

Abstract
Gender-specific differences in longevity are reported across species and are mediated by mechanisms not entirely understood. In C57Bl/6 mice, commonly used in aging research, males typically outlive females. Since in these animals modest but prolonged reduction of core body (Tc) increased life span, we hypothesized that differential Tc may contribute to sex-specific longevity. Here, we compared the circadian profiles of Tc and locomotor activity (LMA) of male and female C57Bl/6 mice. Since Tc and LMA normally change with age, measurements were carried out in young (3 months) as well as in old (24 months) mice. In young females, Tc was influenced by estrous but was overall higher than in males. This difference was larger in old animals after age eliminated the variations associated with estrous. Although temperature homeostasis is regulated centrally by the sexually dimorphic hypothalamic preoptic area, these differences were uniquely dependent on the gonads. In fact, bilateral gonadectomy abolished the effects of estrous and increased resting Tc in males eliminating all sex-specific differences in Tc and LMA. These effects were only partially mimicked by hormonal replacement as Tc was affected by progesterone and to a lesser extent by estrogen but not by testosterone. Thus, gonadal-dependent modulation of Tc may be one of the physiological parameters contributing to gender-specific differences in longevity.
Electronic supplementary material
The online version of this article (doi:10.1007/s11357-010-9164-6) contains supplementary material, which is available to authorized users.
Keywords: Temperature, Gender, Mouse, Gonadectomy, Locomotor activity
Introduction
Temperature reduction prolongs lifespan and retards aging poikilotherms (Liu and Walford 1966; Conti et al. 2006). In homeotherms, core body temperature (Tc) correlates directly with calorie intake and inversely with longevity as Tc is reduced during dietary restriction and is elevated during feeding (Rikke and Johnson 2004, 2007; van Baak 2008). It was eventually shown that a modest (0.5°C) but prolonged reduction of Tc increased median life expectancy in mice independently of calorie restriction (Conti et al. 2006). This suggested that Tc might contribute to longevity and aging in homeotherms as an independent physiological variable and showed that over time, even small changes in Tc can have a significant impact on biological functions. Studies investigating the contribution of Tc to longevity in homeotherms remain few in number. Among the difficulties is the fact that Tc is subject to considerable variation (for a comprehensive review of temperature regulation in rodents see Gordon 1993). For instance, in mice, Tc follows a typical circadian profile with difference of up to 2°C between the light and the dark part of the day when the animals are resting or active, respectively. In addition, the circadian profile of Tc can be modulated differently by several environmental or physiological factors that must be taken into account, particularly when investigating modest differences. These include ambient temperature, humidity, food availability, locomotor activity (LMA), sex, as well as age that affects temperature stability to an extent that can reliably predict approaching death (Reynolds et al. 1985).
Differences in life span between males and females are reported across several species. In mice, the majority of the longevity and aging studies reported that males outlived females (Brown-Borg et al. 1996; Turturro et al. 1999; Coschigano et al. 2000, 2003; Flurkey et al. 2001, 2002; Holzenberger et al. 2003; Kurosu et al. 2005; Schriner et al. 2005; Ali et al. 2006; Conti et al. 2006; Selman et al. 2008; Ladiges et al. 2009). Differences between sexes were also reported in several genetic mouse models of longevity in which life span increase was typically larger in females (Brown-Borg et al. 1996; Flurkey et al. 2001, 2002; Coschigano et al. 2003; Holzenberger et al. 2003; Kurosu et al. 2005; Schriner et al. 2005; Conti et al. 2006; Selman et al. 2008; Ladiges et al. 2009). Several mechanisms have been proposed to explain these differences. They include asymmetric inheritance of sex chromosomes, maternal effects, exposure to different environmental factors, reduced free oxidative damage, and differences in physiology (Ali et al. 2006; Tower and Arbeitman 2009). Tc was never evaluated as a possible contributor.
Here, we investigated the effects of age on Tc and LMA in the C57Bl/6 mice commonly used in aging studies. Recording was carried out continuously by radiotelemety over several days in controlled constant environmental conditions on young adults (3 months old) and old (24 months old) male and female animals. The sex-specific differences were also investigated in mice that were subject to gonadectomy, sham operation, or hormonal replacement.
The findings describe how age and gender affect Tc and suggest that Tc may be one of the variables contributing to gender-specific differences in longevity. They also provide a reference for investigating the contribution of Tc to longevity and aging in mice.
Methods
Animals and surgeries
All procedures were approved by the IACUC of The Scripps Research Institute. Young (∼3 months old) and old (∼24 months old) C57Bl/6 female and male mice were used for all experiments. Animals were housed singly with “Aspen” Sani Chip Bedding (Harlan) and kept at 25°C, 30% humidity, and light/dark cycle of 12:12 h with lights on at 6:00 am defined as a zeitgeber time 0 (ZT0). Food (11% fat Harlan Teklad 7004) and water were ad libitum.
For all surgical procedures, animals were anesthetized with isoflurane (induction 4–5%, maintenance 1–1.5%) and were maintained on a warm heating pad during recovery and allowed to recover in a clean/no bedded cage.
For telemetry, radiotelemetric transmitters sterilized in quatracide (TA10TA-F20, Data Sciences, Inc, St Paul MN; size: approximately 20 × 10 × 7 mm, weight ∼3.4 g) were implanted in the peritoneum as previously described (Conti et al. 2006). Briefly, an incision was made along the abdominal midline, and the sterile transmitter was placed in the abdominal cavity. The anterior wall was sutured with absorbable coated vicryl 5–0, and the skin was sutured with vicryl rapide, a rapidly absorbed, synthetic, braided suture which elicits lower tissue reaction. The transmitters were kept off until the beginning of experiments. For recording, single-caged animals were positioned on single telemetric receivers (RPC-1, Data Sciences, Inc, St Paul, MN, USA). Animals were allowed to recover for 10 days before data were collected. The transmitters were activated 24 h prior to recording, and data were collected every 5 min.
Castration was performed as previously described (Carandente et al. 1982; Paul et al. 2006). Briefly, the anesthetized animal was placed in dorsal recumbency, and the scrotal area was carefully shaved. Prior to draping the site, sterile gauze was placed over the incision site. A small (approx. 1 cm) midline incision was made in the scrotum and subsequently through the tunica. The testicle was exteriorized and a clamp was placed across the spermatic vessels, which was ligated and cut across the top, enabling removal of the testicle. The ligated vessels were allowed to pass back through the tunica incision subsequently closed with one or two simple interrupted 5–0 sutures. The procedure was repeated for removal of the second testicle.
Ovariectomy was performed as previously described (Carandente et al. 1982; Paul et al. 2006). Briefly, the anesthetized animal was placed in ventral recumbency, and the mid-dorsal region was carefully shaved. A small (approx. 0.5 cm) lateral incision was made through the skin of the back of the mouse in the mid-lumbar region. The skin was slid from side to side to locate the position of the ovaries beneath the peritoneal wall. While holding the abdominal wall up, a small incision was made in the abdominal wall over the ovarian fat pad and the ovary exteriorized. The ligaments were carefully torn to release the ovary. A clamp was placed across the tip of the uterine horn, and a ligature was placed just underneath the tip of the uterine horn. The ovary/oviduct was removed by cutting above the ligature. The uterine horn was repositioned into the body cavity. The abdominal wall was closed with one or two simple interrupted sutures. This procedure was repeated for removal of the second ovary. The skin incision was closed with vicryl rapide suture.
For hormone replacement, commercially available sterile slow release pellets were purchased from Innovative Research of America (Sarasota, FL, USA): 17β-estradiol (0.5 mg/21 days), 5α-dihydrotestosterone (5 mg/21 days), and progesterone (10 mg/21 days). Pellets or their correspondent placebo were implanted subcutaneously as previously described (Conti 2000; Sugama et al. 2000). Briefly, an incision was made along the dorsal midline immediately posterior to the scapulae, just large enough to allow passage of the device. The sterile pellet was placed under the skin in the subcutaneous region. Absorbable sutures were used to close the dorsal incision, and the skin was closed with non-absorbable sutures. In order to minimize the incidence of infection, topical antibiotic treatments (Bacitracin Zinc Ointment) were applied after completion of surgical procedures.
Hormones measurement
Plasma estradiol, testosterone, and progesterone analysis were measured on plasma using 17-β-estradiol (IBL America IB79103), progesterone (IBL America IB79105), and testosterone (IBL America IB79106) ELISA kit, respectively, following the manufacturer’s instructions. Measurements were carried out in a single assay for each hormone. Inter-assay coefficient of variation was 1.58% for testosterone, 9.43% for progesterone, and 3.78% for 17-beta-estradiol.
Estrous cycle determination
Estrous cycle was determined by measuring the level of circulating hormones and by histological analysis of vaginal smear by lavage performed in the first 3 h of the dark cycle using a modified version of the procedure described by Goldman et al. (2007). Since telemetric recording is performed for 10 s every 5 min, to eliminate disturbance in the Tc profile (Supplemental Figure 1), samples were collected within 15 s immediately after the end of one recording point, allowing more than 4 min recovery time before the next telemetric recording. Briefly, the mouse was gently raised holding it by its tail and the vagina was flushed with 5 μl of sterile saline using a smooth and tapered micropipette tip. The possibility that the treatment was followed by induced pseudo-pregnancy induced by cervical stimulation was monitored by telemetry as it would mimic persistent diestrus. Collected samples were placed on a slide and were evaluated immediately, then fixed, dried, and stained for storage and subsequent examination using a standard laboratory light microscope with a ×100 total magnification. Cytology was used to define cycle stage as described by Dalal et al. (2001), and stage assignment was based on classification of cells and relative abundance of each cell type (percentage of parabasal, intermediate, cornified, and neutrophils). Proestrus is the presence of nucleated and cornified epithelial cells with some polymorphonuclear lymphocytes (PMNs) in the early stage (>25% basal epithelial cells, <10% neutrophils). Estrous is the cornified epithelial cells predominated with only few nucleated cells seen in the early stage (>50% superficial epithelial cells, zero neutrophils, and anucleated cornified cells). Metestrus is the cornified epithelial cells and PMNs predominated, with some nucleated epithelial cells. Diestrus is the cells that consisted primarily of PMNs with some epithelial cells (>50% neutrophils).
Data analysis
To determine the presence of statistically significant rhythmicity in the data set for each animal, the X2 periodogram procedure was used (Sokolove and Bushell 1978), and a numerical index of the robustness of a rhythm, QP, was obtained (Refinetti 2003) using a software program to calculate the circadian period (plus mean, amplitude, and acrophase) by the cosinor method provided by Dr. Roberto Refinetti. The QP statistic was calculated for each individual using the 5 days of data (1,440 data points). For data sets of this size, a perfect wave (such as a mathematically generated cosine wave) produces a QP value of 1,440. Three other parameters of the rhythms were analyzed: mean level, amplitude, and acrophase (i.e., the time of the daily peak). The mean level of the rhythms was computed as the arithmetic mean of the 288 daily measurements (at 0.083 h intervals) for each individual. The mean for each animal over the 5 days was then calculated. Likewise, the range of excursion was computed as the difference between the highest and the lowest temperature each day for each individual, and the mean for each animal was calculated. The acrophase of the rhythm was calculated by the method of the single cosinor for each day for each animal, and the mean for each animal was then computed. Statistical analysis was performed by t test. Additional statistical analysis involved comparison of group means by analysis of variance, post hoc pairwise comparisons by Tukey’s HSD test, and computation of correlation coefficients by the principle of the least squares (Refinetti 2003) and significances determined at P < 0.05.
Results
Young males vs young females
Comparative analysis of the recording of Tc and LMA in male and female C57Bl/6 mice over 5 days demonstrated the existence of sex-specific differences. Analysis was carried out on n = 6 per group. Since no difference between naive animals or animals subject to sham operation for the gonadectomy experiments was observed (not shown), data are reported for sham-operated animals. The average profile for each sex is presented in Fig. 1, and cosinor analysis is reported in Table 1.
Fig. 1.

Representative profiles of Tc and LMA in young animals over 5 days of continuous recording (n = 6 per group, SEM was not shown, *p < 0.05). a Tc and LMA of 3-month-old female mice (3 mo F); days of estrous phase are indicated; lines indicate level of resting Tc at day of estrous or during the other days of recording; reversal of the bimodal profile corresponding to elevated LMA and Tc at estrous are indicated with circles (see text for further details). b Tc and LMA of 3-month-old male mice (3 mo M). A line indicates that the value of resting Tc is constant over several days of recording. c Comparison of Tc and LMA of 3-month-old male and female mice. Statistically significant differences are indicated with stars
Table 1.
Cosinor analysis of circadian Tc and LMA profile of 3- and 24-month-old female and male mice following gonadectomy or sham operation
| 3 months | 3 months M | 24 months F | 24 months M | ||||
|---|---|---|---|---|---|---|---|
| Proestrus | Estrus | Metestrus | Diestrus | ||||
| Mesor | |||||||
| Tc | 36.83 (0.07) | 37.03 (0.06) | 36.85 (0.05) | 36.78 (0.06) | 36.51 (0.06) | 36.86 (0.09) | 36.46 (0.04) |
| Tc (GNDX) | 36.78 (0.04) | 36.88 (0.04) | 36.87(0.03) | 36.84 (0.04) | 36.89 (0.09) | ||
| LMA | 8.47 (0.90) | 8.26 (0.75) | 6.54 (0.37) | 7.59 (0.62) | 5.28 (0.22) | 3.98 (0.32) | 2.98 (0.51) |
| LMA (GNDX) | 5.26 (0.36) | 5.22 (0.45) | 5.23 (0.22) | 5.03 (0.21) | 5.46 (0.28) | ||
| Amplitude | |||||||
| Tc | 0.93 (0.04) | 0.95 (0.03) | 0.51 (0.03) | 0.70 (0.04) | 0.90 (0.02) | 0.48 (0.05) | 0.73 (0.44) |
| Tc (GNDX) | 0.56 (0.04) | 0.577 (0.02) | 0.57 (0.03) | 0.53 (0.03) | 0.58 (0.04) | ||
| LMA | 7.30 (0.80) | 8.28 (0.95) | 4.74 (0.34) | 5.97 (0.53) | 4.38 (0.28) | 1.58 (0.22) | 1.73 (0.46) |
| LMA (GNDX) | 3.71 (0.42) | 3.62 (0.47) | 3.69 (0.31) | 3.33 (0.28) | 3.74 (0.27) | ||
| Acrophase | |||||||
| Tc | 259.91 (5.39) | 254.61 (3.01) | 284.50 (7.44) | 268.25 (6.69) | 281.17 (4.71) | 280.34 (6.59) | 292.00 (4.13) |
| Tc (GNDX) | 251.40 (5.81) | 252.67 (3.77) | 248.60 (2.67) | 253.53 (4.14) | 284.53 (3.77) | ||
| LMA | 275.00 (5.23) | 282.15 (3.54) | 272.36 (3.93) | 277.60 (5.48) | 292.97 (6.84) | 211.67 (35.5) | 246.00 (33.01) |
| LMA (GNDX) | 267.22 (5.64) | 268.30 (5.33) | 267.75 (4.08) | 270.30 (3.75) | 292.76 (5.37) | ||
GNDX gonadectomy, F female, M male
For female mice, Tc and LMA showed a pattern that was affected by the estrous cycle (Fig. 1A, Table 1). Estrous cycle phases were determined by histological examination of vaginal pap smears (not shown) as well as circulating hormone levels (see below). We first determined the estrous day, then calculated retrogradely and anterogradely the days of proestrus, metestrus, and diestrus. Animals were not synchronized, and alignment of their Tc profiles was performed after individual vaginal and hormonal determination of each phase.
For male mice, Tc and LMA showed a consistent and reproducible pattern with a dark phase characterized by a biphasic profile of Tc and LMA (Fig. 1B, Table 1). Compared with males, female mice had (1) elevated Tc (0.2–0.5°C higher) and LMA (30% higher); (2) sustained LMA and Tc during the dark part at estrus, transforming the typical biphasic profile into monophasic; (3) higher LMA at proestrus and estrus.
Effects of age
Females
Comparison of Tc and LMA between 3- (n = 6) and 24-month (n = 6) females revealed that old females do not display the changes specific to each estrous phase observed in young animals but have a highly reproducible circadian profile, similar to that of ovariectomized young females (see below) (Fig. 2 A, A’). Histological and hormonal analysis confirmed that old females used in this study did not have estrous cycle (not shown). Like in old males, in females age was associated with a reduction of the rate of Tc increase at dark onset: old females increased their Tc from 36.6 ± 0.09°C to 37.7 ± 0.11°C in ∼4 h at a rate of 0.04°C/min, or 0.25°C/h, approximately four times the rate observed in young females with maximal Tc reached with approximately 3 h (3 h and 20 min) delay. This was associated with a delay in the onset of LMA at dark and with an overall reduction of its value (approximately three orders of magnitude) compared with young animals. Average maximal temperature was however only marginally different between young (37.8 ± 0.1°C) and old (37.7 ± 0.1°C) female mice.
Fig. 2.

Effects of age on the profiles of Tc and LMA recorded over 5 days (A to C) or over 24 h (A’ to C’). (A, A’) Comparison of Tc and LMA in 3- and 24-month-old female mice. (B, B’) Comparison of Tc and LMA in 3- and 24-month-old male mice. (C, C’) Comparison of Tc and LMA in 24-month-old male and female mice (n = 6, 6; *p < 0.05). Statistically significant differences are indicated with stars
Older females also had increased Tc (about 0.5°C higher than young naive animals) during the first half of the light phase.
Males
Comparison of Tc and LMA between 3- (n = 6) and 24-month (n = 6) males revealed significant differences during the dark/active phase (Fig. 2, Table 1). In old mice, the phases of Tc increased after dark onset from 35.9 ± 0.06°C to 37.5 ± 0.11°C in ∼4 h at a rate of 0.06°C/min, or 0.37°C/h, approximately three times slower than in young mice. During the dark part of day, Tc in old mice maintained a biphasic profile, but the two maximal peaks of Tc at end of approximately 0.5°C are lower than in young individuals (p < 0.05). In addition, due to delayed onset and to the reduction in the rate of Tc, the first peak of Tc in old mice was delayed approximately 3 h (3 h and 15 min) compared with young animals. LMA during the first 2 h of dark onset increased only marginally from the resting phase in old mice (1.8 ± 0.1 counts/5 min) to 2.8 ± 0.5 counts/5 min and subsequently increased with a rate approximately four times slower than that of young animals to a maximum of 9.3 ± 1.5 counts/min, approximately half of the value reached by young mice. The second peak of activity was not delayed but was fourfold lower than that of young mice. Consequently, aging effectively influenced the circadian profile of Tc and LMA by reducing mesor, amplitude, and acrophase in LMA while reducing amplitude and increasing achrophase of Tc (Table 1). No significant differences in either Tc or LMA of young and old mice were recorded during the light/inactive part of day or during the transition phases of Tc reduction.
Old males vs old females
Comparison of 24-month-old female (n = 6) and male (n = 6) mice demonstrated that the two sexes maintain similar circadian profiles, but resting Tc was 0.6 ± 0.01°C higher than in males (p < 0.01) (Fig. 3 C, C’, Table 1). During the dark/active part of day, old males and females had similar Tc, except for the value of the second maximal peak that was 0.3°C higher in females. No statistically significant changes were observed in LMA throughout 24 h of recording.
Fig. 3.

Effects of gonadectomy (GNDX) on the profiles of Tc and LMA recorded over 5 days (A to C) or over 24 h (A’ to C’). (A and A’) effects of gonadectomy. The 24-h profile of sham females includes estrous-dependent variations. (B, B’) effects of castration. (C, C’) Comparison of Tc and LMA profile in males and females after bilateral gonadectomy (n = 6, 6; *p < 0.05). Statistically significant differences are indicated with stars
Effects of gonadectomy
Tc and LMA were investigated in male and female mice subject to gonadectomy or to sham operation (n = 5 to 7 per group) (Supplemental Figure 2, Supplemental Table 1)
Females
In females, bilateral ovariectomy (OVX) effectively reduced the serum level of progesterone (ovariectomized, 0.2 ± 161 ng/ml; sham proestrus, 2.5 ± 0.01 ng/ml; sham estrous 2.6 ± 1.1 ng/ml; sham metestrus, 12.7 ± 5.6 ng/ml; sham diestrus, 7.7 ± 4.4 ng/ml; F(4,20) = 1.61, TukeyHSD: p < 0.05) and 17-β estradiol (ovariectomized, 22.2 ± 8.8 ng/ml; sham proestrus, 2.01 ± 0.5 ng/ml; sham estrous, 4.2 ± 2.1 ng/ml; sham metestrus, 3.2 ± 1.4 ng/ml; sham diestrus, 0.01 ± 0.2 ng/ml; F(4,20) = 8.14 TukeyHSD: p < 0.05). OVX had the following effects on different phases (Fig. 3): (1) increased averaged Tc at rest on proestrus (OVX 36.31 ± 0.09°C vs sham 36.1 ± 0.14°C, t test p < 0.05), estrus (OVX 36.4 ± 0.09°C vs sham 36.3 ± 0.11°C, t test p < 0.05), and diestrus (OVX 36.3 ± 0.10°C vs sham 36.2 ± 0.13°C, t test p < 0.05) despite a reduced motor activity; (2) eliminated the monophasic Tc and LMA profile observed in the dark phase of estrus (Fig. 2); (3) reduced LMA during the dark phase of day (proestrus OVX 7.4 ± 1.7, sham 15.7 ± 3, p < 0.05; estrous OVX 7.7 ± 1.6, sham 14.6 ± 3.3, p < 0.05; metestrus OVX 7.4 ± 1.5, sham 10.6 ± 3.0, p < 0.05; diestrus OVX 7.2 ± 1.4, sham 12.3 ± 2.8, p < 0.05).
Males
Bilateral castration reduced the levels of testosterone compared in male mice (15.1 ± 1.4 pg/ml vs 9.8 ± 1.03, t test *p < 0.05 in sham-operated vs castrated, respectively) and resulted in 0.5–0.7°C elevation of Tc exclusively at rest. The amplitude of the LMA remained unchanged. These changes resulted in increased mesor and decreases in amplitude without change in acrophase (Table 1). Eventually, Tc and LMA profiles after gonadectomy did not differ substantially except in the value of highest Tc reached at dark onset that was 0.32°C higher in males (Fig. 3 C, C’).
Discussion
Comparison of Tc and LMA profiles across sexes is particularly difficult in young mice because in females they vary with days of estrous cycle (Weinert 1994; Weinert et al. 2004). This becomes evident when comparing the recordings of Tc of both sexes over several days. The two profiles partially overlap, indicating that proper comparative Tc measurement depends on time of day and on day of recording. However, Tc was overall higher in female mice. This difference, previously reported in young mice (Yang et al. 2007), was found here to be also present and increased in old animals at rest. This suggests that female mice may have a higher Tc than males throughout the adult lifetime, an observation that is important when considering the possible effects of reduced Tc on lifespan. However, it should be noted that previous colonic temperature measurements indicate that middle age C57Bl/6J mice may experience transient body temperature disturbances (Talan and Engel 1986), a variable that was not assessed in our study. Apart from this important difference on resting Tc, the other effects of age on LMA and Tc were similar in both sexes. Compared with young, old animals showed reduced LMA, delayed onset, and lowered rate of LMA and Tc increased at dark affecting primarily circadian rhythmicity, significantly reducing their amplitude. These findings were consistent with measurements in mice previously reported where it was demonstrate that the effects of age on Tc were primarily due to reduced LMA (Weinert 2010). However, they differ from what was recently reported in rats where it was reported that 24-month-old animals had elevated daytime Tc without differences in LMA compared with younger rats (Gordon 2008). In addition, although it was reported that circadian rhythm of old organisms is mostly phase-advanced (Weinert 2010), in this study, male and female mice initiate their LMA later compared with young individuals.
Temperature homeostasis is essentially maintained by regulating heat generation and heat dissipation. In homeotherms, including humans and mice, this vital property is regulated centrally, by specific brain regions and neurons (Morrison et al. 2008; Nakamura and Morrison 2008). These include the preoptic area (POA) of the hypothalamus, a region where a network comprising warm as well as cold sensitive and temperature insensitive neurons contribute to the regulation of temperature homeostasis (Hammel 1965; Hori et al. 1999; Griffin et al. 2001; Abe et al. 2003; Tabarean 2005; Boulant 2006). Since the POA is a sexually dimorphic region, we investigated whether Tc differences could still be observed following surgical castration. As expected, bilateral ovariectomy eliminated the estrous-associated variations in females, reducing LMA and Tc at dark and increasing Tc at rest. Resting Tc was also elevated in males after bilateral castration to a level equal to that of ovariectomized females. As a result, gender-specific differences in Tc and LMA were virtually eliminated, and the only statistically significant difference remained in the maximal Tc value that reached at dark. To the best of our knowledge, this is the first demonstration that bilateral ovariectomy eliminated sex-specific differences in Tc and LMA. The findings show that temperature homeostasis is modulated centrally by similar mechanisms in both sexes and that sexual dimorphism of POA does not contribute to gender-specific differences in Tc in the absence of gonads. It also demonstrates that gonads can exert a reversible rather than a permanent influence on Tc, and suggests that age-related changes in gonadal functions can influence temperature homeostasis in a fashion that can be different in males and females. In fact, the increase of Tc difference between sexes observed in old mice can be attributed to the fact that age mimicked the effects of gonadectomy in female mice but not in males.
Sex hormones were proposed to influence Tc by direct action on the POA neurons (Crews et al. 1996; Coomber et al. 1997; Kruijver and Swaab 2002). Receptors for progesterone, testosterone, and estrogens were all found expressed in POA, where their ligands act preferentially, but not exclusively, on temperature-sensitive neurons (Silva and Boulant 1986; Tsai et al. 1988, 1992). However, only the effects of progesterone on basal Tc are believed to be centrally mediated, whereas estrogen and testosterone are proposed to participate in thermoregulatory response to changes in environmental temperature (Brown et al. 1970; Marrone et al. 1976; Fregly et al. 1979; Gangwar 1982). This is consistent with our findings that only progesterone, and to a much lesser extent 17β-estradiol, affected Tc. Instead, the effects of castration on resting Tc could not be eliminated with testosterone, suggesting the possibility that these may be modulated by other not yet identified testicular hormones.
Studies on the effects of gonads on longevity and aging found that castration was associated with a reduction of median life span of male rats (Drori and Folman 1986). In female mice, ovariectomy slightly reduced mean life span, and transplantation of young ovaries partially restored reproductive cycling and prolonged longevity in both post-pubertally gonadectomized, as well as in sham-operated animals (Mobbs et al. 1984; Cargill et al. 2003; Mason et al. 2009). These changes were not ascribed to changes in Tc, but were proposed to be due to the cardiovascular effects of estrogen and testosterone (Nieschlag et al. 1993; Silberberg and Magleby 1999), or by the estrogen dependent reduction of free radicals production (Vina et al. 2005a, 2005b; Ali et al. 2006). We and others showed that male C57Bl/6 mice typically outlive females (Brown-Borg et al. 1996; Turturro et al. 1999; Coschigano et al. 2000, 2003; Flurkey et al. 2001; Flurkey et al. 2002; Holzenberger et al. 2003; Kurosu et al. 2005; Schriner et al. 2005; Ali et al. 2006; Conti et al. 2006; Selman et al. 2008; Ladiges et al. 2009). In our previous work with C57Bl/6 mice housed in the exact same conditions of ambient temperature, humidity, light/dark cycle, and diet used in this study, females had ∼20% lower median life expectancy than males (Conti et al. 2006) (Supplemental Figure 3). Here, we found an inverse correlation between Tc and longevity across genders. We also showed that sex-specific differences in Tc are dependent on gonads and are regulated differently by age in male and female mice. This association suggests that gonadal-dependent modulation of Tc may be one of the physiological parameters contributing to gender-specific differences in longevity.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Table 1 Cosinor analysis of circadian Tc and LMA profile female and male mice during hormone replacement. DHT, 5α-dihydrotestosterone; 17beta E, 17β-estradiol; TX, treated. (PDF 49 kb)
{kind=link}
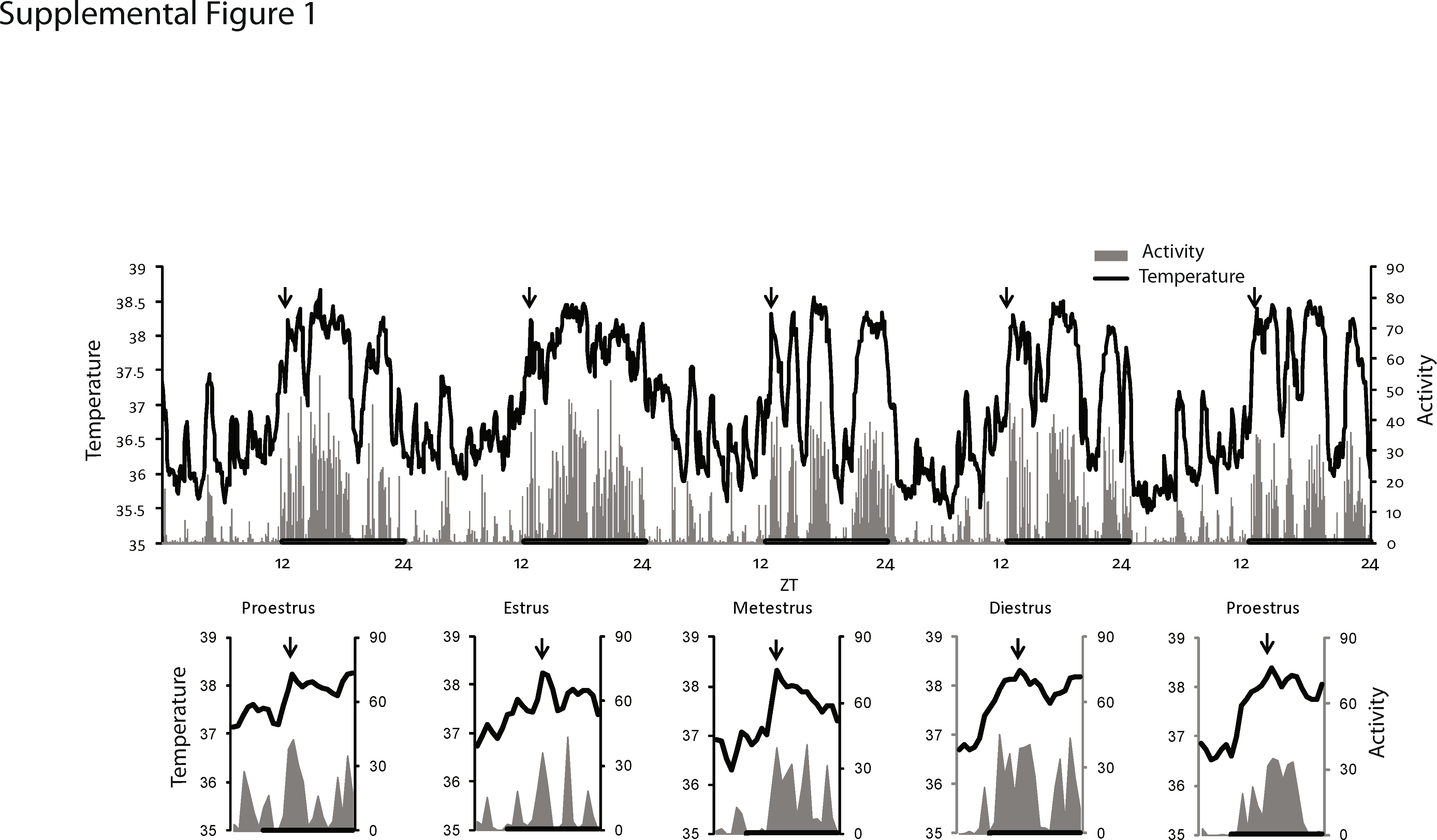
Supplemental Figure 1 Profile of Tc and LMA of 3-month-old female mice indicating (arrows) the time of vaginal swabs collected for histological analysis for determination of estrous phases. A profile comprising 1 h before and 1 h after sample collection is shown in the lower panel. (GIF 171 kb)
{kind=link}
Supplemental Figure 2 Tc and LMA profiles of castrated male and ovariectomized female treated with placebo or 5α-dihydrotestosterone (5αHDT) testosterone, progesterone, or 17β-estradiol (17βE) recorded over 24 h. (n = 6, 6; *p < 0.05). (GIF 311 kb)
{kind=link}
Supplemental Figure 3 Survival curves of C57Bl/6 mice maintained in the same dietary and environmental conditions used for the mice used in the present study, showing that males outlived females. Vertical lines correspond to the age investigated in this study: 3 and 24 months. The curves were extrapolated from the study previously published by us (Conti et al. 2006). (EPS 781 kb) (GIF 37 kb)
Acknowledgments
Supported by The Ellison Medical Foundation and NIH AG028040.
References
- Abe J, Okazawa M, et al. Primary cold-sensitive neurons in acutely dissociated cells of rat hypothalamus. Neurosci Lett. 2003;342(1–2):29–32. doi: 10.1016/S0304-3940(03)00239-8. [DOI] [PubMed] [Google Scholar]
- Ali SS, Xiong C, et al. Gender differences in free radical homeostasis during aging: shorter-lived female C57BL6 mice have increased oxidative stress. Aging Cell. 2006;5(6):565–574. doi: 10.1111/j.1474-9726.2006.00252.x. [DOI] [PubMed] [Google Scholar]
- Boulant JA. Neuronal basis of Hammel's model for set-point thermoregulation. J Appl Physiol. 2006;100(4):1347–1354. doi: 10.1152/japplphysiol.01064.2005. [DOI] [PubMed] [Google Scholar]
- Brown RE, Gander GW, et al. Estrogen and cortisone: effects on thermoregulation in the female rabbit. Proc Soc Exp Biol Med. 1970;134(1):83–86. doi: 10.3181/00379727-134-34733. [DOI] [PubMed] [Google Scholar]
- Brown-Borg HM, Borg KE, et al. Dwarf mice and the ageing process. Nature. 1996;384(6604):33. doi: 10.1038/384033a0. [DOI] [PubMed] [Google Scholar]
- Carandente F, Matteis MA, et al. Circadian temperature rhythm in intact, sham operated, gonadectomized rats. Chronobiologia. 1982;9(2):211–221. [PubMed] [Google Scholar]
- Cargill SL, Carey JR, et al. Age of ovary determines remaining life expectancy in old ovariectomized mice. Aging Cell. 2003;2(3):185–190. doi: 10.1046/j.1474-9728.2003.00049.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conti B, Sugama S, et al. Modulation of IL-18 production in the adrenal cortex following acute ACTH or chronic corticosterone treatment. Neuroimmunomodulation. 2000;8(1):1–7. doi: 10.1159/000026445. [DOI] [PubMed] [Google Scholar]
- Conti B, Sanchez-Alavez M, et al. Transgenic mice with a reduced core body temperature have an increased life span. Science. 2006;314(5800):825–828. doi: 10.1126/science.1132191. [DOI] [PubMed] [Google Scholar]
- Coomber P, Crews D, et al. Independent effects of incubation temperature and gonadal sex on the volume and metabolic capacity of brain nuclei in the leopard gecko (Eublepharis macularius), a lizard with temperature-dependent sex determination. J Comp Neurol. 1997;380(3):409–421. doi: 10.1002/(SICI)1096-9861(19970414)380:3<409::AID-CNE9>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- Coschigano KT, Clemmons D, et al. Assessment of growth parameters and life span of GHR/BP gene-disrupted mice. Endocrinology. 2000;141(7):2608–2613. doi: 10.1210/en.141.7.2608. [DOI] [PubMed] [Google Scholar]
- Coschigano KT, Holland AN, et al. Deletion, but not antagonism, of the mouse growth hormone receptor results in severely decreased body weights, insulin, and insulin-like growth factor I levels and increased life span. Endocrinology. 2003;144(9):3799–3810. doi: 10.1210/en.2003-0374. [DOI] [PubMed] [Google Scholar]
- Crews D, Coomber P, et al. Brain organization in a reptile lacking sex chromosomes: effects of gonadectomy and exogenous testosterone. Horm Behav. 1996;30(4):474–486. doi: 10.1006/hbeh.1996.0051. [DOI] [PubMed] [Google Scholar]
- Dalal SJ, Estep JS, et al. Standardization of the Whitten effect to induce susceptibility to Neisseria gonorrhoeae in female mice. Contemp Top Lab Anim Sci. 2001;40(2):13–17. [PubMed] [Google Scholar]
- Drori D, Folman Y. Interactive environmental and genetic effects on longevity in the male rat: litter size, exercise, electric shocks and castration. Exp Aging Res. 1986;12(2):59–64. doi: 10.1080/03610738608259437. [DOI] [PubMed] [Google Scholar]
- Flurkey K, Papaconstantinou J, et al. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proc Natl Acad Sci USA. 2001;98(12):6736–6741. doi: 10.1073/pnas.111158898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flurkey K, Papaconstantinou J, et al. The Snell dwarf mutation Pit1(dw) can increase life span in mice. Mech Ageing Dev. 2002;123(2–3):121–130. doi: 10.1016/S0047-6374(01)00339-6. [DOI] [PubMed] [Google Scholar]
- Fregly MJ, Kelleher DL, et al. Tolerance of estrogen-treated rats to acute cold exposure. J Appl Physiol. 1979;47(1):59–66. doi: 10.1152/jappl.1979.47.1.59. [DOI] [PubMed] [Google Scholar]
- Gangwar PC. Effects of climate on the thermoregulatory responses of male buffalo (Bubalus-bubalis) Calves supplemented with different levels of testosterone and TDN. Int J Biometeorol. 1982;26(1):73–79. doi: 10.1007/BF02187620. [DOI] [PubMed] [Google Scholar]
- Goldman JM, Murr AS, et al. The rodent estrous cycle: characterization of vaginal cytology and its utility in toxicological studies. Birth Defects Res B Dev Reprod Toxicol. 2007;80(2):84–97. doi: 10.1002/bdrb.20106. [DOI] [PubMed] [Google Scholar]
- Gordon CJ. Temperature regulation in laboratory rodents. New York: Cambridge University Press; 1993. [Google Scholar]
- Gordon CJ. Cardiac and thermal homeostasis in the aging Brown Norway rat. J Gerontol A Biol Sci Med Sci. 2008;63(12):1307–1313. doi: 10.1093/gerona/63.12.1307. [DOI] [PubMed] [Google Scholar]
- Griffin JD, Saper CB, et al. Synaptic and morphological characteristics of temperature-sensitive and -insensitive rat hypothalamic neurones. J Physiol. 2001;537(Pt 2):521–535. doi: 10.1111/j.1469-7793.2001.00521.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammel H. Neurons and temperature regulation. In: Yamamoto WS, Brobeck JR, editors. Physiological controls and regulations. Philadelphia: Saunders; 1965. pp. 71–97. [Google Scholar]
- Holzenberger M, Dupont J, et al. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 2003;421(6919):182–187. doi: 10.1038/nature01298. [DOI] [PubMed] [Google Scholar]
- Hori A, Minato K, et al. Warming-activated channels of warm-sensitive neurons in rat hypothalamic slices. Neurosci Lett. 1999;275(2):93–96. doi: 10.1016/S0304-3940(99)00732-6. [DOI] [PubMed] [Google Scholar]
- Kruijver FP, Swaab DF. Sex hormone receptors are present in the human suprachiasmatic nucleus. Neuroendocrinology. 2002;75(5):296–305. doi: 10.1159/000057339. [DOI] [PubMed] [Google Scholar]
- Kurosu H, Yamamoto M, et al. Suppression of aging in mice by the hormone Klotho. Science. 2005;309(5742):1829–1833. doi: 10.1126/science.1112766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladiges W, Remmen H, et al. Lifespan extension in genetically modified mice. Aging Cell. 2009;8(4):346–352. doi: 10.1111/j.1474-9726.2009.00491.x. [DOI] [PubMed] [Google Scholar]
- Liu RK, Walford RL. Increased growth and life-span with lowered ambient temperature in the annual fish, Cynolebias adloffi. Nature. 1966;212:1277–1278. doi: 10.1038/2121277a0. [DOI] [Google Scholar]
- Marrone BL, Gentry RT, et al. Gonadal hormones and body temperature in rats: effects of estrous cycles, castration and steroid replacement. Physiol Behav. 1976;17(3):419–425. doi: 10.1016/0031-9384(76)90101-3. [DOI] [PubMed] [Google Scholar]
- Mason JB, Cargill SL, et al. Transplantation of young ovaries to old mice increased life span in transplant recipients. J Gerontol A Biol Sci Med Sci. 2009;64(12):1207–1211. doi: 10.1093/gerona/glp134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mobbs CV, Gee DM, et al. Reproductive senescence in female C57BL/6J mice: ovarian impairments and neuroendocrine impairments that are partially reversible and delayable by ovariectomy. Endocrinology. 1984;115(5):1653–1662. doi: 10.1210/endo-115-5-1653. [DOI] [PubMed] [Google Scholar]
- Morrison SF, Nakamura K, et al. Central control of thermogenesis in mammals. Exp Physiol. 2008;93(7):773–797. doi: 10.1113/expphysiol.2007.041848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Morrison SF. A thermosensory pathway that controls body temperature. Nat Neurosci. 2008;11(1):62–71. doi: 10.1038/nn2027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieschlag E, Nieschlag S, et al. Lifespan and testosterone. Nature. 1993;366(6452):215. doi: 10.1038/366215a0. [DOI] [PubMed] [Google Scholar]
- Paul KN, Dugovic C, et al. Diurnal sex differences in the sleep–wake cycle of mice are dependent on gonadal function. Sleep. 2006;29(9):1211–1223. doi: 10.1093/sleep/29.9.1211. [DOI] [PubMed] [Google Scholar]
- Refinetti R. Metabolic heat production, heat loss and the circadian rhythm of body temperature in the rat. Exp Physiol. 2003;88(3):423–429. doi: 10.1113/eph8802521. [DOI] [PubMed] [Google Scholar]
- Reynolds MA, Ingram DK, et al. Relationship of body temperature stability to mortality in aging mice. Mech Ageing Dev. 1985;30(2):143–152. doi: 10.1016/0047-6374(85)90003-X. [DOI] [PubMed] [Google Scholar]
- Rikke BA, Johnson TE. Lower body temperature as a potential mechanism of life extension in homeotherms. Exp Gerontol. 2004;39(6):927–930. doi: 10.1016/j.exger.2004.03.020. [DOI] [PubMed] [Google Scholar]
- Rikke BA, Johnson TE. Physiological genetics of dietary restriction: uncoupling the body temperature and body weight responses. Am J Physiol Regul Integr Comp Physiol. 2007;293(4):R1522–R1527. doi: 10.1152/ajpregu.00215.2007. [DOI] [PubMed] [Google Scholar]
- Schriner SE, Linford NJ, et al. Extension of murine life span by overexpression of catalase targeted to mitochondria. Science. 2005;308(5730):1909–1911. doi: 10.1126/science.1106653. [DOI] [PubMed] [Google Scholar]
- Selman C, Lingard S, et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2008;22(3):807–818. doi: 10.1096/fj.07-9261com. [DOI] [PubMed] [Google Scholar]
- Silberberg SD, Magleby KL. Beating the odds with Big K. Science. 1999;285(5435):1859–1860. doi: 10.1126/science.285.5435.1859. [DOI] [PubMed] [Google Scholar]
- Silva NL, Boulant JA. Effects of testosterone, estradiol, and temperature on neurons in preoptic tissue slices. Am J Physiol. 1986;250(4 Pt 2):R625–R632. doi: 10.1152/ajpregu.1986.250.4.R625. [DOI] [PubMed] [Google Scholar]
- Sokolove PG, Bushell WN. The chi square periodogram: its utility for analysis of circadian rhythms. J Theor Biol. 1978;72(1):131–160. doi: 10.1016/0022-5193(78)90022-X. [DOI] [PubMed] [Google Scholar]
- Sugama S, Kim Y, et al. Tissue-specific expression of rat IL-18 gene and response to adrenocorticotropic hormone treatment. J Immunol. 2000;165(11):6287–6292. doi: 10.4049/jimmunol.165.11.6287. [DOI] [PubMed] [Google Scholar]
- Tabarean IV, Conti B, et al. Electrophysiological properties and thermosensitivity of mouse preoptic and anterior hypothalamic neurons in culture. Neuroscience. 2005;135(2):433–449. doi: 10.1016/j.neuroscience.2005.06.053. [DOI] [PubMed] [Google Scholar]
- Talan MI, Engel BT. Temporal decrease of body temperature in middle-aged C57BL/6J mice. J Gerontol. 1986;41(1):8–12. doi: 10.1093/geronj/41.1.8. [DOI] [PubMed] [Google Scholar]
- Tower J, Arbeitman M. The genetics of gender and life span. J Biol. 2009;8(4):38. doi: 10.1186/jbiol141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai CL, Matsumura K, et al. Effects of progesterone on thermosensitive neurons in preoptic slice preparations. Neurosci Lett. 1988;86(1):56–60. doi: 10.1016/0304-3940(88)90182-6. [DOI] [PubMed] [Google Scholar]
- Tsai CL, Kanosue K, et al. Effects of estradiol treatment on responses of rat preoptic warm sensitive neurons to progesterone in vitro. Neurosci Lett. 1992;136(1):23–26. doi: 10.1016/0304-3940(92)90638-N. [DOI] [PubMed] [Google Scholar]
- Turturro A, Witt WW, et al. Growth curves and survival characteristics of the animals used in the biomarkers of aging program. J Gerontol A Biol Sci Med Sci. 1999;54(11):B492–B501. doi: 10.1093/gerona/54.11.b492. [DOI] [PubMed] [Google Scholar]
- Baak MA. Meal-induced activation of the sympathetic nervous system and its cardiovascular and thermogenic effects in man. Physiol Behav. 2008;94(2):178–186. doi: 10.1016/j.physbeh.2007.12.020. [DOI] [PubMed] [Google Scholar]
- Vina J, Borras C, et al. Why females live longer than males: control of longevity by sex hormones. Sci Aging Knowledge Environ. 2005;23:pe17. doi: 10.1126/sageke.2005.23.pe17. [DOI] [PubMed] [Google Scholar]
- Vina J, Borras C, et al. Why females live longer than males? Importance of the upregulation of longevity-associated genes by oestrogenic compounds. FEBS Lett. 2005;579(12):2541–2545. doi: 10.1016/j.febslet.2005.03.090. [DOI] [PubMed] [Google Scholar]
- Weinert D. Lower variability in female as compared to male laboratory mice: investigations on circadian rhythms. J Exp Anim Sci. 1994;37:121–137. [Google Scholar]
- Weinert D. Circadian temperature variation and ageing. Ageing Res Rev. 2010;9(1):51–60. doi: 10.1016/j.arr.2009.07.003. [DOI] [PubMed] [Google Scholar]
- Weinert D, Waterhouse J, et al. Changes of body temperature and thermoregulation in the course of the ovarian cycle in laboratory mice. Biol Rhythm Res. 2004;35(3):171–185. doi: 10.1080/09291010412331335724. [DOI] [Google Scholar]
- Yang JN, Tiselius C, et al. Sex differences in mouse heart rate and body temperature and in their regulation by adenosine A1 receptors. Acta Physiol Oxf. 2007;190(1):63–75. doi: 10.1111/j.1365-201X.2007.01690.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Below is the link to the electronic supplementary material.
Supplemental Table 1 Cosinor analysis of circadian Tc and LMA profile female and male mice during hormone replacement. DHT, 5α-dihydrotestosterone; 17beta E, 17β-estradiol; TX, treated. (PDF 49 kb)
Supplemental Figure 1 Profile of Tc and LMA of 3-month-old female mice indicating (arrows) the time of vaginal swabs collected for histological analysis for determination of estrous phases. A profile comprising 1 h before and 1 h after sample collection is shown in the lower panel. (GIF 171 kb)
Supplemental Figure 2 Tc and LMA profiles of castrated male and ovariectomized female treated with placebo or 5α-dihydrotestosterone (5αHDT) testosterone, progesterone, or 17β-estradiol (17βE) recorded over 24 h. (n = 6, 6; *p < 0.05). (GIF 311 kb)
Supplemental Figure 3 Survival curves of C57Bl/6 mice maintained in the same dietary and environmental conditions used for the mice used in the present study, showing that males outlived females. Vertical lines correspond to the age investigated in this study: 3 and 24 months. The curves were extrapolated from the study previously published by us (Conti et al. 2006). (EPS 781 kb) (GIF 37 kb)