

Abstract
Elevated interleukin-6 (IL-6), a major mediator of the inflammatory response, has been implicated in androgen receptor (AR) activation, cellular growth and differentiation, plays important roles in the development and progression of prostate cancer, and is a potential target in cancer therapy. Through drug screening using human prostate cancer cells expressing IL-6 autocrine loop, we found that andrographolide, a diterpenoid lactone isolated from a traditional Chinese and Indian medicinal plant Andrographis paniculata, could inhibit IL-6 expression and suppress IL-6–mediated signals. Andrographolide inhibits IL-6 expression at both mRNA and protein levels in a dose-dependent manner. Andrographolide suppresses both IL-6 autocrine loop– and paracrine loop–induced cell signaling including Stat3 and Erk phosphorylation. Furthermore, andrographolide inhibits cell viability and induces apoptotic cell death in both androgen-stimulated and castration-resistant human prostate cancer cells without causing significant toxicity to normal immortalized prostate epithelial cells. Moreover, treatment of andrographolide to mice bearing castration-resistant DU145 human prostate tumors that express constitutive IL-6 autocrine loop significantly suppresses tumor growth. Taken together, these results demonstrate that andrographolide could be developed as a therapeutic agent to treat both androgen-stimulated and castration-resistant prostate cancer possibly by suppressing IL-6 expression and IL-6–induced signaling.
Keywords: prostate cancer, interleukin-6, andrographolide
Introduction
Interleukin-6 (IL-6) is a glycoprotein consisting of 212 amino acids encoded by the IL-6 gene localized to chromosome 7p21-14.1 IL-6 is a pleiotropic cytokine that plays a central role in host defense mechanisms by regulating immune responses and hematopoiesis and inducing either differentiation or growth of a variety of cells. The receptors for the IL-6 family of cytokines (IL-6, IL-11, ciliary neurotrophic factor, oncostatin M, and leukemia inhibitory factor) are composed of an IL-6–specific receptor subunit (α chain) and a signal transducer, gp130 (β chain). The binding of IL-6 to an α chain results in the formation of a hexameric complex containing 2 molecules of each component: IL-6, α chain, and gp130.2
IL-6 has been implicated in the modulation of growth and differentiation in many cancers and is associated with poor prognosis in renal cell carcinoma, ovarian cancer, lymphoma, melanoma, and prostate cancer.3 There is considerable evidence for the involvement of IL-6 in the development and progression of castration-resistant prostate cancer.4-6 The expression of IL-6 and its receptor has been consistently demonstrated in human prostate cancer cell lines and clinical specimens of prostate cancer and benign prostate hyperplasia.7-9 Multiple studies have demonstrated that IL-6 is elevated in the sera of patients with metastatic prostate cancer and that the levels of IL-6 correlate with tumor burden, serum PSA, and clinically evident metastases.10,11 In addition to the clinical data that IL-6 is associated with castration-resistant prostate cancer, experimental studies demonstrate that IL-6 plays a critical role in prostate cancer cell growth and differentiation. Okamoto et al. demonstrated that IL-6 functions as a paracrine growth factor for the human LNCaP androgen-sensitive prostate cancer cells and as an autocrine growth factor for the human DU145 and PC-3 androgen-insensitive prostate cancer cells.12 It has also been reported that IL-6 mediates LNCaP cell growth arrest and induces neuroendocrine differentiation.13-15 Results from a number of groups including ours demonstrate that IL-6 activates AR-mediated gene expression by activation of the AR through a Stat3 pathway in LNCaP cells.16-19 Overexpression of IL-6 enhanced PSA mRNA expression in LNCaP cells and can partially rescue LNCaP cells from growth arrest induced by androgen deprivation therapy. In addition, overexpression of IL-6 protects LNCaP cells from undergoing apoptosis induced by androgen deprivation therapy.20 An elegant study showed that osteoblasts promote prostate cancer cell growth through IL-6–mediated activation of the AR.21 Collectively, these findings suggest that IL-6 can regulate the expression of androgen-responsive genes and promote castration- resistant prostate cancer progression; targeting IL-6 signaling may present an attractive approach for prostate cancer therapy. Evidence suggests that targeting IL-6 signaling using an anti-IL-6 monoclonal antibody induces regression of human prostate cancer xenografts in nude mice.22 Most excitingly, inhibition of IL-6 with CNT0328, an anti-IL-6 monoclonal antibody, suppresses conversion of an androgen-dependent to an androgen-independent phenotype in a prostate cancer xenograft in vivo model.23
Andrographolide is a diterpenoid labdane that is the main bioactive component isolated from a traditional herbal medicinal plant Andrographis paniculata, which is effectively used for the treatment of infection, inflammation, cold, fever, and diarrhea in India and China.24 Numerous studies demonstrate that andrographolide exhibits anticancer activity by inducing apoptosis and cell cycle arrest.25 Andrographolide possesses strong anti-inflammatory activity mainly via inhibition of NF-κB signaling,26 suppression of the expression of inducible nitric oxide synthase,27 and the production of reactive oxygen species.28 Andrographolide can induce cell cycle arrest by increasing expression of p27 and decreasing expression of cyclin-dependent kinase 4.29,30 Andrographolide also triggers apoptosis via the caspase-8–dependent pathway in human cancer cells.31 In murine peritoneal macrophages, andrographolide inhibits the production of TNF-α and interleukin-12 via suppression of the ERK1/2 signaling pathway.32
In the present study, we found that andrographolide could inhibit IL-6 expression and suppress IL-6–mediated signals through drug screening using human prostate cancer cells expressing IL-6 autocrine loop. Andrographolide inhibits IL-6 production in a dose-dependent manner, suppresses IL-6–induced signaling including Stat3, Akt, and ERK1/2, and prevents prostate cancer cell growth in vitro and in vivo.
Results
Blocking IL-6 signaling inhibits prostate cancer cell growth
Previous studies suggest that IL-6 plays critical roles in the activation of androgen receptor and promoting prostate cancer cell growth.16,19,33 To examine the importance of IL-6 signaling in prostate cancer cell growth, IL-6 signaling was blocked in DU145 cells that constitutively express IL-6 autocrine loop using siRNAs specific to IL-6R and gp130. Knockdown of IL-6 signaling by either IL-6R siRNA or gp130 siRNA significantly suppressed DU145 cell growth (Fig. 1), consistent with the essential role of IL-6 in the growth of DU145 cells. These data suggest that IL-6 signaling is a logical target for prostate cancer, and blocking IL-6 expression may have therapeutic effects on prostate cancer.
Figure 1.
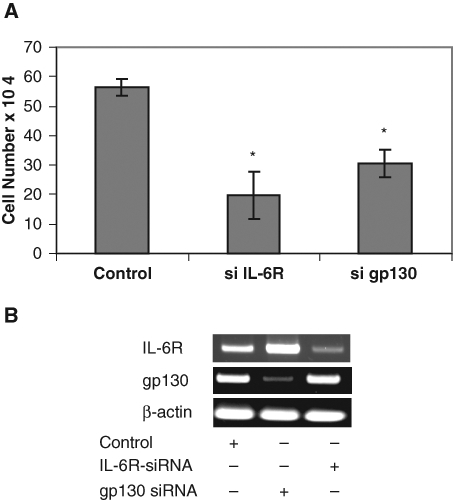
Knockdown of IL-6 signaling suppresses cell growth. (A) DU145 cells were transfected with siRNA specific to IL-6R and gp130 as indicated. GFP siRNA was used as a control. The cells were counted after 2 days following the transfection. (B) Expression of gp130 and IL-6R mRNA was determined by RT-PCR. Asterisks (*) indicate statistical significance.
Andrographolide inhibits IL-6 expression
In an attempt to identify potential inhibitors of IL-6, DU145 human prostate cancer cells that express IL-6 autocrine loop were treated with compounds from the Natural Compound Library (TimTec LLC, Newark, DE) for 24 hours. The Natural Compound Library consisted of 440 natural compounds with minimal purity of 95%. The media were collected and used for measurement of IL-6 protein by ELISA as described previously.33 Compounds that decreased the levels of IL-6 protein by 50% were selected for further characterization. One of the selected compounds is andrographolide (Fig. 2A). Andrographolide exhibits a wide spectrum of biological activities of therapeutic importance including antibacterial, anti-inflammatory, antimalarial, and anticancer properties and is used extensively in the traditional medicine of India and China.24
Figure 2.
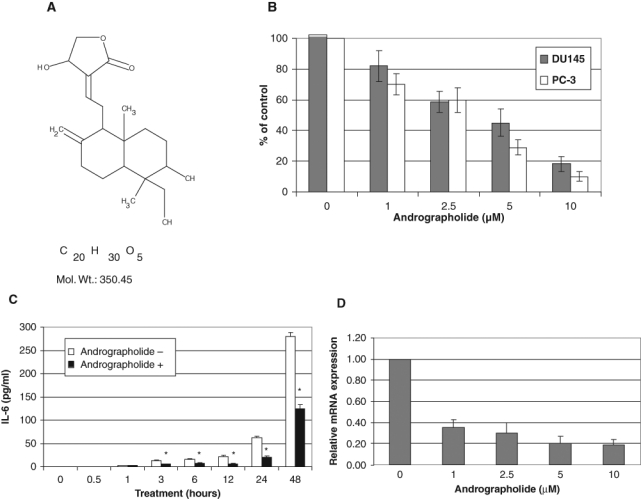
Andrographolide inhibits IL-6 expression. (A) Chemical structure of andrographolide. (B) Effect of andrographolide on IL-6 protein expression. DU145 and PC3 cells were treated with different doses of andrographolide for 24 hours, and the supernatants were used for detection of IL-6 protein expression by ELISA. The results were expressed as percentages compared to the vehicle control. (C) Time course effect of andrographolide on IL-6 protein expression. DU145 cells were cultured in 12-well plates. The media were changed when andrographolide (10 µM) was added to the cells. The media were collected at the time points indicated and used for IL-6 determination by ELISA. (D) Effect of andrographolide on IL-6 mRNA expression by qRT-PCR analysis. DU145 cells were treated with different concentrations of andrographolide as indicated. Total RNA was isolated and subjected to qRT-PCR analysis. Asterisks (*) indicate statistical significance.
We did a dose-response evaluation of andrographolide on IL-6 expression to confirm the initial screening results. Andrographolide inhibits IL-6 protein expression in a dose-dependent manner. The concentration of andrographolide required to decrease the levels of IL-6 protein in DU145 cells by 50% after 24 hours is about 3 µM, while 10 µM andrographolide reduces IL-6 protein level by almost 80% (Fig. 2B). A similar effect was observed in PC3 cells (Fig. 2B). The levels of IL-6 protein expression started to decrease after 3 hours upon andrographolide treatment and continued to decrease after 48 hours (Fig. 2C). Consistent with the protein levels, andrographolide inhibits IL-6 mRNA expression (Fig. 2D). Taken together, these results indicate that andrographolide inhibits IL-6 expression in prostate cancer cells.
Andrographolide antagonizes IL-6–induced signaling
IL-6 activates signaling including Stat3, Akt, and MAPK in prostate cancer cells. To determine whether andrographolide affects constitutive IL-6–induced signaling, DU145 cells that express constitutive IL-6 autocrine loop were treated with increasing doses of andrographolide, and cell lysates were collected and analyzed for protein expression. Andrographolide inhibits Stat3Tyr705 and Erk phosphorylation, whereas Akt phosphorylation was slightly inhibited (Fig. 3A). To determine whether andrographolide inhibits IL-6–stimulated signaling, LNCaP cells that lack IL-6 expression were pretreated with andrographolide for 4 hours, followed by 10 ng/mL of IL-6 treatment for 30 minutes. The cell lysates were collected and analyzed by Western blots. Andrographolide was able to block IL-6–stimulated Stat3Tyr705 phosphorylation, Erk phosphorylation, and Akt phosphorylation (Fig. 3B). These results indicate that andrographolide inhibits both constitutive IL-6 autocrine loop and paracrine IL-6–stimulated signaling.
Figure 3.
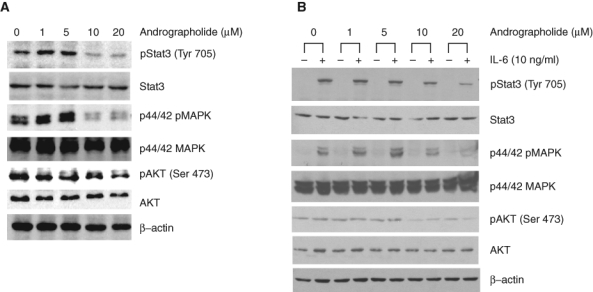
Andrographolide inhibits constitutive and IL-6–induced Stat3, MAPK, and AKT activity of human prostate cancer cells. (A) DU145 cells that express IL-6 autocrine loop were treated with increasing doses of andrographolide for 24 hours, and cell lysates were collected and subjected to Western blot analysis using the antibodies as indicated. (B) LNCaP cells that lack IL-6 but express IL-6 receptor and gp130 were pretreated with andrographolide for 4 hours. Cells were then stimulated with 10 ng/mL of IL-6 for 30 minutes after pretreatment with andrographolide. Whole cell lysates were prepared and subjected to Western blot analysis using antibodies as indicated.
Andrographolide inhibits cell viability and induces apoptosis
To determine whether andrographolide affects prostate cancer cell growth, human prostate cancer cell lines including DU145 and PC-3 that express IL-6 autocrine loop and normal immortalized prostate epithelial PzHPV-7 that lacks IL-6 expression were treated with increasing doses of andrographolide and cell viability determined by trypan blue assays. Andrographolide had little effect on cell viability up to 5 µM at the 48-hour treatment (Fig. 4A). However, andrographolide significantly inhibits DU145 and PC-3 cell growth at a concentration of 10 µM after 48 hours’ treatment (P < 0.001) (Fig. 4A). In contrast, andrographolide had little effect on the growth of the normal immortalized prostate epithelial PzHPV-7 cells up to 10 µM concentration (Fig. 4A). These data suggest that andrographolide may selectively inhibit the growth of prostate cancer cells. We next determined whether andrographolide-mediated growth inhibition is via induction of apoptosis. Apoptotic cell death was determined using the apoptosis-specific ELISA assay to evaluate DNA fragmentation as described previously.20 Andrographolide treatment at a concentration of 10 µM induces significant apoptosis in both DU145 and PC-3 prostate cancer cells but had little effect on PzHPV-7 cells (Fig. 4B). To determine the effects of andrographolide on cell invasion, DU145 cells were treated with different doses of andrographolide, and invasion was determined as the ability of cells to penetrate through Matrigel (BD Biosciences, San Jose, CA) in invasion assay as described previously.34 As shown in Figure 4C, andrographolide inhibited the invasive ability of DU145 cells in vitro. Collectively, our data suggest that andrographolide inhibits prostate cancer cell growth and invasion via induction of apoptosis.
Figure 4.
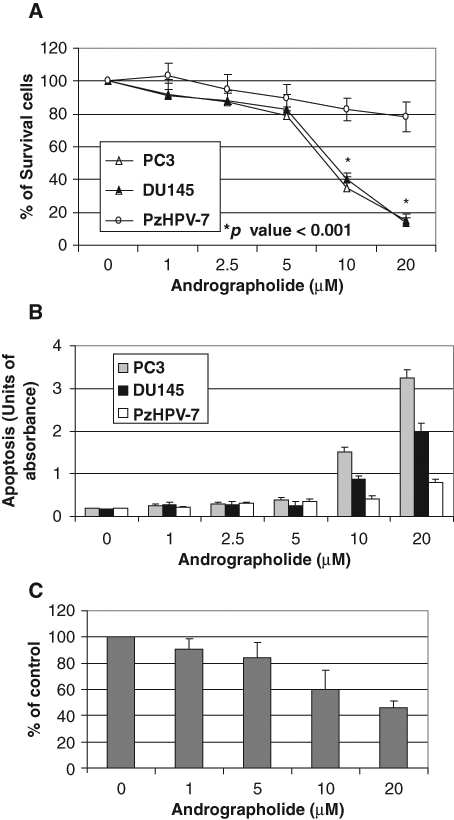
Andrographolide inhibits the growth of prostate cancer cells. (A) Andrographolide selectively inhibits the growth of DU145 and PC-3 prostate cancer cells over normal immortalized prostate epithelial cells. PC-3, DU145, or PZ-HPV7 cells were plated in 12-well plates and treated with various concentrations of andrographolide. After 48 hours, the cells were counted under a microscope. Results are expressed as the average percentage of live cells with standard deviation. Each bar is representative of 3 different experiments. Asterisks (*) indicate statistical significance. (B) Effect on apoptosis. Mononucleosomes and oligonucleosomes in the cytoplasmic fraction were analyzed by cell death detection ELISA as described in Materials and Methods. (C) Andrographolide inhibits DU145 penetration through Matrigel in invasion assay.
Andrographolide suppresses tumor growth
To explore the potential antitumor activity of andrographolide in vivo, we tested andrographolide on DU145 xenografts in male nude mice. Andrographolide (4 mg/kg body weight) was administered intraperitoneally 3 days after ectopic implantation of DU145 cells. Andrographolide treatment delayed tumor growth by 3 weeks and significantly (P < 0.05) reduced tumor volume throughout the experimental period (Fig. 5A). The andrographolide-treated mice gained weight similar to the control-treated mice and exhibited no obvious toxic effects (Fig. 5B).
Figure 5.
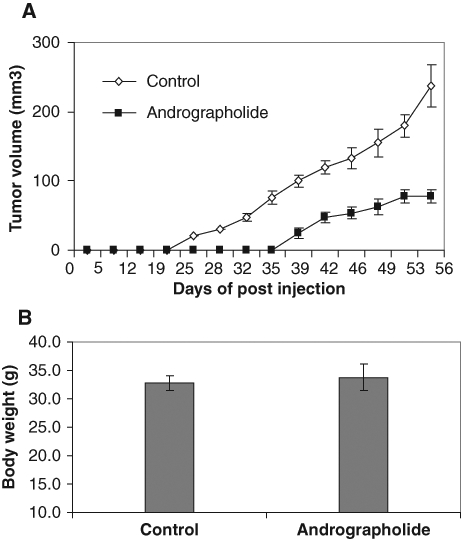
Andrographolide suppresses DU145 xenograft growth in nude mice. Male nude mice were injected subcutaneously with 1 × 106 cells/flank of DU145 cells. The mice were randomly divided into 2 groups with 10 mice each. One group received vehicle only as a control; another group received andrographolide (4 mg/kg of body weight). The treatment (intraperitoneal injection) was performed every other day starting 3 days post–cell inoculation for 8 weeks. (A) Average tumor volume from day 0 to day 56. The tumor volume was measured twice a week using calipers. (B) Average body weight after andrographolide treatment.
Discussion
Considerable evidence from both clinical and experimental studies demonstrated that IL-6 plays a vital role in promoting castration-resistant prostate cancer (CRPC) progression during androgen deprivation therapy. Evidence shows that 1) serum levels of IL-6 are elevated in men with advanced CRPC4-6,35; 2) overexpression of IL-6 enhances castration-resistant growth of the androgen-sensitive human LNCaP and LAPC-4 cells in vitro and in vivo33; 3) IL-6 enhances AR activity and stimulates androgen-responsive gene expression such as prostate-specific antigen (PSA) in prostate cancer (CaP) cells in the castrate environment36,37; 4) IL-6 protects CaP cells from apoptotic cell death and cell cycle arrest initiated by androgen deprivation38; 5) IL-6 increases expression of AR coregulators (p300 and SRC-1) and enhances interaction of AR with coregulators39,40; 6) IL-6 activates several major signaling pathways including JAK-STAT, PI3K-AKT, Src, Etk, β-catenin, erbB1-3, and MAPK in human CaP cells13,18,37,41-44; 7) IL-6 increases the expression of genes encoding many steroidogenic enzymes, including HSD3B2 and AKR1C1-3, which play a critical role in de novo androgen biosynthesis, suggesting that IL-6 may increase the levels of intraprostatic androgens45; 8) IL-6 supports autocrine and paracrine androgen-dependent cell survival in castrate conditions33; and 9) IL-6 increases LNCaP cell resistance to bicalutamide treatment mediated by the coactivator, TIF-2.46 In addition, IL-6 promotes proliferation of neuroendocrine (NE) cells and stimulates the production of neuroendocrine factors to support prostate epithelial cells surviving after androgen deprivation therapy. Collectively, these data suggest that IL-6 potentiates the progression to castration-resistant prostate cancer by sustaining prostate cancer cell survival in an autocrine and paracrine fashion and influencing hypersensitivity of AR and increasing intracrine de novo androgen biosynthesis and coregulator expression. These convincing data suggest that IL-6 signaling is an ideal target for prostate cancer. In fact, inhibition of IL-6 with CNTO328, an anti-IL-6 monoclonal antibody, suppresses conversion of androgen-dependent phenotype to a castration-resistant one in a prostate cancer xenograft in vivo model.23 These results are very encouraging because they indicate that inhibition of IL-6 and its signaling pathway could potentially yield antitumor effects. In this study, we identified andrographolide, a herbal medicine used in India and China, as a potent inhibitor of IL-6 that suppresses human prostate cancer cell growth in vitro and in vivo.
Our findings demonstrate that andrographolide inhibits IL-6 expression at both protein and mRNA levels in DU145 cells. A similar inhibitory effect was observed in PC-3 cells that express constitutive IL-6 autocrine loop, suggesting that inhibition of IL-6 expression by andrographolide is not DU145 cell specific. This phenomenon is consistent with a report showing that andrographolide derivatives inhibit the effects of LPS-induced TNF-α and IL-6 release in mouse macrophages.32 One of the known mechanisms of andrographolide-mediated suppression of inflammation is through inhibition of LPS-induced NF-κB activation in human umbilical vein endothelial cell (HUVEC) human adherent macrophages.26,47
Several signaling pathways have been reported to be crucial for IL-6–mediated AR activation and cellular functions in prostate cancer cells including Stat3, Akt, and MAPK. Among them, Stat3 is particularly involved in IL-6–mediated AR activation, neuroendocrine differentiation, proliferation, and survival upon androgen withdrawal. Our data show that andrographolide reduced not only the levels of Stat3Tyr705 in DU145 cells that express constitutive IL-6 autocrine loop but also the levels of IL-6–induced Stat3Tyr705 phosphorylation in LNCaP cells treated with exogenous IL-6. Furthermore, andrographolide reduces the levels of both constitutive and IL-6–induced p44/42 pMAPK, while slightly inhibiting the levels of pAKT. Similar observations have been previously described in other cell types. Andrographolide treatment inhibits v-Src–induced Erk1/2 phosphorylation and Akt(Ser473) phosphorylation in v-Src–transformed rat kidney epithelial PK3E cells.48
We have identified andrographolide as a potent IL-6 inhibitor that induces apoptosis and suppresses prostate cancer tumor growth. Andrographolide inhibits expression of IL-6 and blocks both autocrine and paracrine IL-6– mediated signaling including Stat3 and ERK1/2. Inhibition of IL-6 by andrographolide correlates with dose-dependent apoptosis induction and growth inhibition in DU145 cells. These data suggest that IL-6 signaling is a logical target for prostate cancer, and blocking IL-6 expression using andrographolide may have therapeutic effects on prostate cancer. In summary, our studies suggest that andrographolide, a herbal medicine used in China and India, may be developed as a potential therapeutic agent to treat prostate cancer.
Materials and Methods
Reagents and cell culture
LNCaP, DU145, and PC-3 cells were maintained in RPMI 1640 supplemented with 10% fetal bovine serum (FBS) and 100 U/mL penicillin and 0.1 mg/mL streptomycin (PS). PZ-HPV7 cells were cultured in keratinocyte-SFM medium supplemented with L- glutamine, EGF, and BPE. The cells were maintained at 37°C in a humidified incubator with 5% carbon dioxide. The Natural Compound Library consisting of 440 natural compounds was purchased from TimTec LLC (Newark, DE). Andrographolide was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in DMSO for in vitro and in vivo bioassays.
Compound screening
DU145 cells were seeded onto 96-well plates at 5,000 cells per well (200 µL) and allowed to grow for 24 hours. The cells were then treated with the compounds at a final concentration of approximately 5 µM and incubated at 37°C for another 24 hours. The media were collected and used for measurement of IL-6 protein by ELISA as described previously.33
Real-time PCR analysis
Total RNA was extracted using Trizol (Invitrogen, Carlsbad, CA) reagent. There was 1 µg of RNA digested with DNase and reverse transcribed with random primers and Im-Prom II Reverse transcriptase (Promega, Madison, WI). The cDNA was subjected to RT-PCR using specific primers for IL-6 (forward: 5′-AAGCCAGAGCTGTGCAGATGAGTA-3′, reverse: 5′-TGTCCTGCAGCCACTGGTTC-3′) and β-actin (forward: 5′-CCCAGCCATGTACGTTGCTA-3′, reverse: 5′-AGGGCATACCCCTCGTAGATG-3′). Quantitative real-time RT-PCR was performed in a 25 µL reaction mixture using SYBR Green IQ supermix (Bio-Rad Laboratories, Hercules, CA) according to the manufacturer’s instructions. Expression levels of IL-6 were normalized to β-actin. The experiment was repeated 3 times with duplicates.
Western blot analysis
Whole cell protein extracts were resolved on SDS-PAGE. Proteins were then transferred to nitrocellulose membrane. After blocking for 1 hour at room temperature in 5% milk in PBS/0.1% Tween-20, membranes were incubated overnight at 4°C with the indicated primary antibodies. Following secondary antibody incubation, immunoreactive proteins were visualized with an enhanced chemiluminescence detection system (Amersham Pharmacia Biotech, Little Chalfont, UK).
Cell growth assay
PC-3, DU145, or PZ-HPV7 cells were seeded on 12-well plates at a density of 1 × 105 cells/well in RPMI 1640 media containing 10% FBS. The cells were treated with increasing doses of andrographolide (0-20 µM), and live cells were counted using trypan blue exclusion after 24 hours. For transfection, DU145 cells were transiently transfected with siRNAs specific for IL-6 receptor or gp130 as described previously.45 The cells were counted after 2 days following the transfection.
Invasion assay
Invasion assays were done according to procedures described previously.34 Briefly, DU145 cells (1 × 105) were suspended in growth medium containing dimethyl sulfoxide or various concentrations of andrographolide into the cell culture inserts containing 8 µmol/L pores for 24-well plates. The inserts were coated with Matrigel (2-3 mg/mL protein) (BD Biosciences) and allowed to solidify before cell plating. The lower chambers were filled with 300 µL of growth medium containing control or drugs of corresponding concentrations. After 72 hours in culture, cells were fixed with 5% glutaraldehyde in PBS and stained with 0.5% toluidine blue in 2% Na2CO3. Only cells that penetrated the membrane were counted in 5 microscopic fields per filter.
Cell death ELISA
Cells were seeded on 12-well plates (1 × 105 cells/well) and treated with vehicle alone or different doses of andrographolide for 48 hours. Mononucleosomes and oligonucleosomes in the cytoplasmic fraction were measured at 405 nm using the Cell Death Detection ELISA kit (Roche, Basel, Switzerland) as described previously.20
In vivo tumor growth assay
DU145 prostate cancer cells were used for this experiment. Twenty male nude mice were randomly divided into 2 groups. The mice were inoculated subcutaneously in the right and left flanks with 1 × 106 viable cells. Three days after injection, one group of mice was treated with andrographolide at 4 mg/kg body weight in 1% DMSO; another group of mice was treated with 1% DMSO as vehicle control. Treatment was performed intraperitoneally every other day for 8 weeks. Body weights were measured before and after the treatment. Tumor volume measurements were taken twice per week after visible tumors appeared. Animals were sacrificed 8 weeks after initial injections, and a final measurement of tumor volume was taken. The volume of the growing tumors was estimated by measuring their dimensions (length × width × depth) with calipers. Mice were used in accordance with the approved Institutional Animal Care and Use Committee Protocol.
Statistical analysis
Data are shown as mean ± standard deviation. Multiple group comparison was performed by 1-way ANOVA followed by the Scheffe procedure for comparison of means. P < 0.05 was considered statistically significant.
Footnotes
This work was supported in part by grants from National Institutes of Health (NIH) CA118887, CA109441, and CA 140468, the National High Technology Joint Research Program of China (No.2006DAI02A02), and Hi-tech Research and Development Program of China Grant (No.2006AA02401).
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
- 1. Akira S, Taga T, Kishimoto T. Interleukin-6 in biology and medicine. Adv Immunol. 1993;54:1-78 [DOI] [PubMed] [Google Scholar]
- 2. Murakami M, Hibi M, Nakagawa N, et al. IL-6-induced homodimerization of gp130 and associated activation of a tyrosine kinase. Science. 1993;260(5115):1808-10 [DOI] [PubMed] [Google Scholar]
- 3. Simpson RJ, Hammacher A, Smith DK, Matthews JM, Ward LD. Interleukin-6: structure-function relationships. Protein Sci. 1997;6(5):929-55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Adler HL, McCurdy MA, Kattan MW, Timme TL, Scardino PT, Thompson TC. Elevated levels of circulating interleukin-6 and transforming growth factor-beta1 in patients with metastatic prostatic carcinoma. J Urol. 1999;161(1):182-7 [PubMed] [Google Scholar]
- 5. Drachenberg DE, Elgamal AA, Rowbotham R, Peterson M, Murphy GP. Circulating levels of interleukin-6 in patients with hormone refractory prostate cancer. Prostate. 1999;41(2):127-33 [DOI] [PubMed] [Google Scholar]
- 6. Nakashima J, Tachibana M, Horiguchi Y, et al. Serum interleukin 6 as a prognostic factor in patients with prostate cancer. Clin Cancer Res. 2000;6(7):2702-6 [PubMed] [Google Scholar]
- 7. Siegall CB, Schwab G, Nordan RP, FitzGerald DJ, Pastan I. Expression of the interleukin 6 receptor and interleukin 6 in prostate carcinoma cells. Cancer Res. 1990;50(24):7786-8 [PubMed] [Google Scholar]
- 8. Siegsmund MJ, Yamazaki H, Pastan I. Interleukin 6 receptor mRNA in prostate carcinomas and benign prostate hyperplasia. J Urol. 1994;151(5):1396-9 [DOI] [PubMed] [Google Scholar]
- 9. Hobisch A, Rogatsch H, Hittmair A, et al. Immunohistochemical localization of interleukin-6 and its receptor in benign, premalignant and malignant prostate tissue. J Pathol. 2000;191(3):239-44 [DOI] [PubMed] [Google Scholar]
- 10. Drachenberg DE, Elgamal AA, Rowbotham R, Peterson M, Murphy GP. Circulating levels of interleukin-6 in patients with hormone refractory prostate cancer. Prostate. 1999;41(2):127-33 [DOI] [PubMed] [Google Scholar]
- 11. Adler HL, McCurdy MA, Kattan MW, Timme TL, Scardino PT, Thompson TC. Elevated levels of circulating interleukin-6 and transforming growth factor-beta1 in patients with metastatic prostatic carcinoma. J Urol. 1999;161(1):182-7 [PubMed] [Google Scholar]
- 12. Okamoto M, Lee C, Oyasu R. Interleukin-6 as a paracrine and autocrine growth factor in human prostatic carcinoma cells in vitro. Cancer Res. 1997;57(1):141-6 [PubMed] [Google Scholar]
- 13. Qiu Y, Robinson D, Pretlow TG, Kung HJ. Etk/Bmx, a tyrosine kinase with a pleckstrin-homology domain, is an effector of phosphatidylinositol 3’-kinase and is involved in interleukin 6-induced neuroendocrine differentiation of prostate cancer cells. Proc Natl Acad Sci U S A. 1998;95(7):3644-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Spiotto MT, Chung TD. STAT3 mediates IL-6-induced neuroendocrine differentiation in prostate cancer cells. Prostate. 2000;42(3):186-95 [DOI] [PubMed] [Google Scholar]
- 15. Deeble PD, Murphy DJ, Parsons SJ, Cox ME. Interleukin-6- and cyclic AMP-mediated signaling potentiates neuroendocrine differentiation of LNCaP prostate tumor cells. Mol Cell Biol. 2001;21(24):8471-82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hobisch A, Eder IE, Putz T, et al. Interleukin-6 regulates prostate-specific protein expression in prostate carcinoma cells by activation of the androgen receptor. Cancer Res. 1998;58(20):4640-5 [PubMed] [Google Scholar]
- 17. Chen T, Wang LH, Farrar WL. Interleukin 6 activates androgen receptor-mediated gene expression through a signal transducer and activator of transcription 3-dependent pathway in LNCaP prostate cancer cells. Cancer Res. 2000;60(8):2132-5 [PubMed] [Google Scholar]
- 18. Lou W, Ni Z, Dyer K, Tweardy DJ, Gao AC. Interleukin-6 induces prostate cancer cell growth accompanied by activation of stat3 signaling pathway. Prostate. 2000;42(3):239-42 [DOI] [PubMed] [Google Scholar]
- 19. Lin DL, Whitney MC, Yao Z, Keller ET. Interleukin-6 induces androgen responsiveness in prostate cancer cells through up-regulation of androgen receptor expression. Clin Cancer Res. 2001;7(6):1773-81 [PubMed] [Google Scholar]
- 20. Lee SO, Lou W, Johnson CS, Trump DL, Gao AC. Interleukin-6 protects LNCaP cells from apoptosis induced by androgen deprivation through the Stat3 pathway. Prostate. 2004;60(3):178-86 [DOI] [PubMed] [Google Scholar]
- 21. Lu Y, Zhang J, Dai J, et al. Osteoblasts induce prostate cancer proliferation and PSA expression through interleukin-6-mediated activation of the androgen receptor. Clin Exp Metastasis. 2004;21(5):399-408 [DOI] [PubMed] [Google Scholar]
- 22. Smith PC, Keller ET. Anti-interleukin-6 monoclonal antibody induces regression of human prostate cancer xenografts in nude mice. Prostate. 2001;48(1):47-53 [DOI] [PubMed] [Google Scholar]
- 23. Wallner L, Dai J, Escara-Wilke J, et al. Inhibition of interleukin-6 with CNTO328, an anti-interleukin-6 monoclonal antibody, inhibits conversion of androgen-dependent prostate cancer to an androgen-independent phenotype in orchiectomized mice. Cancer Res. 2006;66(6):3087-95 [DOI] [PubMed] [Google Scholar]
- 24. Varma A, Padh H, Shrivastava N. Andrographolide: a new plant-derived antineoplastic entity on horizon. Evid Based Complement Alternat Med. 2009. September 14 [Epub ahead of print]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Geethangili M, Rao YK, Fang SH, Tzeng YM. Cytotoxic constituents from Andrographis paniculata induce cell cycle arrest in jurkat cells. Phytother Res. 2008;22(10):1336-41 [DOI] [PubMed] [Google Scholar]
- 26. Xia YF, Ye BQ, Li YD, et al. Andrographolide attenuates inflammation by inhibition of NF-kappa B activation through covalent modification of reduced cysteine 62 of p50. J Immunol. 2004;173(6):4207-17 [DOI] [PubMed] [Google Scholar]
- 27. Iruretagoyena MI, Tobar JA, Gonzalez PA, et al. Andrographolide interferes with T cell activation and reduces experimental autoimmune encephalomyelitis in the mouse. J Pharmacol Exp Ther. 2005;312(1):366-72 [DOI] [PubMed] [Google Scholar]
- 28. Shen YC, Chen CF, Chiou WF. Andrographolide prevents oxygen radical production by human neutrophils: possible mechanism(s) involved in its anti-inflammatory effect. Br J Pharmacol. 2002;135(2):399-406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Shi MD, Lin HH, Lee YC, Chao JK, Lin RA, Chen JH. Inhibition of cell-cycle progression in human colorectal carcinoma Lovo cells by andrographolide. Chem Biol Interact. 2008;174(3):201-10 [DOI] [PubMed] [Google Scholar]
- 30. Rajagopal S, Kumar RA, Deevi DS, Satyanarayana C, Rajagopalan R. Andrographolide, a potential cancer therapeutic agent isolated from Andrographis paniculata. J Exp Ther Oncol. 2003;3(3):147-58 [DOI] [PubMed] [Google Scholar]
- 31. Kim TG, Hwi KK, Hung CS. Morphological and biochemical changes of andrographolide-induced cell death in human prostatic adenocarcinoma PC-3 cells. In Vivo. 2005;19(3):551-7 [PubMed] [Google Scholar]
- 32. Li J, Huang W, Zhang H, Wang X, Zhou H. Synthesis of andrographolide derivatives and their TNF-alpha and IL-6 expression inhibitory activities. Bioorg Med Chem Lett. 2007;17(24):6891-4 [DOI] [PubMed] [Google Scholar]
- 33. Lee SO, Lou W, Hou M, de Miguel F, Gerber L, Gao AC. Interleukin-6 promotes androgen-independent growth in LNCaP human prostate cancer cells. Clin Cancer Res. 2003;9(1):370-6 [PubMed] [Google Scholar]
- 34. Yang JC, Bai L, Yap S, Gao AC, Kung HJ, Evans CP. Effect of the specific Src family kinase inhibitor saracatinib on osteolytic lesions using the PC-3 bone model. Mol Cancer Ther. 2010;9(6):1629-37 [DOI] [PubMed] [Google Scholar]
- 35. Twillie DA, Eisenberger MA, Carducci MA, Hseih WS, Kim WY, Simons JW. Interleukin-6: a candidate mediator of human prostate cancer morbidity. Urology. 1995;45(3):542-9 [DOI] [PubMed] [Google Scholar]
- 36. Culig Z, Bartsch G, Hobisch A. Interleukin-6 regulates androgen receptor activity and prostate cancer cell growth. Mol Cell Endocrinol. 2002;197(1-2):231-8 [DOI] [PubMed] [Google Scholar]
- 37. Liu S, Vinall RL, Tepper C, et al. Inappropriate activation of androgen receptor by relaxin via beta-catenin pathway. Oncogene. 2008;27(4):499-505 [DOI] [PubMed] [Google Scholar]
- 38. Lee SO, Lou W, Johnson CS, Trump DL, Gao AC. Interleukin-6 protects LNCaP cells from apoptosis induced by androgen deprivation through the Stat3 pathway. Prostate. 2004;60(3):178-86 [DOI] [PubMed] [Google Scholar]
- 39. Jia L, Choong CS, Ricciardelli C, Kim J, Tilley WD, Coetzee GA. Androgen receptor signaling: mechanism of interleukin-6 inhibition. Cancer Res. 2004;64(7):2619-26 [DOI] [PubMed] [Google Scholar]
- 40. Debes JD, Comuzzi B, Schmidt LJ, Dehm SM, Culig Z, Tindall DJ. p300 regulates androgen receptor-independent expression of prostate-specific antigen in prostate cancer cells treated chronically with interleukin-6. Cancer Res. 2005;65(13):5965-73 [DOI] [PubMed] [Google Scholar]
- 41. Vinall RL, Tepper CG, Shi XB, Xue LA, Gandour-Edwards R, de Vere White RW. The R273H p53 mutation can facilitate the androgen-independent growth of LNCaP by a mechanism that involves H2 relaxin and its cognate receptor LGR7. Oncogene. 2006;25(14):2082-93 [DOI] [PubMed] [Google Scholar]
- 42. Lee LF, Guan J, Qiu Y, Kung HJ. Neuropeptide-induced androgen independence in prostate cancer cells: roles of nonreceptor tyrosine kinases Etk/Bmx, Src, and focal adhesion kinase. Mol Cell Biol. 2001;21(24):8385-97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Qiu Y, Ravi L, Kung HJ. Requirement of ErbB2 for signalling by interleukin-6 in prostate carcinoma cells. Nature. 1998;393(6680):83-5 [DOI] [PubMed] [Google Scholar]
- 44. Nadiminty N, Lou W, Lee SO, Lin X, Trump DL, Gao AC. Stat3 activation of NF-{kappa}B p100 processing involves CBP/p300-mediated acetylation. Proc Natl Acad Sci U S A. 2006;103(19):7264-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Chun JY, Nadiminty N, Dutt S, et al. Interleukin-6 regulates androgen synthesis in prostate cancer cells. Clin Cancer Res. 2009;15(15):4815-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Feng S, Tang Q, Sun M, Chun JY, Evans CP, Gao AC. Interleukin-6 increases prostate cancer cells resistance to bicalutamide via TIF2. Mol Cancer Ther. 2009;8(3):665-71 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Li YD, Ye BQ, Zheng SX, et al. NF-kappaB transcription factor p50 critically regulates tissue factor in deep vein thrombosis. J Biol Chem. 2009;284(7):4473-83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Liang FP, Lin CH, Kuo CD, Chao HP, Fu SL. Suppression of v-Src transformation by andrographolide via degradation of the v-Src protein and attenuation of the Erk signaling pathway. J Biol Chem. 2008;283(8):5023-33 [DOI] [PubMed] [Google Scholar]