

Abstract
Wind and ocean currents may potentially have important effects on travelling animals, as an animal which does not respond to lateral flow will be drifted from its intended direction of movement. By analysing daily movements of migrating ospreys Pandion haliaetus and marsh harriers Circus aeruginosus, as recorded by satellite telemetry, in relation to global wind data, we showed that these raptors allow on average 47 per cent drift. Furthermore, our analyses revealed significant geographical and temporal variation in the response to crosswinds. During some parts of the migration, the birds drifted and in other parts they compensated or even overcompensated. In some regions, the response of marsh harriers depended on the wind direction. They drifted when the wind came from one side and (over)compensated when the wind came from the opposite side, and this flexible response was different in different geographical regions. These results suggest that migrating raptors modulate their response to crosswinds at different places and times during their travels and show that individual birds use a much more varied repertoire of behavioural responses to wind than hitherto assumed. Our results may also explain why contrasting and variable results have been obtained in previous studies of the effect of wind on bird migration.
Keywords: bird orientation, migration, satellite tracking, wind drift
1. Introduction
Animals often move through or on a medium that is in motion itself. Examples include turtles and fishes swimming in water [1], birds and insects flying in air [2,3] and polar bears walking on drifting ice [4]. The resulting movement of an animal (the track vector) is the sum of the animal vector (heading and speed of the animal) and the flow vector (direction and strength of the flow; [5]). An animal that does not account for lateral flow will be drifted from the intended direction of movement. However, by changing the heading, it is possible to compensate for the flow as long as the speed of locomotion is equal to or larger than the speed of the flow [6] and provided that the animal can estimate the extent and direction of the flow or its own displacement. Relatively fast moving animals can thus decide whether they want to drift, partially drift, compensate (i.e. maintain the intended direction), or overcompensate (figure 1a). Relatively weak fliers, showing flight speeds generally much lower than wind speeds, such as insects, only have a small to moderate possibility to influence their direction of movement while aloft [3,7], apart from selecting circumstances when and where winds are profitable [8].
Figure 1.

(a) Triangles of velocities illustrating the speed and direction of the animal (animal heading), speed and direction of wind (wind vector) and speed and direction of the resulting movement (track) in relation to the direction to the goal (intended direction). An animal which does not account for wind will experience lateral drift. By changing the heading in relation to the direction of the wind the animal can maintain the intended direction (compensation), or even overcompensate. (b) Relationship between perpendicular movement rate and perpendicular wind component (in relation to the general preferred direction) for ospreys (grey dots) and marsh harriers (open dots). Coloured frames indicate segments classified as drift (green panels), compensation (blue panels) and overcompensation (orange panels). Data for regions and seasons were combined (see table 1 for further details).
For birds, geographical wind patterns have a most important effect on the evolution of migration routes [9–12]. However, little is known about how individual birds respond to lateral wind drift during migration (reviews in [2,5,13]). Thorup et al. [13] demonstrated that the effect of wind on migrating raptors was age-dependent, with adults showing partial drift and juveniles almost full drift. Juveniles, who do not migrate towards a specific goal but rather in a certain endogenous direction [14], should invest all their own movement speed in the intended direction, which implies full drift. For experienced adults, who navigate to specific goal areas, partial drift is thought to be an adaptive strategy as this minimizes the duration of the travel [15]. Moreover, for adults, almost full drift is expected far away from the destination, but as they approach their destination, they should gradually compensate more and more. In order to reach a specific goal, they might even have to overcompensate in the end [15]. During barrier crossings, birds should make every effort to move in the intended direction in order to minimize crossing duration, and thus full drift is expected. Finally, (land)birds are expected to compensate and overcompensate near coasts, if they face the risk of being drifted offshore [15].
A general problem in the study of drift is the difficulty of interpreting whether a certain movement of an animal is drift, compensation or overcompensation without knowledge about the intended direction of the animal. A correlation between crosswind and track directions is not per se proof for (partial) wind drift, as such correlations may also arise if populations with different intended track directions selectively travel on days with tailwinds [16,17]. In this paper, we avoid the complications of pseudodrift by analysing daily movements of adult ospreys Pandion haliaetus and marsh harriers Circus aeruginosus tracked by satellite telemetry [18–20]. As these diurnal, long-distance migrants are faithful to breeding and wintering sites, we know the intended direction of migration for each individual. As a consequence, we are able to quantify, on a daily basis, how much the birds deviate from this direction and how this correlates to perpendicular wind components. This allows us to evaluate how much, where, and when the birds drift and (over)compensate along their migratory routes, in relation to barriers such as seas, oceans, mountain ranges and deserts, and also to the distance to the destination. Thus, we have the opportunity to investigate whether birds are flexible in their response to crosswinds, adjusting to local circumstances. More specifically we test the predictions that: (i) Birds drift extensively at the onset of migration (far away from the goal) and (over)compensate during the final approach to the destination, (ii) birds drift fully during barrier crossings, and (iii) birds compensate or overcompensate when they face the risk of being drifted into hazardous habitats.
2. Material and methods
(a). Tracking studies
Between 1996 and 2008 16 adult ospreys and 11 adult marsh harriers were tracked by satellite telemetry between their breeding grounds in Sweden and wintering grounds in (west) Africa. Some of these birds were followed during several years, and thus the entire dataset includes 33 autumn and 12 spring journeys for the ospreys and 24 autumn and 13 spring journeys for the marsh harriers. Different types of satellite transmitters, with different transmission schedules, were used in different years and for different species. The first transmitter models provided locations every third day, whereas later models provided locations almost daily (see [18,20] for details). Transmitters were tracked by the ARGOS system (CLS, Toulouse, France). Validated locations are divided into different classes (A, B and 0–3), corresponding to different location accuracies (see www.argos-system.org/manual). High quality locations (1–3) were always included, low quality locations (0, A, B) only if not obviously off-track (implying unrealistic travel speeds). Tracks were divided into segments spanning one to several days by selecting the location with the highest accuracy for every night (i.e. between the hours of 18.00 and 06.00, when these diurnal migrants are stationary [21,22]). When several locations of the same accuracy were available, the location closest to midnight was selected. Nocturnal migration is very rare in the species studied (less than 1% of all segments [21]), and too few cases of nocturnal migration were recorded to warrant a separate analysis. In order to avoid possible effects of stopover behaviour, we excluded segments shorter than 50 km d−1. Furthermore, we excluded segments longer than 4 days, as wind strength and direction is likely to change during such long time intervals.
(b). Wind data
Wind data were obtained from the NCEP/NCAR Reanalysis project, as provided by the NOAA/OAR/ESRL PSD, Boulder, CO, USA (http://www.cdc.noaa.gov). These data consist of west–east (u-winds) and south–north (v-winds) wind components, which were combined into single wind vectors (i.e. direction and strength of the wind). Wind data were extracted for a pressure level of 925 hPa, which corresponds to an altitude of about 750 m a.s.l. This altitude was chosen as 95 per cent of the movements of ospreys and marsh harriers occur at altitudes between 0 and 1500 m (average altitude is 670 m a.s.l. for 3006 GPS registrations of migrating ospreys and marsh harriers; R.H.G. Klaassen 2007–2009, unpublished data). Wind data were interpolated from the grid data for the midpoint of the segments, at the hours of 06.00, 12.00 and 18.00. Data were averaged in which noon values were given twice as much weight as the morning and afternoon values. In this way, we tried to mimic, in a simplified manner, the wind conditions the birds experienced during the day, given that these birds travel between the hours of 06.00 and 18.00 [21,22].
(c). Analyses
We calculated the overall direction of migration, i.e. the direction between breeding and wintering sites, for each individual bird. Ospreys often have intermediate goals, such as stopover sites in Europe [19]. Thus, for the ospreys, we calculated the overall directions between the breeding and stopover sites and between the stopover and wintering sites separately. We then calculated the forward and perpendicular component of the movement (in km d−1) in relation to the overall intended direction [13]. Similarly, forward and perpendicular components were calculated for the wind vector. To confirm that the birds were affected by wind, we correlated forward rate of movement with the forward wind component (tailwind). Lateral drift was quantified by correlating perpendicular rate of movement with the perpendicular wind component (crosswind).
Based on the relationship between crosswind and perpendicular movement, we distinguished between the following behaviours (also defined in figure 1): (i) drift, i.e. perpendicular movement more than 50 km d−1 or less than −50 km d−1 (i.e. more negative than −50 km d−1) with similar signs for perpendicular movement and perpendicular wind, (ii) compensation or small influence of crosswinds, i.e. perpendicular movement less than 50 km d−1 and more than −50 km d−1, and (iii) overcompensation, i.e. perpendicular movement more than 50 km d−1 or less than −50 km d−1 with opposite signs for perpendicular movement and perpendicular wind. We consider a perpendicular movement of 50 km d−1 to be a substantial deviation from the intended direction of movement for migrating ospreys and marsh harriers, which, on average, cover about 200–250 km d−1 [20,21].
In order to investigate the influence of geographical patterns on the response to crosswinds, we calculated the frequency of drift, compensation and overcompensation within 10° latitudinal bands along the migration routes of the two species. Chi-square tests were then performed to check whether the frequencies of the different behaviours differed between these latitudinal bands. In case the sample size for a certain band was too small, we only distinguished two categories: Europe (latitude north of 36° N) and Africa (latitude south of 36° N).
3. Results
Daily movements of both ospreys and marsh harriers were strongly affected by wind. For every m s−1 of tailwind, the forward rate of movement increased by, on average, 15.3 and 13.8 km d−1 for ospreys and marsh harriers, respectively. The slope of the relationship between perpendicular movement and crosswinds was less steep; for every m s−1 of crosswind, the rate of perpendicular movement increased by, on average, only 6.7 and 6.5 km d−1 for ospreys and marsh harriers, respectively (table 1). Slopes and intercepts for forward and perpendicular regressions did not differ between weak winds (winds less than 3.5 m s−1, equal to the 33rd percentile) and strong winds (winds more than 6.1 m s−1, equal to the 66th percentile; data not shown). Ospreys and marsh harriers thus drifted partially with crosswinds, allowing about 44 and 47 per cent drift, respectively (autumn and spring migration combined). The overall figure for the two species was 47 per cent (table 1).
Table 1.
Slopes and intercepts for the regression of forward rates of movement (km d−1) in relation to forward wind components (m s−1) and of perpendicular rates of movement (km d−1) in relation to perpendicular wind components (m s−1), for ospreys and marsh harriers during different seasons and in different regions. 95 per cent confidence limits are provided for regression slopes. Numbers are printed in bold if there was a significant difference between Europe and Africa in the slope of the perpendicular regression. We also tested whether the slopes of the forward and perpendicular regressions differ from each other (test between slopes; *, p < 0.05, n.s., not significant). The last column provides the estimated drift effect, which is the slope of the perpendicular regression divided by the slope of the forward regression. A ratio equal to 1 means that the perpendicular drift is equal to the effect of wind on forward speed (full drift), a ratio equal to 0 means that there is no perpendicular drift (full compensation), and a ratio in between 0 and 1 implies partial drift.
| n | regression forward movement versus wind |
regression perpendicular movement versus wind |
test between slopes | ratio between perpendicular and forward slope | |||||
|---|---|---|---|---|---|---|---|---|---|
| slope (95% conf. lim.) | intercept (km d−1) | slope (95% conf. lim.) | intercept (km d−1) | ||||||
| osprey | autumn | Europe | 135 | 10.9 (5.7–16.1) | 195.4 | 6.1 (3.7–8.4) | −8.5 | n.s. | 0.56 |
| Africa | 157 | 21.7 (16.6–26.7) | 208.8 | 3.7 (0.27–7.0) | −8.4 | * | 0.17 | ||
| total | 292 | 16.0 (12.6–19.4) | 208.7 | 5.2 (3.2–7.1) | −7.6 | * | 0.33 | ||
| spring | Europe | 58 | 13.4 (4.4–22.5) | 203.5 | 6.3 (1.6–11.0) | 1.7 | n.s. | 0.47 | |
| Africa | 81 | 20.6 (15.6–25.6) | 298.1 | 15.8 (9.1–22.6) | 4.1 | n.s. | 0.77 | ||
| total | 139 | 16.2 (11.4–20.9) | 253.9 | 10.5 (6.6–14.5) | 8.3 | n.s. | 0.65 | ||
| seasons combined | 431 | 15.3 (12.5–18.1) | 222.9 | 6.7 (4.9–8.6) | −3.6 | * | 0.44 | ||
| marsh | autumn | Europe | 136 | 16.4 (12.7–20.2) | 182.7 | 5.9 (4.1–7.6) | −9.5 | * | 0.36 |
| harrier | Africa | 130 | 14.6 (9.4–19.7) | 180.8 | 12.1 (8.0–16.2) | 12.9 | n.s. | 0.83 | |
| total | 266 | 15.7 (12.8–18.6) | 180.9 | 7.2 (5.3–9.0) | −1.7 | * | 0.46 | ||
| spring | Europe | 70 | 8.9 (2.3–15.4) | 192.9 | 1.6 (−0.94–4.0) | 0.1 | * | 0.18 | |
| Africa | 90 | 16.9 (13.1–20.6) | 223.5 | 11.4 (8.4–14.3) | −23.6 | * | 0.67 | ||
| total | 160 | 12.1 (8.7–15.5) | 203.4 | 6.0 (4.1–8.0) | −1.4 | * | 0.50 | ||
| seasons combined | 426 | 13.8 (11.6–16.0) | 190.4 | 6.5 (5.2–7.9) | −1.6 | * | 0.47 | ||
| overall (species and seasons combined) | 857 | 14.2 (12.4–16.0) | 206.5 | 6.6 (5.5–7.2) | −2.6 | * | 0.47 | ||
During migration, the birds sometimes drifted, sometimes compensated, and sometimes overcompensated (figure 2). The frequency distribution of drift, compensation and overcompensation differed significantly between latitudinal intervals in autumn (osprey: 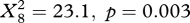 , marsh harrier:
, marsh harrier: 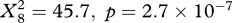 ) as well as in spring (osprey:
) as well as in spring (osprey: 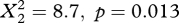 , marsh harrier:
, marsh harrier: 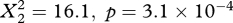 ). As sample sizes were too small for intervals in spring, we only defined two classes in the statistical tests (Europe and Africa; figure 2). At the onset of migration, i.e. far away from the goal, the birds often drifted, particularly in spring, and only rarely overcompensated (both in spring and autumn; figure 2). In contrast, at the end of migration, i.e. close to the goal, the birds rarely drifted (in spring and autumn), whereas overcompensation was frequently observed (only in autumn, especially in marsh harriers; figure 2). Moreover, during crossings of barriers, such as the Alps, the Mediterranean Sea and the Sahara Desert, the birds often overcompensated, and there was no clear dominance of drift in these regions (figure 2). Due to drift at the onset of migration, many marsh harriers came close to the coast of Morocco in spring where they strongly overcompensated.
). As sample sizes were too small for intervals in spring, we only defined two classes in the statistical tests (Europe and Africa; figure 2). At the onset of migration, i.e. far away from the goal, the birds often drifted, particularly in spring, and only rarely overcompensated (both in spring and autumn; figure 2). In contrast, at the end of migration, i.e. close to the goal, the birds rarely drifted (in spring and autumn), whereas overcompensation was frequently observed (only in autumn, especially in marsh harriers; figure 2). Moreover, during crossings of barriers, such as the Alps, the Mediterranean Sea and the Sahara Desert, the birds often overcompensated, and there was no clear dominance of drift in these regions (figure 2). Due to drift at the onset of migration, many marsh harriers came close to the coast of Morocco in spring where they strongly overcompensated.
Figure 2.

Behaviour in relation to crosswinds along the migratory route for two species of raptors. Every segment represents a migratory journey of 1–4 days. The colours of the segments indicate whether it was an occasion of drift (green), compensation (blue), or overcompensation (orange). For classification criteria, see figure 1 and main text. (a) Ospreys in autumn, (b) ospreys in spring, (c) marsh harriers in autumn and (d) marsh harriers in spring. The corresponding bars indicate the frequency distribution of different responses to crosswind for different latitudinal bands (°N, y-axis). Numbers to the right of the bars indicate sample sizes.
This geographical and temporal variation in the response to crosswinds is also apparent from the regression analyses (table 1). As the regression slope between perpendicular wind and perpendicular movement summarizes the overall pattern, a strong element of drift will result in a steeper slope (e.g. ospreys in Africa during spring), whereas a strong element of (over)compensation will result in a more gentle slope (e.g. marsh harriers in Europe during spring). Indeed, there were differences in the slopes for the perpendicular regressions between regions, indicating that the balance between drift and compensation changed throughout the migrations. For three out of four comparisons between regions, the extent of drift was significantly larger in Africa than in Europe (table 1).
In an additional analysis, we investigated the response in behaviour of the raptors to crosswinds coming from different directions, i.e. westerly and easterly winds (figure 3). This analysis revealed that marsh harriers responded differently depending on the wind direction. In northern Europe in autumn, at the onset of migration, the harriers drifted with easterly winds and compensated for westerly winds (latitudinal band 50–60°; frequency distribution of drift, compensation and overcompensation differed between easterly and westerly winds, 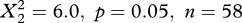 ). In (southern) Spain, the harriers showed the opposite behaviour, drifting with westerly winds and (over)compensating for easterly winds (latitudinal band 30–40°; the frequency distribution differed between easterly and westerly winds,
). In (southern) Spain, the harriers showed the opposite behaviour, drifting with westerly winds and (over)compensating for easterly winds (latitudinal band 30–40°; the frequency distribution differed between easterly and westerly winds, 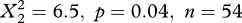 ). No significant differences were found for other latitudinal bands in autumn, whereas no analyses could be done for the southernmost section (10–20°) and for spring owing to small sample sizes. For ospreys we could not detect any differences in the response to westerly and easterly winds for any region or season (see the electronic supplementary material, figure S1).
). No significant differences were found for other latitudinal bands in autumn, whereas no analyses could be done for the southernmost section (10–20°) and for spring owing to small sample sizes. For ospreys we could not detect any differences in the response to westerly and easterly winds for any region or season (see the electronic supplementary material, figure S1).
Figure 3.

Response to crosswinds along the migratory route of marsh harriers, facing negative and positive perpendicular winds (winds from the east and west, respectively). (a) Easterly winds in autumn, (b) westerly winds in autumn, (c) easterly winds in spring and (d) westerly winds in spring. Squares highlight the two areas where the response to crosswinds was dependent on wind direction.
4. Discussion
(a). Response to crosswind
Although Trowbridge published his seminal paper on wind drift in migrating raptors as long ago as in 1902 [23], it was a common belief until the 1960s that migrating birds fly along the shortest possible route between breeding and wintering areas, compensating for drift and thus minimizing the need for complex navigational abilities [5]. However, as evidence for lateral drift accumulated, this viewpoint was revised. Instead, it was believed that nocturnal migrants generally cannot estimate the speed and direction of wind while aloft, resulting in full drift [2]. Diurnal migrants were also expected to drift, though only partially, as this seems to be the optimal strategy in an environment with changing wind conditions [15]. We here open a new chapter to this field of research, showing that individual birds may modulate their response to crosswinds in different areas and at different times during migration.
By correlating perpendicular wind components with perpendicular movement rates, we estimate that adult ospreys and marsh harriers generally allowed about 47 per cent drift. In other words, these raptors generally allow some effect of crosswind (partial drift). The novelty of our study is that the migrating raptors were not static in their response to wind, but changed between different behaviours during different legs of their journeys, including full drift, compensation and overcompensation. In agreement with our first prediction, the tendency to drift was most pronounced at the onset of migration, furthest away from the goal, whereas the birds (over)compensated to a higher degree when approaching their destination. Drift at the onset of migration was stronger in spring than in autumn, which may be explained by the fact that the birds were already relatively close to intermediate goals when they set off in autumn (intermediate goal areas are found in Europe). Contrary to our second prediction, the raptors (over)compensated during barrier crossings. Apparently it is more important for raptors to minimize crossing distance than crossing time. Finally, consistent with our third prediction, the birds strongly compensated near coasts when they faced the risk of being drifted out over the ocean (marsh harriers in spring). Thus, although the raptors did not always respond to crosswinds in the way we had anticipated, their behaviour was remarkably flexible.
An additional explanation for the great flexibility in the response to wind is that travelling animals can deal with lateral drift in very different ways. Firstly, they may compensate instantaneously, by changing their own heading into the wind until the resulting track aligns with the intended direction of movement. Secondly, they may overcompensate after a preceding instance of drift, i.e. overcompensation can also be regarded as a delayed reaction to drift. This is analogous to the idea that birds (partially) drift by strong winds at high altitude during the first part of the day, and correct for the displacement by overcompensating at low altitude, where winds are weaker, during the second part of the day [24]. Finally, birds can also start by overcompensating, anticipating drift during a later migratory leg. Such a strategy requires that wind conditions are known and predictable to the birds. It seems probable that this is the case for the adult, experienced birds which were tracked in this study.
A more detailed analysis revealed the intriguing result that, in certain geographical regions, marsh harriers responded differently to winds coming from different directions. In some regions, the harriers allowed drift if the wind came from one side, but maintained their intended directions when the wind came from the opposite direction. It seems probable that, in these regions, the harriers navigated to some intermediate goal area or tried to circumvent a barrier (for example some water body or a mountain range). It also suggests that the harriers made distinct decisions about whether to drift, compensate or overcompensate. Geographical wind patterns are nevertheless believed to play an important role in shaping migratory routes as they determine the most energy efficient trajectories [11,12]. It is not clear why ospreys did not respond in a similar way to winds coming from different directions. A possible explanation is that the ospreys tracked in this study travelled in a much wider migration corridor than the marsh harriers, making it more difficult to detect such patterns.
(b). Navigation and orientation
The fact that the raptors generally were subjected to wind drift implies that they do not travel between breeding and wintering sites along exactly the same routes. Thus, the birds seem to have good navigational skills, as they manage to return to very specific locations (breeding site, stopover sites, wintering site; [19]). The result that the behaviour of marsh harriers depended on wind direction indicates that the birds tracked were experienced and had good knowledge about their current location and where they wanted to go. This supports the idea that these birds travel by map-based navigation [25].
At some occasions, the raptors maintained their intended direction, i.e. they compensated completely for lateral drift. This implies that the birds were able to estimate drift, most probably by visual cues. Nocturnal migrants cannot use visual cues, particularly during overcast conditions, and are therefore expected to be drifted to a larger extent by crosswinds [2,5]. Nocturnal migrants are thus expected to be more limited in their response to crosswinds, facing a higher risk of being drifted off-course. In strong contrast to insects [26], birds are generally not believed to be capable of estimating the wind direction, for example via turbulence cues. However, the swift Apus apus is a noteworthy exception as swifts seem able to compensate for wind drift during high altitude nocturnal migration [27].
5. Conclusions
On first thought, flow might seem to be a nuisance for a travelling animal, as it may complicate the task to navigate towards a specific location (i.e. wintering or breeding sites). After all, an animal which ignores drift will end up at a place where it did not intend to go. However, flow also provides an opportunity to shorten the duration of a travel, i.e. an animal can actually benefit from it [15,28], and some great migratory journeys would be impossible to make without the assistance of flow [7,29,30]. Our results show that migrating raptors, in general, drift partially by wind. Unexpectedly, the responses of these diurnal migrants were, however, by no means static. Rather, they showed great flexibility in their behaviour. Apart from partial drift, we also regularly observed cases of full drift, compensation and overcompensation. The raptors also seemed to adapt their behaviour to local circumstances; drifting when favourable and compensating or overcompensating when approaching a goal or facing the risk of being blown into hazardous habitats. The flexibility in the response of the birds suggests that migrating raptors have excellent navigational skills and are able to estimate drift while aloft. Finally, flexibility in the behaviour of birds along their migratory routes might also explain why previous studies on how wind affects bird migration have reported contrasting results for different localities and at different times of the year [2,5,31]. Our results thus may help to unite a field of research that has been characterized by much conflicting evidence [32].
Acknowledgements
The study was approved by the Ethical Committees in Uppsala (C179/9) and Malmö/Lund (204-06). Permission to capture the birds was obtained from the Swedish Environmental Protection Agency (412-7093-08 Nv).
We thank Nils Kjellén and Patrik Olofsson for assistance in the field, and Microwave Telemetry Inc. for support and supplying excellent equipment. We wish to acknowledge the use of the Maptool program for graphics in this paper (www.seaturtle.org). We are grateful for the comments provided by Jason Chapman and two anonymous referees. This research was financed through grants from the Crafoord Foundation (to Nils Kjellén), the Swedish Research Council (to T.A. and to R K.) and from the Niels Stensen Foundation (to R.K.).
References
- 1.Lambardi P., Lutjeharms J. R. E., Mencacci R., Hays G. C., Luschi P. 2008. Influence of ocean currents on long-distance movement of leatherback sea turtles in the Southwest Indian Ocean. Mar. Ecol. Prog. Ser. 358, 289–301 10.3354/meps07346 (doi:10.3354/meps07346) [DOI] [Google Scholar]
- 2.Liechti F. 2006. Birds: blowin' by the wind? J. Ornithol. 147, 202–211 10.1007/s10336-006-0061-9 (doi:10.1007/s10336-006-0061-9) [DOI] [Google Scholar]
- 3.Chapman J. W., Reynolds D. R., Mouritsen H., Hill J. K., Riley J. R., Sivell D., Smith A. D., Woidod I. P. 2008. Wind selection and drift compensation optimize migratory pathways in a high-flying moth. Curr. Biol. 18, 514–518 10.1016/j.cub.2008.02.080 (doi:10.1016/j.cub.2008.02.080) [DOI] [PubMed] [Google Scholar]
- 4.Mauritzen M., Derocher A. E., Pavlova O., Oystein W. 2003. Female polar bears, Ursus maritimus, on the Barents Sea drift ice: walking the treadmill. Anim. Behav. 66, 107–113 10.1006/anbe.2003.2171 (doi:10.1006/anbe.2003.2171) [DOI] [Google Scholar]
- 5.Richardson W. J. 1991. Wind and orientation of migrating birds: a review. In Orientation in birds (ed. Berthold P.), pp. 226–249 Basel, Switzerland: Birkhäuser Verlag; [DOI] [PubMed] [Google Scholar]
- 6.Alerstam T. 1978. Analysis and a theory of visible bird migration. Oikos 30, 273–349 10.2307/3543483 (doi:10.2307/3543483) [DOI] [Google Scholar]
- 7.Chapman J. W., Nesbit R. L., Burgin L. E., Reynolds D. R., Smith A. D., Middleton D. R., Hill J. K. 2010. Flight orientation behaviors promote optimal migration trajectories in high-flying insects. Science 327, 682–685 10.1126/science.1182990 (doi:10.1126/science.1182990) [DOI] [PubMed] [Google Scholar]
- 8.Reynolds A. M., Reynolds D. R. 2009. Aphid aerial density profiles are consistent with turbulent advection amplifying flight behaviour: abandoning the epithet ‘passive’. Proc. R. Soc. B 276, 137–143 10.1098/rspb.2008.0880 (doi:10.1098/rspb.2008.0880) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Landsberg H. 1948. Bird migration and pressure patterns. Science 108, 708–709 10.1126/science.108.2817.708-a (doi:10.1126/science.108.2817.708-a) [DOI] [PubMed] [Google Scholar]
- 10.Gauthreaux S. A., Jr, Michi J. E., Belser G. G. 2005. The temporal and spatial structure of the atmosphere and the evolution of bird migration. In Birds of two worlds: the ecology and evolution of migration (eds Greenberg R., Marra P. P.), pp. 182–192 West Sussex, UK: John Hopkins University Press [Google Scholar]
- 11.Felicísimo A. M., Munoz J., González-Solis J. 2008. Ocean surface winds drive dynamics of transoceanic aerial movements. PLoS ONE 3, e2928. 10.1371/journal.pone.0002988 (doi:10.1371/journal.pone.0002988) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Klaassen R. H. G., Strandberg R., Hake M., Olofsson P., Alerstam T. 2010. Loop migration in adult marsh harriers Circus aeruginosus, as revealed by satellite telemetry. J. Avian Biol. 41, 200–207 10.1111/j.1600-048X.2010.05058.x (doi:10.1111/j.1600-048X.2010.05058.x) [DOI] [Google Scholar]
- 13.Thorup K., Alerstam T., Hake M., Kjellén N. 2003. Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proc. R. Soc. Lond. B 270, S8–S11 10.1098/rsbl.2003.0014 (doi:10.1098/rsbl.2003.0014) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gwinner E., Wiltschko W. 1978. Endogenously controlled changes in migratory direction of garden warbler, Sylvia borin. J. Comp. Physiol. 125, 267–273 10.1007/BF00656605 (doi:10.1007/BF00656605) [DOI] [Google Scholar]
- 15.Alerstam T. 1979. Wind as a selective agent in bird migration. Ornis Scand. 10, 76–93 10.2307/3676347 (doi:10.2307/3676347) [DOI] [Google Scholar]
- 16.Evans P. R. 1966. Migration an orientation of passerine night migrants in northeast England. J. Zool. Lond. 150, 319–369 10.1111/j.1469-7998.1966.tb03011.x (doi:10.1111/j.1469-7998.1966.tb03011.x) [DOI] [Google Scholar]
- 17.Nisbet I. C. T., Drury W. H., Jr 1967. Orientation of spring migrants studied by radar. Bird banding 38, 173–186 [Google Scholar]
- 18.Hake M., Kjellén N., Alerstam T. 2001. Satellite tracking of Swedish ospreys Pandion haliaetus: autumn migration routes and orientation. J. Avian Biol. 32, 47–56 10.1034/j.1600-048X.2001.320107.x (doi:10.1034/j.1600-048X.2001.320107.x) [DOI] [Google Scholar]
- 19.Alerstam T., Hake M., Kjellén N. 2006. Temporal and spatial patterns of repeated migratory journeys by ospreys. Anim. Behav. 71, 555–566 10.1016/j.anbehav.2005.05.016 (doi:10.1016/j.anbehav.2005.05.016) [DOI] [Google Scholar]
- 20.Strandberg R., Klaassen R. H. G., Hake M., Olofsson P., Alerstam T. 2008. Complex timing of marsh harrier Circus aeruginosus migration due to pre- and post-migratory movements. Ardea 96, 159–171 [Google Scholar]
- 21.Kjellén N., Hake M., Alerstam T. 2001. Timing and speed of migration in male, female and juvenile ospreys Pandion haliaetus between Sweden and Africa as revealed by field observations, radar and satellite tracking. J. Avian Biol. 32, 57–67 10.1034/j.1600-048X.2001.320108.x (doi:10.1034/j.1600-048X.2001.320108.x) [DOI] [Google Scholar]
- 22.Klaassen R. H. G., Strandberg R., Hake M., Alerstam T. 2008. Flexibility in daily travel routines causes regional variation in bird migration speed. Behav. Ecol. Sociobiol. 62, 1427–1432 10.1007/s00265-008-0572-x (doi:10.1007/s00265-008-0572-x) [DOI] [Google Scholar]
- 23.Trowbrigde C. C. 1902. The relation of wind to bird migration. Am. Nat. 36, 735–753 10.1086/278206 (doi:10.1086/278206) [DOI] [Google Scholar]
- 24.Alerstam T. 1979. Optimal use of wind by migrating birds: combined drift and overcompensation. J. Theor. Biol. 79, 341–353 10.1016/0022-5193(79)90351-5 (doi:10.1016/0022-5193(79)90351-5) [DOI] [PubMed] [Google Scholar]
- 25.Thorup K., Holland R. A. 2009. The bird GPS—long-range navigation in migrants. J. Exp. Biol. 212, 3597–3604 10.1242/jeb.021238 (doi:10.1242/jeb.021238) [DOI] [PubMed] [Google Scholar]
- 26.Reynolds A. M., Reynolds D. R., Smith A. D., Chapman J. W. 2010. A single wind-mediated mechanism explains high-altitude ‘non-goal oriented’ headings and layering of nocturnally-migrating insects. Proc. R. Soc. B 277, 765–772 10.1098/rspb.2009.1221 (doi:10.1098/rspb.2009.1221) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Karlsson H., Henningsson P., Bäckman J., Hedenström A., Alerstam T. 2010. Compensation for wind drift by migrating swifts. Anim. Behav. 80, 399–404 10.1016/j.anbehav.2010.05.023 (doi:10.1016/j.anbehav.2010.05.023) [DOI] [Google Scholar]
- 28.Liechti F., Bruderer B. 1998. The relevance of wind for optimal migration theory. J. Avian Biol. 29, 561–568 10.2307/3677176 (doi:10.2307/3677176) [DOI] [Google Scholar]
- 29.Erni B., Liechti F., Bruderer B. 2005. The role of wind in passerine autumn migration between Europe and Africa. Behav. Ecol. 16, 732–740 10.1093/beheco/ari046 (doi:10.1093/beheco/ari046) [DOI] [Google Scholar]
- 30.Shaffer S. A., et al. 2006. Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc. Natl Acad. Sci. USA 103, 12 799–12 802 10.1073/pnas.0603715103 (doi:10.1073/pnas.0603715103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Alerstam T. 1976. Bird migration in relation to wind and topography. PhD thesis, Lund University, Sweden [Google Scholar]
- 32.Alerstam T. 2006. Conflicting evidence about long-distance animal navigation. Science 313, 791–794 10.1126/science.1129048 (doi:10.1126/science.1129048) [DOI] [PubMed] [Google Scholar]