

Abstract
Background
Fertilization in Caenorhabditis elegans requires functional SPE-9 protein in sperm. SPE-9 is a transmembrane protein with a predicted extracellular domain that contains ten epidermal growth factor (EGF)-like motifs. The presence of these EGF-like motifs suggests that SPE-9 is likely to function in gamete adhesive and/or ligand-receptor interactions.
Results
We obtained specific antisera directed against different regions of SPE-9 in order to determine its subcellular localization. SPE-9 is segregated to spermatids with a pattern that is consistent with localization to the plasma membrane. During spermiogenesis, SPE-9 becomes localized to spiky projections that coalesce to form a pseudopod. This leads to an accumulation of SPE-9 on the pseudopod of mature sperm.
Conclusions
The wild type localization patterns of SPE-9 provide further evidence that like the sperm of other species, C. elegans sperm have molecularly mosaic and dynamic regions. SPE-9 is redistributed by what is likely to be a novel mechanism that is very fast (~5 minutes) and is coincident with dramatic rearrangements in the major sperm protein cytoskeleton. We conclude that SPE-9 ends up in a location on mature sperm where it can function during fertilization and this localization defines the sperm region required for these interactions.
Background
Successful fertilization requires a precise series of cell-cell interactions between gametes [reviewed by [1-6]]. The molecules that mediate these cell-cell interactions need to be present in sperm and oocytes at the right time and place to carry out their functions. Changes in the cellular distribution of gamete proteins could also regulate activity and access to other interacting molecules. Sperm in particular are highly polarized cells with functionally and morphologically distinct surface domains [7,8]. These domains represent the compartmentalization of functions such as motility, energy production and sperm-egg cell surface interacting regions [9]. Many sperm molecules display a restricted distribution that corresponds to various sperm regions and this distribution is likely to be critical for their function [10,11].
The nematode Caenorhabditis elegans is an excellent model system for the study of the molecular mechanisms of fertilization [12,13]. Nematode sperm, like sperm from most species, are highly polarized cells. Unlike flagellated sperm, nematode sperm are amoeboid and have a single pseudopod protruding from the cell body. The pseudopod provides cellular motility, lacks organelles and has a highly dynamic membrane surface [14]. Despite this "engine in front" mode of motility, C. elegans sperm must carry out the same basic functions that are common to all sperm.
A genetic approach and the hermaphrodite/male reproductive biology of C. elegans facilitated the identification of a number of genes required for sperm-oocyte interactions at fertilization. One of these genes encodes SPE-9, a sperm transmembrane protein that contains an array of ten EGF motifs (Fig. 1) [15]. EGF motifs function as extracellular protein-protein interaction domains and are involved in diverse functions such as adhesive and ligand-receptor interactions [16]. In order to determine the distribution of SPE-9 in sperm, we obtained specific antisera directed against two different regions of the protein.
Figure 1.

Schematic representation of SPE-9. Motifs are indicated in the key by grey or black shaded boxes. Regions corresponding to sequences used to generate EX and C sera are indicated by the brackets below the diagram. The extracellular and intracellular domains of SPE-9 are indicated.
Results and Discussion
Specific antisera were obtained for two different regions of SPE-9 denoted EX (extracellular) and C (cytoplasmic) (Fig. 1). The EX sera was directed against a 13 amino acid peptide corresponding to a region between EGF motifs 4 and 5 in the predicted extracellular portion of SPE-9. The C sera was directed against a 22 amino acid peptide corresponding to the predicted cytoplasmic tail of SPE-9. We observed identical staining patterns with both of these antisera.
SPE-9 is segregated to spermatids where, depending on the plane of focus, we see a ring of staining (Fig. 2A,2B,2C,2D). In addition to the identical staining patterns observed with the EX and C antisera, we stained sperm isolated from worms carrying a null allele of spe-9 to further validate specificity. The eb19 allele of spe-9 encodes a premature stop codon before the amino acid sequences used to generate our antibodies [15]. spe-9(eb19) animals are completely sterile and produce spermatids that show no immunoreactivity to both the EX and C sera (Fig. 2E,2F,2G,2H). To make sure that SPE-9 localization patterns are not temperature dependent, we immunostained sperm isolated from worms raised at 16°C, 20°C and 25°C. The staining pattern of SPE-9 was not altered by culture temperature (data not shown). The SPE-9 staining pattern is very different from staining patterns seen with sera that stain internal sperm structures [17,18]. We also labeled spermatids with the monoclonal antibody 1CB4 [19] (Fig. 2I,2J). The epitope recognized by 1CB4 has been shown to reside on membranous organelles (MOs). MOs are located in the cytoplasm close to the plasma membrane. This MO vesicle-staining pattern is distinct from SPE-9 staining (compare 2B and 2D with 2J).
Figure 2.

Immunofluorescence localization of SPE-9 in spermatids. (A, C, E, G, I) Nomarski DIC images of spermatids. (B, D, F, H) Staining for SPE-9 in red. (J) Staining for 1CB4 in green. Staining with anti-SPE-9-EX (A, B) and anti-SPE-9-C (C, D) at this plane of focus show a ring of staining around the cell. No staining is observed in a spe-9(eb19) mutant background for either anti-SPE-9-EX (E, F) or anti-SPE-9-C (G, H). For contrast, 1CB4 stains membranous organelles (MOs) in the cytoplasm (I, J). Bar in I is 5 μm and applies to all panels.
The staining pattern seen for SPE-9 is consistent with the hypothesis that it is a sperm cell surface molecule. All known EGF motif containing proteins function extracellularly [16,20,21]. SPE-9 has a well defined signal sequence and it is most closely related to cell surface Notch/LIN-12/GLP-1 ligands [15]. Finally, if we do not permeabilize cells with detergent we can observe staining with the EX sera but not the C sera or 1CB4 (Fig. 3).
Figure 3.

SPE-9 is a cell surface protein. Non-permeabilized spermatids incubated with anti-SPE-9-EX and 1CB4 (A-C) or anti-SPE-9-C and 1CB4 (D-F). A and D show Nomarski DIC images. B and E show the anti-SPE-9 staining pattern on these cells. C and F show the lack of 1CB4 staining. DAPI staining of nuclei is blue. Dashed lines in C, E and F indicate the positions of the cells. Bar in D is 5 μm and applies to all panels.
In C. elegans, spermiogenesis refers to the conversion of round sessile spermatids to polar motile spermatozoa [reviewed in [22]]. Spermiogenesis occurs in vivo as sperm enter particular regions of the hermaphrodite reproductive tract. Preparations of spermatids can be activated in vitro with various reagents (see below and materials and methods) [23]. Although the molecular nature of this activation pathway is poorly understood, the cellular events of spermiogenesis are fairly well described. These events include the initial polymerization of major sperm protein (MSP) that drives the formation of spike-like protrusions. MSP is the sperm cytoskeletal protein required for pseudopod motility [24]. Shortly after their formation, the spikes fuse to form a mature pseudopod capable of propelling the cell. Additionally, MOs fuse with the plasma membrane. The function of the MOs is not known. However, mutations that block MO fusion produce sperm with short pseudopods, motility defects and result in sterility [25].
Spermiogenesis induced in vivo or in vitro with reagents such as pronase, TEA, or monensin can occur very rapidly (~5 minutes) [23,26-28]. With in vitro activators, it was very difficult to fix cells for immunolocalization and catch transient spikes. Trifluoperazine (TFP) initiates spermiogenesis but induces spikes that do not coalesce into a pseudopod [23]. We found that SPE-9 is localized to the spikes on these arrested cells (Fig. 4A,4B). One concern is that the large and often webbed spikes induced by TFP represent a late spike intermediate. Another way to induce persistent spikes is to treat spermiogenesis mutants such as spe-12 with pronase [23,29]. We found that SPE-9 is localized to spikes induced on spe-12 spermatids treated with pronase (Fig 4C,4D). We were initially concerned that pronase could remove immunoreactive epitopes from the surface of sperm. Under these conditions, EX staining did appear to be slightly reduced while C staining was not reduced consistent with the predicted topology of SPE-9. We conclude that either SPE-9 is a poor substrate for pronase degradation or that the concentrations used in our experiments are not sufficient to remove the bulk of immunoreactive epitopes.
Figure 4.
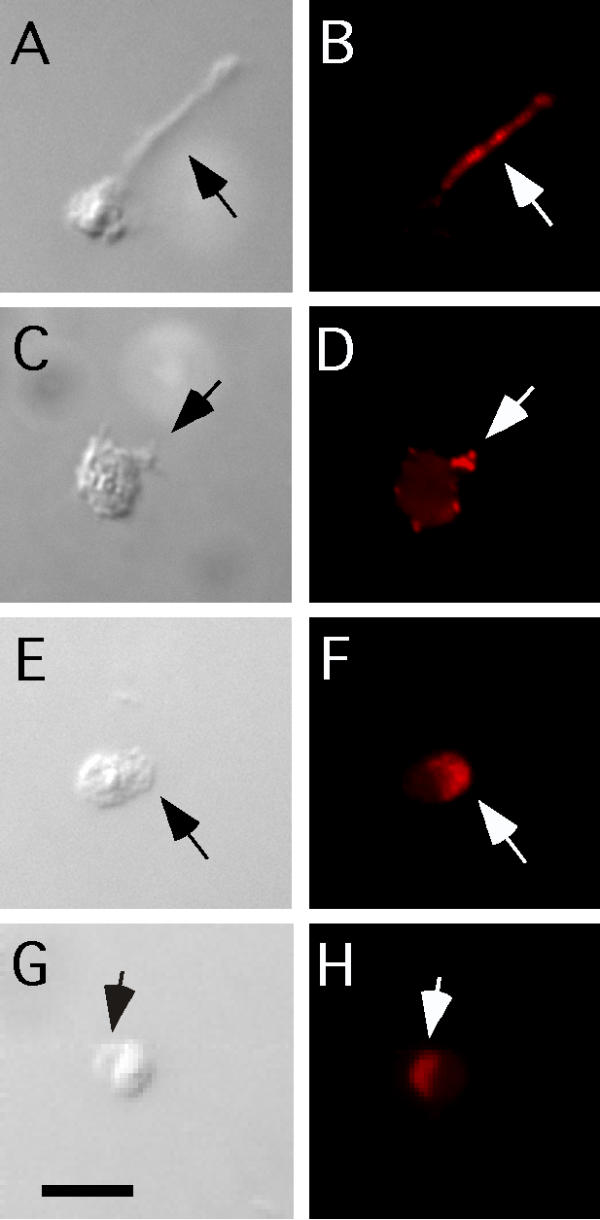
SPE-9 is concentrated in spikes during spermiogenesis and pseudopods in mature sperm. (A, C, E, G) Nomarski DIC images. (B, D, F, H) Staining for SPE-9 in red. Arrows point to spikes or pseudopods. (A, B) Wild-type spermatid activated with TFP. (C, D) spe-12(hc76) spermatid treated with pronase. Note the numerous small spots that correspond to arrested or developing spikes. (E, F) Wild-type sperm activated by TEA treatment and stained with anti-SPE-9-EX. (G, H) Wild-type sperm activated by TEA treatment and stained with anti-SPE-9-C. Bar in G is 5 μm and applies to all panels.
For mature sperm, we found SPE-9 concentrated on the pseudopod (Fig. 4E,4F,4G,4H). We observed the same localization patterns for SPE-9 regardless of the sperm activator used or culture temperature. These results make intuitive sense since SPE-9 is previously localized to spikes that coalesce to form the pseudopod.
Figure 5 shows a schematic summary of SPE-9 distribution. Localization and dynamic redistribution of cell surface components is a fundamental feature of polarized cells that is absolutely required for their proper function in an organism. There are several major mechanisms that could drive the localization of membrane proteins. One mechanism involves the specific distribution of newly synthesized proteins to discrete cell surface domains. The other mechanism involves the redistribution of molecules after insertion into the plasma membrane. The latter mechanism is relevant to SPE-9 since there is no protein synthesis during spermiogenesis or in mature sperm [30,31]. Our data show that SPE-9 is dynamically redistributed through spermiogenesis. This redistribution could occur by trapping SPE-9 in spikes and then pseudopods or through active translocation to these structures. Determining the underlying mechanism of SPE-9 localization will be interesting. The movement of SPE-9 is coincident with major rearrangements of the MSP cytoskeleton. Furthermore, SPE-9 has no known localization sequences.
Figure 5.

Summary of the localization of SPE-9. SPE-9 localization is indicated with red.
For C. elegans, the precise mode of sperm entry into the oocyte is still poorly characterized. An important unanswered question concerns the region of C. elegans sperm (e.g. cell body or pseudopod) that interacts with the egg surface. The requirement of SPE-9 for fertilization and its localization pattern in mature sperm is consistent with the idea that C. elegans sperm interact with the oocyte cell surface "pseudopod first". This idea also makes intuitive sense since the cell body is dragged along and the motility apparatus is in the front. In contrast, flagellated sperm propel the cell body in front and interact with the egg plasma membrane in a spatially restricted manner [5]. In mammals, the sperm cell body is highly mosaic and only an equatorial region of the sperm head is important for sperm binding and fusion with the egg plasma membrane [11,32]. Furthermore, molecules thought to mediate these events are specifically restricted to this region [33].
Conclusion
This is the first report describing the wild type localization pattern of SPE-9 in sperm. The distribution of SPE-9 through spermiogenesis is dynamic. The accumulation of SPE-9 in specific sperm structures is very fast and is coincident with dramatic rearrangements in the major sperm protein cytoskeleton. The mechanism of this redistribution may be novel since SPE-9 has no known localization signals (e.g. PDZ recognition sequence). The concentration of SPE-9 in the pseudopod suggests that this sperm structure is not only important for locomotion but is also important for gamete interactions. Finally, we have generated new reagents that will be useful for the future study of sperm development and function. For instance, SPE-9 antibodies can be used for biochemical studies or as cellular markers for the study of sperm mutants.
Methods
Nematode strains and culture
C. elegans culture and manipulation were essentially as described by Brenner [34]. Strain N2 was considered wild-type. The following mutations were used in this study: spe-9(eb19), spe-12(hc76), him-5(e1490), him-8(e1489). Detailed descriptions of all mutations can be found in L'Hernault [22], Singson et al. [15] or Hodgkin [35]. Animals were cultured at 16°C, 20°C or 25°C. In several experiments, the mutations him-5(e1490) or him-8 (e1489) were used because they produce males at high frequency with no adverse effects on sperm [S.Z. and A.W.S., unpublished observations, [26,36]].
Isolation, in vitro activation and immunofluorescence of sperm
Polyclonal antibodies directed against peptides corresponding to two different regions of the predicted C. elegans SPE-9 amino acid sequence were obtained through Zymed Laboratories (South San Francisco, CA) custom peptide/antibody service. Rabbits were injected with synthetic peptides conjugated to keyhole limpet hemocyanin. Only rabbits with preimmune sera that did not show immunoreactivity on C. elegans spermatids were selected for these experiments. The regions selected for peptide synthesis were chosen based on a lack of homology to other EGF motif-containing proteins, favorable surface probability, antigenic index, hydrophilicity and the location of predicted secondary structures. The sequence of these SPE-9 peptides is as follows: EX, 13 amino acids (C)KNDYNDGKNVNGT in the extracellular region between EGF motif 4 and 5. C, the 22 amino acid cytoplasmic tail (C)SRRRQGRVEEAKKTSEVKTENP (Fig. 1) [15]. The non-coded N-terminal cysteine residues were included for single point site directed conjugation.
Immunofluorescence was based on the protocol described in Arduengo et al. [17]. Young L4 stage males were placed on agar plates and grown without hermaphrodites for 24 hours. About 20 males were dissected for sperm with a needle on a slide containing 10 μl of sperm media (SM) salts (50 mM Hepes, 1 mM MgSO4, 25 mM KCl, 45 mM NaCl, 5 mM CaCl2) plus 10 mM dextrose with or without the in vitro sperm activators pronase E (200 μg/ml), triethanolamine (TEA) (120 mM at pH 7.8), monensin (10-7 M) or Trifluoperazine (TFP) (50 μM) [23,26,28,36,37].
Monoclonal antibody 1CB4 [19] was used for immunofluorescence at 1:20 dilution. Mouse monoclonal primary antibodies were visualized by using fluorescein (FITC)-conjugated affinity purify goat anti-mouse IgG secondary antibody (1:500 in PBS) (Jackson ImmunoResearch Laboratories, Inc. West Grove, PA). Polyclonal antibodies for SPE-9 (C and EX) were generated in rabbits (see above) and detected with a rhodamine (TRITC)-conjugated affinity purify goat anti-rabbit IgG secondary antibody (1:1000 in PBS) (Jackson ImmunoResearch Laboratories, Inc. West Grove, PA). Affinity purified or crude extracts of the SPE-9 sera, EX or C, were used at either 1:1000 or 1:100 dilutions. All microscopy employed a Zeiss Axioplan microscope fitted with a X100 Plan Neofluor objective and micrographs were taken through the appropriate filters. Images were captured on a SensiCam or Optronics digital camera and edited in Adobe Photoshop 7 and Deneba Systems Canvas 8.
Authors' contributions
S.Z. carried out all immunofluorescence experiments. All experiments were conducted in the laboratory of A.W.S. Peptides for antibody production were determined by A.W.S. while working as a postdoctoral fellow in the laboratory of S.W.L. Antibodies were also obtained by A.W.S. while working as a postdoctoral fellow in the laboratory of S.W.L. The manuscript was written by A.W.S.
Acknowledgments
Acknowledgements
We would like to thank Kim McKim, Chris Rongo and Andrea Dean for critical comments on the manuscript. We thank members of the Singson lab for assistance with figures, helpful discussions and comments. The Caenorhabditis Genetic Center provided some nematode strains, and it is funded by the NIH National Center for Research Resources (NCRR). Grants to A.W.S. were from the National Institutes of Health (R01 GM63089-01), the National Science Foundation (IBN-0000182), and the Charles & Johanna Busch Biomedical Fund. A grant to S.W.L. was from the National Science Foundation (IBN-9631102).
Contributor Information
Sonia Zannoni, Email: szannoni@hotmail.com.
Steven W L'Hernault, Email: bioslh@biology.emory.edu.
Andrew W Singson, Email: singson@waksman.rutgers.edu.
References
- Primakoff P, Myles DG. Penetration, adhesion, and fusion in mammalian sperm-egg interaction. Science. 2002;296:2183–2185. doi: 10.1126/science.1072029. [DOI] [PubMed] [Google Scholar]
- Evans Janice P., Florman Harvey M. The state of the union: the cell biology of fertilization. Nature Cell Biology & Nature Medicine fertility supplement. 2002. pp. s57–s63. [DOI] [PubMed]
- Singson A, Zannoni S, Kadandale P. Molecules that function in the steps of fertilization. Cytokine Growth Factor Rev. 2001;12:299–304. doi: 10.1016/S1359-6101(01)00013-2. [DOI] [PubMed] [Google Scholar]
- Vacquier Victor D. Evolution of gamete recognition proteins. Science. 1998;281:1995–1998. doi: 10.1126/science.281.5385.1995. [DOI] [PubMed] [Google Scholar]
- Yanagimachi R. Mammalian fertilization. In: E Knobil and J D Neill, editor. The Physiology of Reproduction. New York, Raven Press; 1994. pp. 189–317. [Google Scholar]
- Talbot P, Shur BD, Myles DG. Cell adhesion and fertilization: steps in oocyte transport, sperm-zona pellucida interactions, and sperm-egg fusion. Biol Reprod. 2003;68:1–9. doi: 10.1095/biolreprod.102.007856. [DOI] [PubMed] [Google Scholar]
- Flesch FM, Gadella BM. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim Biophys Acta. 2000;1469:197–235. doi: 10.1016/S0304-4157(00)00018-6. [DOI] [PubMed] [Google Scholar]
- Myles DG, Koppel DE, Cowan AE, Phelps BM, Primakoff P. Rearrangement of sperm surface antigens prior to fertilization. Ann N Y Acad Sci. 1987;513:262–273. doi: 10.1111/j.1749-6632.1987.tb25014.x. [DOI] [PubMed] [Google Scholar]
- Travis Alexander J., Kopf Gregory S. The spermatozoon as machine: Compartmentalized metabolic and signaling pathways bridge cellular structure and function. In: DeJonge C and Barratt C L R, editor. Assisted Reproductive Technology. Cambridge, University Press, Inc.; 2002. pp. 26–39. [Google Scholar]
- Primakoff P, Myles DG. A map of the guinea pig sperm surface constructed with monoclonal antibodies. Dev Biol. 1983;98:417–428. doi: 10.1016/0012-1606(83)90371-8. [DOI] [PubMed] [Google Scholar]
- Ramalho-Santos Joao, Schatten Gerald, Moreno Picardo. Control of membrane fusion during spermiogenesis and the acrosome reaction. Biology of Reproduction. 2002;67:1043–1051. doi: 10.1095/biolreprod67.4.1043. [DOI] [PubMed] [Google Scholar]
- Singson A. Every sperm is sacred: fertilization in Caenorhabditis elegans. Dev Biol. 2001;230:101–109. doi: 10.1006/dbio.2000.0118. [DOI] [PubMed] [Google Scholar]
- L'Hernault Steven W., Singson Andrew. Developmental genetics of spermatogenesis in the nematode Caenorhabditis elegans. In: Erwin Goldberg, editor. The Testis: From Stem Cell to Sperm Function, Serono Symposium USA. New York, Springer-Verlag Inc.; 2000. pp. 109–119. [Google Scholar]
- Roberts Thomas M., Ward Samuel. Membrane flow during nematode spermiogenesis. The Journal of Cell Biology. 1982;92:113–120. doi: 10.1083/jcb.92.1.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singson Andrew, Mercer Kristina B., L'Hernault Steven W. The C. elegans spe-9 gene encodes a sperm transmembrane protein that contains EGF-like repeats and is required for fertilization. Cell. 1998;93:71–79. doi: 10.1016/s0092-8674(00)81147-2. [DOI] [PubMed] [Google Scholar]
- Campbell Iain D., Bork Peer. Epidermal growth factor-like modules. Current Opinions in Structural Biology. 1993;3:385–392. [Google Scholar]
- Arduengo PM, Appleberry OK, Chuang P, L'Hernault SW. The presenilin protein family member SPE-4 localizes to an ER / Golgi derived organelle and is required for proper cytoplasmic partitioning during C. elegans spermatogenesis. Journal of Cell Science. 1998;111:3645–3654. doi: 10.1242/jcs.111.24.3645. [DOI] [PubMed] [Google Scholar]
- Roberts TM, Pavalko FM, Ward S. Membrane and cytoplasmic proteins are transported in the same organell complex during nematode spermatogenesis. Journal of Cell Biology. 1986;102:1787–1796. doi: 10.1083/jcb.102.5.1787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okamoto H, Thomson JN. Monoclonal antibodies which distinguish certain classes of neuronal and supporting cells in the nervous tissue of the nematode Caenorhabditis elegans. J Neurosci. 1985;5:643–653. doi: 10.1523/JNEUROSCI.05-03-00643.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Appella E, Weber IT, Blasi F. Structure and function of epidermal growth factor-like regions in proteins. FEBS Letters. 1988;231:1–4. doi: 10.1016/0014-5793(88)80690-2. [DOI] [PubMed] [Google Scholar]
- Davis CG. The many faces of epidermal growth factor repeats. New Biologist. 1990;2:410–419. [PubMed] [Google Scholar]
- L'Hernault Steven W. Spermatogenesis. In: Donald L Riddle, Thomas Blumenthal, Barbara J Meyer and James R Priess, editor. C Elegans II. Cold Spring Harbor, Cold Spring Harbor Laboratory; 1997. pp. 271–294. [PubMed] [Google Scholar]
- Shakes Diane C., Ward Samuel. Initiation of spermiogenesis in C. elegans: a pharmacological and genetic analysis. Dev Biol. 1989;134:189–200. doi: 10.1016/0012-1606(89)90088-2. [DOI] [PubMed] [Google Scholar]
- Roberts TM, Stewart M. Acting like actin. The dynamics of the nematode major sperm protein (msp) cytoskeleton indicate a push-pull mechanism for amoeboid cell motility. J Cell Biol. 2000;149:7–12. doi: 10.1083/jcb.149.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Achanzar William E., Ward Samuel. A nematode gene required for sperm vesicle fusion. Journal of Cell Science. 1997;110:1073–1081. doi: 10.1242/jcs.110.9.1073. [DOI] [PubMed] [Google Scholar]
- Nelson GA, Ward S. Vesicle fusion, pseudopod extension and amoeboid motility are induced in nematode spermatids by the ionophore monensin. Cell. 1980;19:457–464. doi: 10.1016/0092-8674(80)90520-6. [DOI] [PubMed] [Google Scholar]
- Ward Samuel, Carrel John S. Fertilization and sperm competition in the nematode Caenorhabditis elegans. Dev Biol. 1979;73:304–321. doi: 10.1016/0012-1606(79)90069-1. [DOI] [PubMed] [Google Scholar]
- Ward Samuel, Hogan Eileen, Nelson Gregory A. The initiation of spermiogenesis in the nematode Caenorhabditis elegans. Dev Biol. 1983;98:70–79. doi: 10.1016/0012-1606(83)90336-6. [DOI] [PubMed] [Google Scholar]
- Nance J, Minniti AN, Sadler C, Ward S. spe-12 encodes a sperm cell surface protein that promotes spermiogenesis in Caenorhabditis elegans. Genetics. 1999;152:209–220. doi: 10.1093/genetics/152.1.209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavalko FM, Roberts TM. Posttranslational insertion of a membrane protein on Caenorhabditis elegans sperm occurs without de novo protein synthesis. J Cell Biochem. 1989;41:57–70. doi: 10.1002/jcb.240410203. [DOI] [PubMed] [Google Scholar]
- Ward S. The asymmetric localization of gene products during the development of Caenorhabditis elegans spermatozoa. In: J Gall, editor. Gametogenesis and the Early Embryo. New York, A. R. Liss; 1986. pp. 55–75. [Google Scholar]
- Myles Diana G. Molecular mechanisms of sperm-egg membrane binding and fusion in mammals. Dev Biol. 1993;158:35–45. doi: 10.1006/dbio.1993.1166. [DOI] [PubMed] [Google Scholar]
- Primakoff P, Hyatt H, Tredick-Kline J. Identification and purification of a sperm surface protein with a potential role in sperm-egg membrane fusion. J Cell Biol. 1987;104:141–149. doi: 10.1083/jcb.104.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenner Sydney. The genetics of Caenorhabditis elegans. Genetics. 1974;77:71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodgkin Jonathan. Appendix 1, Genetics. In: Donald L Riddle, Thomas Blumenthal, Barbara J Meyer and James R Priess, editor. C elegans II. Cold Spring Harbor, Cold Spring Harbor Laboratory Press; 1997. pp. 881–1048. [Google Scholar]
- L'Hernault SW, Roberts TM. Cell biology of nematode sperm. Methods in Cell Biology. 1995;48:273–301. doi: 10.1016/s0091-679x(08)61392-8. [DOI] [PubMed] [Google Scholar]
- Machaca K, DeFelice LJ, L'Hernault SW. A novel chloride channel localizes to Caenorhabditis elegans spermatids and chloride channel blockers induce spermatid differentiation. Dev Biol. 1996;176:1–16. doi: 10.1006/dbio.1996.9999. [DOI] [PubMed] [Google Scholar]