

Abstract
The human endometrium is an important site of innate immune defence, giving protection against uterine infection. Such protection is critical to successful implantation and pregnancy. Infection is a major cause of preterm birth and can also cause infertility and ectopic pregnancy. Natural anti-microbial peptides are key mediators of the innate immune system. These peptides, between them, have anti-bacterial, anti-fungal and anti-viral activity and are expressed at epithelial surfaces throughout the female genital tract. Two families of natural anti-microbials, the defensins and the whey acidic protein (WAP) motif proteins, appear to be prominent in endometrium. The human endometrial epithelium expresses beta-defensins 1–4 and the WAP motif protein, secretory leukocyte protease inhibitor. Each beta-defensin has a different expression profile in relation to the stage of the menstrual cycle, providing potential protection throughout the cycle. Secretory leukocyte protease inhibitor is expressed during the secretory phase of the cycle and has a range of possible roles including anti-protease and anti-microbial activity as well as having effects on epithelial cell growth. The leukocyte populations in the endometrium are also a source of anti-microbial production. Neutrophils are a particularly rich source of alpha-defensins, lactoferrin, lysozyme and the WAP motif protein, elafin. The presence of neutrophils during menstruation will enhance anti-microbial protection at a time when the epithelial barrier is disrupted. Several other anti-microbials including the natural killer cell product, granulysin, are likely to have a role in endometrium. The sequential production of natural anti-microbial peptides by the endometrium throughout the menstrual cycle and at other sites in the female genital tract will offer protection from many pathogens, including those that are sexually transmitted.
Introduction
The key role of the human endometrium is to orchestrate the events that lead to fertilization, implantation and pregnancy. The prevention of uterine infection is crucial to successful human reproduction and although the endometrium must function as an efficient mucosal barrier the passage of spermatozoa has to be accommodated. The innate immune system in endometrium, as elsewhere, must protect against infection while also signalling the presence of a pathogen to the acquired immune system in the event that infection does occur. Natural anti-microbials are gene-encoded peptides that are key mediators of the innate immune system and the primary focus of this review is to describe their expression in the human endometrium
The menstrual cycle
The human endometrium undergoes characteristic cyclical changes in response to the steroid hormones, oestradiol and progesterone. These have been described in detail by Noyes et al [1]. In the first half of the menstrual cycle (proliferative phase, days 4–13), when oestradiol concentrations are increasing due to the development of a follicle, the endometrium undergoes proliferation. Ovulation occurs on day 14 of the cycle, the corpus luteum forms and progesterone concentrations increase thereafter. Under the influence of progesterone endometrial growth ceases and differentiation occurs in preparation for implantation and pregnancy. The secretory function of the endometrial glands increases with maximal secretion in the mid secretory phase (days 19–23) around day 20. In the late secretory phase (days 24–28) spiral arteriole differentiation occurs and the stromal cells in immediate proximity to these vessels differentiate (decidualize). The nuclei of these cells become enlarged and the cytoplasmic volume increases. These changes also occur in the stromal cells below the surface epithelium and, if pregnancy occurs, the entire endometrium decidualizes. In the absence of implantation menstruation occurs resulting in tissue breakdown and repair allowing regeneration of the endometrium for the subsequent cycle. There are also cyclical changes to the leukocyte populations present in endometrium and these are described below.
Natural anti-microbials
The two major families of natural anti-microbials that will be discussed in relation to endometrium are (i). defensins and (ii). whey acidic protein (WAP) motif containing proteins. Studies in other mucosal systems such as the respiratory tract [2] strongly suggest that other anti-microbials will also be active within the uterus.
Defensins
Defensins are a family of small cationic proteins that have been shown to confer the host with anti-bacterial, anti-fungal and anti-viral protection. Defensins have six cysteine residues that form three disulphide bonds and they are divided into two main groups on the basis of the position of these bonds: α-defensins are found in neutrophils (human neutrophil peptides (HNP)1-4) and at epithelial sites (human defensin (HD) 5 and 6) while the β-defensins (human β defensins (HBD)) are found mainly at epithelial surfaces e.g. gut, lung [3,4]. It has recently been reported that there are 28 defensin-like sequences in the human genome [5] but currently only HBD1-6 have been characterized [6-11]. The β-defensins further subdivide into those that are constitutively expressed (e.g. HBD1) and those that are induced upon challenge with inflammatory or pathogen-derived stimuli [9,10,12-14]. In addition to their role as anti-microbials, the β-defensins are chemoattractants allowing an interaction between the innate and the acquired immune systems [8,9,15]. The expression, regulation and role of the β-defensins at non-reproductive sites has been reviewed elsewhere [16-18].
WAP motif containing proteins
Proteins containing the WAP motif have a 50 amino acid domain, which forms a tight core containing four disulphide bonds [19]. This family includes secretory leukocyte protease inhibitor (SLPI), elafin, eppin and HE4 [20]. SLPI and elafin are the most widely studied WAP proteins. These two molecules share 40% homology, with eight identical cysteines and five identical prolines, and both are expressed at mucosal surfaces where they have anti-protease and anti-microbial actions [21-26]. Both proteins inhibit serine proteases with SLPI inhibiting several proteases including neutrophil elastase and cathepsin G [27] while elafin regulates only neutrophil elastase and proteinase 3 [28-30]. These actions combined with their anti-microbial activity allow SLPI and elafin to protect epithelial surfaces during infection and inflammation. The similarity between SLPI, elafin and other proteins containing the WAP motif suggests that they may also have anti-protease and anti-microbial actions at epithelial surfaces [20].
Natural anti-microbial expression by the human endometrium
There are two main sites of natural anti-microbial expression in the endometrium: the epithelium and the leukocyte populations that are an important component of the cell types constituting the endometrium.
Natural anti-microbial expression by the endometrial epithelium
As at other mucosal surfaces the epithelial cell mediates the initial response of the endometrium to infection [31]. In 1997 and 1998 the first reports of defensin expression in the human endometrium were published. These described the expression of the α-defensin, HD5, and the β-defensin, HBD1, in the endometrial epithelium [32-34]. Subsequently, messenger RNA (mRNA) for HBD1-4 has been detected in endometrium and it has been shown that each defensin has a unique temporal expression profile [35,36]. HBD1 and 3 are expressed at highest levels during the secretory phase [35,36]. A similar pattern of expression has been reported for HD5 [33]. In contrast, HBD2 mRNA expression shows a dramatic peak during menstruation while HBD4 is expressed mainly in the proliferative phase [35,36]. These data are summarised in Figure 1. The reasons for these differential patterns of expression have not been determined and may relate to differences in anti-microbial activity of members of the defensin family or to their other activities such as chemotaxis. Acquisition of sexually transmitted infections is reported to be influenced by oestradiol and progesterone [37] and changes to the expression of innate immune molecules such as defensins may contribute to this. As in other epithelial cells, expression of both HBD2 and 3 in endometrial epithelial cells can be upregulated in response to inflammatory stimuli suggesting that these defensins will be induced in response to infection [36,38].
Figure 1.
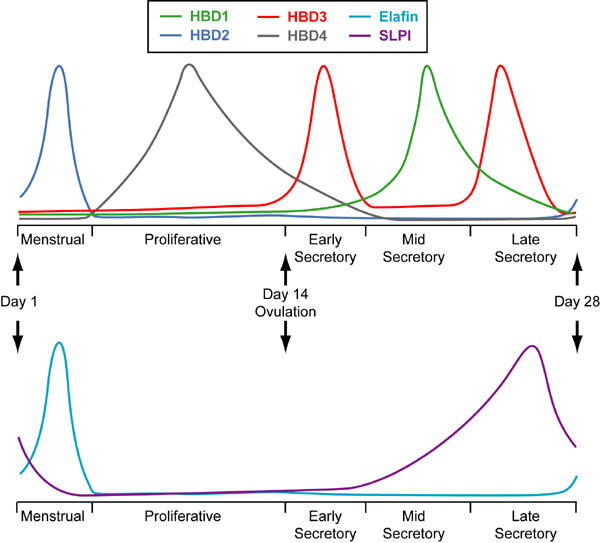
Representation of peak expression of the β-defensins, HBD1-4, and the WAP proteins, SLPI and elafin, in endometrium throughout the menstrual cycle. HBD1, HBD3 and SLPI are maximally expressed during the secretory phase. In contrast, expression of HBD4 is highest during the proliferative phase while HBD2 and elafin expression peaks during menstruation.
The WAP protein, SLPI, has also been detected in the endometrial epithelium with peak expression in the mid-late secretory phase (Figure 2a) [39]. This protein may have important actions during the window of implantation. SLPI has several anti-inflammmatory actions including inhibition of the proinflammatory transcription factor, nuclear factor κB [40], and this combined with its anti-protease activity may prevent an excessive inflammatory response from occurring in the uterus at the time of implantation and during early pregnancy. In addition, SLPI has recently been suggested to have an effect on epithelial growth. Proepithelin (PEPI) is an epithelial growth factor that can be converted to a growth inhibitor, epithelin (EPI), by elastase. SLPI can form a complex with PEPI preventing its cleavage by elastase and hence, promote epithelial growth [41]. In vitro studies using the Ishikawa epithelial cell line (derived from an endometrial adenocarcinoma) have shown that SLPI can also increase expression of the cyclin D1 gene while inhibiting expression of anti-proliferative genes such as insulin-like growth factor binding protein-3 and lysyl oxidase [42]. The anti-microbial actions of SLPI are likely to be particularly important in the protection of the uterus from infection during implantation and early pregnancy. Indeed, it has been shown that the antibacterial activity of apical secretions from polarized endometrial epithelial cells can be reduced by incubation with an anti-SLPI antibody confirming that SLPI contributes to the antibacterial defences at the epithelial surface at least in vitro [43]. SLPI concentrations and the ratio of SLPI to elastase have also been found to increase in the peritoneal fluid of women suffering from endometriosis. Local peritoneal inflammation is one of the factors thought to be responsible for the symptoms of this disease and it has been suggested that SLPI concentrations are increased in response to this and that the anti-inflammatory and anti-protease actions of SLPI may limit the inflammatory process [44].
Figure 2.
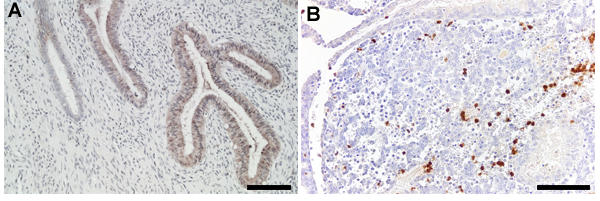
Immunohistochemical localization of the WAP proteins, SLPI and elafin, in human endometrium. (A). SLPI immunolocalization in endometrium from the mid secretory phase. Immunoreactivity is present in the glandular epithelium and secretions. (B). Elafin immunolocalization in menstrual endometrium. Immunoreactivity is present in neutrophils infiltrating the endometrial stroma. Scale bars = 100 μm.
It is likely that the endometrial epithelium will also express additional natural anti-microbials. For example, the iron-binding protein lactoferrin is expressed by the endometrial epithelium [45]. Lactoferrin has anti-microbial actions [46] and has been shown to be oestrogen regulated with maximal expression in the proliferative phase [47]. Lactoferrin has been shown to act in synergy with SLPI [48] and it is likely that interactions between natural anti-microbials in endometrium will also be important.
Natural anti-microbial expression by endometrial leukocytes
Leukocyte populations are an important component of the endometrium, constituting 8.2% of the stroma during the proliferative phase with this figure increasing to 31.7% in first trimester decidua [49]. The presence of several leukocyte populations in human endometrium is cycle dependent. The populations of neutrophils, uterine NK cells (uNK), macrophages and mast cells are relevant to endometrial anti-microbial expression. uNK cells are present in endometrium in the mid-late secretory phase of the menstrual cycle and are the major leukocyte population in first trimester decidua accounting for 70% of the leukocytes present at this time [50]. They are believed to have a role in implantation, decidualization and placentation [51]. uNK cells differ from the main population of NK cells in peripheral blood as they are CD56bright [51]. The presence of these cells in endometrium is dependent on progesterone although this may be an indirect action, as uNK cells do not express the nuclear progesterone receptor [52]. These cells express receptors for the glucocorticoid receptor and oestrogen receptor β suggesting that they do respond to these steroid hormones [53]. Macrophages are present throughout the cycle with a small increase in number premenstrually [50,54]. There are also increased numbers present in early pregnancy with a high concentration found at the implantation site [55]. Immediately prior to menstruation there is an influx of neutrophils into the endometrium [56]. Leukoctyes are involved in the tissue breakdown and repair that occurs at this time [57]. Infiltration of endometrium with both neutrophils and macrophages around the time of menstruation is coincident with falling progesterone concentrations. The effects of progesterone withdrawal on these leukocyte populations is believed to be mediated by increased expression of inflammatory mediators such as interleukin-8 and monocyte chemoattractant protein-1 in the endometrium premenstrually [58-60]. Mast cells are also present throughout the menstrual cycle, although in smaller numbers, and are activated at menstruation [61].
Neutrophils are a rich source of natural anti-microbial molecules (Figure 3). The infiltration of the endometrium by neutrophils during menstruation will enhance natural anti-microbial protection at a time when the epithelial barrier is disrupted. In addition, neutrophils will rapidly infiltrate during infection, again increasing the innate defences at an appropriate time. Peripheral blood neutrophils have been shown to express α-defensins, the cathelicidin, human cationic anti-microbial peptide 18 (hCAP-18), lactoferrin and lysozyme [62-64]. It is likely that endometrial neutrophils will also express these molecules although this has not yet been documented. Moreover, it is possible that the cytokine environment in the uterus may, at certain stages of the menstrual cycle, modulate neutrophil granule release. The WAP protein, elafin, has been detected in endometrial neutrophils (Figure 2b) [65]. Elafin mRNA and protein expression rises dramatically around the time of menstruation and immunohistochemical studies have determined that the cellular source of endometrial elafin is the neutrophil. The differing expression profiles of the two WAP proteins, SLPI and elafin (Figures 1 and 2), in endometrium suggest that they have independent roles. The anti-protease actions of elafin may aid tissue repair during menstruation while the protein's anti-microbial activity will increase innate immune defences at this time.
Figure 3.

Natural anti-microbial expression in the human neutrophil. Neutrophils contain two types of granules that contain natural anti-microbials. The azurophil granules contain α-defensins while specific granules contain lactoferrin and the cathelicidin, hCAP-18. Lysozyme is present in both granule types.
Granulysin is an anti-microbial molecule that is stored in the granules of cytolytic T cells and peripheral blood NK cells [66]. Granulysin mRNA has been found to be maximally expressed in endometrium from the late secretory phase suggesting that the uNK cell is also a source of this anti-microbial [35]. Expression of granulysin during the late secretory phase may provide additional protection around the time of implantation or in its absence, menstruation.
It is also likely that endometrial macrophages and mast cells will express natural anti-microbial peptides. There are reports that HBD1 and 2 are expressed by blood monocytes and alveolar macrophages [67] and that hCAP-18 is expressed by human skin mast cells [68]. However, localization of these peptides to endometrial leukocytes has not yet been described.
Importance of natural anti-microbial expression to reproductive health
Mucosal surfaces of the endometrium do not act in isolation to protect against infection. Anti-microbial peptides are expressed at sites throughout the female reproductive tract (Table 1) in order to protect the upper genital tract and, in pregnancy, the developing fetus, from infection. The consequences of such an infection include infertility, ectopic pregnancy and preterm birth [69,70]. However, little is known about the effects of contraceptive use on natural anti-microbial expression.
Table 1.
Natural anti-microbial expression in the female reproductive tract.
| Site of expression | Anti-microbial | Reference |
| Vagina | HD5 | [33] |
| HBD11 | [34] | |
| Elafin2 | [81] | |
| hCAP-18 | [82] | |
| Cervix | HD5 | [32,33] |
| HBD11 | [34] | |
| Elafin2 | [81] | |
| SLPI2 | [83,84] | |
| hCAP-18 | [82] | |
| Fallopian tube | HD5 (variable) | [33] |
| HBD11 | [34] | |
| SLPI2 | [85] | |
| First trimester decidua | SLPI2 | [39] |
| Amnion/Amniotic fluid | SLPI2 | [86,87] |
| Lactoferrin | [88] | |
| Term decidua | SLPI2 | [86] |
| Lactoferrin | [88] | |
| Chorion | HD5 | [32] |
| Placenta | HNP1 or 3 | [89] |
| HBD11 | [90] | |
| HBD31 | [91] |
1chemoattractant role 2anti-protease activity
The combined oral contraceptive pill (COCP) can alter the susceptibility of users to pelvic infection. For example, the prevalence of infection of the cervix with Chlamydia is increased [71] while symptomatic Chlamydial infection of the upper genital tract is decreased in COCP users compared to non-users [72]. Oral contraceptive users are also at higher risk of gonococcal infections [73]. These differences are likely to be due to several factors including changes to the viscosity of cervical mucus, alterations to cell mediated immunity and decreased menstrual blood flow [37,74]. Changes to innate immune factors are also likely to be important. Expression of the β-defensins, HBD1-4, is suppressed in COCP users compared to peak expression of these defensins in non-users suggesting that innate immune defences are altered [35,36]. This may have important consequences and it is clear that future contraceptive design should ascertain the effects on anti-microbial molecules. New contraceptives should not have a deleterious effect on the innate immune system and ideally, might enhance its actions.
Uterine infection is a major contributory factor in preterm births [70]. There are studies that suggest that the expression of anti-microbial molecules is upregulated in response to this situation. Concentrations of lactoferrin increase in amniotic fluid in patients with intra-amniotic infections, whether in preterm or term labour [75]. In addition, levels of the α-defensins, HNP1-3, are increased 4–24 fold in patients in preterm labour suffering from infection [76]. It is not yet clear whether or not any defects in expression of anti-microbial molecules exist that may increase susceptibility to uterine infections during pregnancy. The role of anti-microbials in the acquisition of sexually transmitted infections has also not been fully established although one study reports a decrease in SLPI concentrations in vaginal fluid from women suffering from lower genital tract infections [77]. In contrast, there are reports to suggest that high concentrations of SLPI may be beneficial in the limitation of spread of HIV. SLPI has been shown to be the active component of saliva that inhibits infection of monocytes with HIV [78] and there is evidence to suggest that high concentrations of SLPI in vaginal fluid reduce the likelihood that an infant will be infected with HIV during vaginal delivery [79]. Similarly, high concentrations of SLPI in infant saliva are reported to offer protection against HIV transmission during breast-feeding [80].
Conclusions
The human endometrium, in common with other mucosal surfaces, produces a wide array of natural anti-microbial peptides with the appearance of these peptides appearing to be governed by the stage of the menstrual cycle. These are expressed by both epithelial cells and endometrial leukocytes and have the potential to provide protection throughout the menstrual cycle and in pregnancy. The actions of these molecules may limit the spread of sexually transmitted infections and prevent against the uterine infections that cause preterm birth.
Contributor Information
Anne E King, Email: a.king@hrsu.mrc.ac.uk.
Hilary OD Critchley, Email: hilary.critchley@ed.ac.uk.
Rodney W Kelly, Email: r.kelly@hrsu.mrc.ac.uk.
References
- Noyes RW, Hertig AT, Rock J. Dating the Endometrial Biopsy. Fertil Steril. 1950;1:3–25. doi: 10.1016/j.fertnstert.2019.08.079. [DOI] [PubMed] [Google Scholar]
- Cole AM, Liao HI, Stuchlik O, Tilan J, Pohl J, Ganz T. Cationic polypeptides are required for antibacterial activity of human airway fluid. J Immunol. 2002;169:6985–6991. doi: 10.4049/jimmunol.169.12.6985. [DOI] [PubMed] [Google Scholar]
- Hancock RE, Diamond G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000;8:402–410. doi: 10.1016/S0966-842X(00)01823-0. [DOI] [PubMed] [Google Scholar]
- Huttner KM, Bevins CL. Antimicrobial peptides as mediators of epithelial host defense. Pediatr Res. 1999;45:785–794. doi: 10.1203/00006450-199906000-00001. [DOI] [PubMed] [Google Scholar]
- Schutte BC, Mitros JP, Bartlett JA, Walters JD, Jia HP, Welsh MJ, Casavant TL, McCray P. B., Jr. Discovery of five conserved beta -defensin gene clusters using a computational search strategy. Proc Natl Acad Sci U S A. 2002;99:2129–2133. doi: 10.1073/pnas.042692699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bals R, Wang X, Wu Z, Freeman T, Bafna V, Zasloff M, Wilson JM. Human beta-defensin 2 is a salt-sensitive peptide antibiotic expressed in human lung. J Clin Invest. 1998;102:874–880. doi: 10.1172/JCI2410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bensch KW, Raida M, Magert HJ, Schulz-Knappe P, Forssmann WG. hBD-1: a novel beta-defensin from human plasma. FEBS Lett. 1995;368:331–335. doi: 10.1016/0014-5793(95)00687-5. [DOI] [PubMed] [Google Scholar]
- Garcia JR, Jaumann F, Schulz S, Krause A, Rodriguez-Jimenez J, Forssmann U, Adermann K, Kluver E, Vogelmeier C, Becker D, Hedrich R, Forssmann WG, Bals R. Identification of a novel, multifunctional beta-defensin (human beta-defensin 3) with specific antimicrobial activityIts interaction with plasma membranes of Xenopus oocytes and the induction of macrophage chemoattraction. Cell Tissue Res. 2001;306:257–264. doi: 10.1007/s004410100433. [DOI] [PubMed] [Google Scholar]
- Garcia JR, Krause A, Schulz S, Rodriguez-Jimenez FJ, Kluver E, Adermann K, Forssmann U, Frimpong-Boateng A, Bals R, Forssmann WG. Human beta-defensin 4: a novel inducible peptide with a specific salt- sensitive spectrum of antimicrobial activity. Faseb J. 2001;15:1819–1821. [PubMed] [Google Scholar]
- Harder J, Bartels J, Christophers E, Schroder JM. Isolation and characterization of human beta -defensin-3, a novel human inducible peptide antibiotic. J Biol Chem. 2001;276:5707–5713. doi: 10.1074/jbc.M008557200. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y, Nagase T, Makita R, Fukuhara S, Tomita T, Tominaga T, Kurihara H, Ouchi Y. Identification of multiple novel epididymis-specific beta-defensin isoforms in humans and mice. J Immunol. 2002;169:2516–2523. doi: 10.4049/jimmunol.169.5.2516. [DOI] [PubMed] [Google Scholar]
- Harder J, Meyer-Hoffert U, Teran LM, Schwichtenberg L, Bartels J, Maune S, Schroder JM. Mucoid Pseudomonas aeruginosa, TNF-alpha, and IL-1beta, but not IL-6, induce human beta-defensin-2 in respiratory epithelia. Am J Respir Cell Mol Biol. 2000;22:714–721. doi: 10.1165/ajrcmb.22.6.4023. [DOI] [PubMed] [Google Scholar]
- Krisanaprakornkit S, Weinberg A, Perez CN, Dale BA. Expression of the peptide antibiotic human beta-defensin 1 in cultured gingival epithelial cells and gingival tissue. Infect Immun. 1998;66:4222–4228. doi: 10.1128/iai.66.9.4222-4228.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Neil DA, Porter E, Elewaut D, Anderson GM, Eckmann L, Ganz T, Kagnoff MF. Expression and regulation of the human b-defensins hBD1 and hBD2 in intestinal epithelium. J Immunol. 1999;163:6718–6724. [PubMed] [Google Scholar]
- Yang D, Chertov O, Bykovskaia SN, Chen Q, Buffo MJ, Shogan J, Anderson M, Schroder JM, Wang JM, Howard OM, Oppenheim JJ. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science. 1999;286:525–528. doi: 10.1126/science.286.5439.525. [DOI] [PubMed] [Google Scholar]
- Dale BA, Krisanaprakornkit S. Defensin antimicrobial peptides in the oral cavity. J Oral Pathol Med. 2001;30:321–327. doi: 10.1034/j.1600-0714.2001.300601.x. [DOI] [PubMed] [Google Scholar]
- Fellermann K, Stange EF. Defensins -- innate immunity at the epithelial frontier. Eur J Gastroenterol Hepatol. 2001;13:771–776. doi: 10.1097/00042737-200107000-00003. [DOI] [PubMed] [Google Scholar]
- Schutte BC, McCray P. B., Jr. [beta]-defensins in lung host defense. Annu Rev Physiol. 2002;64:709–748. doi: 10.1146/annurev.physiol.64.081501.134340. [DOI] [PubMed] [Google Scholar]
- Hennighausen LG, Sippel AE. Mouse whey acidic protein is a novel member of the family of 'four-disulfide core' proteins. Nucleic Acids Res. 1982;10:2677–2684. doi: 10.1093/nar/10.8.2677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clauss A, Lilja H, Lundwall A. A locus on human chromosome 20 contains several genes expressing protease inhibitor domains with homology to whey acidic protein. Biochem J. 2002;368:233–242. doi: 10.1042/BJ20020869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiemstra PS, Maassen RJ, Stolk J, Heinzel-Wieland R, Steffens GJ, Dijkman JH. Antibacterial activity of antileukoprotease. Infect Immun. 1996;64:4520–4524. doi: 10.1128/iai.64.11.4520-4524.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sallenave JM. The role of secretory leukocyte proteinase inhibitor and elafin (elastase-specific inhibitor/skin-derived antileukoprotease) as alarm antiproteinases in inflammatory lung disease. Respir Res. 2000;1:87–92. doi: 10.1186/rr18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sallenave JM. Antimicrobial activity of antiproteinases. Biochem Soc Trans. 2002;30:111–115. doi: 10.1042/0300-5127:0300111. [DOI] [PubMed] [Google Scholar]
- Simpson AJ, Maxwell AI, Govan JR, Haslett C, Sallenave JM. Elafin (elastase-specific inhibitor) has anti-microbial activity against gram-positive and gram-negative respiratory pathogens. FEBS Lett. 1999;452:309–313. doi: 10.1016/S0014-5793(99)00670-5. [DOI] [PubMed] [Google Scholar]
- Tomee JF, Koeter GH, Hiemstra PS, Kauffman HF. Secretory leukoprotease inhibitor: a native antimicrobial protein presenting a new therapeutic option? Thorax. 1998;53:114–116. doi: 10.1136/thx.53.2.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiedow O, Harder J, Bartels J, Streit V, Christophers E. Antileukoprotease in human skin: an antibiotic peptide constitutively produced by keratinocytes. Biochem Biophys Res Commun. 1998;248:904–909. doi: 10.1006/bbrc.1998.9069. [DOI] [PubMed] [Google Scholar]
- Thompson RC, Ohlsson K. Isolation, properties, and complete amino acid sequence of human secretory leukocyte protease inhibitor, a potent inhibitor of leukocyte elastase. Proc Natl Acad Sci USA. 1986;83:6692–6696. doi: 10.1073/pnas.83.18.6692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sallenave JM, Ryle AP. Purification and characterization of elastase-specific inhibitor. Sequence homology with mucus proteinase inhibitor. Biol Chem Hoppe Seyler. 1991;372:13–21. doi: 10.1515/bchm3.1991.372.1.13. [DOI] [PubMed] [Google Scholar]
- Wiedow O, Luademann J, Utecht B. Elafin is a potent inhibitor of proteinase 3. Biochem Biophys Res Commun. 1991;174:6–10. doi: 10.1016/0006-291x(91)90476-n. [DOI] [PubMed] [Google Scholar]
- Wiedow O, Schroder JM, Gregory H, Young JA, Christophers E. Elafin: an elastase-specific inhibitor of human skin. Purification, characterization, and complete amino acid sequence. J Biol Chem. 1990;265:14791–14795. [PubMed] [Google Scholar]
- Quayle AJ. The innate and early immune response to pathogen challenge in the female genital tract and the pivotal role of epithelial cells. J Reprod Immunol. 2002;57:61–79. doi: 10.1016/S0165-0378(02)00019-0. [DOI] [PubMed] [Google Scholar]
- Svinarich DM, Wolf NA, Gomez R, Gonik B, Romero R. Detection of human defensin 5 in reproductive tissues. Am J Obstet Gynecol. 1997;176:470–475. doi: 10.1016/s0002-9378(97)70517-9. [DOI] [PubMed] [Google Scholar]
- Quayle AJ, Porter EM, Nussbaum AA, Wang YM, Brabec C, Yip KP, Mok SC. Gene expression, immunolocalization, and secretion of human defensin-5 in human female reproductive tract. Am J Pathol. 1998;152:1247–1258. [PMC free article] [PubMed] [Google Scholar]
- Valore EV, Park CH, Quayle AJ, Wiles KR, McCray P. B., Jr., Ganz T. Human beta-defensin-1: an antimicrobial peptide of urogenital tissues. J Clin Invest. 1998;101:1633–1642. doi: 10.1172/JCI1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleming DC, King AE, Williams ARW, Critchley HOD, Kelly RW. Hormonal contraception can suppress natural antimicrobial gene transcription in human endometrium. Fertil Steril. 2003;79:856–863. doi: 10.1016/S0015-0282(02)04930-0. [DOI] [PubMed] [Google Scholar]
- King AE, Fleming DC, Critchley HOD, Kelly RW. Differential expression of the natural antimicrobials, beta-defensins 3 and 4, in human endometrium. J Reprod Immunol. 2003;In press doi: 10.1016/s0165-0378(02)00083-9. [DOI] [PubMed] [Google Scholar]
- Sonnex C. Influence of ovarian hormones on urogenital infection. Sex Transm Infect. 1998;74:11–19. doi: 10.1136/sti.74.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King AE, Fleming DC, Critchley HOD, Kelly RW. Regulation of natural antibiotic expression by inflammatory mediators and mimics of infection in human endometrial epithelial cells. Mol Hum Reprod. 2002;8:341–349. doi: 10.1093/molehr/8.4.341. [DOI] [PubMed] [Google Scholar]
- King AE, Critchley HOD, Kelly RW. Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial protective role. Mol Hum Reprod. 2000;6:191–196. doi: 10.1093/molehr/6.2.191. [DOI] [PubMed] [Google Scholar]
- Lentsch AB, Jordan JA, Czermak BJ, Diehl KM, Younkin EM, Sarma V, Ward PA. Inhibition of NF-kappaB activation and augmentation of IkappaBbeta by secretory leukocyte protease inhibitor during lung inflammation. Am J Pathol. 1999;154:239–247. doi: 10.1016/s0002-9440(10)65270-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J, Nathan C, Jin W, Sim D, Ashcroft GS, Wahl SM, Lacomis L, Erdjument-Bromage H, Tempst P, Wright CD, Ding A. Conversion of Proepithelin to Epithelins. Roles of SLPI and Elastase in Host Defense and Wound Repair. Cell. 2002;111:867–878. doi: 10.1016/s0092-8674(02)01141-8. [DOI] [PubMed] [Google Scholar]
- Zhang D, Simmen RC, Michel FJ, Zhao G, Vale-Cruz D, Simmen FA. Secretory leukocyte protease inhibitor mediates proliferation of human endometrial epithelial cells by positive and negative regulation of growth-associated genes. J Biol Chem. 2002;277:29999–30009. doi: 10.1074/jbc.M203503200. [DOI] [PubMed] [Google Scholar]
- Fahey JV, Wira CR. Effect of menstrual status on antibacterial activity and secretory leukocyte protease inhibitor production by human uterine epithelial cells in culture. J Infect Dis. 2002;185:1606–1613. doi: 10.1086/340512. [DOI] [PubMed] [Google Scholar]
- Shimoya K, Moriyama A, Ogata I, Nobunaga T, Koyama M, Azuma C, Murata Y. Increased concentrations of secretory leukocyte protease inhibitor in peritoneal fluid of women with endometriosis. Mol Hum Reprod. 2000;6:829–834. doi: 10.1093/molehr/6.9.829. [DOI] [PubMed] [Google Scholar]
- Masson PL, Heremans JF, Ferin J. Presence of an Iron-binding protein (lactoferrin) in the genital tract of the human female. I. Its immunohistochemical localization in the endometrium. Fertil Steril. 1968;19:679–689. doi: 10.1016/s0015-0282(16)36781-4. [DOI] [PubMed] [Google Scholar]
- Baveye S, Elass E, Mazurier J, Spik G, Legrand D. Lactoferrin: a multifunctional glycoprotein involved in the modulation of the inflammatory process. Clin Chem Lab Med. 1999;37:281–286. doi: 10.1515/CCLM.1999.049. [DOI] [PubMed] [Google Scholar]
- Teng CT, Gladwell W, Beard C, Walmer D, Teng CS, Brenner R. Lactoferrin gene expression is estrogen responsive in human and rhesus monkey endometrium. Mol Hum Reprod. 2002;8:58–67. doi: 10.1093/molehr/8.1.58. [DOI] [PubMed] [Google Scholar]
- Singh PK, Tack BF, McCray PB Jr., Welsh MJ. Synergistic and additive killing by antimicrobial factors found in human airway surface liquid. Am J Physiol Lung Cell Mol Physiol. 2000;279:L799–805. doi: 10.1152/ajplung.2000.279.5.L799. [DOI] [PubMed] [Google Scholar]
- Bulmer JN, Longfellow M, Ritson A. Leukocytes and resident blood cells in endometrium. Ann N Y Acad Sci. 1991;622:57–68. doi: 10.1111/j.1749-6632.1991.tb37850.x. [DOI] [PubMed] [Google Scholar]
- Loke YW, King A. Uterine Mucosal Lymphocytes. In: Loke YW and King A, editor. Human Implantation, Cell Biology and Immunology. Cambridge, Cambridge University Press; 1995. pp. 102–129. [Google Scholar]
- King A. Uterine leukocytes and decidualization. Hum Reprod Update. 2000;6:28–36. doi: 10.1093/humupd/6.1.28. [DOI] [PubMed] [Google Scholar]
- King A, Gardner L, Loke YW. Evaluation of oestrogen and progesterone receptor expression in uterine mucosal lymphocytes. Hum Reprod. 1996;11:1079–1082. doi: 10.1093/oxfordjournals.humrep.a019300. [DOI] [PubMed] [Google Scholar]
- Henderson TA, Saunders PT, Moffett-King A, Groome NP, Critchley HO. Steroid receptor expression in uterine natural killer cells. J Clin Endocrinol Metab. 2003;88:440–449. doi: 10.1210/jc.2002-021174. [DOI] [PubMed] [Google Scholar]
- Bulmer JN, Lunny DP, Hagin SV. Immunohistochemical characterization of stromal leucocytes in nonpregnant human endometrium. Am J Reprod Immunol Microbiol. 1988;17:83–90. doi: 10.1111/j.1600-0897.1988.tb00208.x. [DOI] [PubMed] [Google Scholar]
- Kabawat SE, Mostoufi-Zadeh M, Driscoll SG, Bhan AK. Implantation site in normal pregnancy. A study with monoclonal antibodies. Am J Pathol. 1985;118:76–84. [PMC free article] [PubMed] [Google Scholar]
- Poropatich C, Rojas M, Silverberg SG. Polymorphonuclear leukocytes in the endometrium during the normal menstrual cycle. Int J Gynecol Pathol. 1987;6:230–234. doi: 10.1097/00004347-198709000-00004. [DOI] [PubMed] [Google Scholar]
- Salamonsen LA, Lathbury LJ. Endometrial leukocytes and menstruation. Hum Reprod Update. 2000;6:16–27. doi: 10.1093/humupd/6.1.16. [DOI] [PubMed] [Google Scholar]
- Critchley HOD, Jones RL, Lea RG, Drudy TA, Kelly RW, Williams AR, Baird DT. Role of inflammatory mediators in human endometrium during progesterone withdrawal and early pregnancy. J Clin Endocrinol Metab. 1999;84:240–248. doi: 10.1210/jc.84.1.240. [DOI] [PubMed] [Google Scholar]
- Critchley HOD, Kelly RW, Lea RG, Drudy TA, Jones RL, Baird DT. Sex steroid regulation of leukocyte traffic in human decidua. Hum Reprod. 1996;11:2257–2262. doi: 10.1093/oxfordjournals.humrep.a019086. [DOI] [PubMed] [Google Scholar]
- Jones RL, Kelly RW, Critchley HOD. Chemokine and cyclooxygenase-2 expression in human endometrium coincides with leukocyte accumulation. Hum Reprod. 1997;12:1300–1306. doi: 10.1093/humrep/12.6.1300. [DOI] [PubMed] [Google Scholar]
- Jeziorska M, Salamonsen LA, Woolley DE. Mast cell and eosinophil distribution and activation in human endometrium throughout the menstrual cycle. Biol Reprod. 1995;53:312–320. doi: 10.1095/biolreprod53.2.312. [DOI] [PubMed] [Google Scholar]
- Cowland JB, Johnsen AH, Borregaard N. hCAP-18, a cathelin/pro-bactenecin-like protein of human neutrophil specific granules. FEBS Lett. 1995;368:173–176. doi: 10.1016/0014-5793(95)00634-L. [DOI] [PubMed] [Google Scholar]
- Ganz T, Selsted ME, Szklarek D, Harwig SS, Daher K, Bainton DF, Lehrer RI. Defensins. Natural peptide antibiotics of human neutrophils. J Clin Invest. 1985;76:1427–1435. doi: 10.1172/JCI112120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spitznagel JK, Dalldorf FG, Leffell MS, Folds JD, Welsh IR, Cooney MH, Martin LE. Character of azurophil and specific granules purified from human polymorphonuclear leukocytes. Lab Invest. 1974;30:774–785. [PubMed] [Google Scholar]
- King AE, Critchley HOD, Sallenave JM, Kelly RW. Elafin in human endometrium: An anti-protease and anti-microbial molecule expressed during menstruation. J Clin Endocrinol Metab. 2003;In press doi: 10.1210/jc.2003-030239. [DOI] [PubMed] [Google Scholar]
- Krensky AM. Granulysin: a novel antimicrobial peptide of cytolytic T lymphocytes and natural killer cells. Biochem Pharmacol. 2000;59:317–320. doi: 10.1016/S0006-2952(99)00177-X. [DOI] [PubMed] [Google Scholar]
- Duits LA, Ravensbergen B, Rademaker M, Hiemstra PS, Nibbering PH. Expression of beta-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Immunology. 2002;106:517–525. doi: 10.1046/j.1365-2567.2002.01430.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Nardo A, Vitiello A, Gallo RL. Cutting edge: mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J Immunol. 2003;170:2274–2278. doi: 10.4049/jimmunol.170.5.2274. [DOI] [PubMed] [Google Scholar]
- Cates W., Jr., Wasserheit JN. Genital chlamydial infections: epidemiology and reproductive sequelae. Am J Obstet Gynecol. 1991;164:1771–1781. doi: 10.1016/0002-9378(91)90559-a. [DOI] [PubMed] [Google Scholar]
- Romero R, Sirtori M, Oyarzun E, Avila C, Mazor M, Callahan R, Sabo V, Athanassiadis AP, Hobbins JC. Infection and labor. V. Prevalence, microbiology, and clinical significance of intraamniotic infection in women with preterm labor and intact membranes. Am J Obstet Gynecol. 1989;161:817–824. doi: 10.1016/0002-9378(89)90409-2. [DOI] [PubMed] [Google Scholar]
- Cottingham J, Hunter D. Chlamydia trachomatis and oral contraceptive use: a quantitative review. Genitourin Med. 1992;68:209–216. doi: 10.1136/sti.68.4.209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolner-Hanssen P, Eschenbach DA, Paavonen J, Kiviat N, Stevens CE, Critchlow C, DeRouen T, Holmes KK. Decreased risk of symptomatic chlamydial pelvic inflammatory disease associated with oral contraceptive use. Jama. 1990;263:54–59. doi: 10.1001/jama.263.1.54. [DOI] [PubMed] [Google Scholar]
- Louv WC, Austin H, Perlman J, Alexander WJ. Oral contraceptive use and the risk of chlamydial and gonococcal infections. Am J Obstet Gynecol. 1989;160:396–402. doi: 10.1016/0002-9378(89)90456-0. [DOI] [PubMed] [Google Scholar]
- McGregor JA, Hammill HA. Contraception and sexually transmitted diseases: interactions and opportunities. Am J Obstet Gynecol. 1993;168:2033–2041. doi: 10.1016/s0002-9378(12)90946-1. [DOI] [PubMed] [Google Scholar]
- Pacora P, Maymon E, Gervasi MT, Gomez R, Edwin SS, Yoon BH, Romero R. Lactoferrin in intrauterine infection, human parturition, and rupture of fetal membranes. Am J Obstet Gynecol. 2000;183:904–910. doi: 10.1067/mob.2000.108882. [DOI] [PubMed] [Google Scholar]
- Heine RP, Wiesenfeld H, Mortimer L, Greig PC. Amniotic fluid defensins: potential markers of subclinical intrauterine infection. Clin Infect Dis. 1998;27:513–518. doi: 10.1086/514691. [DOI] [PubMed] [Google Scholar]
- Draper DL, Landers DV, Krohn MA, Hillier SL, Wiesenfeld HC, Heine RP. Levels of vaginal secretory leukocyte protease inhibitor are decreased in women with lower reproductive tract infections. Am J Obstet Gynecol. 2000;183:1243–1248. doi: 10.1067/mob.2000.107383. [DOI] [PubMed] [Google Scholar]
- McNeely TB, Dealy M, Dripps DJ, Orenstein JM, Eisenberg SP, Wahl SM. Secretory leukocyte protease inhibitor: a human saliva protein exhibiting anti-human immunodeficiency virus 1 activity in vitro. J Clin Invest. 1995;96:456–464. doi: 10.1172/JCI118056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillay K, Coutsoudis A, Agadzi-Naqvi AK, Kuhn L, Coovadia HM, Janoff EN. Secretory leukocyte protease inhibitor in vaginal fluids and perinatal human immunodeficiency virus type 1 transmission. J Infect Dis. 2001;183:653–656. doi: 10.1086/318535. [DOI] [PubMed] [Google Scholar]
- Farquhar C, VanCott TC, Mbori-Ngacha DA, Horani L, Bosire RK, Kreiss JK, Richardson BA, John-Stewart GC. Salivary Secretory Leukocyte Protease Inhibitor Is Associated with Reduced Transmission of Human Immunodeficiency Virus Type 1 through Breast Milk. J Infect Dis. 2002;186:1173–1176. doi: 10.1086/343805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfundt R, van Ruissen F, van Vlijmen-Willems IM, Alkemade HA, Zeeuwen PL, Jap PH, Dijkman H, Fransen J, Croes H, van Erp PE, Schalkwijk J. Constitutive and inducible expression of SKALP/elafin provides anti-elastase defense in human epithelia. J Clin Invest. 1996;98:1389–1399. doi: 10.1172/JCI118926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frohm Nilsson M, Sandstedt B, Sorensen O, Weber G, Borregaard N, Stahle-Backdahl M. The human cationic antimicrobial protein (hCAP18), a peptide antibiotic, is widely expressed in human squamous epithelia and colocalizes with interleukin-6. Infect Immun. 1999;67:2561–2566. doi: 10.1128/iai.67.5.2561-2566.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casslen B, Rosengren M, Ohlsson K. Localization and quantitation of a low molecular weight proteinase inhibitor, antileukoprotease, in the human uterus. Hoppe Seylers Z Physiol Chem. 1981;362:953–961. doi: 10.1515/bchm2.1981.362.2.953. [DOI] [PubMed] [Google Scholar]
- Moriyama A, Shimoya K, Ogata I, Kimura T, Nakamura T, Wada H, Ohashi K, Azuma C, Saji F, Murata Y. Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle. Mol Hum Reprod. 1999;5:656–661. doi: 10.1093/molehr/5.7.656. [DOI] [PubMed] [Google Scholar]
- Ota Y, Shimoya K, Zhang Q, Moriyama A, Chin R, Tenma K, Kimura T, Koyama M, Azuma C, Murata Y. The expression of secretory leukocyte protease inhibitor (SLPI) in the Fallopian tube: SLPI protects the acrosome reaction of sperm from inhibitory effects of elastase. Hum Reprod. 2002;17:2517–2522. doi: 10.1093/humrep/17.10.2517. [DOI] [PubMed] [Google Scholar]
- Denison FC, Kelly RW, Calder AA, Riley SC. Secretory leukocyte protease inhibitor concentration increases in amniotic fluid with the onset of labour in women: characterization of sites of release within the uterus. J Endocrinol. 1999;161:299–306. doi: 10.1677/joe.0.1610299. [DOI] [PubMed] [Google Scholar]
- Zhang Q, Shimoya K, Moriyama A, Yamanaka K, Nakajima A, Nobunaga T, Koyama M, Azuma C, Murata Y. Production of secretory leukocyte protease inhibitor by human amniotic membranes and regulation of its concentration in amniotic fluid. Mol Hum Reprod. 2001;7:573–579. doi: 10.1093/molehr/7.6.573. [DOI] [PubMed] [Google Scholar]
- Niemela A, Kulomaa M, Vija P, Tuohimaa P, Saarikoski S. Lactoferrin in human amniotic fluid. Hum Reprod. 1989;4:99–101. doi: 10.1093/oxfordjournals.humrep.a136854. [DOI] [PubMed] [Google Scholar]
- Svinarich DM, Gomez R, Romero R. Detection of human defensins in the placenta. Am J Reprod Immunol. 1997;38:252–255. doi: 10.1111/j.1600-0897.1997.tb00511.x. [DOI] [PubMed] [Google Scholar]
- Zhao C, Wang I, Lehrer RI. Widespread expression of beta-defensin hBD-1 in human secretory glands and epithelial cells. FEBS Lett. 1996;396:319–322. doi: 10.1016/0014-5793(96)01123-4. [DOI] [PubMed] [Google Scholar]
- Jia HP, Schutte BC, Schudy A, Linzmeier R, Guthmiller JM, Johnson GK, Tack BF, Mitros JP, Rosenthal A, Ganz T, McCray P. B., Jr. Discovery of new human beta-defensins using a genomics-based approach. Gene. 2001;263:211–218. doi: 10.1016/S0378-1119(00)00569-2. [DOI] [PubMed] [Google Scholar]