

Purpose of work
To show biological activity of carboxymethylated α-(1 → 3)-d-glucans isolated from the selected macromycetes fungi on human tumor and normal cells.
Water-insoluble, alkali-soluble polysaccharides (WIP) were isolated from fruiting bodies of four macromycetes fungi: Lentinus edodes, Pleurotus ostreatus, Piptoporus betulinus and Laetiporus sulphureus. The structure of the polysaccharides was determined using composition analysis, methylation analysis, fourier transform infrared spectroscopy, and nuclear magnetic resonance spectroscopy. The chemical and spectroscopic investigations indicated that the polysaccharides were an α-(1 → 3)-d-glucans. A biological activity analysis of the carboxymethylated (CM) α-(1 → 3)-d-glucans was based on an assessment of their cytotoxic, mitochondrial metabolism-modulating, and free radical scavenging effects. The cytotoxic activity of the CM-glucans was concentration- and cell-type-dependent. The tested CM-glucans, generally, did not have a free radical scavenging effect. The CM-α-(1 → 3)-d-glucans isolated from the selected macromycetes fungi are biologically active and may therefore be used as diet or therapy supplements.
Keywords: Carboxymethylation, Cytotoxicity, α-(1 → 3)-d-Glucan, Human cell cultures, Macromycetes fungi
Introduction
Currently, there is tendency to introduce natural substances to human therapies. One of the important natural resources of biologically active substances are the macromycetes fungi, amongst which almost 700 species possess significant pharmacological activity. The diverse biological activities of macromycetes fungi include antibacterial, antiviral, antitumor, immunosuppressive, antiallergic, and antisclerotic effects. Moreover, substances derived from these fungi may act as supplements for lowering blood pressure, antioxidants, and hepatoprotective and antiinflammatory factors (Lindequist et al. 2005). Among these compounds, polysaccharides, which belong mainly to β- and α-d-glucans, play a special role in immunomodulation and antitumor effects. Nowadays, the best analyzed fungal polysaccharides are β-d-glucans (e.g., lentinan, schizophyllan and grifolan) and their peptide/protein derivatives (Bohn and BeMiller 1995; Ng and Yap 2002; Sarkar et al. 1993). The properties of these agents include stimulation of hematopoietic stem cells, activation of the alternative complement pathway, and activation of immune cells such as lymphocytes, macrophages, DC, NK cells, Th cells, Tc cells, and B cells (Moradali et al. 2007). Compared with the intensive investigations of β-d-glucans, the biological activity of α-(1 → 3)-d-glucan, which is especially apparent after chemical and structural modification (carboxymethylation, sulfation, aminopropylation, or hydroxyethylation) (Bao et al. 2001), has been reported only rarely in the literature. There are only a few studies indicating that CM-α-(1 → 3)-d-glucan preparations from various macromycetes fungi may exhibit antitumor or immunomodulatory effects, have been explored (Kiho et al. 1994).
In the present study, we analyzed the structure of water-insoluble, alkali-soluble polysaccharides (WIP) isolated from fruiting bodies of Lentinus edodes, Pleurotus ostreatus, Piptoporus betulinus, and Laetiporus sulphureus and their biological activity, i.e. cytotoxicity, mitochondrial metabolism-modulating effects, and free radical scavenging activity, of their CM products against two tumor cell lines, human cervical carcinoma (HeLa) and human acute T cell leukemia (Jurkat, Clone E6-1), and normal human skin fibroblasts (HSF).
Materials and methods
Fungi
The fruiting bodies of Laetiporus sulphureus (Bull.: Fr.) Murrill and Piptoporus betulinus (Bull.: Fr.), representing a combination of young and aged basidiocarps, were collected from deciduous trees grown in Lublin and environs, Poland. The fungi were identified by Dr. Z. Flisińska, Department of Botany and Mycology, Maria Curie-Skłodowska University, Lublin, Poland. Voucher specimens are deposited in the Department of Industrial Microbiology, Maria Curie-Skłodowska University, Poland. Cultivated mushrooms [Lentinus edodes (Berk.) Pegler and Pleurotus ostreatus (Jacq.: Fr.) P. Kumm.] were purchased in a supermarket.
Isolation of the alkali-soluble polysaccharides
Fresh fruiting bodies of the selected macromycetes fungi were lyophilized and milled. The dried material was used for the isolation of the alkali-soluble α-glucans according to a method described by Kiho et al. (1994). The dried material (100 g) was successively extracted with methanol, NaCl (9 g l−1), hot water, Na2CO3 (50 g l−1), and 1 M NaOH containing 0.2 g NaBH4 l−1 for 24 h at room temperature. The final extracts were neutralized with 1 M HCl constant mixing. The water-insoluble fractions were washed with water, collected by centrifugation, and lyophilized. The yields of WIP isolated from L. edodes, P. ostreatus, P. betulinus and L. sulphureus were 9, 6.1, 24, and 57% of dry fungal mass, respectively.
Carboxymethylation
Because the isolated α-(1 → 3)-d-glucans were insoluble in water, they were converted into water soluble fractions by carboxymethylation according to the procedure of Sasaki et al. (1979). The degree of substitution (DS) of the carboxymethyl groups was determined after total hydrolysis by 1H NMR spectroscopy (Ho and Klosiewicz 1980). Spectra were recorded with an Avance (300 MHz) spectrometer (Bruker BioSpin) at 40°C.
Sugar analysis
For sugar analysis, the polysaccharides were hydrolyzed with 2 M trifluoroacetic acid (TFA) (100°C, 4 h). The sugars were converted into alditol acetates (Russa et al. 1995). Methylation was done according to the method of Hakomori (1964), and the methylated polysaccharides were purified on a Sep-Pak C18 cartridge (York et al. 1986). The resulting material was hydrolyzed in 2 M TFA (100°C, 4 h) and reduced with NaBD4. The partially methylated alditols were converted into acetate derivatives. 1H NMR spectra of the polysaccharide preparations were recorded as before at 60°C. The 1H chemical shift was obtained using acetone (δ H – 2.225 ppm) as the internal standard. An infrared absorption spectrum (FT-IR) between 400 and 4,000 cm−1 was recorded with the specimen prepared as a KBr disk.
Preparation of CM-glucans for experiments
CM-α-(1 → 3)-d-Glucans (100 mg) were dissolved in 10 ml 0.85% (w/v) NaCl, divided into portions and frozen. For experiments, glucans were prepared at 25–250 μg ml−1.
Cell culture
Three human cell lines were used. The HSF cells were prepared by the liquid overlay method in the Department of Virology and Immunology, University of Maria Curie-Skłodowska in Lublin, Poland. Skin fragments had been freshly excised from healthy donors. Outgrowths of skin fibroblasts were separated, cultured, and used for the experiments. Human Negroid cervix epitheloid carcinoma (HeLa) cells (ECACC No. 93021013) were cultured as monolayers in 25 cm2 culture flasks. Jurkat, Clone E6-1 acute T cell leukemia (ATCC No. TIB-152) cells were cultured in suspension. All cell lines were maintained in RPMI 1640 medium supplemented with 10% FBS (fetal bovine serum) (v/v) and antibiotics (100 U penicillin ml−1, 100 μg streptomycin ml−1) at 37°C in a humidified atmosphere with 5% CO2.
Incubation of cells with CM-α-(1 → 3)-d-glucans
The total number of cells was estimated by counting in haemocytometer. A 100 μl of cell suspension (2 × 105 cells ml−1) was added to appropriate wells of 96-well flat-bottomed microtitre plates (MTT and NR methods). After 24 h the medium was discarded and a fresh one containing 2% (v/v) FBS and appropriate concentrations of CM-α-(1 → 3)-d-glucans was added. In case of Jurkat cells, medium was changed after centrifugation of the plates (1,000 rpm for 5 min). As control, cells cultured in 100 μl medium supplemented with 2% (v/v) FBS was used. The total cell number was equivalent to these in the sample wells. An additional control, consisting of culture medium with 2% (v/v) FBS and appropriate CM-α-(1 → 3)-d-glucan concentrations, was used in the MTT test to evaluate MTT reduction by CM-glucans themselves. The incubation was performed for further 24 h and cytotoxicity of CM-α-(1 → 3)-d-glucans was estimated with spectrophotometric methods (MTT and NR assays).
MTT, NR-uptake, and DPPH• assays
MTT, Neutral Red (NR) uptake and DPPH• free radical scavenging assays have been performed as described previously (Paduch et al. 2008). All these methods are spectrophotometric. The MTT test is based on the conversion by viable cells of the yellow tetrazolium salt to purple crystals of formazan. The reaction is catalyzed by mitochondrial succinyl dehydrogenase. The NR cytotoxicity assay is based on the uptake and lysosomal accumulation of the supravital dye, neutral red.
Results and discussion
Polysaccharide analysis
Monosaccharide analysis (Table 1) showed that all the tested WIP consisted principally of glucose with some mannose and xylose (1.5–4.5%) and trace amounts of galactose. Linkage analysis showed that (1 → 3)-linked Glcp is the major chain constituent (67–91%), in addition to (1 → 4)-linked Glcp (3–27%) (Table 2). The FT-IR spectra (Fig. 1a) suggested the presence of α-(1 → 3) linkages. All spectra showed two bands at 848.6–849.02 and 822.99–823.38 cm−1, which indicated that the isolated materials contained α-(1 → 3)-d-glucans (Jelsma and Kreger 1979). The 1H NMR spectra of the polysaccharide preparations (Fig. 1b) showed a signal resonance (at δ 5.54–5.61 ppm) assigned to α-(1 → 3)-d-glucosidic linkages. The above data indicate that the WIP extracted from fruiting bodies of the selected macromycetes fungi were α-(1 → 3)-d-glucans. To increase solubility of the α-glucans in water, the carboxymethylation was applied. This procedure gave CM-α-(1 → 3)-d-glucans with the following DS: 0.96, 0.81, 0.65, and 0.57 for Lentinus edodes, Pleurotus ostreatus, Piptoporus betulinus, and Laetiporus sulphureus, respectively. The CM derivatives were used for further experiments.
Table 1.
Glycosyl residue composition of water-insoluble polysaccharides
| Species | Glc (%) | Man (%) | Xyl (%) | Gal (%) |
|---|---|---|---|---|
| L. edodes | 75.9 | 19.5 | 4.5 | Trace |
| P. ostreatus | 84.8 | 10.8 | 4.3 | – |
| P. betulinus | 94.3 | 4.1 | 1.5 | – |
| L. sulphureus | 93.4 | 5 | 1.5 | – |
Table 2.
Summary of methylation analysis of water-insoluble polysaccharides extracted from macromycetes fungi
| Species | Linkages | Feature | Molar amounts (%) |
|---|---|---|---|
| L. edodes | Glcp-(1→ | Non-reducing end | 0.9 |
| →3)-Glcp-(1→ | α-(1→3)-Bond | 67 | |
| →4)-Glcp-(1→ | α-(1→4)-Bond | 27.3 | |
| →3,6)-Glcp-(1→ | Branching site | 0.9 | |
| →4,6)-Glcp-(1→ | Branching site | 3.9 | |
| P. ostreatus | Glcp-(1→ | Non-reducing end | 4 |
| →3)-Glcp-(1→ | α-(1→3)-Bond | 82.8 | |
| →4)-Glcp-(1→ | α-(1→4)-Bond | 7.4 | |
| →3,6)-Glcp-(1→ | Branching site | 4.5 | |
| →4,6)-Glcp-(1→ | Branching site | 1.3 | |
| P. betulinus | Glcp-(1→ | Non-reducing end | 3.3 |
| →3)-Glcp-(1→ | α-(1→3)-Bond | 84.6 | |
| →4)-Glcp-(1→ | α-(1→4)-Bond | 6 | |
| →3,6)-Glcp-(1→ | Branching site | 1.9 | |
| →4,6)-Glcp-(1→ | Branching site | 4.2 | |
| L. sulphureus | Glcp-(1→ | Non-reducing end | 3.3 |
| →3)-Glcp-(1→ | α-(1→3)-Bond | 91.2 | |
| →4)-Glcp-(1→ | α-(1→4)-Bond | 3 | |
| →3,6)-Glcp-(1→ | Branching site | 1.5 | |
| →4,6)-Glcp-(1→ | Branching site | 1 |
Fig. 1.
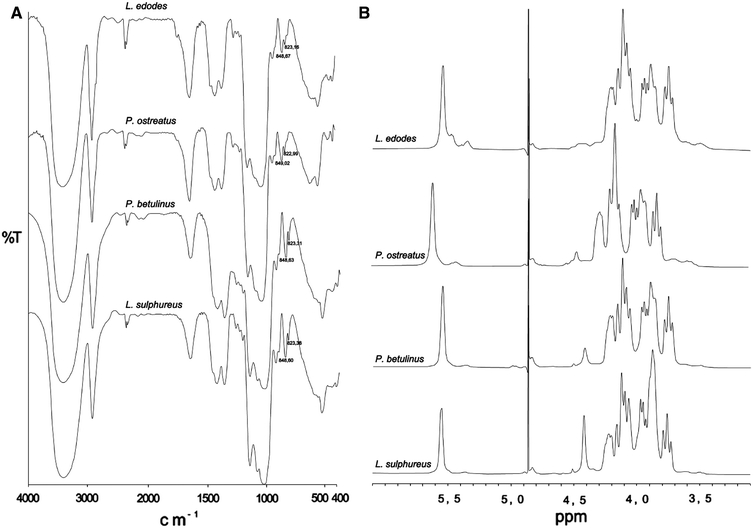
FTIR (a) and 1H NMR (b) spectra of water-insoluble, alkali-soluble polysaccharides (WIP) obtained from macromycetes fungi, in 0.9 M NaOH in D2O at 60°C
Viability of cells analysis and free radical scavenging activity
Before introducing new substances into clinical application, they should be examined both in in vitro and in vivo tests. One of the in vitro methods is potential cytotoxic or cytostatic activity of tested compounds analysis. It should be performed to establish whether they can be safely used in the treatment and if so, which concentrations are adequate to treat certain diseases. Therefore, the intention of carrying out cytotoxicity tests in vitro is to initially determine whether analyzed compounds are active towards, e.g. transformed cells and eventual injurious action or influence on proper metabolic functions of normal cells.
In the present study, the cytotoxic activity of CM-α-(1 → 3)-d-glucans was measured using the MTT and NR assays. As a positive control we used cisplatin. The IC50 values (drug concentration inhibiting 50% of cell growth) for the MTT test were 2.45, 1.76, >30 μg ml−1 for HeLa, Jurkat and HSF cells, respectively. For the NR assay they were 0.92, 0.53,>25 μg ml−1 for HeLa, Jurkat and HSF cells, respectively.
MTT analysis of cell viability showed that the CM-α-(1 → 3)-d-glucans, at 25–250 μg ml−1, decreased the metabolism of normal HSF cells only slightly (Fig. 2), but had a considerable effect on the viability of the tumor cells HeLa and Jurkat. CM polymers from L. edodes and P. ostreatus decreased the intensity of the mitochondrial metabolism of HSF cells by approximately 30 and 25%, respectively. The CM-glucans from P. betulinus and L. sulphureus did not show any cytotoxic activity against normal cells.
Fig. 2.
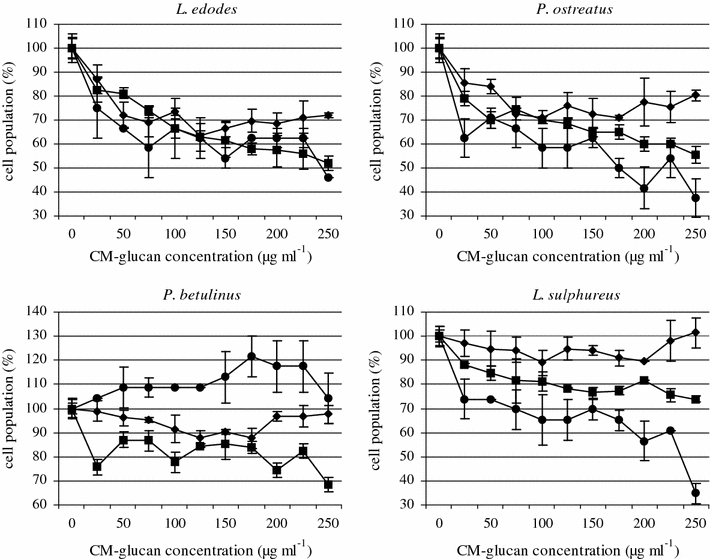
The effect of 24 h treatment of Hela (filled square), Jurkat (filled circle) and HSF (filled diamond) cells (2 × 105 cells ml−1) with CM-α-(1→3)-d-glucan from macromycetes fungi. The MTT assay. Bars represent the standard deviations of three independent experiments. The results are shown as percentage of the control, arbitrarily set to 100% (Y axis). The graph describes changes in cell population after incubation with CM-glucans
All the tested CM-α-(1 → 3)-d-glucans were toxic against the HeLa tumor cell line. Tumor cell viablility decreased on average by 35% after incubation with L. edodes and P. ostreatus. HeLa cell viability, after incubation with CM-glucans from P. betulinus and L. sulphureus, was 80% regardless of the applied CM-glucan concentration. The tested CM-α-(1 → 3)-d-glucans showed the highest toxicity when applied to Jurkat human leukemia cells. Their viability decreased by about 40% when the CM-glucans isolated from L. edodes and P. ostreatus were used. The CM-glucan from P. ostreatus induced this effect at the lowest of the applied concentrations (25 μg ml−1), while the CM-polymer from L. edodes was most toxic at 75 μg ml−1. The CM-α-(1 → 3)-d-glucans from L. sulphureus decreased the viability of Jurkat cells on average by 30%. By contrast, the CM-glucan from P. betulinus, stimulated the mitochondrial metabolism of tumor cells.
An NR-uptake assay showed that the tested CM-α-(1 → 3)-d-glucans stimulated HSF cell proliferation. Higher CM-glucan concentrations (>75 μg ml−1) also induced Jurkat leukemia cell proliferation and viability (Fig. 3). A high viability and proliferative activity of normal cells after incubation with the CM-glucans was detected, though the polymers from P. ostreatus and L. sulphureus had a slightly weaker effect than the CM-glucans from L. edodes i P. betulinus. For all the tested CM-α-(1 → 3)-d-glucans, a maximum stimulatory effect was caused by CM-glucan concentrations which did not exceed 100 μg ml−1.
Fig. 3.
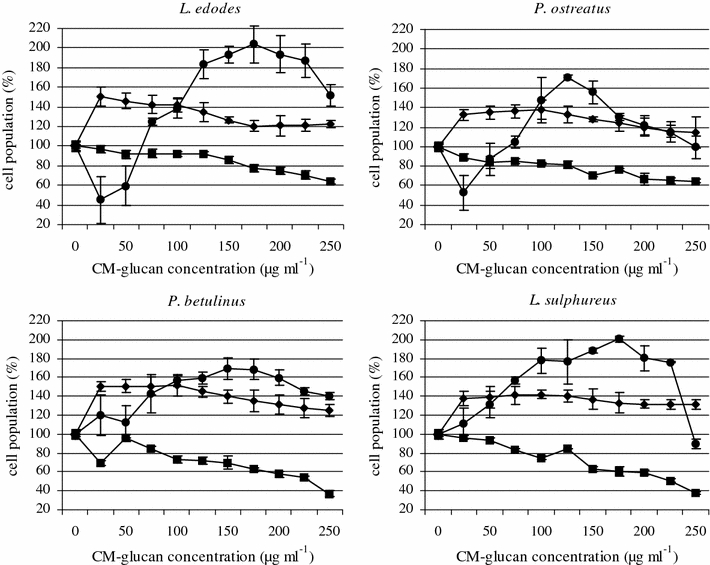
The effect of 24 h treatment of Hela (filled square), Jurkat (filled circle) and HSF (filled diamond) cells (2 × 105 cells ml−1) with CM-α-(1→3)-d-glucan from macromycetes fungi. The NR assay. Bars represent the standard deviations of three independent experiments. The results are shown as percentage of the control, arbitrarily set to 100% (Y axis). The graph describes changes in cell population after incubation with CM-glucans
On the basis of these results, one may suggest that the values obtained in the MTT assay should be interpreted only as indicating mitochondrial metabolism impairment rather than a toxic effect. The NR analysis confirmed the toxicity of all tested CM-α-(1 → 3)-d-glucans against HeLa cells. The viability of these cells decreased to 50% when the CM-glucans (200 μg ml−1) from P. betulinus and L. sulphureus were used. The CM-glucans from L. edodes and P. ostreatus acted more weakly, decreasing the viability of HeLa cells by 30 and 40%, respectively. Such decreases in viability may be associated with the ability of the modified α-glucan derivatives to alter the structure of membrane proteins in tumor cells. This may lead to adhesion disturbances, as was shown for sulphuric derivatives of α-glucans by Zhang and Cheung (2002). Possibly the carboxymethylated α-glucans use the same mechanism.
Although the addition of CM-glucans generally induced the proliferation of Jurkat cells, low concentrations (25 and 50 μg ml−1) of L. edodes and P. ostreatus decreased leukemia cell viability by about 50%. The most powerful activity was detected for the CM-glucans isolated from L. edodes and L. sulphureus. This effect may be explained by specific receptor binding by α-glucans and stimulatory or inhibitory signal transduction. As it has already been shown, the molecule involved in this process may be dectin-1. Although dectin-1 is a main receptor of monocytes, macrophages, and neutrophils for β-glucan, some α-glucans also bind to this molecule and provide a proliferation modulating signal (Rappleye et al. 2007; Taylor et al. 2002). The anti-tumor effect of the polysaccharides could be a consequence of stimulation of the cell-mediated immune response (Li et al. 2008). For example, a polysaccharide-rich fraction from Ganoderma lucidum had a strong stimulatory effect on both macrophages and T-lymphocytes (Liu et al. 2004). Besides, polysaccharides can, in a dose-dependent manner, directly inhibit the proliferation of leukemia, lymphoma, or selected solid tumor-derived cell lines (Ho et al. 2004). These effects are strongly associated with local cytokine network but also depend on the cell-cycle stage.
Induction of cell-mediated immune responses by glucans are mainly based on the production of interferon-γ (IFN-γ) and interleukin-2 (IL-2) which cause increased T cell proliferation (Ho et al. 2004). Because Jurkat, Clone E6-1 cells are known to produce large amounts of IL-2 after stimulation, the addition of CM-glucan in our study may have induced cell proliferation, as was found in NR test. Moreover, the different effect of CM-glucans on the proliferation and viability of solid tumor-derived HeLa cells and Jurkat cells could be a result of differences in growth conditions. Cells growing in suspension and those growing in an adherent mode show different sensitivities to cytotoxic or stimulatory effects of substances. This may be associated with differences in the length of cell cycle phases or the presence of specific receptors, e.g., adhesion molecules, in cellular membranes. It may, therefore, determine differences in intracellular signal transduction, which may influence the sensitivity of HeLa and Jurkat cells to the tested CM-glucans.
The CM-α-(1 → 3)-d-glucans tested in the present study did not show free radical scavenging activity (DPPH method). However, the lowest concentrations (25 μg ml−1) of the CM-glucans isolated from L. edodes, P. betulinus and L. sulphureus showed a slight scavenging action (Table 3).
Table 3.
DPPH scavenging effect (%)
| Concentration (μg ml−1) | L. edodes | P. ostreatus | P. betulinus | L. sulphureus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % of control | SD | P | % of control | SD | P | % of control | SD | P | % of control | S.D. | P | |
| 25 | 4 | 1.8 | ** | 4 | 0.9 | ** | 1.3 | 0.5 | * | −0.3 | 0.6 | |
| 50 | −3.2 | 1.5 | −2.7 | 0.9 | * | −0.5 | 1.1 | −1.3 | 0.7 | |||
| 75 | −3.2 | 0.6 | ** | −3.5 | 0.5 | ** | −0.5 | 2.1 | −0.9 | 0.8 | ||
| 100 | −3.2 | 0.6 | *** | −3.3 | 0.9 | ** | −0.6 | 0.9 | −1.5 | 0.8 | ||
| 125 | −3.2 | 1.4 | −4.1 | 1.6 | * | −0.4 | 0.7 | −1.6 | 0.3 | ** | ||
| 150 | −2.8 | 1.2 | −4 | 1.3 | * | −0.2 | 0.9 | −1.6 | 0.4 | ** | ||
| 175 | −3 | 1.7 | −3.9 | 1.8 | * | −0.1 | 0.5 | −2.2 | 0.9 | * | ||
| 200 | −2.8 | 0.9 | * | −4.9 | 1.4 | ** | −1.9 | 2.4 | −2.4 | 0.5 | ** | |
Negative values represent the % of ROS reduction, positive values should be considered as lack of radical scavenging. P < 0.05 (*), P < 0.001 (**), P < 0.0001 (***) significance compared to control. Results are shown as mean ± SD of three independent experiments
The % of DPPH• radical reduced by CM-α-(1→3)-d-glucan from macromycetes fungi is compared to the control (0% of reduction)a
In conclusion: carboxymethylated fungal α-(1 → 3)-d-glucans have a biological activity potential, i.e. they express cytotoxic or mitochondrial metabolism-modulating effects but generally do not show free radical scavenging activity. These activities, however, should be further tested to be considered for application.
Conflict of interest
The authors declare that they have no conflicts of interest.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
References
- Bao X, Duan J, Fang X, Fang J. Chemical modifications of the (1 → 3)-α-d-glucan from spores of Ganoderma lucidum and investigation of their physicochemical properties and immunological activity. Carbohydr Res. 2001;336:127–140. doi: 10.1016/S0008-6215(01)00238-5. [DOI] [PubMed] [Google Scholar]
- Bohn JA, BeMiller JN. (1–3)-β-d-glucans as biological response modifiers: a review of structure-functional activity relationships. Carbohydr Polym. 1995;28:3–14. doi: 10.1016/0144-8617(95)00076-3. [DOI] [Google Scholar]
- Hakomori S. Rapid permethylation of glycolipids and polysaccharides catalysed by methyl carbon. J Biochem. 1964;55:205–208. [PubMed] [Google Scholar]
- Ho FFL, Klosiewicz DL. Proton nuclear magnetic spectroscopy for determination of substituents and their distribution in carboxymethylcellulose. Anal Chem. 1980;52:913–916. doi: 10.1021/ac50056a032. [DOI] [Google Scholar]
- Ho CY, Lau CBS, Kim CF, Leung KN, Fung KP, Tse TF, Chan HHL, Chow MSS. Differential effect of Coriolus versicolor (Yunzhi) extract on cytokine production by murine lymphocytes in vitro. Int Immunopharmacol. 2004;4:1549–1557. doi: 10.1016/j.intimp.2004.07.021. [DOI] [PubMed] [Google Scholar]
- Jelsma J, Kreger DR. Polymorphism in crystalline (1 → 3)-α-d-glucan from fungal cell-walls. Carbohydr Res. 1979;71:51–64. doi: 10.1016/S0008-6215(00)86060-7. [DOI] [PubMed] [Google Scholar]
- Kiho T, Yoshida I, Katsuragawa M, Sakushima M, Usui S, Ukai S. Polysaccharides in Fungi XXXIV: a polysaccharide from fruiting bodies of Amanitamuscaria and the antitumor activities of its carboxymethylated product. Biol Pharm Bull. 1994;17:1460–1462. doi: 10.1248/bpb.17.1460. [DOI] [PubMed] [Google Scholar]
- Li SG, Wang DG, Tian W, Wang XX, Zhao JX, Liu Z, Chen R. Characterization and anti-tumor activity of a polysaccharide from Hedysarum polybotrys Hand.-Mazz. Carbohydr Polymers. 2008;73:344–350. doi: 10.1016/j.carbpol.2007.12.001. [DOI] [Google Scholar]
- Lindequist U, Niedermeyer THJ, Julich WD. The pharmacological potential of mushrooms. eCAM. 2005;2:285–299. doi: 10.1093/ecam/neh107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu JJ, Huang TS, Hsu ML, Chen ChCh, Lin WS, Lue FJ, Chang WH. Antitumor effects of the partially purified polysaccharides from Antrodia camphorata and the mechanism of its action. Toxicol Appl Pharmacol. 2004;201:186–193. doi: 10.1016/j.taap.2004.05.016. [DOI] [PubMed] [Google Scholar]
- Moradali MF, Mostafavi H, Ghods S, Hedjaroude GA. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi) Int Immunopharmacol. 2007;7:701–724. doi: 10.1016/j.intimp.2007.01.008. [DOI] [PubMed] [Google Scholar]
- Ng ML, Yap AT. Inhibition of human colon carcinoma development by lentinan from shiitake mushrooms (Lentinus edodes) J Altern Complement Med. 2002;8:581–589. doi: 10.1089/107555302320825093. [DOI] [PubMed] [Google Scholar]
- Paduch R, Matysik G, Wójciak-Kosior M, Kandefer-Szerszeń M, Skalska-Kamińska A, Nowak-Kryska M, Niedziela P. Lamium album extracts express free radical scavenging and cytotoxic activities. Pol J Environ Stud. 2008;17:569–580. [Google Scholar]
- Rappleye CA, Eissenberg LG, Goldman WE. Histoplasma capsulatum α-(1 → 3)-glucan blocks innate recognition by the β-glucan receptor. PNAS. 2007;104:1366–1370. doi: 10.1073/pnas.0609848104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russa R, Urbanik-Sypniewska T, Lindström K, Mayer H. Chemical characterization of two lipopolysaccharide species isolated from Rhizobium loti NZP2213. Arch Microbiol. 1995;163:345–351. doi: 10.1007/BF00404207. [DOI] [PubMed] [Google Scholar]
- Sarkar S, Koga J, Whitley RJ, Chatterjee S. Antiviral effect of the extract of culture medium of Lentinus edodes mycelia on the replication of herpes simplex virus 1. Antivir Res. 1993;20:293–303. doi: 10.1016/0166-3542(93)90073-R. [DOI] [PubMed] [Google Scholar]
- Sasaki T, Abiko N, Nitta K. Antitumor activity of carboxymethyloglucans obtained by carboxymethylation of (1 → 3)-β-d-glucan from Alcaligenes faecalis var myxogenes IFO13140. Eur J Cancer Clin Oncol. 1979;15:211–215. doi: 10.1016/0014-2964(79)90062-8. [DOI] [PubMed] [Google Scholar]
- Taylor PR, Brown GD, Reid DM, Willment JA, Martinez-Pomares L, Gordon S, Wong SYC. The β-glucan receptor, dectin-1, is predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J Immunol. 2002;169:3876–3882. doi: 10.4049/jimmunol.169.7.3876. [DOI] [PubMed] [Google Scholar]
- York WS, Darvill AG, McNeil M, Stevenson TT, Albersheim P (1986) Isolation and characterization of plant cell walls and cell wall components. In: Weissbach A, Weissbach H (eds) Methods enzymol. Academic Press, London 118:3–40
- Zhang P, Cheung PCK. Evaluation o-sulfated Lentinus edodes α-(1 → 3)-d-glucan as a potential antitumor agent. Biosci Biotechnol Biochem. 2002;66:1052–1056. doi: 10.1271/bbb.66.1052. [DOI] [PubMed] [Google Scholar]