

Abstract
Objective
The current study tested whether neural activity in response to messages designed to help smokers quit could predict smoking reduction, above and beyond self-report.
Design
Using neural activity in an a priori region of interest (a subregion of medial prefrontal cortex [MPFC]), in response to ads designed to help smokers quit smoking, we prospectively predicted reductions in smoking in a community sample of smokers (N = 28) who were attempting to quit smoking. Smoking was assessed via expired carbon monoxide (CO; a biological measure of recent smoking) at baseline and 1 month following exposure to professionally developed quitting ads.
Results
A positive relationship was observed between activity in the MPFC region of interest and successful quitting (increased activity in MPFC was associated with a greater decrease in expired CO). The addition of neural activity to a model predicting changes in CO from self-reported intentions, self-efficacy, and ability to relate to the messages significantly improved model fit, doubling the variance explained ( ).
Conclusion: Neural activity is a useful complement to existing self-report measures. In this investigation, we extend prior work predicting behavior change based on neural activity in response to persuasive media to an important health domain and discuss potential psychological interpretations of the brain–behavior link. Our results support a novel use of neuroimaging technology for understanding the psychology of behavior change and facilitating health promotion.
Keywords: functional magnetic resonance imaging (fMRI), neuroimaging, behavior change, smoking
We are exposed to messages designed to change our health behaviors nearly every day; parents and friends may encourage us, doctors may instruct us, and public health media campaigns try to persuade us to make healthier lifestyle choices. In attempting to understand the circumstances under which these types of messages are likely to result in behavior change, prominent persuasion and behavior change theories have demonstrated that people’s self-reported intentions and self-efficacy predict message-consistent behavior change (Ajzen, 1991; Chaiken, Liberman, & Eagly, 1989; Fishbein & Ajzen, 1975; Fishbein et al., 2001; Petty & Cacioppo, 1986; Rosenstock, 1966; Strecher & Rosenstock, 1997). However, models based on self-report measures have limitations. For example, one recent meta-analysis reported that the theory of planned behavior accounted for 27% of the variability in people’s behavior, leaving nearly three quarters of the variability unexplained (Armitage & Conner, 2001), and a separate meta-analysis reported a slightly weaker average intention– behavior link (Webb & Sheeran, 2006). Presumably, predictive efficacy is partially limited by moderating events and changing circumstances that occur after predictions can be made and thus cannot be accounted for in such models. Nevertheless, it is also likely that a share of unexplained variance could be captured with new measurement instruments that do not share some of the biases associated with self-report methods.
Neuroimaging as a Tool to Predict Behavior Change
Some of the difficulty in predicting behavior change following persuasive messages, and consequently identifying messages that are likely to have the greatest impact on health behavior change, may stem from limitations in people’s ability to produce accurate self-reports in the context of laboratory or focus group settings or in retrospective surveys. For example, limitations and biases may arise because of social desirability effects (Booth-Kewley, Larson, & Miyoshi, 2007; Edwards, 1953) or when people do not have conscious access to the factors that lead them to change their behaviors (Nisbett & Wilson, 1977). Furthermore, when asked, people may confabulate reasons that did not actually factor in the behavior change (Wilson & Nisbett, 1978). Likewise, introspection during stimulus presentation may change the experience (Wilson & Schooler, 1991).
Among methods for circumventing such challenges (Hurlburt & Heavey, 2001; Nederhof, 2006), functional magnetic resonance imaging (fMRI) provides a way of monitoring neural responses to persuasive messages in the moment that effects initially take hold (Lieberman, 2010). As such, it provides a complementary data source on how people process messages (Chua, Liberzon, Welsh, & Strecher, 2009; Chua, Polk, Welsh, Liberzon, & Strecher, 2009) that may allow us to more accurately predict behavior change following message exposure (Falk, Berkman, Harrison, Mann, & Lieberman, 2010). Neuroimaging has been used successfully in past investigations to link neural activity during cognitive tasks to changes in substance use behaviors (Brewer, Worhunsky, Carroll, Rounsaville, & Potenza, 2008; Kosten et al., 2006; Paulus, Tapert, & Schuckit, 2005), as well as lower level attention, memory, and reward processes involved in smoking and smoking cessation (Brody et al., 2002, 2007; Mendrek et al., 2006; Weinstein & Cox, 2006; Xu et al., 2007), but has not previously been used to predict changes in smoking reduction in response to persuasive messages. In the context of assessing responses to persuasive messages, understanding the neural precursors of behavior change may ultimately allow us to select among messages those that are most likely to show specific, desired effects and are ultimately more effective in helping people change their behaviors.
The Present Approach
As a starting point, one recent neuroimaging study demonstrated that neural activity during initial exposure to persuasive messages can predict variability in behavior change that is not predicted by self-report measures such as attitudes and intentions (Falk et al., 2010). More specifically, Falk and colleagues (2010) reported that activity in a neural region associated with self-related processing (a subregion of medial prefrontal cortex [MPFC]) was associated with changes in sunscreen use from the week before the scan to the week after, above and beyond people’s attitudes and intentions to use sunscreen.
In the present study, we extended this work to the context of antitobacco messages and subsequent smoking reduction. This represents an important step forward theoretically, given that smoking reduction is a more motivationally relevant and complex behavior change than increased sunscreen use. This also represents an important practical step, given that smoking is the leading cause of preventable morbidity and mortality in the United States and as such is a public health priority (Centers for Disease Control and Prevention, 2008, 2010; Office of the Surgeon General, 2004). Evidence suggests that mass media campaigns can be used to facilitate quitting (National Cancer Institute, 2008; Popham et al., 1993; Vallone et al., 2010). However, at present the underlying mechanisms that lead messages to be successful are not fully understood.
From a theoretical standpoint, we were also interested in whether neural activity could predict independent variance above and beyond self-report measures. We chose to examine intentions and self-efficacy as self-report variables that have been demonstrated to predict substantial variance in behavior change and are present in several major theories of health behavior change (Fishbein et al., 2001). Interrogating neural activity simultaneously with known self-report precursors of behavior change (e.g., intention and self-efficacy) will not only allow us to gain a deeper understanding of the brain– behavior link, but may also help us understand the interrelationships between these psychological constructs and behavioral outcomes of interest.
In addition, given that Falk and colleagues (2010) speculated that activity in MPFC might index either an explicit or implicit connection between persuasive messages designed to change behavior and the self, we included a measure of participants’ explicit ability to relate to the messages. This measure may help us narrow our understanding of whether the variability in behavior change explained by neural activity in our MPFC region of interest (ROI) is reducible to information obtainable through explicit self-report.
Lastly, self-reports of smoking behavior (either through global recall or through timeline follow-back) are prone to a number of cognitive biases (Hammersley, 1994; Pierce, 2009; Shiffman, 2009). For example, smokers exhibit a tendency to report that they have smoked numbers of cigarettes that cluster around certain round numbers (e.g., 10, 20, the number of cigarettes in a pack), even though this is not reflective of actual behavior (Klesges, Debon, & Ray, 1995). Likewise, bias may arise because of selfpresentation concerns or other factors such as people’s motivation to appear consistent with their stated intentions (Cialdini & Goldstein, 2004). Therefore, although having limitations of its own (discussed in the Limitations section of this article), we elected to use a biological indicator of recent smoking (expired carbon monoxide [CO]) as our primary dependent variable.
By combining data obtained through fMRI, self-report surveys, and biological indicators of cigarette smoking, we were able to link neural responses during exposure to health messages to the real-world behaviors that follow. More specifically, we demonstrate that activity in an a priori region of MPFC can be used to explain variability in expired CO in the context of smoking reduction.
Method
Participants
Thirty-one right-handed participants (15 women) were recruited from the American Lung Association’s Freedom From Smoking program, an 8-week professional-led, group-based smoking cessation program, that was standard to all subjects (aside from recruitment, there was no connection between the program and this study). Two male participants were excluded from behavioral follow-up analyses because of missing endpoint data, and one male participant was excluded because of excessive head motion during the fMRI scanner session, resulting in a final sample of 28 participants.
All participants were heavy smokers who intended to quit. Participants were considered heavy smokers if they smoked at least 10 cigarettes per day, 7 days per week, for at least 1 year, and had urinary cotinine levels of at least 1,000 ng/ml. On average, participants smoked 21 cigarettes per day (SD = 10.0). In addition to enrollment in a cessation program, quitting intentions were assessed via scores greater than 9 of 10 on the Contemplation Ladder, a single-item measure of intentions to quit (Biener & Abrams, 1991), thus holding baseline intentions to quit relatively constant across this sample. Participants met standard criteria for fMRI scanning; participants were excluded if they were left-handed, did not speak English, were pregnant or claustrophobic, or had any other condition contraindicated for MRI. Participants were also excluded if they consumed more than 10 alcoholic drinks per week or had any of the following conditions: dependence on substances other than nicotine, dependence on substances within 1 year of the scan date, neurological or psychiatric disorders, or cardiovascular disease. Complete methodological details and behavior change data are available in Berkman, Dickenson, Falk, and Lieberman (in press).
Participants varied in age from 28 to 69 years (M = 45 years, SD = 10.1), and had been smoking from 11 to 53 years (M = 28.4 years, SD = 2.0). Participants were ethnically diverse: 50% Caucasian, 25% Hispanic, 21% African American, and 4% other; and socioeconomically diverse: participant mean annual income = $31,070 (range = $0–$200,000); 57% received some form of government assistance; 60% completed some form of college, and 28% received a bachelor’s degree or higher. Participants were paid $80 for completion of the fMRI portion of the study. All participants provided written informed consent that was approved by the University of California, Los Angeles Institutional Review Board.
Materials and Procedure
On arrival, participants gave consent and were screened for use of following illicit drugs with a urine test (Syva RapidTest d.a.u. 5, Dade Behring Inc., Cupertino, CA): amphetamines, cocaine, marijuana, opiates, and PCP.
Baseline smoking measures
Participants completed self-report measures of smoking history, nicotine dependence, cravings, and intentions to quit, as well as exhaled CO, a measure of recent smoking (Microsmokerlyzer, Bedfont Scientific Ltd., Kent, England). To minimize withdrawal effects during the scan, participants smoked a cigarette within 1 hr of the beginning of the scan.1 Next, participants received verbal instructions and completed a practice version of the scanner task. Participants also completed a variety of other questionnaires that are not relevant to the present hypotheses.
The ads task
Professionally developed TV commercials designed to help smokers quit smoking were obtained from public health agencies and foundations including the American Legacy Foundation, the California Department of Public Health, the Massachusetts Department of Public Health, and the Louisiana Public Health Institute. Discussions with experts at these agencies narrowed the pool of ads to those that would be most relevant to smokers who were trying to quit smoking. Ads focusing on prevention or that did not target current smokers were excluded. Sixteen final video-based ads that specifically targeted smokers and encouraged quitting were included. Content included testimonials focusing on danger to the smoker (Massachusetts’ “Fight for Your Life” Campaign), humorous ads emphasizing the social value of quitting (Louisiana’s Finger Puppet Campaign), the danger of second-hand smoke to others (California Department of Public Health), encouragement to relearn behaviors without cigarettes (American Legacy Foundation’s Ex Campaign: “Re- Learn”), and ads empathizing with the difficulty of quitting and suggesting resources to help (American Legacy Foundation’s Ex Campaign: “Direct Response”). All ads were 30 s long, with the exception of two ads, which were 15 s long.
fMRI procedure
Stimuli were presented using fMRI scanner compatible LCD goggles, and responses were recorded using a scanner compatible button box. Foam padding was used to reduce head motion. During the primary task, each participant watched a series of 16 ads (each of which was designed to help smokers quit smoking) while neural activity was recorded using fMRI. The order of ads was counterbalanced across subjects. Following the presentation of each ad, participants rated the extent to which the ad promoted a sense of self-efficacy (“This ad makes me feel that I can quit”), increased intentions to quit (“This ad makes me more determined to quit”), and self-relevance (“I can relate to this ad”). All ratings were made on a 4-point scale (anchors: disagree strongly, disagree somewhat, agree somewhat, agree strongly). Participants were given 4 s to make each rating. Ads and ratings were interspersed with rest periods in which participants viewed a fixation cross and were instructed to clear their minds. Additional 15-s fixation-cross rest periods were interspersed every four blocks to allow the hemodynamic response to return to baseline.
fMRI data acquisition
Brain imaging data were acquired on a 3T Siemens Trio scanner at the UCLA Ahmanson-Lovelace Brain Mapping Center. High-resolution structural T2-weighted echo-planar images (spin-echo; TR = 5,000 ms; TE = 34 ms; matrix size 128 × 128; 34 axial slices; FOV = 192 mm; 4 mm thick) were acquired coplanar with the functional scans. One functional scan lasting 11.5 min (351 volumes) was acquired during the task (echo-planar T2*-weighted gradient-echo; TR = 2,000 ms; TE = 30 ms; flip angle = 90°; matrix size 64 × 64; 34 axial slices; FOV = 192 mm; 4 mm thick).
Follow-up smoking measures
Approximately 1 month following the baseline portion of the study, an in-person follow-up was conducted in the field, including a biological verification of participants’ self-reported smoking (through expired CO). At this stage, two participants were unable to be reached.
Data Analysis
fMRI data analysis
The imaging data were preprocessed using a combination of FSL tools (FMRIB Software Library, Oxford University, Oxford, England) and SPM8 (Wellcome Department of Cognitive Neurology, Institute for Neurology, London, England). All images were brain-extracted using FSL’s Brain Extraction Tool and realigned within runs using FSL’s Motion Correction using FMRIB’s Linear Image Registration Tool, then checked for residual motion and noise spikes using a custom automated diagnostic tool (thresholded at 2-mm motion or 2% global signal change from one image to the next). At this stage, one participant was excluded because of extreme head motion. In SPM8, all functional and anatomical images were reoriented to set the origin to the anterior commissure and the horizontal (y) axis parallel to the AC–PC line. Also in SPM8, functional images were corrected for slice acquisition timing differences within volumes, realigned within and between runs to correct for residual head motion, and coregistered to the matched-bandwidth structural scan using a six-parameter rigid body transformation. The coregistered structural scan was then normalized into the Montreal Neurological Institute standard stereotactic space, and these parameters were applied to all functional images. Finally, the normalized functional images were smoothed using an 8-mm full width at half maximum Gaussian kernel. All functional imaging results are reported in Montreal Neurological Institute coordinates.
The task was modeled separately for each subject, using a blocked design in SPM5 (Wellcome Department of Cognitive Neurology, Institute for Neurology, London, England). Initial analyses modeled ad exposure compared with a rest period. Response periods were modeled as effects of no interest in the present analysis given that our primary interest lay in understanding realtime neural responses to the ads. A corresponding random effects model averaged across results at the single-subject level.
A priori ROI
The primary ROI was constructed using Marsbar (Brett, Anton, Valabregue, & Poline, 2002), based on prior work predicting behavior change from neural activity (Falk et al., 2010). The ROI encompassed a ventral subregion of MPFC that was most highly associated with behavior change in work by Falk and colleagues (2010; see Figure 1). Thus, we refined our region of interest to capitalize on the whole-brain exploratory search conducted in that prior investigation. Average parameter estimates of activity in this ROI were extracted at the group level using Marsbar.
Figure 1.

Medial prefrontal cortex (MPFC) region of interest used to predict behavior change. This particular region of MPFC was selected a priori based on prior work linking neural activity to behavior change in the context of a simpler health behavior (sunscreen use) over a 1-week period. In the current investigation, neural activity in this region also predicts smoking reduction in a group of smokers over the course of a month, above and beyond their self-reported intentions to quit, self-efficacy to quit, and ability to relate to ads designed to help people quit smoking.
Associations between neural activity during ad exposure and changes in CO
We used changes in expired CO from baseline to endpoint as our primary proxy for behavior change; change was calculated as the difference between endpoint expired CO and baseline expired CO (positive values indicate more successful quitting). To determine whether activity during ad exposure in our a priori ROI was associated with behavior change, we regressed behavior change scores onto parameter estimates of activity from the ROI. To determine whether neural activity explained variability in behavior change that was not explained by self-report measures of intention, self-efficacy, and ability to relate to each ad, we also entered these measures into a regression model, predicting behavior change from neural activity, controlling for all self-report measures collected in the scanner.
Complementary whole-brain searches were conducted with a voxel-wise threshold of p < .005 combined with a minimum cluster size of 18 in MPFC (given our a priori hypothesis) and 42 in the rest of the brain, corresponding to p < .05, False Discovery Rate (FDR) corrected for each search space, based on a Monte Carlo simulation implemented using AlphaSim in the software package AFNI (http://afni.nimh.gov/afni/doc/manual/AlphaSim).
More specifically, in these whole-brain searches, we regressed neural activity onto changes in CO, as well as changes in CO, controlling for self-report measures. A design matrix was constructed for each subject at the single-subject level comparing activity while viewing the ads to activity at rest. At the group level, activity during the ads (compared with rest) was then correlated with subsequent changes in expired CO (see Table S1, available online as supplemental material). A parallel regression was run at the group level assessing the relationship between activity during ad exposure and changes in CO, controlling for intentions, selfefficacy, and average ability to relate to the ads.
Results
Smoking Behavior Change
At baseline, participants were all heavy smokers (see recruitment criteria) and smoked an average of 21.12 (SD = 10.04) cigarettes per day, corresponding to a baseline expired CO average of 19.21 ppm (SD = 11.37). At 1-month follow-up, participants smoked an average of 5.00 (SD = 5.42) cigarettes per day, corresponding to an endpoint expired CO average of 12.07 ppm (SD = 10.72). This represented a significant decline: average change in expired CO = 7.14 ppm (SD = 14.15), t(27) = 2.672, p = .013; average change in cigarettes = 16.13 cigarettes per day (SD = 11.30), t(27) = 7.55, p < .01 (Berkman et al., in press). The correlation between self-report smoking behavior and our biological measure of expired CO was significant both at baseline (r = .44, p = .02) and endpoint (r = .48, p = .009).2
Behavioral Responses to the Ads Task
One-sample t tests comparing neutral on a 4-point scale from strongly disagree to strongly agree suggested that, on average, participants could relate to the ads: M = 2.85, SD = 0.44, t(27) = 4.225, p < .001; exposure to ads significantly reinforced participants’ intentions to quit smoking: M = 2.80, SD = 0.56, t(27) = 2.757, p = .010; and significantly increased their self-efficacy to quit: M = 2.75, SD = 0.49, t(27) = 2.712, p = .012.
Associations Between Self-Reported Intentions, Self-Efficacy, and Expired CO
The averaged values for intention and self-efficacy measures were highly correlated with one another between subjects (r = .93, p < .001), and each was significantly correlated with the degree to which participants could relate to the ads as a whole (rintention_relate = .442, p = .018; rself-efficacy_relate = .53, p = .004). Likewise, within subjects, each of the self-report measures was significantly correlated, p < .05 (the average within-subjects correlation between individual ratings of intention and self-efficacy was .62, between intention and ability to relate was .55, and between self-efficacy and ability to relate was .58). Given the extremely high degree of multicolinearity between intention and self-efficacy between subjects, a composite intention/self-efficacy measure was calculated as the average of the two measures, within subjects, for the purpose of multiple regression analyses predicting behavior change (thus “self-report” measures referred to below include a composite intention/self-efficacy variable and the relate variable). Table 1 includes correlations between each of the self-report measures and neural activity, between subjects.
Table 1.
Correlations Between Self-Report Measures of Intentions, Self-Efficacy, Ability to Relate to the Ads, and Neural Activity in Medial Prefrontal Cortex Region of Interest (MPFC ROI)
| Measure | Self-efficacy | Intentions | Ability to relate |
|---|---|---|---|
| Intentions | .927** | ||
| Ability to relate | .525** | .442* | |
| MPFC ROI | .053 | .032 | .164 |
Note. Results suggest that whereas the self-report measures are related to one another, they are each uncorrelated with estimates of neural activity across subjects.
p < .05.
p < .005.
Neural Activity During Ad Exposure Predicts Subsequent Changes in Expired CO
To determine whether neural activity in our a priori hypothesized MPFC ROI was associated with behavior change, we regressed changes in expired CO from baseline to 1 month postscan onto parameter estimates of activity during ad exposure compared with rest. Neural activity in this ROI during ad exposure significantly predicted behavior change, both before, β = .42, t(26) = 2.35, p = .027, and after, β = .45, t(24) = 2.75, p = .011, controlling for self-reported intentions to quit, self-efficacy to quit, and ability to relate to each ad (see Figure 2b) such that increased activity in MPFC was associated with greater declines in expired CO.
Figure 2.
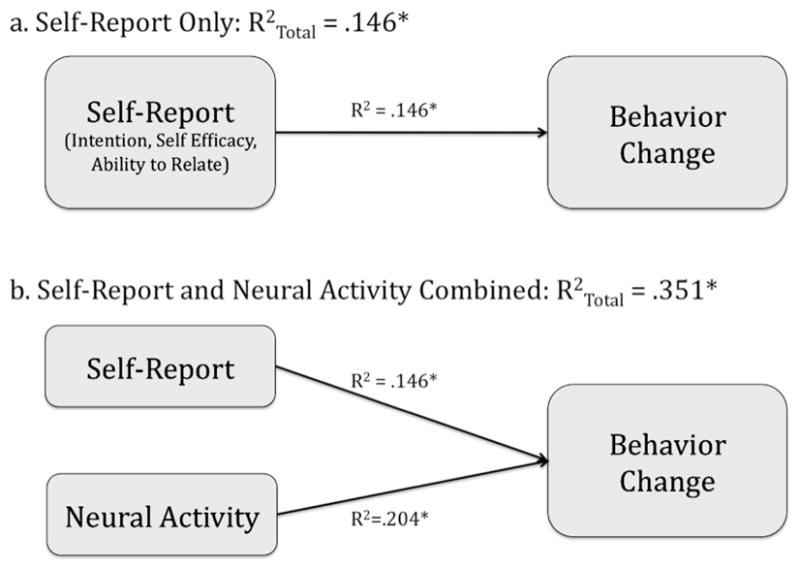
Variance explained by self-report alone and self-report combined with neural activity in medial prefrontal cortex (MPFC). (a) A regression model including self-reported intentions, self-efficacy, and ability to relate to messages predicted 14.6% of the variance in behavior change. (b) A regression model including self-report measures plus neural activity during ad exposure in MPFC predicted 35% of the variance in behavior change ( ). * p < .05.
Complementary whole-brain analyses also identified a small number of other regions associated with behavior change outside of our hypothesized MPFC ROI. Three clusters of neural activity in the second most common region implicated in self-related processing—medial precuneus/posterior cingulate; peak cluster 1: t(26) = 3.67; peak cluster 2: t(26) = 4.77; peak cluster 3: t(26) = 3.97; ps < .005—and a region involved in motor planning— supplementary motor area; t(26) = 3.75, p < .005—were the regions most highly associated with behavior change. A complete table of results, before and after controlling for self-report variables, is available in the supplemental online materials.
To determine the amount of variability in behavior change that could be explained above and beyond self-report, we tested a series of linear regression models, successively adding variables to the model. R2 values were compared for models with self-report variables alone (R = .382, R2 = .146) and with neural activity in our functionally defined ROI plus all self-report measures (R = .592, R2 = .351). This comparison revealed that an additional 20% of the variability in participants’ behavior change could be explained using neural activity, above and beyond variables accounted for in traditional models of behavior change, R2 change = .20, F(1, 24) = 7.55, p = .011. Put another way, the addition of neural activity to the model more than doubles the amount of variance explained and represents a significant improvement in the model (see Table 2; see Figure 2b). Conversely, when neural activity from our functional ROI was first entered into the model, followed by self-report predictors (intention, self-efficacy, and ability to relate to the messages), a marginally significant improvement was observed, R2 change including intention, self-efficacy, and relate variables in addition to medial prefrontal cortex = .175, F(1, 24) = 3.24, p = .057.
Table 2.
Results of Hierarchical Regression Predicting Changes in Smoking as Indicated by Expired Carbon Monoxide (CO), From Self-Report Measures Alone (Intentions, Self-Efficacy, Ability to Relate to Ads), and Then Combining Self-Report Measures With Neural Activity in an A Priori Defined Region of Medial Prefrontal Cortex
| Step | R | R2 | df error | F | Significant model | R2 change | F change | Significant F change |
|---|---|---|---|---|---|---|---|---|
| 1: Self-report | .382 | .146 | 25 | 2.14 | .138 | |||
| 2: Self-report and neural activity | .592 | .351 | 24 | 4.32 | .014 | .204 | 7.554 | 0.011 |
Thus, neural activity and self-report predict independent variance in changes in expired CO; neural activity during exposure to ads designed to help smokers quit smoking more than doubles the variability explained in subsequent behavior change (as inferred by CO scores), as compared with the self-report measures used alone, and significantly improves model fit. The combination of all self-report measures collected plus neural activity results in the greatest proportion of variability explained, with the fMRI and self-report contributing equal portions of independent variance in this case.
Discussion
In this investigation, we extended prior results demonstrating the ability of neural responses to persuasive messages to predict real-world outcomes to the context of smoking reduction. We targeted smokers who were already taking action to quit, holding stage of change constant across our sample. We also extended prior results by demonstrating that the variability in behavior change predicted by neuroimaging data is independent of self-efficacy and ability to relate to the messages viewed, in addition to being independent of measures captured in prior studies such as intentions to change.
In this article, we have reported that activity in an identical subregion of MPFC that was associated with behavior change in a prior, independent study of sunscreen behavior change (Falk et al., 2010) was associated with changes in expired CO following exposure to professionally developed quitting ads. This relationship (increased MPFC → greater decrease in expired CO) remained significant after controlling for self-reported intentions and self-efficacy, two common measures used to predict behavior change, as well as participants’ self-reported ability to relate to the ads. Consistent with prior findings (e.g., Falk et al., 2010) that found that neural measures explained an additional 23% in behavior change, neural activity in our ROI explained an additional 20% of the variability in expired CO, above and beyond self-reported intentions, self-efficacy, and ability to relate to the ads, doubling the variability explained in comparison to traditional self-report alone.
In considering the psychological mechanisms that might link the observed neural activity to behavior change, we hypothesize a self-processing mechanism; activity in MPFC is implicated in nearly all studies of self-related processing (Lieberman, 2010). The idea that self-processing may link neural activity to behavior change is also consistent with research suggesting that people who become more absorbed in the narrative of antismoking commercials report increased benefit (Dunlop, Wakefield, & Kashima, 2008), that self-relevant messages are more effective than messages targeting generic individuals (Dietz, Delva, Woolley, & Russello, 2008; Strecher et al., 2008; Strecher, Shiffman, & West, 2005), and that tailored messages activate MPFC more than messages that are not personally tailored (Chua, Liberzon, et al., 2009). These findings are also consistent with theories that highlight selfrelated processes as predictors of behavior change (Ajzen & Fishbein, 1980; Fishbein et al., 2001; Strecher & Rosenstock, 1997).
Given that several prominent theories of behavior change (Ajzen & Fishbein, 1980; Fishbein & Ajzen, 1975; Fishbein et al., 2001; Strecher & Rosenstock, 1997) touch on self-related processing of different varieties, prior work (Falk et al., 2010) speculated that activity in our MPFC ROI might reflect activation of either implicit or explicit connections between message content and the self. In the current investigation, we tested one measure of explicit connection between the self and the message (ability to relate to the message). We found that self-reported ability to relate to messages predicted variability independent of neural activity in MPFC. This explicit measure may be useful in understanding the process of behavior change; however, it does not explain the MPFC–behavior relationship. Our findings do not rule out a different form of self-related processing, and informal debriefing with subjects several weeks postscan suggested that many of the ads that did not seem immediately relevant at the time of the scan emerged as especially relevant and helpful as they engaged in the process of quitting. One interpretation that is consistent with these participant observations is that MPFC activity in this context reflects an implicit connection between the self and the behavior in question (in this case quitting); the particular ventral subregion of MPFC targeted in our investigation has also been implicated in implicit valuation and affective judgments, independent of conscious awareness (Moran, Heatherton, & Kelley, 2009; Rameson, Satpute, & Lieberman, 2010).
It is also possible that neural activity in MPFC may capture a different type of process that is distinct from intention, self-efficacy, and ability to relate to message content, which can be pinpointed in future investigations. Regions of MPFC in the vicinity of the region observed by Falk and colleagues (2010) to predict behavior change have been associated with framing effects (Chua, Liberzon, et al., 2009; Chua, Polk, et al., 2009), implicit preferences (McClure et al., 2004), consideration of personally relevant future goals (D’Argembeau et al., 2010), and value of stimuli in terms of expected outcomes with respect to the current situation (Cunningham, Zelazo, Packer, & Van Bavel, 2007).
Building on evidence that MPFC activity is associated with envisioning personal goals when envisioning future events (D’Argembeau et al., 2010), as well as weighing costs and benefits of stimuli against current experience (Cunningham et al., 2007), it is possible that increased MPFC activity in our ROI supports envisioning oneself carrying out the behavior in question. This interpretation is consistent with classic work suggesting the importance of providing a specific action plan in changing behavior (Dabbs & Leventhal, 1966; Leventhal, Singer, & Jones, 1965; Leventhal, Watts, & Pagano, 1967). This interpretation is also consistent with the observed relationship between activity in other self-related processing regions (e.g., medial precuneus/posteriorcingulate) as well as a region involved in motor planning (supplementary motor area) in whole-brain regressions exploring regions associated with behavior change (Kosten et al., 2006; McClernon, Kozink, Lutz, & Rose, 2009). This coordinated activity might index the extent to which antismoking ads prompt participants to engage in planning of specific personal actions needed to carry out the behavior in question.3 Thus, it is possible that although we have focused initially on MPFC as producing a signal that explains a sizable proportion of variability in behavior change, exploring coordinated activity in MPFC, medial precuneus/posterior-cingulate, and supplementary motor area could explain an even greater proportion of variability in subsequent behavior change; more ventral posterior cingulate activity has been observed in other studies in which neural activity during cognitive tasks was associated with subsequent changes in substance use (Brewer et al., 2008; Paulus et al., 2005), and might be explored as an additional ROI in future studies linking cognitive control to successful quitting. Just as we built on prior work to refine our ability to predict behavior change, regions identified in whole-brain searches in this investigation can serve to develop even more targeted and sophisticated ROIs in future work.
Limitations
Future work may also benefit by addressing limitations of the current study. In particular, given that nicotine alters cerebral blood flow and vascular resistance (Hall, 1972; Miyazaki, 1969; Skinhoj, Olesen, & Paulson, 1973), that the effects may differ across the brain (Jacobsen et al., 2002), and that acute nicotine administration causes its own changes in BOLD signal (Kumari et al., 2003; Stein et al., 1998), the neural activation observed in this study could have been influenced by recency of smoking prior to the scan; future work will benefit by more precisely measuring and controlling time between nicotine administration and task performance. Likewise, it should be noted that the expired CO measure is sensitive to a number of factors other than recent smoking; for example, the measure is sensitive to the recency of smoking (which we attempted to control by allowing all participants to smoke directly before the scan), the brand of CO machine (which was standardized across the study), running and cardiovascular fitness (future investigations might benefit from explicitly recording information pertaining to changes in physical activity habits; however, in the present sample, we are unaware of any major changes in these habits among our participants), and proximity to car emissions (which again could be more precisely measured in future investigations, but is unlikely to have shifted dramatically within our smokers over the span of the study), among other factors. Furthermore, expired CO is sensitive to the proximity to others smoking, which we believe is an advantage to this measure in the current investigation because reducing both one’s own smoking as well as exposure to others who are smoking are important to long-term health. Expired CO presents the advantage of being a biological index of recent smoking, and parallels other investigations using fMRI to examine outcomes in cocaine-dependent patients (Brewer et al., 2008) and to test compliance in other fMRI/smoking investigations (McClernon et al., 2009); however, as in the present study, future investigations will also benefit from observing the similarities and differences between such biological measures and more traditional self-report measures. To the extent that these issues were present in the current data set, each would have impaired our ability to predict outcomes from brain data. However, controlling such factors in the future may allow investigators to predict outcomes more precisely.
Conclusion
The results of this investigation provide convergent evidence that neural activity can predict behavior change, above and beyond self-report. Given that self-report and neural activity explain independent portions of the variability in behavior change, neuroimaging methods can be viewed as a complement to existing self-report methods, indexing processes that may be inaccessible to conscious awareness or otherwise uncaptured through self-report. The specific psychological mechanisms linking neural activity to behavior change will be of interest to future investigations, where tests of affective versus cognitive and implicit versus explicit self-processes, as well as the role of envisioning the future self in the context of goals may yield especially fruitful results. Networks of activity including regions from both the self-related processing network and areas implicated in motor planning may also serve as particularly useful targets for future research. Finally, pinpointing psychological mechanisms that link neural activity to behavior change will help us connect our understanding of the brain to classic work in health psychology. This will also be of use in updating our understanding of behavior change and generating novel self-report measures that could be used in a wider context to explain variability that is currently accounted for using fMRI. The current results bring us one step closer to the ability to use fMRI to select messages that are most likely to change behavior both at the individual and population levels, and further suggest that brain data may provide information that introspection does not.
Acknowledgments
The authors wish to thank: Janna Dickenson and the staff of the UCLA Brainmapping Center for their tremendous assistance; Robert Kaplan and John Pierce for helpful discussions and guidance; For helpful discussion and provision of stimulus materials: Donna Valone, Kristen McCausland, Jeff Costantino and the American Legacy Foundation; Jason Melancon, Shawna Shields and Rosalind Bello and the Louisiana Public Health Institute/Louisiana Campaign for Tobacco-Free Living; Colleen Stevens and the California Department of Public Health; and Greg Connolly at the Harvard School of Public Health.
Footnotes
Attempts were made to standardize the procedural order such that participants smoked as closely as possible to the start of the scan. Some fluctuation in time from the last cigarette to start of the ads task did occur given the variability in setting up participants in the scanner, need to repeat instructions, and time needed for participants to acclimate to using the button box.
As reported elsewhere (Berkman et al., in press), due to nonnormality of the self-report (number of cigarettes) variables, baseline and endpoint number of cigarettes were log-transformed prior to conducting statistical tests.
It is interesting to note that activity in these regions is also associated with substance-cue potentiation in abstinent smokers viewing smoking-related cues (McClernon et al., 2009), as well as relapse in cocaine-dependent patients who viewed cocaine-related cues (Kosten et al., 2006). It is possible that responses to cues prompting quitting and cues prompting activation of substance use activate planning routines that would predict future behavior.
Contributor Information
Emily B. Falk, Department of Communication Studies, Research Center for Group Dynamics/Institute for Social Research, and Department of Psychology, University of Michigan
Elliot T. Berkman, Department of Psychology, University of Oregon
Danielle Whalen, Department of Psychology, University of California, Los Angeles.
Matthew D. Lieberman, Department of Psychology, University of California, Los Angeles
References
- Ajzen I. The theory of planned behavior. Organizational Behavior and Human Decision Processes. 1991;50:179–211. [Google Scholar]
- Ajzen I, Fishbein M. Understanding attitudes and predicting social behavior. Englewood Cliffs, NJ: Prentice Hall; 1980. [Google Scholar]
- Armitage CJ, Conner M. Efficacy of the theory of planned behavior: A meta-analytic review. British Journal of Social Psychology. 2001;40:471–499. doi: 10.1348/014466601164939. [DOI] [PubMed] [Google Scholar]
- Berkman ET, Dickenson J, Falk EB, Lieberman MD. Using SMS text messaging to assess moderators of smoking reduction: Validating a new tool for ecological measurement of health behaviors. (in press) Manuscript submitted for publication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biener L, Abrams DB. The Contemplation Ladder: Validation of a measure of readiness to consider smoking cessation. Health Psychology. 1991;10:360–365. doi: 10.1037//0278-6133.10.5.360. [DOI] [PubMed] [Google Scholar]
- Booth-Kewley S, Larson GE, Miyoshi DK. Social desirability effects on computerized and paper-and-pencil questionnaires. Computers in Human Behavior. 2007;21:463–477. [Google Scholar]
- Brett M, Anton J, Valabregue R, Poline J. Region of interest analysis using an SPM toolbox. Paper presented at the 8th International Conference on Functional Mapping of the Human Brain; Sendai, Japan. 2002. Jun, Available on CD-ROM in NeuroImage. [Google Scholar]
- Brewer JA, Worhunsky PD, Carroll KM, Rounsaville BJ, Potenza MN. Pretreatment brain activation during Stroop task is associated with outcomes in cocaine-dependent patients. Biological Psychiatry. 2008;64:998–1004. doi: 10.1016/j.biopsych.2008.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brody AL, Mandelkern MA, London ED, Childress AR, Lee GS, Bota RG, Jarvik ME. Brain metabolic changes during cigarette craving. Archives of General Psychiatry. 2002;59:1162–1172. doi: 10.1001/archpsyc.59.12.1162. [DOI] [PubMed] [Google Scholar]
- Brody AL, Mandelkern MA, Olmstead RE, Jou J, Tiongson E, Allen V, Cohen MS. Neural substrates of resisting craving during cigarette cue exposure. Biological Psychiatry. 2007;62:642–651. doi: 10.1016/j.biopsych.2006.10.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Centers for Disease Control and Prevention. Annual smokingattributable mortality, years of potential life lost, and productivity losses— United States, 2000–2004. Morbidity and Mortality Weekly Report. 2008;57:1226–1228. [PubMed] [Google Scholar]
- Centers for Disease Control and Prevention. Health effects of cigarette smoking. 2010 Retrieved from http://www.cdc.gov/tobacco/data_statistics/fact_sheets/health_effects/effects_cig_smoking/index.htm#definition.
- Chaiken S, Liberman A, Eagly AH. Heuristic and systematic information processing within and beyond the persuasion context. In: Uleman JS, Bargh JA, editors. Unintended thought. New York: Guilford Press; 1989. pp. 212–252. [Google Scholar]
- Chua H, Liberzon I, Welsh R, Strecher V. Neural correlates of message tailoring and self-relatedness in smoking cessation programming. Biological Psychiatry. 2009;65:165–168. doi: 10.1016/j.biopsych.2008.08.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chua H, Polk T, Welsh R, Liberzon I, Strecher V. Neural responses to elements of a web-based smoking cessation program. Studies in Health Technology and Informatics. 2009;144:174–178. [PMC free article] [PubMed] [Google Scholar]
- Cialdini R, Goldstein N. Social influence: Compliance and conformity. Annual Review of Psychology. 2004;55:591–621. doi: 10.1146/annurev.psych.55.090902.142015. [DOI] [PubMed] [Google Scholar]
- Cunningham W, Zelazo P, Packer DJ, Van Bavel JJ. The iterative reprocessing model: A multilevel framework for attitudes and evaluation. Social Cognition. 2007;25:736–760. [Google Scholar]
- D’Argembeau A, Stawarczyk D, Majerus S, Collette F, Van der Linden M, Feyers D, Salmon E. The neural basis of personal goal processing when envisioning future events. Journal of Cognitive Neuroscience. 2010;22:1701–1713. doi: 10.1162/jocn.2009.21314. [DOI] [PubMed] [Google Scholar]
- Dabbs JM, Jr, Leventhal H. Effects of varying the recommendations in a fear-arousing communication. Journal of Personality and Social Psychology. 1966;4:525–531. doi: 10.1037/h0021190. [DOI] [PubMed] [Google Scholar]
- Dietz NA, Delva J, Woolley ME, Russello L. The reach of a youth-oriented anti-tobacco media campaign on adult smokers. Drug and Alcohol Dependence. 2008;93:180–184. doi: 10.1016/j.drugalcdep.2007.08.019. [DOI] [PubMed] [Google Scholar]
- Dunlop SM, Wakefield M, Kashima Y. The contribution of antismoking advertising to quitting: Intra- and interpersonal processes. Journal of Health Communication. 2008;13:250–266. doi: 10.1080/10810730801985301. [DOI] [PubMed] [Google Scholar]
- Edwards AL. The relationship between the judged desirability of a trait and the probability that the trait will be endorsed. Journal of Applied Psychology. 1953;37:90–93. [Google Scholar]
- Falk EB, Berkman ET, Harrison B, Mann T, Lieberman MD. Predicting persuasion-induced behavior change from the brain. Journal of Neuroscience. 2010;30:8421–8424. doi: 10.1523/JNEUROSCI.0063-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fishbein M, Ajzen I. Belief, attitude, intention, and behavior: An introduction to theory and research. Reading, MA: Addison Wesley; 1975. [Google Scholar]
- Fishbein M, Triandis HC, Kanfer FH, Becker M, Middlestadt SE, Eichler A. Factors influencing behavior and behavior change. In: Baum A, Revenson TA, Singer JE, editors. Handbook of health psychology. Mahwah, NJ: Erlbaum; 2001. pp. 3–16. [Google Scholar]
- Hall GH. Effects of nicotine, carbon monoxide and tobacco smoke on regional blood flow in the cerebral cortex. European Journal of Pharmacology. 1972;19:385–388. doi: 10.1016/0014-2999(72)90106-9. [DOI] [PubMed] [Google Scholar]
- Hammersley R. A digest of memory phenomena for addiction research. Addiction. 1994;89:283–293. doi: 10.1111/j.1360-0443.1994.tb00890.x. [DOI] [PubMed] [Google Scholar]
- Hurlburt RT, Heavey CL. Telling what we know: Describing inner experience. Trends in Cognitive Sciences. 2001;5:400–403. doi: 10.1016/s1364-6613(00)01724-1. [DOI] [PubMed] [Google Scholar]
- Jacobsen LK, Gore JC, Skudlarski P, Lacadie CM, Jatlow P, Krystal JH. Impact of intravenous nicotine on BOLD signal response to photic stimulation. Magnetic Resonance Imaging. 2002;20:141–145. doi: 10.1016/s0730-725x(02)00494-0. [DOI] [PubMed] [Google Scholar]
- Klesges RC, Debon M, Ray JW. Are self-reports of smoking rate biased? Evidence from the Second National Health and Nutrition Examination Survey. Journal of Clinical Epidemiology. 1995;48:1225–1233. doi: 10.1016/0895-4356(95)00020-5. [DOI] [PubMed] [Google Scholar]
- Kosten TR, Scanley BE, Tucker KA, Oliveto A, Prince C, Sinha R, Wexler BE. Cue-induced brain activity changes and relapse in cocaine-dependent patients. Neuropsychopharmacology. 2006;31:644–650. doi: 10.1038/sj.npp.1300851. [DOI] [PubMed] [Google Scholar]
- Kumari V, Gray JA, ffytche DH, Mitterschiffthaler MT, Das M, Zachariah E, Sharma T. Cognitive effects of nicotine in humans: An fMRI study. NeuroImage. 2003;19:1002–1013. doi: 10.1016/s1053-8119(03)00110-1. [DOI] [PubMed] [Google Scholar]
- Leventhal H, Singer R, Jones S. Effects of fear and specificity of recommendation upon attitudes and behavior. Journal of Personality and Social Psychology. 1965;34:20–29. doi: 10.1037/h0022089. [DOI] [PubMed] [Google Scholar]
- Leventhal H, Watts JC, Pagano F. Effects of fear and instructions on how to cope with danger. Journal of Personality and Social Psychology. 1967;6:313–321. doi: 10.1037/h0021222. [DOI] [PubMed] [Google Scholar]
- Lieberman M. Social cognitive neuroscience. In: Fiske S, Gilbert D, Lindzey G, editors. Handbook of social psychology. 5. New York: McGraw-Hill; 2010. pp. 143–193. [Google Scholar]
- McClernon FJ, Kozink RV, Lutz AM, Rose JE. 24-h smoking abstinence potentiates fMRI-BOLD activation to smoking cues in cerebral cortex and dorsal striatum. Psychopharmacology (Berl) 2009;204:25–35. doi: 10.1007/s00213-008-1436-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClure S, Li J, Tomlin D, Cypert K, Montague L, Montague P. Neural correlates of behavioral preference for culturally familiar drinks. Neuron. 2004;44:379–387. doi: 10.1016/j.neuron.2004.09.019. [DOI] [PubMed] [Google Scholar]
- Mendrek A, Monterosso J, Simon SL, Jarvik M, Brody A, Olmstead R, London ED. Working memory in cigarette smokers: Comparison to non-smokers and effects of abstinence. Addictive Behaviors. 2006;31:833–844. doi: 10.1016/j.addbeh.2005.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyazaki M. Circulatory effect of cigarette smoking, with special reference to the effect on cerebral hemodynamics. Japanese Circulation Journal. 1969;33:907–912. doi: 10.1253/jcj.33.907. [DOI] [PubMed] [Google Scholar]
- Moran JM, Heatherton TF, Kelley WM. Modulation of cortical midline structures by implicit and explicit self-relevance evaluation. Society for Neuroscience. 2009;4:197–211. doi: 10.1080/17470910802250519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- National Cancer Institute. NCI Tobacco Control Monograph Series No. 19. Washington, DC: U.S. Department of Health and Human Services; 2008. The role of the media in promoting and reducing tobacco use. [Google Scholar]
- Nederhof AJ. Methods of coping with social desirability bias: A review. European Journal of Social Psychology. 2006;15:263–280. [Google Scholar]
- Nisbett R, Wilson T. Telling more than we can know: Verbal reports on mental processes. Psychological Review. 1977;84:231–259. [Google Scholar]
- Office of the Surgeon General. United States Surgeon General’s Report: The health consequences of smoking. Washington, DC: U.S. Department of Health and Human Services; 2004. [Google Scholar]
- Paulus MP, Tapert SF, Schuckit MA. Neural activation patterns of methamphetamine-dependent subjects during decision making predict relapse. Archives of General Psychiatry. 2005;62:761–768. doi: 10.1001/archpsyc.62.7.761. [DOI] [PubMed] [Google Scholar]
- Petty RE, Cacioppo JT. The elaboration likelihood model of persuasion. In: Berkowitz L, editor. Advances in experimental social psychology. Vol. 19. Orlando, FL: Academic Press; 1986. pp. 123–205. [Google Scholar]
- Pierce JP. Electronic recording, self-report, and bias in measuring cigarette consumption. Health Psychology. 2009;28:527–528. doi: 10.1037/a0016188. [DOI] [PubMed] [Google Scholar]
- Popham W, Potter L, Bal D, Johnson M, Duerr J, Quinn V. Do anti-smoking media campaigns help smokers quit? Public Health Reports. 1993;108:510–513. [PMC free article] [PubMed] [Google Scholar]
- Rameson LT, Satpute AB, Lieberman MD. The neural correlates of implicit and explicit self-relevant processing. NeuroImage. 2010;50:701–708. doi: 10.1016/j.neuroimage.2009.12.098. [DOI] [PubMed] [Google Scholar]
- Rosenstock IM. Why people use health services. Milbank Memorial Fund Quarterly. 1966;44:94–124. [PubMed] [Google Scholar]
- Shiffman S. Commentary on Herd & Borland (2009) and Herd et al. (2009): Illuminating the course and dynamics of smoking cessation. Addiction. 2009;104:2100–2101. doi: 10.1111/j.1360-0443.2009.02799.x. [DOI] [PubMed] [Google Scholar]
- Skinhoj E, Olesen J, Paulson OB. Influence of smoking and nicotine on cerebral blood flow and metabolic rate of oxygen in man. Journal of Applied Physiology. 1973;35:820–822. doi: 10.1152/jappl.1973.35.6.820. [DOI] [PubMed] [Google Scholar]
- Stein EA, Pankiewicz J, Harsch HH, Cho JK, Fuller SA, Hoffmann RG, Bloom AS. Nicotine-induced limbic cortical activation in the human brain: A functional MRI study. The American Journal of Psychiatry. 1998;155:1009–1015. doi: 10.1176/ajp.155.8.1009. [DOI] [PubMed] [Google Scholar]
- Strecher VJ, McClure JB, Alexander GL, Chakraborty B, Nair VN, Konkel JM, Pomerleau OF. Web-based smokingcessation programs: Results of a randomized trial. American Journal of Preventive Medicine. 2008;34:373–381. doi: 10.1016/j.amepre.2007.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strecher VJ, Rosenstock IM. The health belief model. In: Baum A, Newman S, Weinman J, West R, McManus C, editors. Cambridge handbook of psychology, health and medicine. Cambridge, England: Cambridge University Press; 1997. pp. 113–117. [Google Scholar]
- Strecher VJ, Shiffman S, West R. Randomized controlled trial of a web-based computer-tailored smoking cessation program as a supplement to nicotine patch therapy. Addiction. 2005;100:682–688. doi: 10.1111/j.1360-0443.2005.01093.x. [DOI] [PubMed] [Google Scholar]
- Vallone DM, Duke JC, Mowery PD, McCausland KL, Xiao H, Costantino JC, Allen JA. The impact of EX: Results from a pilot smoking-cessation media campaign. American Journal of Preventive Medicine. 2010;38(Suppl. 3):S312–S318. doi: 10.1016/j.amepre.2009.11.021. [DOI] [PubMed] [Google Scholar]
- Webb T, Sheeran P. Does changing behavioral intentions engender behavior change? A meta-analysis of the experimental evidence. Psychological Bulletin. 2006;132:249–268. doi: 10.1037/0033-2909.132.2.249. [DOI] [PubMed] [Google Scholar]
- Weinstein A, Cox WM. Cognitive processing of drug-related stimuli: The role of memory and attention. Journal of Psychopharmacology. 2006;20:850–859. doi: 10.1177/0269881106061116. [DOI] [PubMed] [Google Scholar]
- Wilson T, Nisbett RE. The accuracy of verbal reports about the effects of stimuli on evaluations and behavior. Social Psychology. 1978;41:118–131. [Google Scholar]
- Wilson T, Schooler J. Thinking too much: Introspection can reduce the quality of preferences and decisions. Journal of Personality and Social Psychology. 1991;60:181–192. doi: 10.1037//0022-3514.60.2.181. [DOI] [PubMed] [Google Scholar]
- Xu J, Mendrek A, Cohen MS, Monterosso J, Simon S, Jarvik M, London ED. Effect of cigarette smoking on prefrontal cortical function in nondeprived smokers performing the Stroop task. Neuropsychopharmacology. 2007;32:1421–1428. doi: 10.1038/sj.npp.1301272. [DOI] [PMC free article] [PubMed] [Google Scholar]