

Abstract
Purpose
The purpose of this study was to determine if loss of serine protease HtrA1 in endometrial cancer will promote the invasive potential of EC cell lines.
Experimental design
Western blot analysis and immunohistochemistry methods were used to determine HtrA1 expression in EC cell lines and primary tumors, respectively. Migration, invasion assays and in vivo xenograft experiment were performed to compare the extent of metastasis between HtrA1 expressing and HtrA-1 knocked down clones.
Results
Western blot analysis of HtrA1 in 13 EC cell lines revealed complete loss of HtrA1 expression in all 7 papillary serous EC cell lines. Downregulation of HtrA1 in Hec1A and Hec1B cell lines resulted in a 3-4 fold increase in the invasive potential. Exogenous expression of HtrA1 in Ark 1 and Ark 2 cells resulted in 3-4 fold decrease in both invasive and migration potential of these cells. There was an increased rate of metastasis to the lungs associated with HtrA1 downregulation in Hec1B cells compared to control cells with endogenous HtrA1 expression. Enhanced expression of HtrA1 in Ark 2 cells resulted in significantly less tumor nodules metastasizing to the lungs compared to parental or protease deficient (SA mutant) Ark 2 cells. Immunohistochemical (IHC) analysis showed 57% (105/184) of primary EC tumors had low HtrA1 expression. The association of low HtrA1 expression with high-grade endometrioid tumors was statistically significant (p=0.016).
Conclusions
Collectively, these data indicate loss of HtrA1 may contribute to the aggressiveness and metastatic ability of endometrial tumors.
Keywords: HtrA1, metastasis, Endometrial Cancer
Introduction
The American Cancer Society predicts that 41,200 women will be diagnosed with endometrial cancer, and 7,350 women will die of the disease in 2006 (American Cancer Society, 2006). While highly amenable to curative surgical treatment, with the overall 5-year survival rate of 86%, metastatic disease remains the most significant contributor to morbidity and mortality in patients with endometrial cancer. Those patients who present with advanced or recurrent disease will require surgical therapy as well as adjuvant therapy with little impact on overall survival. While multiple mechanisms influence tumor aggressiveness and tendency for recurrence, metastatic capability is the most important factor influencing patient survival. Understanding the molecular and biochemical mechanisms resulting in a metastatic phenotype is expected to fuel the development of new therapeutic approaches.
HtrA1 is one of the four members of the HtrA family of serine proteases. These members share two highly conserved domains, the PDZ binding domain and the trypsin-like catalytic domain (1). Bacterial HtrA proteases serve dual functions; one as chaperone proteins at low/normal temperature and as proteases at high temperatures (2). Second, under conditions of cellular stress, the members of this protein family play a role in activating the cellular stress response and exhibit evidence of increasing proteolytic activity (3-6). Originally identified in the bacteria (7), homologs of this protein family has been identified in vertebrae, invertebrate and humans, after loss of its expression was noted in transformed fibroblasts (1). Thus far, HtrA1 has been implicated in osteoarthritis (8), Alzheimer's disease (9), macular degeneration (10) and appears to play a role in some neuromuscular diseases, in particular muscular dystrophy (11).
Chien et al. identified HtrA1 as a proapoptotic downregulated gene in several types of malignancies including ovarian cancer (12). Additionally, Baldi et al. (13) showed that HtrA1 acts as a metastatic tumor suppressor in malignant melanoma, in vitro. Using immunohistochemistry, Bowden et al. measured protein expression levels of HtrA1, HtrA2, and HtrA3 in progressive grades of endometrial adenocarcinoma, showing considerably less staining for all three HtrA proteins in grade 3 carcinoma (14). HtrA1 has also been shown to play a role in inducing cell death of cancer cells as well as a modulator of differential response to chemotherapy (3). While these studies imply a role for HtrA1 as a pro-apoptotic tumor suppressor, its functional significance in endometrial carcinoma remains undefined. In this study, we have identified additional role for HtrA1 in invasion and migration in vitro and in vivo and tested whether HtrA1 expression in primary tumor samples correlates with clinical outcome. Identification of HtrA1 as a modulator of metastatic potential would have potential clinical implications for patient risk stratification to identify those at greater risk for metastatic or advanced disease in endometrial cancer.
Materials and Methods
Cell culture
Ark 1 and Ark 2 uterine papillary serous cell lines were maintained in DMEM/F12 serum with 10% FBS. Hec1A and Hec1B endometrioid-type cell lines were maintained in DMEM/F12 serum with 10% FBS.
Antibodies
An affinity-purified polyclonal rabbit anti-human HtrA1 antibody was raised against polypeptide corresponding to amino acids 161-480 of HtrA1 (8). A monoclonal antibody against ß-actin was purchased from Sigma-Aldrich.
Plasmids
Plasmids encoding wild-type (WT) or mutant S328A (SA) HtrA1 were generated by PCR cloning into pcDNA3.1. These plasmids had been established and described by Hu et al, 1998 (8)and used by Chien et al 2004 (12). Empty vector pcDNA3.1 was used as the control vector.
Transient transfection with siRNA
HtrA1 was transiently downregulated in Hec1A and Hec1B cell lines. Scrambled control siRNA (5′-Fluo-UCCUGCUGGAGCCUCAUGUTT-3′) and HtrA1 siRNA targeting the 3′UTR (5′-Fluo-CGGCCGAAGUUGCCUCUUUTT-3′) were purchased from Sigma-Aldrich and transfected in OptiMEM using Oligofectamine (Invitrogen) per manufacturer's instructions. For optimal transfection conditions to achieve >80-90% downregulation, Hec1A cells were plated at 40,000 cells/well in a 24-well plate or 200,000 cells/well in a 6-well plate while Hec 1B cells were plated at 30,000 cell/well in a 24-well plate or 150,000 cells/well in a 6-well plate. Western immunoblot analysis using whole cell lysate was performed 48hrs after transfection to confirm efficiency of downregulation.
Establishment of stable transfectants
HtrA1 expressed with induced in Ark1 and Ark2 cell lines through the establishment of stable clones. Exponentially growing Ark1 and Ark2 cells in 35mm dishes were washed with serum-free media and incubated for 15 minutes with a reaction solution containing 1μg of plasmid (WT HtrA1, SA HtrA1, or empty vector pcDNA3.1 control), 6μl of Plus reagent (Invitrogen, Carlsbad, CA, USA), and 4 μl of Lipofectamine. After incubation for 3hrs, complete medium with 10% serum was added. 24hrs after transfection, cells were transferred to 100mm plate. 48hrs after transfection, G418 (Geneticin) was added at a final concentration of 500 μg/ml or 900 μg/ml to select stable Ark1 and Ark2 transfectants, respectively. Clones were screened by western blot analysis and selected for migration and invasion assays based on sufficient HtrA1 expression.
Western blot analysis
Using Laemmli buffer containing fresh 2.5% ß-mercaptoethanol, cells were lysed and boiled for 5 minutes. Western immunoblot analysis was performed on whole-cell lysates using the affinity-purified polyclonal rabbit anti-human HtrA1 antibody at 1:500 dilution. Loading control was confirmed using anti-ß-actin antibody at 1:5000 dilution.
Migration and invasion assay
The migratory and invasive capacity of stable Ark1 and Ark2 clones (empty vector control (EV), HtrA1 SA, and HtrA1 WT) and transiently transfected Hec1A and Hec1B (scr-control, siRNA) was analyzed as described previously (3). Experimental conditions were optimized to required cell number and necessary time for migration or invasion. Cell migration was evaluated using transwell plates (24-well format, 8-μm pore, Millipore). To measure clonal cell invasion, Matrigel Invasion chambers (24-well format, 8-μm pore, BD Biosciences, Clontech) were used. 7.5 × 104 cells were seeded in the upper chamber in serum-free media. Media containing 10% fetal bovine serum will be placed in the lower chamber as a chemo-attractant. After a 12 hr 37°C incubation period (to measure transwell migration) or a 24hr incubation period (to measure Matrigel invasion), cells on the upper surface of the membrane filter were removed using a cotton-wool swab. Cells which migrated/invaded through to the lower surface were fixed with 3.7% formaldehyde, stained with methyl violet, and counted in 4 low-power fields (magnification 20X). Mean number of net migrating or invading cells ± SD were determined and expressed as % of control vector. Experiments were all performed in triplicate.
Scratch Assay
HtrA1 targeting shRNA (shRNA1 and shRNA2) and non-targeting control shRNA (NTC) were purchased from Sigma-Aldrich. shRNA downregulated clones were generated as previously described (15). Stable batch clones of NTC, shRNA1 and shRNA2 in Hec1A and Hec1B were plated (20,000 cell/well in 24-well plate). Scratch assay to determine migration of cells was performed as previously described (15).
In vivo metastatic model
After obtaining the approval from the institutional animal care and use committee, 8-week old SCID mice were injected with 1 million cells (in 200 μl PBS) via tail vein, and lung metastases determined 8 week after the injection. Lungs were collected at the time of necropsy, and gross examination of metastatic nodules was conducted. Lungs were then fixed in formalin, embedded in paraffin, sectioned at 8-10 μm, and stained with hematoxylin and eosin. With the help of pathologist, Dr. Amy Clayton, metastatic nodules in lungs were examined and recorded. Fisher's exact test was used to detect any significant relationships between dichotomous variable (metastasis or no metastasis) and mouse groups (WT, SA, and EV) using a significance level of 0.05 (Software JMP).
Patient population
A population of 184 patients with endometrial cancer was randomly selected from a total of 1,109 patients who had hysterectomy at Mayo Clinic between January 1984 and December 1996. The mean age of the 184 patients was 66.1 years (±11.3). Clinical characteristics of the patients are summarized in Table 1.
Table 1.
Clinical characteristics of patients
| Characteristics | Number (%) | |
|---|---|---|
| Stage | I-II | 142 (77) |
| III-IV | 42 (23) | |
|
| ||
| Depth of Myometrial Invasion | None | 37 (20) |
| Any | 147 (80) | |
|
| ||
| Histologic Grade | 1 | 65 (35) |
| 2 | 75 (41) | |
| 3 | 44 (24) | |
|
| ||
| Histologic Subtype | Endometrioid or Mucinous | 171 (93) |
| Non-endometrioid | 13 (7) | |
Tissue microarray block construction
After approval was obtained from the Mayo Clinic Institutional Review board, endometrial cancer tumor specimens were used to construct the array. Histologic sections were screened and areas of representative tumor were marked. Using an automated construction device, three tissue cores (0.6 mm in diameter) were taken from each tumor sample and placed in a new 240-capacity recipient paraffin block. Liver cores were used as fiducial markers and controls for immunohistochemical reactions. 4-μm sections were then cut from the TMA paraffin block and transferred to a slide for immunohistochemical staining.
Immunohistochemistry
The specificity of affinity-purified polyclonal antibodies for HtrA1 was described in previous studies (3). As per the guidelines described by De Luca et al (16), immunohistochemical staining for HtrA1 was performed incubating the TMA sections with polyclonal rabbit anti-human HtrA1 antibody at 1:50 dilution.
TMA Digital Analysis
Using the Bliss “Virtual Microscopy” microscope and computer system (Bacus Laboratories) digital image analysis was performed after IHC.
Two reviewers (SAM and gynecologic pathologist, AC) independently scored each tissue core based on intensity of HtrA1 expression. Cells with golden-brown membranous and/or cytoplasmic staining were considered HtrA1 positive. HtrA1 expression was scored based on intensity of staining with a score of 0=none, 1=weak, 2=moderate, 3=strong staining. As triplicate cores were taken from each tumor specimen, tumor core scores were averaged to determine an overall HtrA1 intensity staining score for each patient sample. A separate dependent collaborator (AM) retrieved the clinical data.
Analysis of HtrA1 expression and clinical parameters in endometrial cancer
To explore the relationship of staining with clinical outcome, the four possible outcomes for staining (0, 1, 2, and 3) were dichotomized into two groups, high (intensity score of 2 or 3) versus low (intensity score of 0 or 1) HtrA1 expression. Fisher's exact test was used to detect any significant relationships between these dichotomous variables and clinical parameters including stage, grade, myometrial invasion, histologic subtype using a significance level of 0.05 (Software JMP). Kaplan-Meier estimates were used to determine association of HtrA1 expression (high or low) with progression free and overall survival (Software JMP).
Results
Expression pattern of HtrA1 in normal endometrium and endometrial cancer cell lines
Although HtrA1 is known to be expressed in a wide range of normal human tissue including proliferative endometrium and first-trimester placenta (16, 17), establishment of its expression pattern in EC cell lines is limited. Immunohistochemical staining on proliferative endometrium revealed HtrA1 is expressed in the normal tissue (Figure 1A). Immunoblot analysis of HtrA1 expression in endometrial cancer cell lines showed loss of HtrA1 expression in more than 50% (7/13) of these cell lines. Interestingly, there is universal loss of HtrA1 expression in the uterine papillary serous cell lines, an aggressive histologic subtype of endometrial cancer (Figure 1B). On the other hand, Western blot analysis of the six endometrioid-type endometrial cancer cell lines showed variable levels of HtrA1 expression (Figure 1B).
Figure 1.

HtrA1 is expressed in normal endometrium, with varying degrees in endometrioid adenocarcinomas and lost in uterine papillary serous carcinoma. (A) Immunohistochemistry shows htrA1 is expression in normal endometrium (arrows). (B) Western blot analysis of HtrA1 expression in endometrial cell lines with β-actin loading control (lower panel).
Downregulation of HtrA1 promotes invasion, in vitro
One of the hallmarks of advanced and/or metastatic carcinoma is the ability of tumor cells to invade and infiltrate surrounding tissue as well as spread to distant sites. Loss of expression of HtrA1 has been associated with tumors of greater metastatic potential (13, 14, 18). Induction of HtrA1 expression in metastatic melanoma cell lines has been shown to inhibit cell proliferation and invasion (13). To establish what effect downregulation of HtrA1 has on cellular migratory and invasive capacity of EC cell lines, HtrA1 expression was downregulated with siRNA in Hec1A and Hec1B which express endogenous HtrA1 (Figure 1B). HtrA1 was downregulated by transient transfection with HtrA1 siRNA (1900si) or scrambled siRNA (1900scr) for 48 hours. Downregulation of HtrA1 expression by siRNA was validated by Western blot analysis (Insets in Figures 2A and B). Transwell migration assays using fetal bovine serum as the chemo-attractant were then performed with the HtrA1 downregulated and control cells for 12 hours. The migratory capacities of Hec1A and Hec1B cells were not affected by downregulation of HtrA1 (siRNA) as compared to scrambled siRNA controls (Figure 2A & B). To determine the effect of HtrA1 down-regulation on the ability of these cells to invade, Matrigel invasion assays were performed, again using fetal bovine serum as the chemo-attractant over a 24-hour time period. The HtrA1-silenced (siRNA) Hec1A and Hec1B cells exhibited 3-4 fold higher invasive ability compared to the scrambled siRNA-control cells (Figure 2A & B). These data indicate that loss of HtrA1 expression promotes invasion but not migration in Hec1A and Hec1B EC cell lines. However, stable downregulation of HtrA1 with two different shRNAs, shRNA 1 and shRNA 2 (Figures 2C and D) in Hec1A and Hec1B cells resulted in increased cell migration as determined by scratch assay.
Figure 2.
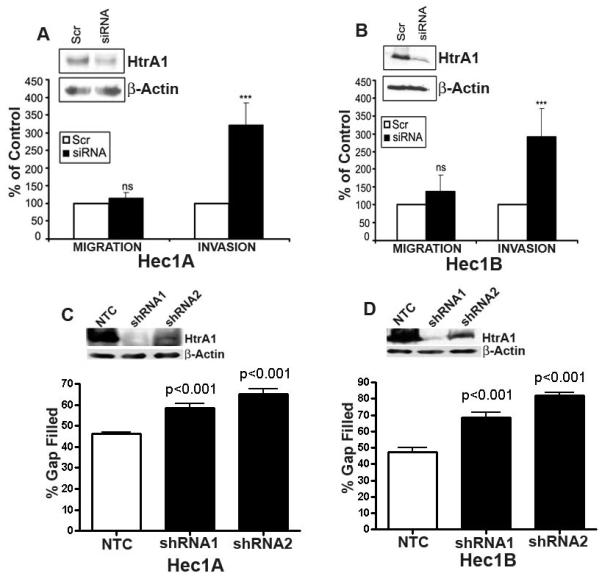
Downregulation of HtrA1 in Hec1A and 1B cells promoted invasion. (A and B) Results from matrigel invasion assay show increased number of Hec1A and Hec1B cells invading through the basement membrane (expressed as % of control cells) in scr control compared to HtrA1siRNA transfected cells respectively. There was minimal effect on the migratory capacity. Insets in (A and B) shows efficient downregulation of HtrA1 with HtrA1 siRNA compared to scr control by immunoblot analysis with β-actin loading control (lower panel) in Hec1A and Hec1B cells respectively. NS- Not significant, **P =<0.01 and *P=<0.05 between HtrA1 siRNA and scr control. (C and D) Results from scratch assay of Hec1A and Hec1B with two different shRNAs against HtrA1. The insets in C and D show the levels of stable downregulation of HtrA1 In these cells.
Forced overexpression of HtrA1 inhibits migration & invasion, in vitro
With the establishment that HtrA1 downregulation enhanced cell invasion, the effect of ectopic expression of HtrA1 was then investigated. Two uterine papillary serous cell lines which lack endogenous HtrA1, Ark1 and Ark2 (Figure 1B), were used for generating stable clones expressing wild-type HtrA1 (WT), catalytic site mutant HtrA1 (SA), or empty vector (EV). HtrA1 expression was validated by Western immunoblot analysis (Ark 1, Figure 3A, inset; Ark 2, Figure 3B, inset). These stable clones were used to perform transwell migration assays using fetal bovine serum as the chemo-attractant to investigate the effect of forced expression of HtrA1 expression on cellular migration. The migratory capacity of Ark1 and Ark2 cells was inhibited by forced overexpression of HtrA1 (wild-type WT clones) as compared to EV-controls (Figures 3A & B) or protease mutant HtrA1 (SA) clones (Figure 3B). To determine the effect HtrA1 expression had on the ability of these cells to invade, Matrigel invasion assays were performed, again using fetal bovine serum as the chemo-attractant over a 24-hour time period. The ability of HtrA1-expressing (WT) Ark1 and Ark2 clones to invade was inhibited by 60% compared to the invasive ability of EV (Figure 3A, B) and SA control clones (Figure 3B).
Figure 3.

Exogenous expression of HtrA1 inhibits migration and invasion of Ark 1 and Ark 2 cells in vitro. Results from matrigel invasion and migration assay show decreased number of Ark 1 and Ark 2 cells invading through the basement membrane (expressed as % of control cells) in with WT HtrA1 expressing cells compared to EV or SA mutant HtrA1 transfected controls respectively. Insets in (A and B) shows the expression of WT HtrA1 in Ark 1 clones 11 and 14 and Ark 2 clones 13, 16,and 19. Mutated HtrA1 in SA clones 17 and 19 in Ark 2 cells and absence of expression in Ark 1 and Ark 2 EV1 and EV3 cells by immunoblot analysis with β-actin loading control (lower panel) respectively. NS- Not significant, **P =<0.01 and *P=<0.05 between HtrA1 WT and Vector (V1) controls.
No significant difference in proliferation rate was observed in MTT reduction assay of clonal lines with and without HtrA1 expression over a period of 8 days (Data not shown). This suggests that the increased invasion and migration of HtrA1 non-expressing cells was not due to increased proliferation rate of the cells. Collectively, these results indicate that exogenous expression of HtrA1 significantly inhibits both migration and invasion in EC cancer cell lines.
HtrA1 expression decreases lung metastasis of ARK-2 Cells in SCID Mice
In an in vivo experimental metastasis model, exponentially growing HtrA1 WT, SA mutant and V1 stable clones of Ark2 cells (Figure 4A) were injected into the tail veins of 8-week old female SCID mice and metastasis to the lung was examined. Although lung is not a typical site for implantation of papillary serous and endometrial adenocarcinomas, Ark2 cells were able to form micrometastases in the lungs of mice (Figure 4B). 7 of 11 mice injected with vector clonal group (no HtrA1 expression) developed lung micrometastases in 8 weeks period, while only 1 of 11 mice injected with wild type HtrA1 clone, and 2 of 11 in SA mutant clones developed lung micrometastases (Figure 4C). Immunohistochemical staining for HtrA1 in WT and SA clones showed the presence of HtrA1, while all of the V clones was negative suggesting that these tumor cells did not express HtrA1 (Fig 4B lower panel). This strongly supports the role of HtrA1 as a metastatic tumor suppressor.
Figure 4.
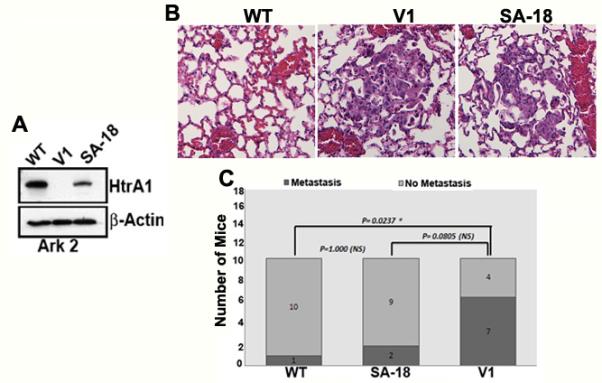
Exogenously expressed HtrA1 inhibits lung metastases of endometrial cancer cells. (A) Western blot analysis of HtrA1 expression in Ark 2 clonal lines stably transfected with wild type HtrA1 (WT), protease mutant HtrA1 (SA 18) or empty vector (V1). (B) Representative figure showing micrometastasis of Ark 2 cells in the lung in V1 and SA 18 clonal lines but not in WT clone (Top Panel). (C). Metastasis to lung was attenuated in Ark 2 cells expressing wild type HtrA1 compared to control Ark 2 cells transfected with empty vector. P-values were calculated by Pearson χ2 analysis.
To further confirm our hypothesis, we generated HtrA1 knocked-down pooled stable clones in Hec1B endometrial cancer cells which express endogenous HtrA1 (Figure 1B), using lentiviral-based shRNA targeted against HtrA1. Non-target shRNA transduced control batch clones were also selected. HtrA1 knocked down pooled stable clones (shRNA1) with significant downregulation of HtrA1 and its corresponding non-target control (NTC) (Figure 5A) were injected into the tail vein of SCID mice as before. 5 weeks later, the mice were sacrificed, dissected, and lungs were harvested and processed. Grossly visible metastatic nodules were noted on all lung tissues obtained from mice injected with shRNA HtrA1 knockout clones (shRNA11) cells (Figure 5B, lower panel). In contrast, lungs from mice injected with HtrA1 expressing clones (NTC) had minimal or no visible metastatic nodules. Histological examination of the H&E stained sections of the lung tissue also supported this conclusion (Figure 5B, top panel). Collectively, these data clearly shows that loss of HtrA1 in endometrial cancer cells is associated with increased propensity of these cells to metastasize.
Figure 5.
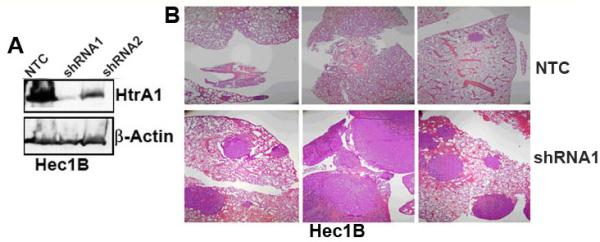
Downregulation of endogenous HtrA1 in Hec1B promotes lung metastasis of endometrial cancer cells. (A) Western blot analysis of HtrA1 expression in Hec1B clonal lines stably transfected with non-targeting shRNA (NT) or with two different shRNAs targeting HtrA1 (shRNA 1 and shRNA 2). (B) Hematoxylin and Eosin staining of lung tissue from mice injected via tail vein with stable pool of Hec1B cells expressing non-targeted shRNA (NT) or HtrA1-targeted shRNA (shRNA1).
HtrA1 expression is lost in high-grade versus low-grade endometrial cancer
Having established that loss of HtrA1 expression is associated with enhanced invasive ability while forced expression of the protein inhibits migration and invasion, in vitro and in vivo, next we wanted to correlate this observation clinically. Using tissue microarrays containing triplicate core biopsies from the tumors of endometrial cancer patients, immunohistochemical analysis for HtrA1 was performed. HtrA1 expression was scored by two independent, blinded individuals based on intensity of staining with a score of 0=none, 1=weak, 2=moderate, 3=strong staining. Concordance analysis using JMP (version 8), we found 90% agreement among triplicates with an overall kappa of 0.91.
Clinical outcomes including stage, grade, myometrial invasion, histologic subtype, disease-free and overall survival were correlated with intensity of HtrA1 expression. The four possible outcomes for staining (0, 1, 2, and 3) were dichotomized into two groups, high (intensity score of 2 or 3) versus low (intensity score of 0 or 1) HtrA1 expression (Figure 6A). Fisher's exact test was used to detect any significant relationships between these dichotomous variables and clinical parameters. 184 primary endometrial tumors were available for analysis. HtrA1 was absent or weak in 108/184 (57.1%) of tumors while protein expression was moderate or strongly expressed in 79/184 (42.8%) of tumors. When tumor expression level of HtrA1 was correlated with clinical parameters (Table 2), only high-grade (Grade 3) tumors were associated with a statistically significant loss of HtrA1 expression compared to low grade (Grades 1-2) at a p-value of 0.016. Interestingly, there was a trend towards lower HtrA1 expression levels with increasing myometrial invasion.
Figure 6.

Immunohistochemical analysis of HtrA1 expression in tissue microarrays. (A) A representative endometrial cancer tissue showing absence of HtrA1 staining. (B) A representative endometrial cancer tissue showing intense HtrA1 staining.
Kaplan-Meier survival analysis was performed to evaluate HtrA1 expression level with progression-free and overall patient survival (C and D respectively). Although not statistically significant, there is a trend towards improved PFS and OS in patients with tumor expressing high levels of HtrA1 compared to low levels based on IHC.
Table 2.
Association of HtrA1 expression level with clinical parameters
| Clinical Parameters | Patient No. (%) |
HtrA1 High No. (%) |
HtrA1 Low No. (%) |
p-value |
|---|---|---|---|---|
| Overall | 184 | 79 (42.9) | 105 (57.1) | |
|
| ||||
| Stage | ||||
| I | 136 | 57 | 79 | |
| II-IV | 48 | 22 | 26 | 0.64 |
|
| ||||
| Myometrial Invasion | ||||
| ≤ 50% | 146 | 66 | 80 | |
| > 50% | 38 | 13 | 25 | 0.22 |
| < 66% | 151 | 69 | 82 | |
| ≥ 66% | 33 | 10 | 23 | 0.11 |
|
| ||||
| Histologic Grade | ||||
| 1-2 | 140 | 67 | 73 | |
| 3 | 44 | 12 | 32 | 0.016 |
|
| ||||
| Histology Type | ||||
| Endometriod | 164 | 67 | 97 | |
| Non-endometrioid | 20 | 12 | 8 | 0.1 |
Discussion
Carcinoma of the endometrium, a common tumor of the female reproductive tract, presents with symptoms early in the disease process thereby lending itself to early diagnosis, treatment, and frequent cure. However, of those individuals who present with disease at an advanced stage or with metastatic, recurrent disease, subsequent treatment and prognosis is limited. The majority of mortality from endometrial carcinoma, unfortunately, arises from a small percentage of patients who present with this advanced stage disease. Identifying those patients, therefore, who are at risk for metastatic disease could offer opportunities for targeted surveillance and alterations in treatment. Moreover, understanding the molecular events that regulate cellular ability to locally invade and spread to distant sites provides the key to understanding development of a metastatic phenotype. One such step has been the recognition of HtrA1 as a potential metastatic tumor suppressor gene (Baldi et al 2002; Esposito et al, 2006). In lung cancer and endometrial cancer tumor specimens, decreased HtrA1 is associated with higher grade tumor (14, 18). Additionally, lymph node metastases and metastatic lesions from primary lung and melanoma, respectively, express lower levels of HtrA1 than the primary sites (13, 18). In vitro assessment has shown that upregulation of HtrA1 protein in a malignant melanoma cell line inhibits cell proliferation while exogenous expression induces cell death in an ovarian cancer cell line with subsequent downregulation promoting anchorage-independent growth (12, 13, 18). We have further expanded these previous observations by providing in vivo data demonstrating the metastatic tumor suppression of HtrA1 in endometrial cancer cell lines.
Previously published work has shown the HtrA1 is highly expressed in the normal, proliferative endometrium (16). Through immunohistochemical staining of benign, proliferative endometrium, our work confirms the expression of HtrA1 in normal endometrium. Further screening of established endometrial cancer cell line shows an interesting pattern of HtrA1 expression. There is universal absence of expression of HtrA1 in the uterine papillary serous cell lines while all of the endometrioid-type endometrial cancer cell lines express HtrA1 to varying degrees. Translating this to the clinical situation, uterine papillary serous carcinoma is a highly aggressive, estrogen-independent Type II endometrial carcinoma which arises in a background of atrophic endometrium, appears histologically similar to and mimics the behavior of papillary serous carcinoma of the ovary (19). On the other hand, the more common endometrioid-type endometrial adenocarcinoma is an estrogen-dependent Type I endometrial cancer which arises from a background of endometrial hyperplasia, is typically low-grade and diagnosed at an early stage. There is a well-documented, clear progression from normal endometrium to atypical endometrial hyperplasia, to invasive endometrioid-type adenocarcinoma (19). Universal expression of HtrA1 in type I endometrial cancer cell lines which most closely resemble normal proliferative endometrium versus universal absence of HtrA1 in type II endometrial cancer cell lines which more closely resemble serous ovarian carcinoma than normal endometrium may be more reflective of the molecular differences between these two tumor types than a true reflection of the impact of HtrA1. These results are consistent with our unpublished observation that HtrA1 expression is regulated by steroid hormones.
On the other hand, the true functional significance of HtrA1 has yet to be elucidated. There is growing evidence to support a role of HtrA1 in metastatic tumor potential but its mechanism of action is unknown. In an effort to understand the role of HtrA1, in general, and more specifically, its impact in endometrial cancer, we explored the impact of this protein through forced upregulation and downregulation. In order to address the potential biological significance of the loss of expression of HtrA1 in tumor cells, HtrA1 was constitutively downregulated in two endometrioid-type endometrial cancer cell lines, Hec1A and Hec1B. Initial immunoblot analysis had established that these cell lines expressed high levels of endogenous HtrA1. Downregulation of HtrA1 in Hec1A and Hec1B after transient transfection increased invasion through Matrigel but did not enhance Transwell migration. However, we observed significant increase in cell migration following stable knockdown of HtrA1 in the same cell lines, suggesting that stable knockdown of HtrA1 may lead adaptation of cells to a more migratory phenotype which was not observed with transient knockdown. Conversely, expression of HtrA1 was induced in two uterine papillary serous cell lines, Ark1 and Ark2, which immunoblot analysis had established did not express endogenous HtrA1. Stable Ark1 and Ark2 clones expressing HtrA1 showed inhibition of Transwell migration and Matrigel invasion compared to stable empty vector clones or clones expressing catalytically inactive mutant HtrA1 (SA). Therefore, suppression of HtrA1 appears to contribute to a metastatic phenotype through enhanced ability of a cell to migrate and invade, in vitro. The discordant effect of HtrA1 on cell migration in endometrial cancer cell lines could be the results of differences in genetic backgrounds of type I and type II carcinomas or the artificial results of forced expression in Ark1 and Ark2. Nonetheless, we observed consistent effect of HtrA1 on Matrigel invasion in all four cell lines tested, i.e., down-regulation of HtrA1 promotes invasion whereas upregulation of HtrA1 inhibits invasion.
Since expression of HtrA1 appeared to inhibit migration and invasion of endometrial cancer cells in vitro, we further defined the impact of HtrA1 expression on in vivo metastatic models and on the clinical outcome of patients with primary endometrial cancer. We found a statistically significant increase in metastatic potential of endometrial cancer cell lines with decreased HtrA1 expression. Consistent with these results, forced expression of HtrA1 in endometrial cancer cell lines decreases metastatic potential of endometrial cancer cells. Interestingly, lung micrometastases we observed in one mouse injected with Ark2 cell line stably transfected with wild-type HtrA1 continued to express HtrA1. It is likely that these cells escape HtrA1-mediated suppression of lung metastasis as a result of alterations in downstream molecular pathways mediated by HtrA1. We also observed reduced lung metastasis in Ark2 cell line stably transfected with protease wild-type HtrA1. These results suggest that protease activity is important in regulating specific steps in hematogenous metastasis. It should be noted that reduced lung metastasis was also observed in mice injected with Ark2 cell stably transfected with protease mutant HtrA1, albeit it does not rise to the levels of statistical significance. Current study may not have sufficient power to detect the significant differences in metastatic behavior produced by protease mutant HtrA1. Finally, we observed a statistically significant association of HtrA1 downregulation in primary tumors with increased grade of tumor. Additionally, there was a trend towards increased overall survival and prolonged progression-free survival in patients with tumor expressing high levels of HtrA1 compared to low levels. However, the overall good prognosis of endometrial cancer with limited recurrence events limits the overall conclusions that can be drawn from this analysis. It does, however, correlate with findings reported by Bowden et al illustrating down-regulation of HtrA1 with increasing grades of endometrial cancer. In agreement with our in vitro studies showing that loss of HtrA1 enhances cell invasion was the observation of a trend towards decreased HtrA1 expression in tumors with a greater percentage of myometrial invasion.
The finding that HtrA1 downregulation corresponds to poor clinical outcome and is associated with increased tumorigenesis and metastatic potential in vitro and in vivo, strengthens the previous observation that loss of HtrA1 expression is associated with metastatic tumors in melanoma and lung cancer (13, 18). Further studies to elucidate the underlying mechanisms associated with HtrA1 downregulation and metastasis can be explored. It is hoped that an understanding of the molecular mechanisms resulting in metastatic disease, a major contributor to cancer deaths, in general, and mortality from endometrial cancer, specifically, will lead to the successful development of therapies which inhibit this process.
STATEMENT OF TRANSLATIONAL RELEVANCE.
A better understanding of the molecular mechanisms associated with metastatic disease, a major contributing factor for cancer-associated mortality, may facilitate the successful development of therapies that inhibit metastatic processes and reduce the risk of recurrence. Our finding that HtrA1 is significantly down-regulated in high-grade endometrial cancer (EC) compared to low-grade EC supports the translational relevance of HtrA1 as a potential biomarker in aiding tumor grade classification. In addition, the fact that HtrA1 down-regulation is associated with increased tumorigenesis and metastatic potential in vitro and in vivo, is consistent with its putative role as a metastatic tumor suppressor. Enhanced expression of HtrA1 by epigenetic agents, anti-microRNAs, small molecules, and viral-mediated gene delivery approaches that upregulate HtrA1 expression in target cells could become a viable treatment strategy. Such approaches could potentially be useful as a risk reduction prophylaxis and may be amenable to patients at high-risk for recurrent or metastatic disease.
Acknowledgements
We thank the members of the Shridhar laboratory for stimulating discussions.
FUNDING
This work was supported by grants from National Cancer Institute, 1R01CA123249 (to VS and JC), the Mayo Clinic Bernard and Edith Waterman Center for Cancer Genetics and the Mayo Foundation (to V.S.).
REFERENCES
- 1.Zumbrunn J, Trueb B. Primary structure of a putative serine protease specific for IGF- binding proteins. FEBS Lett. 1996;398:187–92. doi: 10.1016/s0014-5793(96)01229-x. [DOI] [PubMed] [Google Scholar]
- 2.Clausen T, Southan C, Ehrmann M. The HtrA family of proteases: implications for protein composition and cell fate. Mol Cell. 2002;10:443–55. doi: 10.1016/s1097-2765(02)00658-5. [DOI] [PubMed] [Google Scholar]
- 3.Chien J, Aletti G, Baldi A, et al. Serine protease HtrA1 modulates chemotherapy-induced cytotoxicity. J Clin Invest. 2006;116:1994–2004. doi: 10.1172/JCI27698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Faccio L, Fusco C, Chen A, Martinotti S, Bonventre JV, Zervos AS. Characterization of a novel human serine protease that has extensive homology to bacterial heat shock endoprotease HtrA and is regulated by kidney ischemia. J Biol Chem. 2000;275:2581–8. doi: 10.1074/jbc.275.4.2581. [DOI] [PubMed] [Google Scholar]
- 5.Gray CW, Ward RV, Karran E, et al. Characterization of human HtrA2, a novel serine protease involved in the mammalian cellular stress response. Eur J Biochem. 2000;267:5699–710. doi: 10.1046/j.1432-1327.2000.01589.x. [DOI] [PubMed] [Google Scholar]
- 6.Liu ML, Liu MJ, Kim JM, Kim HJ, Kim JH, Hong ST. HtrA2 interacts with A beta peptide but does not directly alter its production or degradation. Molecules and cells. 2005;20:83–9. [PubMed] [Google Scholar]
- 7.Pallen MJ, Wren BW. The HtrA family of serine proteases. Mol Microbiol. 1997;26:209–21. doi: 10.1046/j.1365-2958.1997.5601928.x. [DOI] [PubMed] [Google Scholar]
- 8.Hu SI, Carozza M, Klein M, Nantermet P, Luk D, Crowl RM. Human HtrA, an evolutionarily conserved serine protease identified as a differentially expressed gene product in osteoarthritic cartilage. J Biol Chem. 1998;273:34406–12. doi: 10.1074/jbc.273.51.34406. [DOI] [PubMed] [Google Scholar]
- 9.Grau S, Richards PJ, Kerr B, et al. The role of human HtrA1 in arthritic disease. The Journal of biological chemistry. 2006;281:6124–9. doi: 10.1074/jbc.M500361200. [DOI] [PubMed] [Google Scholar]
- 10.Yang Z, Camp NJ, Sun H, et al. A variant of the HTRA1 gene increases susceptibility to age-related macular degeneration. Science. 2006;314:992–3. doi: 10.1126/science.1133811. [DOI] [PubMed] [Google Scholar]
- 11.Bakay M, Zhao P, Chen J, Hoffman EP. A web-accessible complete transcriptome of normal human and DMD muscle. Neuromuscul Disord. 2002;12(Suppl 1):S125–41. doi: 10.1016/s0960-8966(02)00093-7. [DOI] [PubMed] [Google Scholar]
- 12.Chien J, Staub J, Hu SI, et al. A candidate tumor suppressor HtrA1 is downregulated in ovarian cancer. Oncogene. 2004;23:1636–44. doi: 10.1038/sj.onc.1207271. [DOI] [PubMed] [Google Scholar]
- 13.Baldi A, De Luca A, Morini M, et al. The HtrA1 serine protease is down-regulated during human melanoma progression and represses growth of metastatic melanoma cells. Oncogene. 2002;21:6684–8. doi: 10.1038/sj.onc.1205911. [DOI] [PubMed] [Google Scholar]
- 14.Bowden MA, Di Nezza-Cossens LA, Jobling T, Salamonsen LA, Nie G. Serine proteases HTRA1 and HTRA3 are down-regulated with increasing grades of human endometrial cancer. Gynecol Oncol. 2006;103:253–60. doi: 10.1016/j.ygyno.2006.03.006. [DOI] [PubMed] [Google Scholar]
- 15.Chien J, Ota T, Aletti G, et al. Serine protease HtrA1 associates with microtubules and inhibits cell migration. Molecular and cellular biology. 2009;29:4177–87. doi: 10.1128/MCB.00035-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.De Luca A, De Falco M, Severino A, et al. Distribution of the serine protease HtrA1 in normal human tissues. J Histochem Cytochem. 2003;51:1279–84. doi: 10.1177/002215540305101004. [DOI] [PubMed] [Google Scholar]
- 17.Nie GY, Li Y, Minoura H, et al. A novel serine protease of the mammalian HtrA family is up-regulated in mouse uterus coinciding with placentation. Mol Hum Reprod. 2003;9:279–90. doi: 10.1093/molehr/gag036. [DOI] [PubMed] [Google Scholar]
- 18.Esposito V, Campioni M, De Luca A, et al. Analysis of HtrA1 serine protease expression in human lung cancer. Anticancer Res. 2006;26:3455–9. [PubMed] [Google Scholar]
- 19.Lax SF, Kurman RJ. A dualistic model for endometrial carcinogenesis based on immunohistochemical and molecular genetic analyses. Verhandlungen der Deutschen Gesellschaft fur Pathologie. 1997;81:228–32. [PubMed] [Google Scholar]