom MH, Lilenfeld LR, et al. Alterations in serotonin activity and psychiatric symptoms after recovery from bulimia nervosa. Arch Gen Psychiatry 1998;55:927-35. [DOI] [PubMed]
- 39.Wolfe BE, Metzger ED, Levine JM, Finkelstein DM, Cooper TB, Jimerson DC. Serotonin function following remission from bulimia nervosa. Neuropsychopharmacology 2000;22:257-63. [DOI] [PubMed]
- 40.Kaye WH, Frank GK, Meltzer CM, Price JC, McConaha CW, Crossan PJ, et al. Altered serotonin 2A receptor activity in women who have recovered from bulimia nervosa. Am J Psychiatry 2001;158:1152-5. [DOI] [PubMed]
- 41.Goodwin GM, Fraser S, Stump K, Fairburn CG, Elliot JM, Cowen PJ. Dieting and weight loss in volunteers increases the number of alpha2-adenoreceptors and 5-HT receptors on blood platelets without effect on [3H]-imipramine binding. J Affective Disord 1987;12:267-74. [DOI] [PubMed]
- 42.Huether G, Zhou D, Schmidt S, Wiltfang J, Rüther E. Long-term food restriction down regulates the density of serotonin transporters in the rat frontal cortex. Biol Psychiatry 1997; 41:1174-80. [DOI] [PubMed]
- 43.Cowen PJ, Clifford EM, Walsh AES, Williams C, Fairburn CG. Moderate dieting causes 5-HT2C receptor supersensitivity. Psychol Med 1996;26:1155-9. [DOI] [PubMed]
- 44.Kaye WH, Weltzin TE. Neurochemistry of bulimia nervosa. J Clin Psychiatry 1991;52(Suppl):21-8. [PubMed]
- 45.Eichelman BS. Bridges from the animal laboratory to the study of violent or criminal individuals. In: Hodgins S, editor. Mental disorders and crime. New York: Sage; 1993. p. 194-207.
- 46.Morand C, Young SN, Ervin FR. Clinical response of aggressive schizophrenics to oral tryptophan. Biol Psychiatry 1983; 18: 575-8. [PubMed]
- 47.Valzelli L. Reflections on experimental and human pathology of aggression. Prog Neuropsychopharmacol Biol Psychiatry 1984; 8: 311-25. [PubMed]
- 48.Åsberg M, Schalling D, Träskman-Bendz L, Wägner A. Psychobiology of suicide, impulsivity and related phenomena. In: Meltzer HY, editor. Psychopharmacology: third generation of progress. New York: Raven Press; 1987. p. 655-88.
- 49.Coccaro EF, Siever LJ, Klar HM, Cochrane K, Cooper TB, Mohs RC, et al. Serotonergic studies in patients with affective and personality disorders: correlates with suicidal and impulsive aggressive behaviour. Arch Gen Psychiatry 1989;46:587-99. [DOI] [PubMed]
- 50.Swedo SE, Leonard HL, Krusei MJP, Rettew DC, Listwak SJ, Berrettini W, et al. Cerebrospinal fluid neurochemistry in children and adolescents with obsessive-compulsive disorder. Arch Gen Psychiatry 1992;49:29-36. [DOI] [PubMed]
- 51.Fineberg NA, Roberts A, Montgomery SA, Cowen PJ. Brain 5-HT function in obsessive-compulsive disorder. prolactin responses to d-fenfluramine. Br J Psychiatry 1998;171:280-2. [DOI] [PubMed]
- 52.Baumgarten HG, Grosdanovic Z. Role of serotonin in obsessive-compulsive disorder. Br J Psychiatry 1998;173(Suppl 35): 13-20. [PubMed]
- 53.McBride PA, De Meo MD, Sweeney JA, Halper J, Mann JJ, Shear MK. Neuroendocrine and behavioral responses to challenge with the indirect serotonin agonist dl-fenfluramine in adults with obsessive-compulsive disorder. Biol Psychiatry 1992; 31:19-34. [DOI] [PubMed]
- 54.Altemus M, Swedo SE, Leonard HL, Richter D, Rubinow DR, Potter WZ, et al. Changes in cerebrospinal fluid neurochemistry during treatment of obsessive-compulsive disorder with clomipramine. Arch Gen Psychiatry 1994;51:846-9. [DOI] [PubMed]
- 55.Goodman WK, McDougle CJ, Price LH, Barr LC, Hills OF, Caplik JF, et al. M-Chlorophenylpiperazine in patients with obsessive-compulsive disorder: absence of symptom exacerbation. Biol Psychiatry 1995;38:138-49. [DOI] [PubMed]
- 56.Germine RH, Goddard AW, Woods SW, Charnet DS, Henninger GR. Anger and anxiety responses to m-chlorophenylpiperazine in generalized anxiety disorder. Biol Psychiatry 1992;32:457-61. [DOI] [PubMed]
- 57.Mundo E, Bellodi L, Smeraldy E. Effects of acute intravenous clomipramine in obsessive-compulsive symptoms and response to chronic treatment. Biol Psychiatry 1995;28:525-31. [DOI] [PubMed]
- 58.Garvey MJ, Noyes R Jr, Woodman C, Laukes C. Relationship of generalized anxiety symptoms to urinary 5-hydroxyindoleacetic acid and vanillylmandelic acid. Psychiatry Res 1995;57:1-5. [DOI] [PubMed]
- 59.Askenazy F, Candito M, Caci H, Myquel M, Chambon P, Darcourt G, et al. Whole blood serotonin content, tryptophan concentrations, and impulsivity in anorexia nervosa. Biol Psychiatry 1998;43:188-95. [DOI] [PubMed]
- 60.Cloninger CR, Svrakic DM, Przybeck TR. A psychobiological model of temperament and character. Arch Gen Psychiatry 1993;50:975-90. [DOI] [PubMed]
- 61.Hollander E. Treatment of obsessive-compulsive spectrum disorders with SSRIs. Br J Psychiatry 1998;35(Suppl):7-12. [PubMed]
- 62.Stein DJ, Trestman RI, Mitropoulou V, Coccaro EF, Hollander E, Siever LJ. Impulsivity and serotonergic function in compulsive personality disorder. J Neuropsychiatry Clin Neurosciences 1996; 8:393-8. [DOI] [PubMed]
- 63.Waller DA, Sheinberg AL, Gullion C, Moeller FG, Cannon DS, Petty F, et al. Impulsivity and neuroendocrine response to buspirone in bulimia nervosa. Biol Psychiatry 1996;39:371-4. [DOI] [PubMed]
- 64.Carrasco JL, Diaz-Marsa M, Hollander E, Cesar J, Saiz-Ruiz J. Decreased platelet monoamine oxidase activity in female bulimia nervosa. Eur Neuropsychopharmacol 2000;10:113-7. [DOI] [PubMed]
- 65.Rinne T, Westenberg HGM, den Boer JA, van den Brink W. Serotonergic blunting of meta-chlorophenylpiperazine (m-CPP) highly correlates with sustained childhood abuse in impulsive and autoaggressive female borderline patients. Biol Psychiatry 2000;47:548-56. [DOI] [PubMed]
- 66.Coccaro EF, Kavoussi RJ, Sheline YI, Lish JD, Cszernansky JG. Impulsive aggression in personality disorder correlates with tritiated paroxetine binding in the platelet. Arch Gen Psychiatry 1996; 53:531-6. [DOI] [PubMed]
- 67.Simeon D, Stanley B, Frances A, Mann JJ, Winchel R, Stanley M. Self-mutilation in personality disorders: psychological and biological correlates. Am J Psychiatry 1992;149:221-6. [DOI] [PubMed]
- 68.Steiger H, Gauvin L, Israel M, Ng Ying Kin NM, Young SN, Roussin J. Serotonin function, personality-trait variations and childhood abuse in women with bulimia-spectrum eating disorders. J Clin Psychiatry. In press. [DOI] [PubMed]
- 69.Hinney A, Remschmidt H, Hebebrand J. Candidate gene polymorphisms in eating disorders. Eur J Pharmacol 2000;410:147-59. [DOI] [PubMed]
- 70.Enoch MA, Kaye WH, Rotondo A, Greenberg BD, Murphy DL, Goldman D. 5-HT2A promoter polymorphism -1438G/A, anorexia nervosa, and obsessive-compulsive disorder. Lancet 1998;351:1785-6. [DOI] [PubMed]
- 71.Ziegler A, Hebebrand J, Gorg R, Rosenkranz K, Fichter MM, Herpertz-Dahlmann B, et al. Further lack of association between the 5-HT2A gene promoter polymorphism and susceptibility to eating disorders and a meta-analysis pertaining to anorexia nervosa. Mol Psychiatry 1999;4:410-2. [DOI] [PubMed]
- 72.Nacmias B, Ricca V, Tedde A, Mezzani B, Rotella CM, Sorbi S. 5-HT2A receptor gene polymorphism in anorexia nervosa and bulimia nervosa. Neurosci Lett 1999;277:134-6. [DOI] [PubMed]
- 73.Nishiguchi N, Matsushita S, Suzuki K, Murayama M, Shirakawa O, Higuchi S. Association between 5HT2A receptor gene promoter region polymorphism and eating disorders in Japanese patients. Biol Psychiatry 2001;50:123-8. [DOI] [PubMed]
- 74.Di Bella DD, Catalano M, Cavallini MC, Riboldi C, Bellodi I. Serotonin transporter linked polymorphic region in anorexia nervosa and bulimia nervosa. Mol Psychiatry 2000;5:233-41. [DOI] [PubMed]
- 75.Heils A, Teufel A, Petri S, Stober G, Riederer P, Bengel D, et al. Allelic variations of human serotonin transporter gene expression. J Neurochem 1996;66:2621-4. [DOI] [PubMed]
- 76.Bondy B, Erfurth A, deJonge S, Krüger M, Meyer H. Possible association of the short allele of the serotonin transporter promoter gene polymorphism (5-HTTLPR) with violent suicide. Mol Psychiatry 2000;5:193-5. [DOI] [PubMed]
- 77.Lesch KP, Wolozin BL, Murphy DL, Reiderer P. Primary structure of the human platelet serotonin uptake site: identity with the brain serotonin transporter. J Neurochem 1993;60:2319-22. [DOI] [PubMed]
- 78.Steiger H, Joober R, Israël M, Bruce K, Ng Ying Kin NM, Gauvin L, et al. A polymorphism in the promoter region of the serotonin transporter gene (5-HTTLPR) corresponds to impulsivity and reduced paroxetine binding in eating-disordered and normal-eater women [lecture]. Annual meeting of the Eating Disorders Research Society; 2002 Nov 20-23; Charleston, South Carolina.
- 79.de Souza EB. Serotonin and dopamine receptors in the rat pituitary gland: autoradiographic identification, characterization and localization. Endocrinology 1986;119:1534-42. [DOI] [PubMed]
- 80.Fichtner CG, O'Connor FL, Yeoh HC, Arora R, Crayton JW. Hypodensity of platelet serotonin uptake sites in posttraumatic stress disorder: associated clinical features. Life Sci 1995; 57: PL37-44. [DOI] [PubMed]
- 81.Maes M, Lin A, Verkerk R, Delmeire L, Gastel AV, Van der Planken M, et al. Serotonergic and noradrenergic markers of post-traumatic stress disorder with and without major depression. Neuropsychopharmacology 1999;20:188-97. [DOI] [PubMed]
- 82.Steiger H, Gauvin L, Israël M, Koerner N, Ng Ying Kin NM, Paris J, et al. Association of serotonin and cortisol indices with childhood abuse in bulimia nervosa. Arch Gen Psychiatry 2001;58:837-43. [DOI] [PubMed]
- 83.Keel PK, Mitchell JE. Outcome in bulimia nervosa. Am J Psychiatry 1997;154(3):313-21. [DOI] [PubMed]
- 84.Steinhausen HC. The outcome of anorexia nervosa in the 20th century. Am J Psychiatry 2000;159(8):1284-93. [DOI] [PubMed]