

Abstract
Biomass-derived energy offers the potential to increase energy security while mitigating anthropogenic climate change, but a successful path toward increased production requires a thorough accounting of costs and benefits. Until recently, the efficacy of biomass-derived energy has focused primarily on biogeochemical consequences. Here we show that the biogeophysical effects that result from hypothetical conversion of annual to perennial bioenergy crops across the central United States impart a significant local to regional cooling with considerable implications for the reservoir of stored soil water. This cooling effect is related mainly to local increases in transpiration, but also to higher albedo. The reduction in radiative forcing from albedo alone is equivalent to a carbon emissions reduction of  , which is six times larger than the annual biogeochemical effects that arise from offsetting fossil fuel use. Thus, in the near-term, the biogeophysical effects are an important aspect of climate impacts of biofuels, even at the global scale. Locally, the simulated cooling is sufficiently large to partially offset projected warming due to increasing greenhouse gases over the next few decades. These results demonstrate that a thorough evaluation of costs and benefits of bioenergy-related land-use change must include potential impacts on the surface energy and water balance to comprehensively address important concerns for local, regional, and global climate change.
, which is six times larger than the annual biogeochemical effects that arise from offsetting fossil fuel use. Thus, in the near-term, the biogeophysical effects are an important aspect of climate impacts of biofuels, even at the global scale. Locally, the simulated cooling is sufficiently large to partially offset projected warming due to increasing greenhouse gases over the next few decades. These results demonstrate that a thorough evaluation of costs and benefits of bioenergy-related land-use change must include potential impacts on the surface energy and water balance to comprehensively address important concerns for local, regional, and global climate change.
Keywords: regional climate modeling, agriculture, landscape modification, CO2
Securing energy independence and lessening the human fingerprint on climate are two principal motivations behind increased production of bioenergy. Recognition of the full array of costs and benefits of increased production, such as effects on energy and food security, anthropogenic climate change mitigation, and maintenance of biodiversity, will assist in realization of principal objectives (1–8). Prior research gauging the effectiveness of bioenergy has estimated potential impacts based on greenhouse gas (GHG) emission changes through direct or indirect land-use change (LUC) and by means of life cycle analysis (LCA). In addition to impacts on GHGs, LUC also modifies the surface energy and water balance (9), with implications for near-surface temperature and precipitation, and serves as an additional first-order climate forcing on global (10, 11) and regional (9, 12) spatial scales.
One of the main proposed strategies for bioenergy production is widespread planting and harvesting of perennial grasses, such as switchgrass (Panicum virgatum L.) or miscanthus (Miscanthus X giganteus). One LCA study suggested that net GHG savings relative to fossil fuels of greater than 200 g CO2e-C m-2 yr-1 may be expected for biomass (switchgrass) conversion to ethanol (13) (roughly double for hybrid poplar). Potential mitigation, however, is complicated by variability in inventory components and system boundaries (i.e., LCA methodology) that leads to GHG displacement estimates that differ in sign, even for the same species (13–15). Whereas a consensus accounting of carbon savings remains elusive, such analyses should be complemented by consideration of the direct climate effects associated with bioenergy-related LUC, a currently omitted and potentially important environmental consequence. Although such effects may not be global in scale, they could have significant influence on local and regional climates important to millions of people and food production.
Here we use the Weather Research and Forecasting Model (WRF) (16) (see Materials and Methods) to evaluate the climate effects of converting agricultural areas in the central United States to perennial crops. Based on recent side-by-side growing season observations of miscanthus and maize (Zea mays; an annual bioenergy and food crop) (17) we represented the modification of surface vegetation properties by shifting WRF’s default vegetation characteristics (albedo, leaf area index, and vegetation fraction), largely representative of annual cropping systems, over agricultural areas within the central United States (covering 839,000 km2; see Figs. S1 and S2) by 1 mo in each direction (hereafter “Perennials”). That is, vegetation characteristics are advanced by 1 mo, relative to the default, in the spring season (representing earlier green-up) and delayed by 1 mo in the fall season (depicting lagged senescence) (Fig. S2). Vegetation properties were held constant for 2 mo at roughly midway through the growing season (July 31st) to allow for delayed perennial bioenergy crop senescence in the fall. We also performed a pair of additional sensitivity experiments to assess the simulated contribution of albedo (hereafter “Perennials-NoAlb”) and the tendency of perennials to exhibit a deeper rooting depth (hereafter “Perennials-2m”) (18, 19) (Table S1).
Our approach accounts for biogeophysical impacts on climate by considering properties that directly influence the manner in which energy is absorbed at the surface and redistributed to the overlying atmosphere. Modification of the albedo, or the reflectivity of the surface, establishes the surface’s energy availability. In conjunction with the fraction of bare ground covered by vegetation these parameters modulate the partitioning between sensible, latent, and ground heat fluxes. Changes in the green-up and eventual die-down of vegetation, or phenology, will necessarily modify the transport of heat and water vapor to the atmosphere. In addition to the direct impact of the altered landscape, atmospheric feedbacks to modified surface energy inputs may impact cloud formation, with additional consequences for near-surface temperature and precipitation.
The maximum mean growing season near-surface cooling associated with the conversion of annual to perennial crops was nearly 0.9 °C (Fig. 1A). Maintaining albedo at default values decreases the simulated cooling. However, this effect appears to be of secondary importance, on near-surface temperature, compared to impacts of evapotranspiration (ET) (Fig. 1 A and B and Table 1), as more of the incoming energy is used to evaporate water rather than heat the ground in the model experiments with perennial crops. Local growing-season-averaged cooling effects reach 1.5 °C for LUC to a perennial bioenergy crop with a 2 m rooting depth (Fig. 1C and Fig. 2A). Averaged over all land pixels our experiments indicate cooling imparted by perennials between 0.08 °C and 0.16 °C, with mean regional cooling over the growing season exceeding 0.45 °C for all perennial experiments (Table 1).
Fig. 1.

Simulated time mean (APR–OCT) difference in (A) 2-m temperature [°C] (Perennials minus Annuals); (B) as (A) but perennial crop representation does not include albedo modification; (C) as (A) but perennial crop representation includes rooting depth of 2 m.
Table 1.
Mean difference (APR–OCT) response of climate variables between perennials and annuals
| 2 m temp. [°C]: all land | 2 m temp. [°C]: perturbed pixels | 2 m dew-point temp: [°C]: all land | 2 m dew-point temp [°C]: perturbed pixels | ET [mm day-1] | Net surface SW [W m-2] | Net surface LW [W m-2] | |
| Perennials—Annuals | -0.08 | -0.51 | 0.02 | 0.16 | 0.1 | -2.36 | -0.65 |
| Perennials-NoAlb—Annuals | -0.07 | -0.45 | 0.02 | 0.18 | 0.1 | -1.42 | -0.23 |
| Perennials-2m—Annuals | -0.16 | -0.84 | 0.09 | 0.54 | 0.22 | -3.63 | -1.06 |
Where not specified, calculations are for perturbed pixels only. ET, evapotranspiration; SW, shortwave; LW, longwave
Fig. 2.

(A) Simulated evolution of daily mean temperature [°C] difference (Perennials minus Annuals) over grid cells where land surface was perturbed. Blue line: Perennials-2m minus Annuals; red line: Perennials minus Annuals; green line: Perennials-NoAlb minus Annuals. (B) As (A) but for ET [mm day-1]. (C) Simulated evolution of near-surface (top two soil layers: Surface-40 cm) volumetric soil moisture [m3 m-3] averaged over grid cells where land surface was perturbed. Blue line: Perennials-2m; purple line: Annuals; red line: Perennials; green line: Perennials-NoAlb. (D) As (C) but for deep soil (bottom two soil layers: 40–200 cm).
Based on net fossil fuel savings of 140 g CO2e-C m-2 yr-1 (13), we estimate that complete conversion from annual to perennial bioenergy crops over the region would save an additional 13 t C ha-1 yr-1. Accounting for albedo changes resulting from this conversion (i.e., biogeophysical impact) translates to an emissions-equivalent reduction of 78 t C ha-1 (see Materials and Methods for detailed calculation). Conversion from annual to perennial bioenergy crops has a significantly larger impact on global radiative forcing (RF) when including biogeophysical and biogeochemical impacts, together. We estimate that it would take 7 yr for the biogeochemical impacts to surpass biogeophysical effects, for the area of land undergoing conversion in our experiments. Understanding the full spectrum of effects associated with biomass energy expansion will therefore require more than the biogeochemical impact considered to date (20), and must include a careful assessment of potential consequences for local and regional climate (21) and hydrology, as presented here.
Simulated near-surface thermal effects are greatest during spring and fall, with impacts lessened during the summer season (Fig. 2A), when differences in vegetation characteristics between cropping systems approach zero. Given the lack of biogeophysical observational data associated with bioenergy crops, our approach most probably omits additional features associated with summer season variability (e.g., albedo, canopy resistance) whose influence may be substantial (21). Differences in vegetation coverage between annual and perennial bioenergy crops at the start and end of the growing season account for the simulated enhancement in perennial crop ET. Mean ET for perennial crops exceeds that of annuals by 0.1 mm day-1, with shoulder-season effects of considerably greater magnitude (Fig. 2B).
The impact of a deeper rooting depth more than doubles simulated ET relative to the effects of using a 1 m rooting depth. The addition of transpired water moistens the near-surface air and serves as the chief cooling mechanism in our simulations. This transport of water has implications for the reservoir of stored soil water, necessarily decreasing shallow and deep water availability with time (Fig. 2 C and D). While deep water availability displays a progressive decrease with time for Perennials-2m (by the end of the simulation, this difference, relative to Annuals, exceeds 5%) the shallow (i.e., within the top 40 cm of the soil surface) reservoir of soil water increases. The substantial increase in ET (0.22 mm day-1 averaged throughout the growing season; Fig. 2B and Table 1) is sufficient to enhance local recycling of rainfall (Fig. S3), further moistening the near-surface soil. In this sense, our experiments suggest a positive feedback, whereby direct initial cooling resulting from the enhancement of ET (also supported by the increase in 2 m dew-point temperature presented in Table 1) leads to an increase in the vertically integrated cloud and water vapor mixing ratio and rainfall, subsequently reducing net surface shortwave radiation (Table 1), cooling the surface further. The vertical extent of simulated surface cooling extends above the planetary boundary layer and outweighs the impact of enhanced longwave emission due to greater water vapor and cloud mixing ratio, resulting in a net decrease in longwave radiation at the surface for the perennial bioenergy crops (Table 1).
To test the possibility that the ET effect of the perennials is transient, we conducted a three-yr offline simulation using representation of Perennials-2m and Annual crops, with the High-Resolution Land Data Assimilation System (Version 3.1), which runs the Noah land surface model (LSM) in uncoupled mode. The sustained ET difference for each growing season indicates that our simulated ET effect is not transient (Fig. S4).
Whereas the impact of perennials may offset a significant fraction of future greenhouse warming at local scales, it remains small when compared to projected warming from global GHG emissions at large scales (Fig. 3). Locally, and through 2040, the modeled conversion of annual to perennial bioenergy crops balances GHG warming (see Materials and Methods) though this effect occurs largely during spring and fall with reduced impacts during midsummer. Through this time frame the two forcing mechanisms nearly balance each other at about 4 million ha, with GHG-induced warming playing a relatively greater role at larger spatial scales. Through 2100, local LUC effects are greatly diminished, compensating for about one-fourth of projected warming (4 °C) (Fig. 3). Consideration of a lower (i.e., one more consistent with widespread use of biomass-derived energy) emissions trajectory reveals LUC effects that offset nearly one half of projected warming (B1 scenario) at local scales whereas near-surface cooling impacts play a reduced role for higher emissions trajectories (A2 scenario) (Fig. S5).
Fig. 3.

Comparison of near-surface temperature change (April through October) associated with simulated conversion from annual to perennial bioenergy crops (i.e., biogeophysical effect) against projected WCRP CMIP3 warming (i.e., global GHG emissions), with increasing spatial scale (centered on lat: 40.0 °N, longitude: 87.75 °W).
Our experiments suggest that phenological contrasts associated with conversion of annual to perennial bioenergy crops in the central United States may impart significant local and regional influence and through nearly the midcentury are of similar order of magnitude as projected impacts due to rising GHGs. In addition to direct cooling effects, feedbacks related to deep-soil moisture depletion illustrate additional unintended consequences requiring added attention in light of increasing reliance on biomass-derived energy (22–24). By the end of the twenty-first century local cooling effects associated with LUC are much smaller than large-scale warming associated with increased GHGs. Additional biophysical differences between cropping systems (e.g., canopy resistance) during green-up and senescence may lead to further impacts, highlighting the necessity of improving location- and vegetation-specific representation of bioenergy cropping systems. Further work assessing the long-term (i.e., decadal-scale or longer) evolving nature of soil water depletion and associated equilibrium ET resulting from soil moisture/temperature and atmospheric changes are necessary. To more comprehensively gauge the full spectrum of anthropogenic impacts, such forcing that directly influences the scale of human interaction is a necessary component to bridge the large-scale biogeochemical bioenergy impacts studied to date with the local-to-regional-to-global scale biogeophysical effects associated with agricultural practices.
Materials and Methods
WRF Modeling System.
WRF (version 3.1) is a state-of-the-art, fully compressible, nonhydrostatic mesoscale code that has been widely used in a number of applications ranging from urban canopy level modeling, to air quality research, and has been extended to a variety of longer climate scale applications. The vertical coordinate is terrain-following and the horizontal coordinate is staggered on an Arakawa C-grid. The modeling system has a detailed level of complexity, includes multiple parameterization options for convection, radiation, boundary layer, and cloud microphysics, including multiple land-surface model options. We use the 4-layer Noah land surface scheme (25), used operationally at the National Center for Environmental Prediction, to update soil temperature and moisture following model initialization. The Noah land surface model has been widely used in the regional climate modeling community. Important instances include its utility in the development of the 25-yr North American Regional Reanalysis (NARR) atmospheric and land surface hydrology dataset (26), and operation as part of the North American Regional Climate Change Assessment Program (http://narccap.ucar.edu/data/rcm-characteristics.html), which utilizes WRF and its predecessor (MM5) as two of the six atmospheric models used. Of the six atmospheric models, three made use of the Noah LSM to treat interactions between the land surface and overlying atmosphere. For subgrid-scale convective processes, we use a recently modified version of the Kain–Fritsch convective parameterization scheme (27).
We initialize and force all experiments with data extracted from the NARR dataset (26). Initial soil moisture and temperature were also retrieved from NARR, but following the initial model timestep the Noah LSM was used to update these fields and calculate the necessary fluxes required by the Yonsei University Planetary Boundary Layer scheme (28).
Our representation of albedo is based on a monthly, five-yr, 0.144° climatological dataset assumed to be valid at the 15th of each month, with global coverage (16, 29). This albedo dataset implicitly includes variable vegetation density effects, thereby accounting for both soil and vegetated contributions. Total ET, however, is dependant on the vegetation fraction, accounting for both direct (i.e., contribution from bare soil) evaporation as well as contribution from intercepted canopy water and transpired water from canopy and roots. Our representation of vegetation fraction is based on a monthly, five-yr, 0.144° climatological dataset assumed to be valid at the 15th of each month, with global coverage (30).
A detailed inventory of setup options is presented in Table S2.
WRF Simulations.
We conducted four 8 mo simulations initialized on March 1, 1995, and completed on October 31, 1995, characterizing a full growing season (Tables S1 and S2). Our simulation domain consisted of the entire United States, northern Mexico, and southern Canada, and portions of the Atlantic and Pacific oceans (Fig. S1). To represent earlier green-up of perennial bioenergy crops we used the default May 1 biogeophysical characteristics (i.e., those pertaining to “Annuals”) for April 1 Perennials. As with the default vegetation characteristics, which are updated daily, April 2 Perennials biogeophysical characteristics are updated accordingly based upon the May 2 Annuals representation. Updating of vegetation properties for Perennials is maintained such that it lags that of Annuals by 1 mo, until July 1 (corresponding to July 31 for Annuals). Shifting this date back (i.e., corresponding to July 24 for Annuals) and forth (i.e., corresponding to August 7 for Annuals) by one week did not alter the conclusions of our sensitivity experiments. Land surface Perennials representation are maintained at constant values for the ensuing 2 mo with updating resumed once more on September 1 (corresponding to August 1 for Annuals; Perennial(s) senescence is now lagging). Updating of vegetation properties for Perennials is continued, daily (based on the Annuals representation), until October 31 (corresponding to October 1 for Annuals). Monthly mean differences at the start and end of the growing season illustrate phenological contrasts between Annuals and Perennials (Fig. S2).
For most areas of the world, including our study region, albedo is higher for a vegetation canopy compared to soil. For example, it has been shown, using in situ observations, that a maize field experiences an albedo increase relative to the background soil albedo during the growing season (31). Overall, the albedo contrast between biofuel crops and current land cover, which drives the main result related to global climate forcing, is a robust feature that is likely not specific to our particular model or implementation.
The initial month was used as spin-up (for all experiments) with the remaining months of the default simulation (Annuals) evaluated against suitable observations of temperature and precipitation (Fig. S6; see below).
To evaluate the mechanisms behind climate responses, a third scenario that was identical to Perennials but did not include the albedo modification (Perennials-NoAlb) was considered (Table S1). Evaluation of potential consequences of deep root systems associated with perennial crops served as the source of one additional experiment that included a 2-m root depth (Perennials-2m; all other experiments utilized a rooting depth of 1 m).
Evaluation of WRF Simulations.
We evaluate Annuals (which makes use of the 24-class US Geological Survey land use and land cover dataset as the default WRF landscape representation) against suitable observations of gridded temperature and precipitation (Fig. S6). We use the University of Delaware Global Air Temperature data, courtesy of the Earth Systems Research Laboratory (http://www.cdc.noaa.gov) for model comparison to temperature and the Climate Prediction Center US Unified Precipitation data provided by the National Oceanic and Atmospheric Administration/Office of Oceanic and Atmospheric Research/Earth System Research Laboratory (http://www.cdc.noaa.gov) for model comparison to precipitation. We emphasize that the purpose of these sensitivity experiments was not to replicate the climate of the growing season precisely, and consequently no tuning of the model was carried out to improve correspondence to observations. Our primary goal of understanding and quantifying the sensitivity to prescribed biogeophysical landscape change associated with bioenergy crops was enhanced due to usage of spectral nudging (32, 33), a technique recently implemented in the version of WRF utilized for this study. Usage of this method constrains the model solution of the longest atmospheric wavelengths (above the PBL only) to that of the driving boundary fields and leads to more reliable sensitivity results than would otherwise be expected (32, 33).
Differences between modeled and observed temperature for the growing season are least over the eastern one-half of the United States (including the Corn Belt states), southwestern United States, and western United States, with temperature differences generally within 1 °C. WRF does have a positive warm bias over the central and southern plains ranging between 1–3 °C (this warm bias is greatest over parts of Oklahoma and Texas and reaches to nearly 5 °C). Overall, WRF does an excellent job in simulating the broad, continental temperature distribution.
WRF-simulated precipitation over the course of the growing season shows good general agreement, with greater precipitation in the east and less in the west and southwest. WRF properly simulates increased rainfall totals along the Gulf Coast states but misses the observed peak precipitation along Oklahoma, Kansas, Missouri, which instead falls further downstream (along the Corn Belt states). Nevertheless, the broad distribution of precipitation, rather than point-specific totals, is reasonably simulated, and provides confidence in the model’s ability to ably reproduce the United States climate during this time period.
Finally, it is important to highlight that we selected 1995 as a simulation year because external large-scale dynamical forcing agents (e.g., El Nino) were not a primary driving factor in the season’s climatic evolution. Our intent was to quantify bioenergy-related LUC effects separate from other forcing factors.
Carbon Savings Calculations.
Our imposed conversion of annuals to perennials over the central United States encompassed a planted area of 83.9 million ha [this is roughly equivalent to 2016 agricultural area projections within the “Corn Belt,” “Lake States,” and “Northern Plains” states (as defined by the US Department of Agriculture)]. Using a carbon saving estimate (near-term carbon savings of switchgrass [approximately  ] minus carbon savings of corn-soy [approximately 70 g CO2e-C m-2 yr-1], relative to fossil fuels) of 140 g CO2e-C m-2 yr-1 (13), yields a savings of:
] minus carbon savings of corn-soy [approximately 70 g CO2e-C m-2 yr-1], relative to fossil fuels) of 140 g CO2e-C m-2 yr-1 (13), yields a savings of:
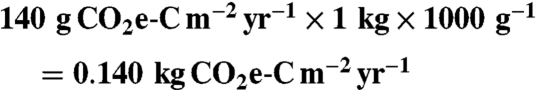 |
 |
 |
Conversion of annual to perennial bioenergy crops over this area would yield a carbon savings of 0.06 ppmv yr-1.
We calculated the contribution of our imposed land-use change to global RF by dividing the local (i.e., over perturbed pixels only) top of the atmosphere (TOA) net shortwave (SW) by the earth’s surface area (34, 35):
 |
Our TOA SW RF value of -3.21 W m-2 represents an entire growing season time average (APR–OCT) difference between Perennials-2m and Annuals. We assume that for the remaining months (i.e., NOV–MAR) snow entirely covers the landscape and reduces RF to zero. This conservative approach yields a global RF of approximately -0.0053 W m-2.
Using a radiative forcing efficiency (α) of 5.35 W m-2 (36) and the previous estimate of CO2 radiative forcing: RF = approximately - 0.0053 W m-2, we obtain:
| [1] |
where C0 is the total (i.e., reference value) CO2 concentration as of 2005 (379 parts per million by volume; 37) and ΔC is the globally averaged atmospheric CO2 concentration change required to give an equivalent global RF to that obtained from our imposed conversion to perennial bioenergy crops. Solving [1] for ΔC yields:
Given our assumption of zero RF for the remaining months:
We estimate that it would take about 6 years for the RF associated with the carbon savings effect (approximately -0.06 ppmv yr-1) to exceed that associated with biogeophysical impacts resulting from a transition to perennial bioenergy crops (approximately -0.37 ppmv). Our estimate of carbon saved is based on the highest GHG displacement values reported in the literature (for switchgrass) and a more conservative value would yield less savings (14) and a longer time period until biogeochemical impacts exceed biogeophysical effects on global RF.
The terrestrial carbon stock change (34) owing to atmospheric CO2 concentration change (i.e., ΔC) may be obtained from:
Where ΔCT is the terrestrial carbon stock change, Mc and Ma are the molecular weights of carbon and dry air, ma is the mass of the atmosphere, the factor of 2 accounts for an airborne emissions fraction of 0.5 (38), and ΔC and C0 are as before.
We calculate ΔCT and estimate the emissions-equivalent of shortwave forcing (34) for both the carbon savings impacts (relative to fossil fuels) and the biogeophysical impacts resulting from conversion to perennial bioenergy crops.
 |
 |
Two of the experiments (Perennials-2m and Annuals) were repeated, identical to previously discussed simulations except for the use of an alternate SW radiation scheme [Rapid Radiation Transfer Model for Global applications(RRTMG)-SW scheme; 16, 39]. Usage of this scheme permitted TOA SW RF as an output parameter (the initial scheme did not) and was used in the carbon savings calculations. Differences between Perennials-2m and Annuals utilizing this radiation scheme were similar to those outlined previously (Fig. S7). For example, near-surface growing season temperature differences (Perennials-2m minus Annuals) over perturbed pixels using the Dudhia SW scheme (Table 1) of -0.84 °C were comparable to those using the RRTMG-SW scheme (difference of -0.72 °C). The similarity in differences despite variability in choice of radiation scheme on climate variables (e.g., soil moisture, near-surface temperature), and associated physical mechanisms, increases confidence in the robustness of our conclusions.
Simulated Rainfall Recycling.
Differences (Perennials-2m minus Annuals) in monthly precipitation are illustrative of enhanced local recycling of rainfall (Fig. S3). Whereas simulated summer (or near-summer) precipitation should be expected to display a greater sensitivity to the choice of convective scheme, the tail end of the growing season (i.e., October) should not, as total precipitation is less reliant on subgrid convective processes and less sensitive to the choice of scheme, as propagating storm systems are innately larger scale.
The majority of the region where we alter land surface properties is rainfed. We do not artificially add irrigation for some experiments while removing it from others and therefore none of our conclusions are based on projected changes in irrigation.
Biogeochemical and Biogeophysical Forcing Comparison.
To project the degree of warming due to increased GHGs that may be offset by the conversion of annual to perennial bioenergy crops we obtained Lawrence Livermore National Laboratory (LLNL)-Reclamation-Santa Clara University downscaled climate projections data derived from the World Climate Research Programme’s (WCRP) Coupled Model Intercomparison Project phase 3 (CMIP3) multimodel dataset, stored and served at the LLNL Green Data Oasis. Multimodel projections corresponding to high (A2), medium (A1b), and low (B1) emission trajectories from 36, 39, and 37 General Circulation Models (for A2, A1b, and B1 scenarios, respectively) of mean April through October temperature change for 2020–2039, 2040–2059, 2060–2079, and 2080–2099 were obtained, and the degree of warming relative to 1980–2000 was calculated for each 2-yr subset. Special Report on Emission Scenarios WCRP CMIP3 data and WRF output data were regridded to identical resolutions to enable direct comparison of forcing agents for the simulation period (April through October). We compared LUC cooling to projected CMIP3 warming across increasing spatial scales starting with the locally greatest effect (centered on lat: 40.0 °N, longitude: 87.75 °W) and proceeded to increasingly larger scales.
Supplementary Material
Acknowledgments.
We thank two anonymous reviewers for their thoughtful critique of the manuscript. This work was made possible through support from the Stanford University Global Climate and Energy Project and National Science Foundation Grant 0934592 at Arizona State University.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1008779108/-/DCSupplemental.
References
- 1.Sustainable Bioenergy: A Framework for Decision Makers. Geneva: United Nations; 2007. Available at http://esa.un.org/un-energy/pdf/susdev.Biofuels.FAO.pdf. [Google Scholar]
- 2.Fargione J, Jason H, Tilman D, Polasky S, Hawthorne P. Land clearing and the biofuel carbon debt. Science. 2008;319:1235–1238. doi: 10.1126/science.1152747. [DOI] [PubMed] [Google Scholar]
- 3.Martindale W, Trewavas A. Fuelling the 9 billion. Nature Biotechnol. 2008;26:1068–1070. doi: 10.1038/nbt1008-1068. [DOI] [PubMed] [Google Scholar]
- 4.Searchinger T, et al. Use of US croplands for biofuels increases greenhouse gases through emissions from land-use change. Science. 2008;319:1238–1240. doi: 10.1126/science.1151861. [DOI] [PubMed] [Google Scholar]
- 5.Hill J, Nelson E, Tilman D, Polasky S, Douglas T. Environmental, economic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc Natl Acad Sci USA. 2006;103:11206–11210. doi: 10.1073/pnas.0604600103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hill J, et al. Climate change and health costs of air emissions from biofuels and gasoline. Proc Natl Acad Sci USA. 2009;106:2077–2082. doi: 10.1073/pnas.0812835106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tilman D, Hill J, Lehman C. Carbon-negative biofuels from low-input high-diversity grassland biomass. Science. 2006;314:1598–1600. doi: 10.1126/science.1133306. [DOI] [PubMed] [Google Scholar]
- 8.Tilman D, et al. Beneficial biofuels—The food, energy, and environment trilemma. Science. 2009;325:270–271. doi: 10.1126/science.1177970. [DOI] [PubMed] [Google Scholar]
- 9.Pielke RA, Sr., et al. Land use and climate change. Science. 2005;310:1625–1626. doi: 10.1126/science.1120529. [DOI] [PubMed] [Google Scholar]
- 10.Foley JA, et al. Global consequences of land-use. Science. 2005;309:570–574. doi: 10.1126/science.1111772. [DOI] [PubMed] [Google Scholar]
- 11.Feddema JJ, et al. The importance of land cover change in simulating future climates. Science. 2005;310:1674–1678. doi: 10.1126/science.1118160. [DOI] [PubMed] [Google Scholar]
- 12.Pielke RA, Sr., et al. An overview of regional land-use and land-cover impacts on rainfall. Tellus B. 2007;59:587–601. [Google Scholar]
- 13.Adler PR, Del Grosso SJ, Parton WJ. Life-cycle assessment of net greenhouse-gas flux for bioenergy cropping systems. Ecol Appl. 2007;17:675–691. doi: 10.1890/05-2018. [DOI] [PubMed] [Google Scholar]
- 14.Davis SC, Anderson-Teixera KJ, DeLucia EH. Life-cycle analysis and the ecology of biofuels. Trends Plant Sci. 2009;14:140–146. doi: 10.1016/j.tplants.2008.12.006. [DOI] [PubMed] [Google Scholar]
- 15.Delucchi MA. Institute of Transportation Studies. Lifecycle Analyses of Biofuels. David, CA: University of California; 2006. Available at http://www.its.ucdavis.edu/publications/2006/UCD-ITS-RR-06-08.pdf. [Google Scholar]
- 16.Skamarock WC, et al. A Description of the Advanced Research WRF Version 3. Boulder, CO: National Center for Atmospheric Research; 2008. [Google Scholar]
- 17.Dohleman FG, Long SP. More productive than maize in the Midwest: How does Miscanthus do it? Plant Physiol. 2009;150:2104–2115. doi: 10.1104/pp.109.139162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Neukirchen D, et al. Spatial and temporal distribution of the root system and root nutrient content of an established Miscanthus crop. Eur J Agron. 1999;11:301–309. [Google Scholar]
- 19.Hall R. Grasses for Energy Production: Hydrological Guidelines. Centre for Ecology and Hydrology; 2003. Available at http://www.bis.gov.uk/files/file14946.pdf. [Google Scholar]
- 20.Field CB, Campbell JE, Lobell DB. Biomass energy: The scale of the potential resource. Trends Ecol Evol. 2007;23:65–72. doi: 10.1016/j.tree.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 21.Georgescu M, Lobell DB, Field CB. Potential impact of US biofuels on regional climate. Geophys Res Lett. 2009;36:L21806. [Google Scholar]
- 22.Delucchi MA. Impact of biofuels on climate change, water use, and land use. Ann NY Acad Sci. 2010;1195:28–45. doi: 10.1111/j.1749-6632.2010.05457.x. [DOI] [PubMed] [Google Scholar]
- 23.Fingerman KR, et al. Accounting for the water impacts of ethanol production. Environ Res Lett. 2010;5:014020. [Google Scholar]
- 24.Vanloocke A, Bernacchi CJ, Twine TE. The impacts of Miscanthus × giganteus production onthe Midwest US hydrologic cycle. Glob Change Biol Bioenergy. 2010;2:180–191. [Google Scholar]
- 25.Chen F, Dudhia J. Coupling an advanced land-surface/hydrology model with the Penn State/NCAR MM5 modeling system. Part I: Model description and implementation. Mon Weather Rev. 2001;129:569–585. [Google Scholar]
- 26.Mesinger F, et al. North American regional reanalysis. B Am Meteorol Soc. 2006;87:343–360. [Google Scholar]
- 27.Kain JS. The Kain–Fritsch convective parameterization: An update. J Appl Meteorol. 2004;43:170–181. [Google Scholar]
- 28.Hong S-Y, Noh Y, Dudhia J. A new vertical diffusion package with an explicit treatment of entrainment processes. Mon Weather Rev. 2006;134:2318–2341. [Google Scholar]
- 29.Csiszar I, Gutman G. Mapping global land surface albedo from NOAA AVHRR. J Geophys Res. 1999;104:6215–6228. [Google Scholar]
- 30.Gutman G, Ignatov A. The derivation of green vegetation fraction from NOAA/AVHRR data for use in numerical weather prediction models. Int J Remote Sens. 1998;19:1533–1543. [Google Scholar]
- 31.Song J. Phenological influences on the albedo of prairie grassland and crop fields. Int J Biometeorol. 1999;42:153–157. [Google Scholar]
- 32.Miguez-Macho G, Stenchikov GL, Robock A. Spectral nudging to eliminate the effects of domain position and geometry in regional climate model simulations. J Geophys Res. 2004;109:D13104. [Google Scholar]
- 33.Miguez-Macho G, Stenchikov GL, Robock A. Regional climate simulations over North America: Interaction of local processes with improved large-scale flow. J Climate. 2005;18:1227–1246. [Google Scholar]
- 34.Betts RA. Offset of potential carbon sink from boreal forestation by decreases in surface albedo. Nature. 2000;408:187–190. doi: 10.1038/35041545. [DOI] [PubMed] [Google Scholar]
- 35.Nemet GF. Net radiative forcing of widespread deployment of photovoltaics. Environ Sci Technol. 2009;43:2173–2178. doi: 10.1021/es801747c. [DOI] [PubMed] [Google Scholar]
- 36.Myhre G, Highwood EJ, Shine KP, Stordal F. New estimates of radiative forcing due to well mixed greenhouse gases. Geophys Res Lett. 1998;25:2715–2718. [Google Scholar]
- 37.Forster P, et al. In: Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Solomon S, et al., editors. Cambridge, UK: Cambridge University Press; 2007. [Google Scholar]
- 38.Canadell JG, et al. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc Natl Acad Sci USA. 2007;104:18866–18870. doi: 10.1073/pnas.0702737104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Iacono MJ, et al. Radiative forcing by long-lived greenhouse gases: Calculations with the AER radiative transfer models. J Geophys Res. 2008;113:D13103. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.