

Abstract
In nutritional epidemiology, it is often assumed that nutrient absorption is proportional to nutrient intake. For several nutrients, including non-haem Fe, this assumption may not hold. Depending on the nutrients ingested with non-haem Fe, its availability for absorption varies greatly. Therefore, using Fe intake to examine associations between Fe and health can impact upon the validity of findings. Previous algorithms that adjust Fe intakes for dietary factors known to affect absorption have been found to underestimate Fe absorption and, in the present study, perform poorly on independent dietary data. We have designed a new algorithm to adjust Fe intakes for the effects of ascorbic acid, meat, fish and poultry, phytate, polyphenols and Ca, incorporating not only absorption data from test meals but also current understanding of Fe absorption. In so doing, we have created a robust and universal Fe algorithm with potential for use in large cohorts. The algorithm described aims not to predict Fe absorption but available Fe in the gut, a measure we believe to be of greater use in epidemiological research. Available Fe is Fe available for absorption from the gastrointestinal tract, taking into account enhancing or inhibiting effects of dietary modifiers. Our algorithm successfully estimated average Fe availability in test meal data used to construct the algorithm and, unlike other algorithms tested, also provided plausible predictions when applied to independent dietary data. Future research is needed to evaluate the extent to which this algorithm is useful in epidemiological research to relate Fe to health outcomes.
Keywords: Iron, Availability, Algorithms, Absorption
Nutritional epidemiology is the study of dietary constituents such as nutrients, foods or food groups and their relationship to biomarkers of health and health outcomes. It is well recognised that the errors in quantifying nutrient intakes, which arise in many ways, can mask associations between diet and health. Accurate assessment and large cohorts are therefore required to detect the effects of diet on health. Critical to using intake data in nutritional epidemiology is the underlying assumption that nutrient intake has some agreement with nutrient exposure at the site of action or, in other words, that absorption shows some proportionality to intake. In the case of non-haem Fe, however, this assumption does not hold.
Non-haem Fe makes up about 90 % of total ingested Fe in an omnivorous Western diet(1) and, depending upon the nutrients with which it is ingested, its availability in the intestinal lumen varies markedly. For example, polyphenols, Ca and phytate, when consumed concurrently with Fe-containing foods, all inhibit Fe absorption, while ascorbic acid, red meat, fish and poultry enhance absorption. Thus, it is commonly recognised that the estimate of Fe intake is inappropriate to measure what is available for the body. For this reason it has been proposed that an algorithm, which takes account of the effects of some of the nutrients co-ingested with Fe, may go some way to restoring the association between ingestion and exposure by providing a value for available or absorbed Fe.
Algorithms may be criticised for generating values that generalise for the entire population and hence are imprecise for individuals. However, it is important to note that the purpose of such algorithms is only to allow Fe to be treated like many other nutrients in large studies and they cannot remove any of the other errors inherent in nutrient intake–health outcome studies. In addition, Conway et al. (2) have summarised further areas where Fe algorithms may be useful including: the identification of new dietary factors that modify Fe absorption, the setting of recommended daily amounts for populations based on the levels of available Fe expected in habitual diets(2) and institutional meal planning. It has been suggested that under certain circumstances, such as in inflammatory bowel disease, an accurate estimate of available Fe could help, alongside haematological parameters, to determine an individual’s Fe status(3).
In addition to dietary factors affecting the intestinal availability of Fe, the host’s Fe status also has a large impact on the extent of absorption through mucosal regulation. Some algorithms therefore include serum ferritin as a proxy for the host’s Fe status, and hence try to predict more accurately an individual’s absorption(4,5). An algorithm that incorporates Fe status, and therefore tries to predict Fe absorption, is unsuitable for epidemiological use because a measure of low absorption, for example, would not distinguish between a subject with a diet low in available Fe and a subject consuming a consistently Fe-rich diet who is therefore Fe replete and has homeostatically low absorption. In contrast, an algorithm that predicts Fe available in the gut rather than Fe absorption distinguishes effectively between the quality of subjects’ diets in terms of their typical exposure to Fe and therefore creates a measure that is useful in epidemiological research. Thankachan et al. (6) recently showed that the effect of enhancers and inhibitors on Fe’s availability is irrespective of a subject’s Fe status.
Most Fe algorithms are based on dietary constituents and have been reviewed by Beard et al. (7). A food group-based algorithm has also recently been published by Conway et al. (2). Overall, predictive abilities of all the algorithms were limited, as they significantly underestimate absorption(7). Fe absorption algorithms are often derived and evaluated on sample test meals that have been designed to have a single absorption-modifier in excess within meals of moderate and uniform portion size. In the present paper we take a new approach and have designed a non-linear equation derived not only from test meal data but also incorporating our knowledge of Fe absorption with the aim of developing an effective tool for dietary epidemiological research.
Experimental methods
Designing the algorithm
Subjects
The data generated from a study designed and carried out by Conway et al. (8) were used in designing the algorithm. In the Conway study, sixty-one subjects were recruited from students and staff at King’s College London. Inclusion criteria were: females aged 18–45 years; Hb < 125 g/l; serum ferritin < 20 μg/l; otherwise healthy; no history of gastrointestinal illness; taking no drugs that may affect Fe absorption; willingness to eat all proposed foods(8). The Conway study, described in detail elsewhere(8), was conducted according to the guidelines laid down in the Declaration of Helsinki and all procedures involving human subjects were approved by the King’s College Research Ethics Committee (7/797). Written informed consent was obtained from all subjects.
Dietary data
Each subject consumed between one and four of eighteen test meals. For each subject, meals were eaten on separate days within a 2-week period. The test meals were designed to have varying Fe availability. Table 1 shows the key dietary enhancers and inhibitors in each test meal. In seventeen of the eighteen meals 10 mg of exogenous iron chloride was added to ensure that Fe absorption could be detected (see below). Fe, Ca, phytate and polyphenol contents of the meals were analysed directly and ascorbic acid was estimated using values from McCance and Widdowson’s The Composition of Foods (9). The test meals used and further details of their composition are described elsewhere(8).
Table 1.
Dietary modifiers in test meals from a study by Conway et al. (8)
| Meal* | Subjects consuming the meal (n) |
Fe (mg) |
Ascorbic acid (mg) |
Animal tissue (g) |
Phytate (mg) |
Ca (mg) |
Polyphenols from tea (mg) |
|---|---|---|---|---|---|---|---|
| 1 | 40 | 13·1 | 260 | 0 | 0 | 188 | 0 |
| 2 | 20 | 14·9 | 129 | 0 | 108 | 172 | 0 |
| 3 | 10 | 16·7 | 87 | 0 | 161 | 176 | 0 |
| 4 | 10 | 16·7 | 87 | 0 | 161 | 176 | 227 |
| 5 | 10 | 11·6 | 82 | 0 | 0 | 85 | 0 |
| 6 | 10 | 11·8 | 84 | 0 | 0 | 460 | 0 |
| 7 | 10 | 11·9 | 85 | 0 | 0 | 98 | 0 |
| 8 | 10 | 12·1 | 87 | 0 | 0 | 473 | 0 |
| 9 | 10 | 14·9 | 129 | 0 | 108 | 481 | 0 |
| 10 | 19 | 13·4 | 129 | 0 | 0 | 171 | 0 |
| 11 | 10 | 13·4 | 129 | 0 | 0 | 480 | 0 |
| 12 | 9 | 12·8 | 44 | 0 | 0 | 205 | 0 |
| 13 | 9 | 12·8 | 44 | 0 | 0 | 205 | 227 |
| 14 | 10 | 13·4 | 8 | 0 | 0 | 171 | 227 |
| 15 | 10 | 11·3 | 8 | 0 | 0 | 72 | 0 |
| 16 | 10 | 11 | 8 | 100 | 0 | 119 | 0 |
| 17 | 11 | 11·3 | 85 | 100 | 0 | 132 | 0 |
| 18 | 10 | 3·1 | 260 | 0 | 0 | 188 | 0 |
In meals 1–17, 10 mg of exogenous iron chloride was added to ensure that Fe absorption could be detected.
Iron absorption measurements
Subjects fasted for 12 h before each test meal. Two baseline blood samples were taken before any food was consumed to determine mean serum Fe. Serum Fe data were measured at a further eight time points over 4 h postprandially and serum Fe curves derived. Fe absorption was calculated from the serum Fe curves obtained using the method described by Conway et al. (8) based on the two parameters of maximum increase in serum Fe concentration and percentage Fe recovery at peak and at fixed time points(8). The 4 h postprandial serum Fe curves were available for a total of 228 test meals. It is important to note that, in subjects who are Fe deficient, absorption will parallel Fe available in the gut because subjects will absorb all the Fe they can. In the present study we have used the assumption that Fe absorption equals available Fe in Fe-deficient subjects and thus used Fe-deficient subjects and absorption data (which can be measured) to generate the algorithm.
Form and estimation of parameters in the algorithm
The following key dietary factors that influence non-haem Fe absorption were selected for inclusion in this algorithm: ascorbic acid(10) and red meat, fish and poultry(10) as enhancers and Ca(11-13), phytate(14-17) and polyphenols(18,19) as inhibitors. Polyphenols from tea only were used to develop the algorithm. Tea is a major source of polyphenols in Western diets, and can be relatively easily calculated from dietary intake data.
The form of the algorithm as a whole (where A is a constant) was decided upon to ensure that predictions of available Fe could never fall below 0 %:
The next step was to select a function to describe the multiplicative relationship between each dietary variable and non-haem Fe availability in the gut. In contrast to previous work, we designed our algorithm based on absorption data from test meals and current biological understanding of Fe absorption. We used the function ‘y = 1 + ln (1 + bx)’ for each dietary enhancer and ‘y = 1/(1 + ln (1 + bx))’ for each inhibitor to describe the multiplicative effect of these factors on non-haem Fe availability in our algorithm (Appendix A Figs. 1 and 2). In the absence of each dietary factor (i.e. x = 0) there is no effect on availability (i.e. multiply by 1). As the intake of the dietary enhancer increases so too does non-haem Fe availability in the gut. The enhancing effect increases at a slower rate until the increased effect of the dietary enhancer on availability is negligible. With increasing intakes of inhibitors, availability falls at a decreasing rate until, as with dietary enhancers, continued increase in intake has minimal further effect on non-haem Fe availability. The functions selected define a multitude of curves, but precisely which curve is used is determined from the test meal data and non-linear regression. We included dietary non-haem Fe in our algorithm in the same manner as an inhibitor. It was felt important to include Fe in the algorithm, which is often not done (see Reddy et al. (4), Monsen et al. (10) and Tseng et al. (20)), because, as Fe intake increases, a proportionally smaller percentage of Fe is absorbed(21). Fe is, itself, an effective inhibitor of percentage Fe availability in the gut.
Once we had selected the dietary variables for inclusion in the algorithm, the form of the algorithm, and the functions to describe the relationships, we used non-linear multiple regression by least-squares analysis to model the combined influence of the selected dietary factors on non-haem Fe absorption and thus estimate the parameters in the algorithm. An additional equation estimates availability of haem Fe at an efficiency of 25 % based on current understanding of haem Fe absorption(22). This, added to availability of non-haem Fe estimated by our algorithm, predicts ‘available Fe’.
Applying the algorithm to independent data
Subjects
Data generated in a study by Cook(3) were used in applying and testing our newly developed algorithm on an independent set of dietary intake data to see if our algorithm at least generated plausible predictions of available Fe. Twenty-eight healthy, free-living control subjects recruited by advertisement in a London newspaper on two consecutive Tuesdays were used. Potential subjects had been screened by telephone to exclude those with known chronic disease, hereditary disorders of Fe metabolism or those taking proton pump inhibitor medication. Pregnant and lactating women were also excluded. Subjects fell within the age range of 18–65 years.
Dietary and iron status data
Meal-by-meal intake data were collected through a specific validated Fe FFQ(23) that was used to assess intakes of dietary Fe and Fe absorption modifiers over the previous month. Recorded intake levels of haem and non-haem Fe, ascorbic acid, red meat, fish and poultry, polyphenols from tea, phytate and Ca had been calculated from the FFQ and the data, yielding 314 separate meals, were used in the present study. Table 2 shows the range of intakes of the key dietary modifiers. Serum ferritin was measured for twenty-six of the subjects.
Table 2.
Intake range of dietary modifiers from a study by Cook et al. (3)
| Dietary variable | Intake range per meal |
|---|---|
| Non-haem Fe (mg) | 0–30 |
| Haem Fe (mg) | 0–10 |
| Ascorbic acid (mg) | 0–329 |
| Red meat, fish and poultry (g) | 0–350 |
| Polyphenols from tea (mg) | 0–740 |
| Phytate (mg) | 0–332 |
| Ca (mg) | 1–2759 |
| Tannic acid (mg)* | 0–111 |
Statistical analysis
We applied the newly developed algorithm and those of Reddy et al. (4) and Hallberg & Hulthén(5), to investigate predicted Fe availability and predicted absorption and to consider whether the algorithms were sufficiently flexible for use with every meal. The algorithms of Reddy et al. (4) and Hallberg & Hulthén(5) are detailed in Appendix B. The algorithms were applied to each meal separately. Non-haem Fe and haem Fe availabilities are predicted independently and then summed to give total available Fe for each meal.
All statistical analysis was performed using Stata software (version 10.0; StataCorp LP, College Station, TX, USA).
Results
Designing the algorithm
The individual distributions, of measured percentage Fe absorption, are shown in Appendix A Fig. 3 for the eighteen different meals. Fe absorption varied across different meals and also between subjects. ANOVA demonstrated that the maximum variation in Fe absorption that could be explained by an algorithm of dietary factors using these data was 47 % (i.e. R2 0·47). Intra-meal variation cannot be explained by dietary factors using these data, therefore 53 % of variation is assumed to come from between-subject differences as well as overall measurement error.
Having established, by a priori criteria, the constraints and form of the model, non-linear regression indicated that the following algorithm provided the closest fit to the data:
where:
and AA is ascorbic acid (mg), AT is red meat, fish and poultry (g), C is Ca (mg), P is phytate (mg), PO is polyphenols from tea (mg), NH is non-haem Fe (mg) and HI is haem Fe (mg).
The relationship between Fe absorption (from serum Fe curves) and predicted available Fe (from the algorithm above) is R2 0·45 (P<0·001; root mean squared error = 5·8). The algorithm explained much of the theoretical maximum for between-meal variation (i.e. R2 0·45 v. a maximum R2 0·47).
When comparing two meals in the Conway et al. study(8), which were identical with the exception of Fe content (meal 1 v. meal 18; Table 1), the meal with the higher Fe content had lower percentage Fe absorption, supporting the inclusion of dietary non-haem Fe as an inhibitor of percentage available Fe in this algorithm (Appendix A Fig. 3).
Applying the algorithm
For the algorithm developed here all meals from the study by Cook(3) could be modelled (n 314) and the range of predicted Fe availabilities was 5–35 % (Table 3). Predictions of available Fe (i.e. excluding adjustments for individuals’ serum ferritin from the algorithms), using the Reddy et al. algorithm(4) and the Hallberg & Hulthén algorithm(5), ranged from 12–1591 % and 3–355 % respectively (Table 3). Using the Reddy et al. algorithm(4) or the Hallberg & Hulthén algorithm(5) to estimate absorption, eighteen of the 314 meals (5·8 %) could not be predicted because of missing ferritin values for two subjects. Of the remaining 296 meals, the Reddy et al. algorithm(4) predicted Fe absorption exceeding 100 % in fifteen meals (range being 103–410 %) and the range of absorption was 1–97 % for the remaining meals. Similarly, some absorption estimates of greater than 100 % were observed when using the Hallberg & Hulthén algorithm(5). Twenty-six meals had a predicted absorption of over 100 %, while predictions ranged from 0 to 92 % for the remaining 270 meals (Table 3). The median predictions of available Fe were similar for our algorithm and that of Reddy et al. (4).
Table 3.
Comparison of algorithm predictions*
| Our algorithm | Reddy et al. algorithm(4) | Hallberg & Hulthén algorithm(5) | |
|---|---|---|---|
| Percentage Fe absorption† | |||
| Median | – | 15·1 | 21·3 |
| IQR | – | 5·4–42·6 | 5·5–43·5 |
| Range | – | 1·1–409·8 | 0·4–303·6 |
| N | – | 296 | 296 |
| Percentage available Fe‡ | |||
| Median | 19·3 | 24·9 | 47·7 |
| IQR | 16·4–21·6 | 21·7–69·2 | 23·0–77·4 |
| Range | 5·5–34·5 | 12·4–1590·7 | 2·7–355·2 |
| N | 314 | 314 | 314 |
IQR, interquartile range.
Predictions of available Fe and Fe absorption from the Reddy et al. (4)and Hallberg & Hulthén(5) algorithms as well as predictions of available Fe from the algorithm developed in the present study, on data from a study by Cook et al. (3).
The absence of a measure of serum ferritin in two subjects meant that Fe absorption could only be estimated for 296 meals when using the Reddy et al. (4) and Hallberg & Hulthén(5) algorithms which both incorporate ferritin.
Using the assumption that absorption in an Fe-deficient subject equals available Fe, percentage available Fe was calculated using the Hallberg & Hulthén algorithm(5) and the Reddy et al. algorithm(4) by calculating estimates for a hypothetical person with a serum ferritin of 10 μg/l, i.e. as if the subject consuming the meals were Fe deficient (see Appendix B).
Discussion
The discrepancy between dietary Fe intake and the amount of Fe available for use by the body, because of the variation in the proportion of non-haem Fe that can be absorbed, drives the need for an improved measure for use in epidemiological research when examining associations between Fe and health. Previously, algorithms have been developed to predict Fe absorption and this has been seen as advantageous to using intake data alone. However, current published algorithms significantly underestimate Fe absorption(7), have a lack of agreement with one another(7) and can predict impossibly high absorption percentages when used on dietary survey data (Table 3). Furthermore, we believe that predicting absorption is of limited use for population-based research and instead the focus should be on distinguishing between different qualities of diet based on the amount of Fe available in the intestinal lumen. With this in mind, we have developed a new algorithm constructed not solely on the mathematical basis of absorption from test meals but incorporating current biological understanding of how dietary variables affect Fe availability and we aim to predict not absorption, but Fe available in the gut. To predict Fe absorption from a single meal and where biomarkers are considered reliable for Fe status, then an algorithm including a measurement of Fe status, such as ferritin, would be appropriate. However, this is not the reason for which we designed our algorithm and the use of Fe status markers would be counter-productive.
Variation in absorption is dependent in part on dietary factors consumed simultaneously with Fe-containing foods (i.e. available Fe) but it also depends on individual variation. Intra-meal variation (i.e. when different individuals consume the same meal but absorb differing amounts of Fe) cannot be explained by an algorithm of dietary factors. Our algorithm is able to explain 45 % of the variation in this dataset. It is important to note that this R2 value is achieved on individual data. If, instead, individual variation is removed and mean Fe absorption values for each meal are used, as presented by Hallberg & Hulthén for their algorithm(5), then the R2 of our algorithm and that of Hallberg & Hulthén compare well (R2 0·93 and 0·99 respectively; data not shown).
For any algorithm to be of use in epidemiological research it needs to predict Fe availability at all intakes of dietary Fe and dietary modifiers of Fe availability, including values that are outside the range of dietary variables used to construct the algorithm. Because it describes the relationship between availability and dietary variables from biological understanding of Fe absorption, and not solely from test meal data, our algorithm seems applicable to many different studies. Key to this ability are the selected non-linear functions which, we believe, more accurately describe Fe availability compared with previously used functions. As with previous algorithms, an increase in intake of the dietary modifiers leads to an increase in the effect that they have on non-haem Fe availability. However, in our algorithm, as the intake of each dietary modifier increases, the effect on availability increases at a slower rate until, at high levels, the increase in effect of the dietary modifier on availability is minimal. Therefore, estimates of available Fe are maintained at biologically plausible levels, unlike the levels of absorption occasionally observed when using other algorithms(7). Using our algorithm, available Fe predictions exceeding or even approaching 100 % are very unlikely within normal dietary intake ranges. In fact we have simulated over 1·5 million permutations (i.e. meals) with varying composition (ascorbic acid 0–1000 mg; red meat, fish and poultry 0–1000 g; phytate 0–4000 mg; polyphenols from tea 0–2000 mg; Ca 0–2500 mg; non-haem Fe 17–100 mg; haem Fe 0–100 mg) and found that predictions of available Fe by our algorithm for all meals fell within the range of 1 to 67 % (data not shown).
Appendix A Fig. 4 shows the shape of the curve used by Reddy et al. (4) to describe the effect of animal tissue on non-haem Fe absorption in their algorithm. The sharp-rising, ever-increasing curve could result in unrealistic estimates of Fe absorption at very high levels of animal tissue intake. From 380 g of animal tissue upwards, absorption exceeds 100 %. Ascorbic acid is similarly defined, so estimates of absorption at high intakes of ascorbic acid are also likely to exceed biologically plausible levels. When these effects are combined, the effect on absorption is more dramatic, as reported by Beard et al. (7) who calculated predicted absorption to be greater than 100 % in some subjects when using the Reddy et al. algorithm(4). Functions selected for use in other algorithms also fail to support common understandings of Fe absorption, such as the functions selected by Hallberg & Hulthén(5) to define the effect of phytate in their algorithm (Appendix A Fig. 5). In the Hallberg & Hulthén algorithm, in the absence of ascorbic acid, phytate works as an inhibitor of non-haem Fe absorption, which supports conventional theory about the effect of phytate(14-17) (Appendix A Fig. 5). However, even as little as 0·01 mg of ascorbic acid changes this relationship dramatically. At very low levels of ascorbic acid, phytate no longer has an effect on non-haem Fe absorption. At intakes upward of 1 mg of ascorbic acid, phytate enhances Fe absorption (Appendix A Fig. 5). To give perspective, this means that in the Medical Research Council’s National Survey of Health and Development(24) phytate would increase predictions of Fe absorption in two-thirds of all meals consumed in 1999 when using Hallberg and Hulthén’s algorithm (data not shown). This does not support current understanding of the role of phytate in non-haem Fe absorption, which is that it inhibits Fe absorption irrespective of ascorbic acid.
Our algorithm and the two previously developed algorithms, discussed above, were applied to meals from an independent dataset from a study by Cook(3) in which a wider range of intakes of dietary modifiers were consumed compared with those from which our algorithm was created (Tables 1 and 2). We found a plausible range of Fe availability values across all meals (5–35 %). Percentage available Fe and percentage Fe absorption estimated using the other algorithms(4,5) exceeded 100 % in some subjects. Consistent with our discussion above, when using the Reddy et al. algorithm, predictions of non-haem Fe absorption approached or even exceeded 100 % when subjects were consuming relatively large intakes of animal tissue and, similarly, implausibly high absorption predictions were observed when using Hallberg & Hulthén’s algorithm in those individuals with relatively high intakes of animal tissue or ascorbic acid. Because of the short-term influence of the dietary modifiers on non-haem Fe absorption, i.e. only modifiers consumed simultaneously with Fe influence its availability, it is important that the algorithms are applied on a meal or eating-occasion basis, as was done here, and not, for example, on daily totals.
We support the view of Beiseigel et al. (25) that more research is required to validate an analytical method for measuring polyphenols that influence Fe availability. Our data demonstrated that while polyphenols from tea inhibited Fe availability, the same inhibition was not observed when looking at the total polyphenol content of the meals. Similarly, Hallberg & Hulthén(5) found that tea and coffee inhibit Fe absorption while Reddy et al. (4) found that the polyphenol content of mixed meals did not. The absence of beverages other than tea in our test meals meant that only the effect of polyphenols from tea was included in our algorithm, although we believe that this equation could be equally applied to coffee or other polyphenol-containing beverages. The absence of information on the polyphenol and phytate content of foods in many food databases needs to be rectified through more comprehensive analysis of these compounds in foods and the incorporation into food composition databases. The importance of both polyphenols and phytate means that their inhibiting effect on non-haem Fe absorption should not be ignored simply because of limited data.
Several studies have shown meat, fish and poultry to affect the absorption of non-haem Fe(26,27). However, little is known about the mechanism by which this occurs or differences in effect, if any, between meats and/or fish. For a review, see Layrisse et al. (28). For the purpose of our algorithm, we assume the effect of all red meats, poultry and fish to be equal. Further work is needed to determine if this is a valid assumption(28).
The algorithm presented here is relatively straightforward with the effect of each dietary modifier of non-haem Fe availability acting independently. In contrast, the more complex algorithm of Hallberg & Hulthén(5) attempts to adjust for interactions between dietary variables and this may be a limitation of our work and an area that could be further refined. We investigated how different available Fe was to Fe intake in the dataset of Cook(3); the two are highly correlated (r 0·89). This does not preclude the possibility that available Fe might relate quite differently to a health outcome compared with Fe intake.
The present investigation was able to show that our algorithm yields accurate estimates of Fe availability in the dataset used to construct the algorithm and provides plausible predictions of Fe availability using real dietary data while other algorithms do not. The next step is to test the validity of these predictions in an independent dataset in which dietary data and Fe absorption data are both present; future work will focus on this. The new algorithm described in the present paper estimates available Fe in the intestine within a more physiological plausible range than earlier algorithms. We believe that our algorithm could be a useful tool in epidemiological research to examine relationships between dietary Fe and health.
Acknowledgements
We thank Professor John Beard (Pennsylvania State University) for guidance on the application of the Hallberg and Hulthén algorithm. We thank Dr William Cook and colleagues (Medical Research Council (MRC) Human Nutrition Research) and Professor Catherine Geissler, Dr Miranda Lomer and colleagues (King’s College, London) for access to data from their recent studies. We are grateful to Dr Adrian Mander (MRC Human Nutrition Research) for input and discussion on algorithm design.
The algorithm in the paper has previously been included in conference proceedings.
A. P. R. is supported by a MRC PhD studentship, held at the MRC Collaborative Centre for Human Nutrition Research. The present study received no other specific grant from any funding agency in the public, commercial or not-for-profit sectors.
A. P. R. and J. J. P. conceived of and designed the research. R. E. C. was responsible for acquisition of data. A. P. R. and M. D. C. conducted the analysis design and interpretation. A. P. R. drafted and revised the manuscript. All authors discussed the core issues and helped to revise the manuscript.
Appendix A
Fig. 1.
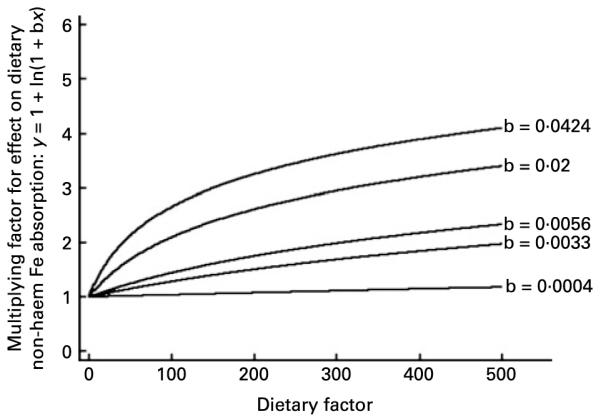
The effect of a dietary enhancer. The function ‘y = 1 + ln (1 + bx)’ is shown for different example values of b. We selected this function to describe the effect of dietary enhancers on dietary non-haem Fe availability in the algorithm designed in the present study. As shown in the Results section, the curve ‘b = 0·0056’ shows the precise fit for the relationship between ascorbic acid (mg) and dietary non-haem Fe availability in our algorithm.
Fig. 2.
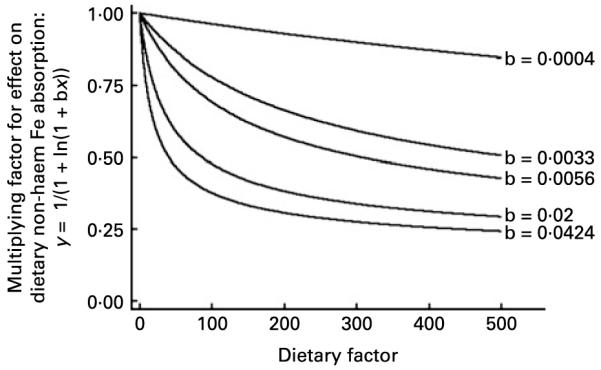
The effect of a dietary inhibitor. The function ‘y = 1/(1 + ln(1 + bx))’ is shown for example values of b. We selected this function to describe the effect of dietary inhibitors on dietary non-haem Fe availability in the algorithm designed in the present study. As shown in the Results section, the curve ‘b = 0·0424’ shows the precise fit for the relationship between dietary non-haem Fe intake (mg) and dietary non-haem Fe availability in this algorithm (i.e. inhibition of percentage Fe availability by Fe itself). Likewise, the line ‘b = 0·0033’ shows the precise relationship between non-haem Fe availability and phytate.
Fig. 3.
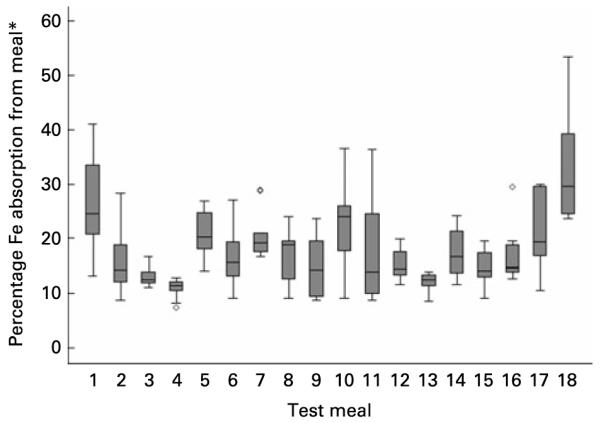
Variation in Fe absorption. Box and whisker plot showing the intra- and inter-meal variation in dietary non-haem Fe absorption across eighteen test meals from a study by Conway et al. (8). The central line is the median; the box represents the interquartile range; the whisker shows the normal range values; ○, outliers. * Estimated using serum Fe data and the Conway et al. method(8).
Fig. 4.
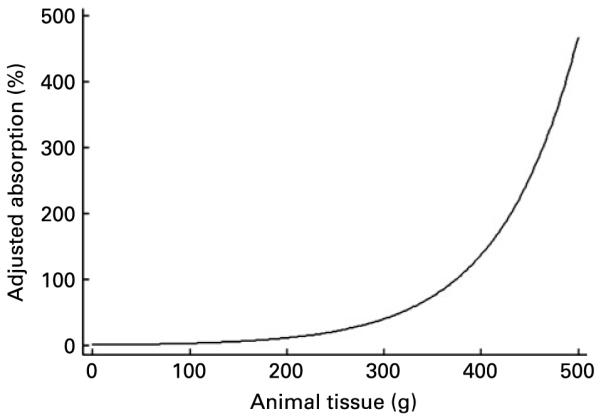
The effect of animal tissue on Fe absorption. The function shown was used to define the effect of animal tissue on percentage Fe absorption in the Reddy et al. algorithm(4). Ascorbic acid and phytic acid intake are assumed to be 0 in this representation.
Fig. 5.
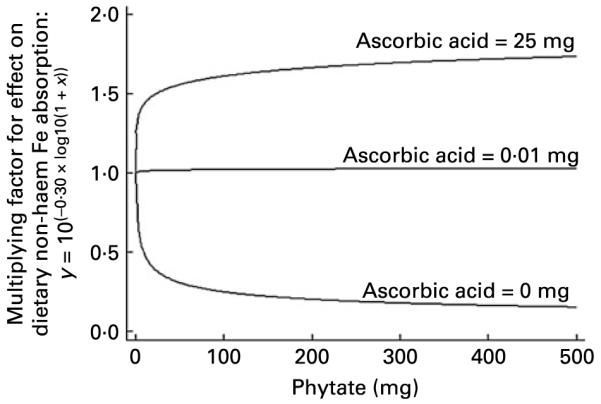
The effect of phytate on Fe absorption. The function shown was used to define the effect of phytate on percentage Fe absorption at different intakes of ascorbic acid in the Hallberg & Hulthén algorithm(5).
Appendix B
Reddy et al. algorithm(4)
where vitC is ascorbic acid (mg), phytate is phytate (mg), mfp is meat, fish and poultry (g) and ferritin is serum ferritin (μg/l).
Hallberg & Hulthén algorithm(5)
where vitC is ascorbic acid (mg), phytate is phytate (mg), calcium is Ca (mg), mfp is meat, fish and poultry (g), tan is tannic acid (mg), soy is soya protein (g), egg is number of eggs, alcohol is presence of alcohol (yes = 1, no = 0), ferritin is serum ferritin (μg/l) and log is log10.
Using the assumption that absorption in an Fe-deficient subject equals available Fe, percentage available Fe was calculated using the Hallberg & Hulthén algorithm(5) and the Reddy et al. algorithm(4) by calculating estimates for a hypothetical person with a serum ferritin of 10 μg/l, i.e. as if the subject consuming the meals were Fe deficient.
N.B. The algorithm presented here is the authors’ interpretation of the algorithm presented by Hallberg & Hulthén (i.e. we have included brackets where we expect they should be and, without which, the algorithm cannot be used) and in places is not identical to their published algorithm(5), even allowing for their subsequent published changes which were also due to errors in the original algorithm(29-31).
No haem Fe was present in the meals we tested, so Hallberg and Hulthén’s haem equation has not been included here.
Footnotes
There are no conflicts of interest for any author.
References
- 1.Reddy MB, Hurrell RF, Cook JD. Meat consumption in a varied diet marginally influences nonheme iron absorption in normal individuals. J Nutr. 2006;136:576–581. doi: 10.1093/jn/136.3.576. [DOI] [PubMed] [Google Scholar]
- 2.Conway RE, Powell JJ, Geissler CA. A food-group based algorithm to predict non-heme iron absorption. Int J Food Sci Nutr. 2007;58:29–41. doi: 10.1080/09637480601121250. [DOI] [PubMed] [Google Scholar]
- 3.Cook WB. Iron Absorption in Health and Inflammatory Bowel Disease. University of Cambridge; Cambridge, UK: 2009. [Google Scholar]
- 4.Reddy MB, Hurrell RF, Cook JD. Estimation of nonheme-iron bioavailability from meal composition. Am J Clin Nutr. 2000;71:937–943. doi: 10.1093/ajcn/71.4.937. [DOI] [PubMed] [Google Scholar]
- 5.Hallberg L, Hulthén L. Prediction of dietary iron absorption: an algorithm for calculating absorption and bio-availability of dietary iron. Am J Clin Nutr. 2000;71:1147–1160. doi: 10.1093/ajcn/71.5.1147. [DOI] [PubMed] [Google Scholar]
- 6.Thankachan P, Walczyk T, Muthayya S, et al. Iron absorption in young Indian women: the interaction of iron status with the influence of tea and ascorbic acid. Am J Clin Nutr. 2008;87:881–886. doi: 10.1093/ajcn/87.4.881. [DOI] [PubMed] [Google Scholar]
- 7.Beard JL, Murray-Kolb LE, Haas JD, et al. Iron absorption prediction equations lack agreement and underestimate iron absorption. J Nutr. 2007;137:1741–1746. doi: 10.1093/jn/137.7.1741. [DOI] [PubMed] [Google Scholar]
- 8.Conway RE, Geissler CA, Hider RC, et al. Serum iron curves can be used to estimate dietary iron bioavailability in humans. J Nutr. 2006;136:1910–1914. doi: 10.1093/jn/136.7.1910. [DOI] [PubMed] [Google Scholar]
- 9.Holland B, Welch A, Unwin I, et al. McCance and Widdowson’s The Composition of Foods. 5th ed. The Royal Society of Chemistry and Ministry of Agriculture, Fisheries and Foods; Cambridge, UK: 1991. [Google Scholar]
- 10.Monsen ER, Hallberg L, Layrisse M, et al. Estimation of available dietary iron. Am J Clin Nutr. 1978;31:134–141. doi: 10.1093/ajcn/31.1.134. [DOI] [PubMed] [Google Scholar]
- 11.Monsen ER, Cook JD. Food iron absorption in human subjects. IV. The effects of calcium and phosphate salts on the absorption of nonheme iron. Am J Clin Nutr. 1976;29:1142–1148. doi: 10.1093/ajcn/29.10.1142. [DOI] [PubMed] [Google Scholar]
- 12.Hallberg L, Brune M, Erlandsson M, et al. Calcium: effect of different amounts on nonheme- and heme-iron absorption in humans. Am J Clin Nutr. 1991;53:112–119. doi: 10.1093/ajcn/53.1.112. [DOI] [PubMed] [Google Scholar]
- 13.Cook JD, Dassenko SA, Whittaker P. Calcium supplementation: effect on iron absorption. Am J Clin Nutr. 1991;53:106–111. doi: 10.1093/ajcn/53.1.106. [DOI] [PubMed] [Google Scholar]
- 14.Hallberg L, Rossander L, Skanberg AB. Phytates and the inhibitory effect of bran on iron absorption in man. Am J Clin Nutr. 1987;45:988–996. doi: 10.1093/ajcn/45.5.988. [DOI] [PubMed] [Google Scholar]
- 15.Hallberg L, Brune M, Rossander L. Iron absorption in man: ascorbic acid and dose-dependent inhibition by phytate. Am J Clin Nutr. 1989;49:140–144. doi: 10.1093/ajcn/49.1.140. [DOI] [PubMed] [Google Scholar]
- 16.Siegenberg D, Baynes RD, Bothwell TH, et al. Ascorbic acid prevents the dose-dependent inhibitory effects of polyphenols and phytates on nonheme-iron absorption. Am J Clin Nutr. 1991;53:537–541. doi: 10.1093/ajcn/53.2.537. [DOI] [PubMed] [Google Scholar]
- 17.Reddy MB, Hurrell RF, Juillerat MA, et al. The influence of different protein sources on phytate inhibition of nonheme-iron absorption in humans. Am J Clin Nutr. 1996;63:203–207. doi: 10.1093/ajcn/63.2.203. [DOI] [PubMed] [Google Scholar]
- 18.Brune M, Rossander L, Hallberg L. Iron absorption and phenolic compounds: importance of different phenolic structures. Eur J Clin Nutr. 1989;43:547–557. [PubMed] [Google Scholar]
- 19.Tuntawiroon M, Sritongkul N, Brune M, et al. Dose-dependent inhibitory effect of phenolic compounds in foods on nonheme-iron absorption in men. Am J Clin Nutr. 1991;53:554–557. doi: 10.1093/ajcn/53.2.554. [DOI] [PubMed] [Google Scholar]
- 20.Tseng M, Chakraborty H, Robinson DT, et al. Adjustment of iron intake for dietary enhancers and inhibitors in population studies: bioavailable iron in rural and urban residing Russian women and children. J Nutr. 1997;127:1456–1468. doi: 10.1093/jn/127.8.1456. [DOI] [PubMed] [Google Scholar]
- 21.Hoglund S, Reizenstein P. Studies in iron absorption. V. Effect of gastrointestinal factors on iron absorption. Blood. 1969;34:496–504. [PubMed] [Google Scholar]
- 22.Hunt JR. Dietary and physiological factors that affect the absorption and bioavailability of iron. Int J Vitam Nutr Res. 2005;75:375–384. doi: 10.1024/0300-9831.75.6.375. [DOI] [PubMed] [Google Scholar]
- 23.Heath AL, Skeaff CM, Gibson RS. The relative validity of a computerized food frequency questionnaire for estimating intake of dietary iron and its absorption modifiers. Eur J Clin Nutr. 2000;54:592–599. doi: 10.1038/sj.ejcn.1601063. [DOI] [PubMed] [Google Scholar]
- 24.Prynne CJ, McCarron A, Wadsworth MEJ, et al. Dietary fibre and phytate, a balancing act. Results from three timepoints in a British Birth Cohort. Br J Nutr. 2009 doi: 10.1017/S0007114509991644. In the Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Beiseigel JM, Hunt JR, Glahn RP, et al. Iron bio-availability from maize and beans: a comparison of human measurements with Caco-2 cell and algorithm predictions. Am J Clin Nutr. 2007;86:388–396. doi: 10.1093/ajcn/86.2.388. [DOI] [PubMed] [Google Scholar]
- 26.Cook JD, Monsen ER. Food iron absorption in human subjects. III. Comparison of the effect of animal proteins on nonheme iron absorption. Am J Clin Nutr. 1976;29:859–867. doi: 10.1093/ajcn/29.8.859. [DOI] [PubMed] [Google Scholar]
- 27.Hallberg L. Bioavailability of dietary iron in man. Annu Rev Nutr. 1981;1:123–147. doi: 10.1146/annurev.nu.01.070181.001011. [DOI] [PubMed] [Google Scholar]
- 28.Layrisse M, Martinez-Torres C, Roche M. Effect of interaction of various foods on iron absorption. Am J Clin Nutr. 1968;21:1175–1183. doi: 10.1093/ajcn/21.10.1175. [DOI] [PubMed] [Google Scholar]
- 29.Hallberg L, Hulthen L. Erratum for Prediction of dietary iron absorption: an algorithm for calculating absorption and bioavailability of dietary iron. Am J Clin Nutr. 2000;72:1242. doi: 10.1093/ajcn/71.5.1147. [DOI] [PubMed] [Google Scholar]
- 30.Hallberg L, Hulthen L. Erratum for Prediction of dietary iron absorption: an algorithm for calculating absorption and bioavailability of dietary iron. Am J Clin Nutr. 2001;74:274. doi: 10.1093/ajcn/71.5.1147. [DOI] [PubMed] [Google Scholar]
- 31.Hallberg L, Hulthen L. Erratum for Prediction of dietary iron absorption: an algorithm for calculating absorption and bioavailability of dietary iron. Am J Clin Nutr. 2006;84:1253. doi: 10.1093/ajcn/71.5.1147. [DOI] [PubMed] [Google Scholar]