

Abstract
Foot-and-mouth disease (FMD) continues to be a significant threat to the health and economic value of livestock species. This acute infection is caused by the highly contagious FMD virus (FMDV), which infects cloven-hoofed animals, including large and small ruminants and swine. Current vaccine strategies are all directed toward the induction of neutralizing antibody responses. However, the role of cytotoxic T lymphocytes (CTLs) has not received a great deal of attention, in part because of the technical difficulties associated with establishing a reliable assay of cell killing for this highly cytopathic virus. Here, we have used recombinant human adenovirus vectors as a means of delivering FMDV antigens in a T cell-directed vaccine in pigs. We tested the hypothesis that impaired processing of the FMDV capsid would enhance cytolytic activity, presumably by targeting all proteins for degradation and effectively increasing the class I major histocompatibility complex (MHC)/FMDV peptide concentration for stimulation of a CTL response. We compared such a T cell-targeting vaccine with the parental vaccine, previously shown to effectively induce a neutralizing antibody response. Our results show induction of FMDV-specific CD8+ CTL killing of MHC-matched target cells in an antigen-specific manner. Further, we confirm these results by MHC tetramer staining. This work presents the first demonstration of FMDV-specific CTL killing and confirmation by MHC tetramer staining in response to vaccination against FMDV.
Foot-and-mouth disease (FMD) is a highly infectious viral disease that affects cloven-hoofed animals and remains an important threat to livestock throughout much of the world (reviewed in reference 18). Many species of wild animals are susceptible to infection, but agricultural concerns are particularly focused on swine, cattle, sheep, and goats. This acute disease is characterized by fever and viremia that last 1 to 2 days, the formation of vesicular lesions in the mouth and on the feet and teats, lethargy, lameness, and loss of meat and milk production. Clinical signs of disease resolve within 7 to 10 days; however, an asymptomatic persistent infection (carrier state) can develop, particularly in previously vaccinated cattle (14, 17). The etiological cause of disease is FMD virus (FMDV), which has a single-stranded, positive-sense RNA genome of 8 kb. FMDV is a member of the Picornaviridae family, genus Aphthovirus. Following entry of the virus into the cell and uncoating, the released viral genome serves as an mRNA for the translation of viral proteins. Initially synthesized as a single polypeptide, the protein is subsequently cleaved by viral proteases, including leader (Lpro) and 3Cpro, into mature structural and nonstructural proteins (53).
FMDV is highly cytopathic, and several viral proteins are known to play a role in disrupting the host immune response (reviewed in reference 19). Lpro cleaves the host translation initiation factor eIF4G, which plays a critical role in cap-dependent mRNA translation (12). An internal ribosomal entry site (IRES) in the 5′ untranslated region of the FMDV genome allows synthesis of viral proteins to continue while effectively shutting down host protein synthesis (2, 27). Lpro also promotes the degradation of NF-κB, a regulator of beta interferon (IFN-β) transcription (10). The combined actions of Lpro allow FMDV to block synthesis of the antiviral proteins IFN-α and -β, to which FMDV is sensitive (7, 9). In addition to the role of 3Cpro in cleaving the viral polyprotein into mature protein products, it has been shown to induce the cleavage of eIF4A and eIF4G (3). This protease has also been implicated in the cleavage of histone H3 (15, 54), thus further inhibiting host transcription and translation. In addition, both the precursor protein 2BC and the coexpressed mature products, 2B and 2C, are involved in disruption of the cellular secretory pathway (33, 34). Combined with the loss of host protein synthesis, this disruption of the secretory pathway appears to be responsible for inhibition of new MHC class I translation and surface expression in FMDV-infected cells (51).
The immunopathology of this acute infection includes a transient lymphopenia (1, 13). Viral infection was shown to lead to inhibition of the type I IFN response of multiple populations of dendritic cells (36, 37). In vitro, natural killer (NK) cells have been shown to lyse FMDV-infected cells (56). However, analysis of cells from FMDV-infected animals shows a loss of NK cell function during the acute phase of infection (55) (reviewed in reference 16).
The development of neutralizing antibodies plays a central role in FMDV immunity and often correlates with protection of vaccinated animals against live virus challenge (32). Currently available vaccines consist of killed virus in adjuvant and are effective at providing protection but do not allow for the “differentiation of infected from vaccinated animals” (DIVA) (48). A leading candidate for a next-generation FMDV vaccine is a replication-defective human adenovirus 5 (Ad5) viral vector delivering FMDV capsid precursor (P1) and 3Cpro, an “empty capsid” vaccine. This platform has DIVA capability and is very compatible with diagnostic tests in use at present. The parent polypeptide is processed by the 3Cpro into the protein components of the capsid (reviewed in references 20 and 48). This vaccine strategy is effective in swine (30, 31, 35) and cattle (39). However, like the killed-virus vaccine and natural infection, neutralizing antibodies raised against the vaccine strain do not provide cross-serotype protection. This limits the value of vaccination during an FMDV outbreak or in disease eradication (48), as virus strain matching is required for vaccine efficacy.
CD8+ cytotoxic T lymphocytes (CTLs) play an important role in adaptive immunity and are capable of directly killing virus-infected cells in an antigen-specific manner through the release of perforin and granzymes as well as through Fas-Fas ligand interactions (reviewed in references 5 and 58). Unlike antibodies which recognize complex antigens, CTLs recognize antigens that have been processed and cleaved into 8- to 12-amino-acid peptides by the proteasome and then loaded into MHC class I molecules in the lumen of the endoplasmic reticulum (reviewed in reference 46). These peptide/MHC complexes are then transported to the surface of the infected cell, where they are displayed to T cell surveillance. CTL recognition occurs when an appropriate T cell receptor (TCR), in association with the CD8 molecule, stably binds to the peptide/MHC complex, resulting in stimulatory signaling cascades within the CTL (58). The ability of CTLs to recognize epitopes that remain hidden to neutralizing antibodies makes them attractive candidates for enhancing FMDV immunity.
In contrast to the focus on neutralizing antibodies, the role of CTLs in FMDV immunity has gone largely unexplored. The ability of the virus to inhibit MHC synthesis and transport and the speed with which the virus is cleared presumably result in limited antigen exposure for MHC class I-restricted T cells. In addition to the virus-induced lymphopenia, data from in vitro studies would predict that FMDV-specific CTLs are not elicited by natural FMDV infection if MHC class I expression is blocked in vivo as has been described in vitro (51). This would make FMDV-infected cells invisible to class I-restricted CTLs. Given the cytopathic nature of the virus, this hypothesis has gone largely untested, as FMDV-infected cells are rapidly lysed in vitro and cannot be used as target cells in assays for antigen-specific cell lysis.
We have overcome these limitations with two alternative approaches. First, we employed recombinant Ad5 FMDV vaccine technology as a method for delivering FMDV antigens to both stimulating and target cells for the in vitro CTL killing assay. Here, we describe our development of an assay using swine leukocyte antigen (SLA) homozygous pigs and the MHC-matched PK (15) cells as target cells. Second, in order to maximize the CTL response, we used a vaccine construct containing a mutation in 3Cpro that inhibits its ability to process P1 into mature capsids. This approach tests the hypothesis that one result of reduced processing of the primary polypeptide precursor would be an increase in the pool of FMDV proteins available for loading of FMDV-derived peptides into MHC class I molecules. This is then predicted to enhance stimulation of CTLs specific for FMDV peptides.
Further, to confirm that CTL killing was antigen specific and MHC restricted, we developed class I MHC tetramers using the NetMHCpan prediction algorithm developed for human class I MHC (38) and extended to other species, including swine (reference 25 and this report). Here we report that the T cell-targeting vaccine elicited a greater overall CTL killing response than the parental vaccine and correlated with the induction of FMDV-specific CTLs by MHC tetramer staining.
MATERIALS AND METHODS
Cell lines.
Porcine kidney [PK(15)] cells (ATCC CCL-33) and PK(15)-EGFP cells (described below) were maintained in minimal essential medium (MEM) (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, Waltham, MA) and 3.4 mM l-glutamine, 3 mM dextrose, 1 mM sodium pyruvate, 5% sodium bicarbonate, nonessential and essential amino acids, 2-mercaptoethanol, and antibiotics (MEM-10). Baby hamster kidney (BHK-21) cells (ATCC CCL-10) were maintained in basal medium Eagle (BME) (Invitrogen, Carlsbad, CA) supplemented with 10% FBS, 10% tryptose phosphate broth, 2 mM l-glutamine, and antibiotics (BME-10). Human 293 cells (ATCC CRL-1573) were maintained in MEM supplemented with 10% FBS, 2 mM l-glutamine, and antibiotics. Peripheral blood mononuclear cells (PBMCs) were maintained in Roswell Park Memorial Institute (RPMI)-1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% FBS, 3.4 mM l-glutamine, 3 mM dextrose, 1 mM sodium pyruvate, 5% sodium bicarbonate, nonessential and essential amino acids, 2-mercaptoethanol, and antibiotics. IBRS2 (swine kidney) cells were maintained in MEM supplemented with 10% FBS, 2 mM l-glutamine, and antibiotics.
Animals.
Yucatan 4b.0/4b.0 (24) pigs were obtained from Sinclair Research, Inc., Columbia, MO. Animals were held for at least 1 week before being entered into a study. All animal procedures were reviewed and approved by the Plum Island Institutional Animal Care and Use Committee.
Antibodies.
A mouse monoclonal antibody (MAb) against porcine CD6, clone MIL8, was purchased from AbD Serotec (Raleigh, NC). Mouse MAbs against porcine CD8 (R-phycoerythrin [RPE], clone 76-2-11) and CD4 (fluorescein isothiocyanate [FITC], clone 74-12-4) were purchased from BD Biosciences (San Jose, CA). Monoclonal anti-FMDV-VP1 (clone 6HC4.1.3 [45]) and anti-VP2 (VP0) (clone F1412SA [57]) were used in combination to probe for Western blotting. In addition, a mixture of rabbit polyclonal antibodies raised against the FMDV capsid proteins (VP1, VP2, and VP3) were used for Western blotting (43). Anti-tubulin-α MAb was used for control staining of the Western blot (clone DM1A from Thermo Scientific, Fremont, CA).
Viruses.
The production of viruses Ad5A24 (Ad5-FMDV-B), Ad5VSV-G (Ad5-VSV-G) (35), and Ad5-Blue (Ad5-Empty) (35) have been previously described. A mutation in the active site of the 3Cpro gene (21) was subcloned into pAd5-A24, replacing the parental, wild-type 3Cpro, and was termed pAd5A24-3C-mut. Production of this new adenovirus, Ad5A24-3C-mut (Ad5-FMDV-T), was otherwise identical to that of the parental virus. All adenoviruses were grown in 293 cells. FMDV, strain A24 Cruzeiro, was propagated in BHK-21 cells.
Western blotting.
IBRS2 cells were seeded in a 96-well microplate at 20,000 cells/well in 50 μl supplemented, phenol red-free MEM and incubated for 24 h at 37°C. Following attachment, cells were infected with adenovirus constructs at a multiplicity of infection (MOI) of 20, or with FMDV A24 at an MOI of 10, and then incubated for 24 h or 5 h, respectively. Whole-cell lysates were prepared, and protein expression was evaluated by separation by SDS-PAGE followed by Western blotting.
Vaccination.
Six Yucatan 4b.0/4b.0 pigs were divided into two groups of three pigs each and vaccinated with either Ad5-FMDV-B or Ad5-FMDV-T. PBS was administered to one additional pig as a negative control. For both vaccination and boosting, animals were given a total of 5 × 109 PFU of the appropriate adenovirus. Virus was delivered intradermally in a total of 2 ml to four sites on the neck (0.5 ml/site) using the DERMA-VAC NF needle-free system (Merial, Duluth, GA). Pigs were boosted 12 weeks postvaccination and again 11 or 13 months postvaccination.
Isolation of PBMCs.
Porcine blood was collected in Vacutainer tubes containing heparin. Typically, 20 ml of blood was diluted with 14 ml of phosphate-buffered saline (PBS), followed by the addition of a 12-ml underlay of Lymphoprep (Axis-Shield, Oslo, Norway), and then centrifuged at 1,000 × g for 20 min at room temperature. The band of peripheral blood mononuclear cells (PBMCs) was harvested and washed three times with PBS, followed by suspension in RPMI.
Stimulator/target cell preparation.
PK(15) cells were chosen because they are MHC haplotype 4b.0/4.b0 (23), matching the MHC locus of the Yucatan swine under study. To prepare stimulator cells, PK(15) cells were seeded into 24-well plates and allowed to attach overnight. Cell medium was removed and cells were infected with adenovirus at an MOI of 10 in 0.2 ml of RPMI 1640 (nonsupplemented) for 1 h at 37°C with occasional rocking. For target cells, we developed PK(15)-EGFP cells. pEGFP-C1 (Clontec, Mountain View, CA) was linearized by digestion with PciI (New England BioLabs, Ipswich, MA) and transfected into PK(15) cells using the ProFection mammalian transfection system (Promega, Madison, WI). Stably transfected cells were selected by growth in the presence of Geneticin (500 μg/ml) (Invitrogen, Carlsbad, CA) and cloned by two rounds of limiting dilution. For the assay, 2 × 106 cells were infected with the appropriate virus at an MOI of 10 for 1 h in a low volume of MEM. Target cells were then given MEM-10 and incubated overnight at 37°C.
CD6+ purification.
PBMCs isolated from vaccinated pigs were suspended at a concentration of 107 cells/ml, and 1 ml was added to each well of infected PK(15) cells (typically 10 to 16 replicate wells), followed by the addition of 1 ml/well of RPMI 1640 (supplemented). Plates were incubated at 37°C for 3 to 5 days. Stimulated PBMCs were recovered from wells and reisolated on Lymphoprep in order to remove dead cells. To positively select CD6+ cells, purified PBMCs were labeled with mouse anti-porcine CD6 MAb (1 μl/106 cells) followed by MACS goat anti-mouse IgG microbeads and separated on LS columns using a MidiMACS magnetic separator (Miltenyi Biotec, Gladbach, Germany), according to the manufacturer's instructions, resulting in approximately 95% CD6+ cells (data not shown).
Cytotoxic T lymphocyte killing assay.
In vitro expanded, CD6+-enriched cells were suspended in supplemented RPMI 1640 at a concentration of 107 cells/ml, plated (100 μl/well) in a low-binding 96-well plate (Thermo Fisher Scientific, Waltham, MA), and diluted 2-fold to obtain final effector-to-target-cell (E:T) ratios of 50:1, 25:1, 12:1, and 6:1. Trypsinized PK(15)-EGFP target cells were suspended at 105 cells/ml and added to the effector cells (100 μl/well). In order to discriminate dead target cells from live cells, 5 μl of 7-amino-actinomycin D (7-AAD) (BD Biosciences, San Jose, CA) was added to each well. The plate was centrifuged at 233 × g for 1 min and then incubated at 37°C for 4 h. Following incubation, cells were resuspended and data were acquired by a FACSCalibur flow cytometer with a high-throughput sampler (HTS) (BD Biosciences, San Jose, CA). Data were analyzed using CellQuest Pro software (BD Biosciences, San Jose, CA) by gating on PK(15)-EGFP cells and determining the percentage of 7-AAD-positive (lysed) cells. Background lysis was determined by 7-AAD staining of target cells in the absence of effectors, and this percentage was subtracted from all groups.
Serum neutralization assay.
Endpoint titers of serum neutralization of FMDV A24 were determined according to the procedure described by Pacheco and coworkers (40). Briefly, 4-fold serum dilutions were incubated at 37°C with 100 50% tissue culture infective doses (TCID50) of FMDV serotype A24 for 1 h in a microtiter plate, after which BHK-21 cells were added (104 cells/well). Following incubation for 3 days, wells were scored for cytopathic effect and endpoint titers were calculated, which represent the reciprocal of the log dilution of serum able to neutralize 100 TCID50 of virus.
Tetramer construction and staining.
Peptides and recombinant SLA-1*0401 and human β2-microglobulin (β2m) were produced and complexed into tetramers as described previously (28) with minor modifications. Briefly, recombinant β2m and peptides were added to 10-fold and 40-fold excesses, respectively, of SLA-1*0401 in a reaction buffer containing a cocktail of protease inhibitors [0.68 mM EDTA, 4.8 μM pepstatin A, 94.4 μM 1-10 phenanthroline, 0.1 mM N-ethylmaleimide, 8.9 μM tosyl-l-lysine chloromethyl ketone (TLCK), 9.3 μM tosyl-l-phenylalanine chloromethyl ketone (TPCK), 66.8 μM phenylmethylsulfonyl fluoride (PMSF)] followed by the addition of streptavidin-allophycocyanin (SA-APC). For tetramer staining, 2 × 105 to 5 × 105 cells were washed with cold FACS buffer (PBS, 0.3% bovine serum albumin, 0.1% sodium azide) and resuspended in solution containing tetramer previously mixed 1:1 with 2× FACS buffer. Cells were incubated at room temperature with rocking for 20 min and then washed three times in cold FACS buffer. Cells were resuspended in FACS buffer containing fluorescent MAbs against porcine CD8 and CD4 and then incubated at room temperature for 20 min. After being washed three times in FACS buffer, cells were fixed in PBS containing 1% formaldehyde and stored at 4°C until being analyzed by flow cytometry.
RESULTS
Mutation of FMDV 3C protease results in reduced capsid processing and production of anti-FMDV antibodies.
Previous work by Moraes and coworkers demonstrated that vaccination of swine with Ad5A24 (Ad5-FMDV-B) resulted in the generation of a protective anti-FMDV antibody response (35). Mutation of the 3C protease resulting in an amino acid change (C163S) is known to inhibit processing of P1 into mature capsid proteins (21). A second adenovirus construct with the same P1 polyprotein derived from FMDV strain A24 and the mutated 3C protease was made, which we designate Ad5-FMDV-T. This construct was predicted to provide enhanced stimulation of a CTL response based on previous work that demonstrated a lack of antibody response to a similar construct made for FMDV strain A12 (31).
To demonstrate the reduced activity of 3C in this new construct, IBRS2 cells were infected with Ad5-FMDV-B, Ad5-FMDV-T, Ad5-Empty, or FMDV strain A24. Cell lysates were analyzed by SDS-PAGE followed by Western blotting. We detected the presence of P1 cleavage products VP0, VP3, and VP1 in the lysates of cells infected with either FMDV or Ad5-FMDV-B, with little or no P1 precursor (Fig. 1). In contrast, cells infected with Ad5-FMDV-T showed a significant decrease in the presence of VP0, VP3, and VP1, with significantly more unprocessed P1 (Fig. 1), suggesting a reduction of 3Cpro activity and consequent accumulation of P1.
FIG. 1.

Reduced capsid processing of Ad5-FMDV-T. IBRS2 cells were infected with the indicated virus as described in Materials and Methods. Following infection, cell lysates were made and proteins were separated by SDS-PAGE and visualized by Western blotting using either MAbs against VP2 (visualized as the VP0 precursor) and VP1 (left) or a mix of rabbit polyclonal antibodies against FMDV capsid proteins (right).
Sera from pigs vaccinated and boosted with Ad5-FMDV-B and Ad5-FMDV-T were tested for FMDV neutralizing antibodies as described in Materials and Methods. All three pigs vaccinated with Ad5-FMDV-B generated neutralizing antibodies following priming, with elevated titers following two rounds of boosting (Fig. 2). In contrast, of the three pigs vaccinated with Ad5-FMDV-T, only p945 generated detectable neutralizing antibodies following priming. Both p945 and p947 had serum neutralizing antibodies following boost, with the titer for p947 remaining relatively low. Neutralizing antibodies were never detected in serum from p925 or from the PBS negative control, p863. These data suggest that Ad5-FMDV-T is less effective than Ad5-FMDV-B at eliciting neutralizing antibodies, presumably due to less efficient processing of P1 by 3Cpro.
FIG. 2.

Reduced neutralizing antibodies in Ad5-FMDV-T-vaccinated animals. Serum samples from pigs vaccinated with PBS, Ad5-FMDV-B, or Ad5-FMDV-T were collected before and following priming and boosting (solid bars, prevaccination; open bars, preboost; striped bars, pre-second boost; shaded bars, final). Serum neutralizing titers (SNT) were determined as described in Materials and Methods. Final serum was collected 26 days or more postboost.
Establishment of an FMDV CTL assay.
The animals vaccinated with either Ad5-FMDV-T, Ad5-FMDV-B, or PBS were boosted 12 weeks postvaccination and analyzed. PBMCs were collected 24 days postboost and stimulated for 5 days in the presence of PK(15) cells infected with Ad5-FMDV-T and then tested for cytolytic activity against target cells that were either mock infected or infected with Ad5-FMDV-T. Consistent with our hypothesis, we observed the greatest antigen-specific cytolysis by cells derived from pigs vaccinated with Ad5-FMDV-T (Fig. 3). Due to low effector cell numbers, we were unable to evaluate the response against mock-infected cells for p864 and p925, and thus we omitted data from these two animals from Fig. 3. In spite of the observed elevation of antigen-specific cytolysis, with the exception of p945 we observed poor titration of target cell killing across several effector-to-target-cell (E:T) ratios (data not shown). A similar lack of titration of cytolysis by bulk PBMCs was previously observed by Denyer and coworkers, who demonstrated that purification of CD6+ cells by positive selection prior to incubating with target cells resulted in titration of cytolytic activity across E:T ratios (11).
FIG. 3.

Elevated CTL activity in Ad5-FMDV-T-vaccinated animals. PBMCs were isolated from vaccinated animals, stimulated in the presence of Ad5-FMDV-T-infected PK(15) cells for 5 days, and then assayed for cytolytic activity against Ad5-FMDV-T-infected or mock-infected target cells, as described in Materials and Methods. The mean and standard deviation of results for duplicate wells of an E:T ratio of 50:1 are shown.
Based on this information, we enriched for CD6+ cells following antigen stimulation in all further experiments, as described in Materials and Methods. PBMCs isolated from pigs vaccinated with Ad5-FMDV-T and enriched for CD6 expression exhibited enhanced cytolysis of Ad5-FMDV-T-infected target cells that titrated across E:T ratios, compared to mock or Ad5-Empty-infected cells (Fig. 4). The level of killing detected by this refined assay was slightly reduced, likely due to the elimination of NK cells from the responding population. The animal vaccinated with PBS showed no specific killing (see Fig. S1 in the supplemental material). In subsequent figures, we show the data for the 50:1 E:T ratio only, with the cytolysis of Ad5-Empty-infected cells subtracted from each group, though all four E:T ratios were always run. Finally, with this new assay, we tested the kinetics of the expansion culture over the course of 5 days and found that the highest level of killing, with minimum background, was detected on day 3 (data not shown), so all expansion cultures were done for 3 days.
FIG. 4.
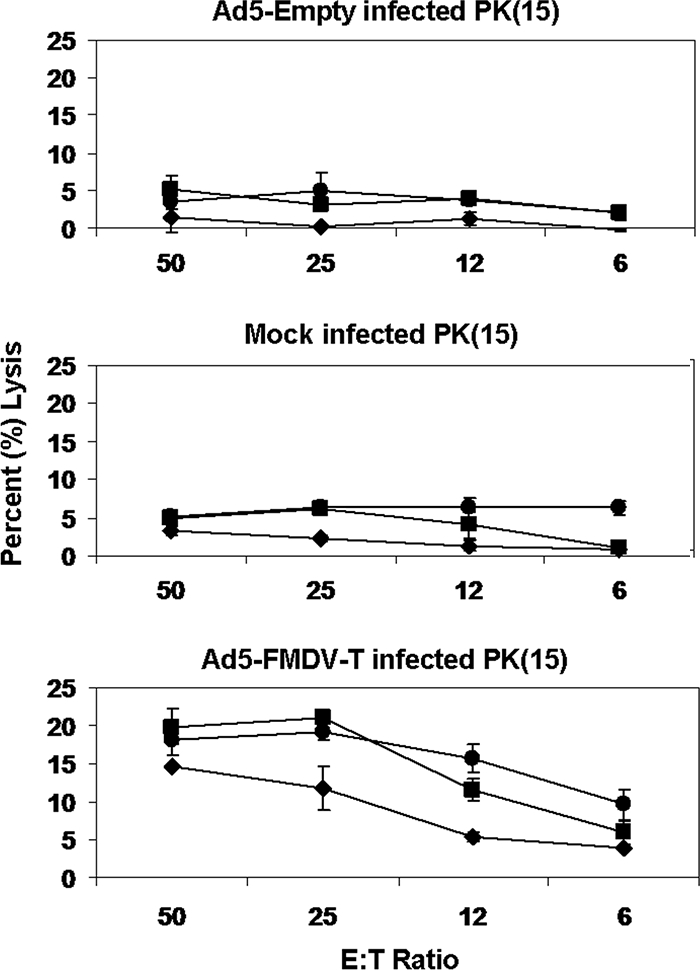
Titration of CD6+ CTL activity. PBMCs isolated from pigs vaccinated with Ad5-FMDV-T (⧫, p925; ▪, p945; •, p947) were stimulated with Ad5-FMDV-T-infected cells for 3 days and then enriched for CD6 surface expression. Effector and target cells were mixed at E:T ratios of 50:1, 25:1, 12:1, and 6:1. Cytolysis of target cells was measured by flow cytometry with target cells mock infected or infected with Ad5-Empty or Ad5-FMDV-T. All error bars represent the standard deviation of results for duplicate wells.
Specificity of CTL cytolytic activity for FMDV P1.
The development of this CTL assay was achieved with these vaccinated and boosted animals such that by 9 months following the first boost, antigen-specific cytolysis was difficult to detect, suggesting a waning of CTL immunity. Therefore, all pigs were boosted a second time with the vaccinating virus construct (p864 to p866 with Ad5-FMDV-B and p925, p945, and p947 with Ad5-FMDV-T) and activation of CTL killing was tracked using the assay.
In order to test the ability of the different adenovirus constructs to stimulate CTLs, PBMCs isolated from pigs 4 days after boosting with Ad5-FMDV-B were stimulated with cells infected with either Ad5-FMDV-B or Ad5-FMDV-T. Following enrichment for CD6+ cells, the effector cells were mixed with target cells infected with either Ad5-FMDV-B, Ad5-FMDV-T, Ad5-Empty, or Ad5-VSV-G or mock infected. Ad5-VSV-G contains the gene for glycoprotein G of vesicular stomatitis virus, strain New Jersey (VSV-NJ), and served as a negative-control antigen. CD6+ cells stimulated with Ad5-FMDV-B, compared to those stimulated with Ad5-FMDV-T, appeared to show less overall target cell killing, but there was no statistical difference between these groups (Fig. 5). Specificity for FMDV P1 was shown by little or no cytolysis of target cells infected with Ad5-VSV-G or mock infected (Fig. 5), eliminating the possibility that expression of any adenovirus-delivered transgene may contribute to cell lysis independent of antigen specificity.
FIG. 5.

Differential antigen stimulation. PBMCs from Ad5-FMDV-B-vaccinated pigs were isolated 4 days postboost and stimulated for 3 days with PK(15) cells infected with either Ad5-FMDV-T or Ad5-FMDV-B. Following CD6+ purification, effector cells were assayed for cytolysis of target cells infected with Ad5-FMDV-T (solid bars), Ad5-FMDV-B (open bars), or Ad5-VSV-G (shaded bars) or mock-infected (striped bars) cells. Data are shown relative to Ad5-Empty. The mean and standard deviation of results for triplicate wells are indicated.
Vaccine targeting T cells induces an enhanced CTL response.
Following the second boost of animals, PBMCs were obtained on days 4, 8, 14, and 28 and CTL killing activity for each pig was assayed. Antigen-specific cytolysis remained low throughout the experiment in animals vaccinated with the Ad5-FMDV-B construct. In Fig. 6, a small rise in antigen-specific killing on day 4 after the second boost is apparent. Low CTL lysis was observed when analyzing cells taken the day of the boost or 8 and 14 days following the boost. These data suggest that Ad5-FMDV-B is a poor inducer of CTLs. Data from all time points are shown relative to Ad5-Empty, except for data for day 14, which are shown relative to Ad5-VSV-G because small numbers of effector cells isolated necessitated omitting mock and Ad5-Empty target groups.
FIG. 6.

CD6+ CTL activity generated following second boost. PBMCs from Ad5-FMDV-B (p864, p865, and p866)- or Ad5-FMDV-T (p925, p945 and p947)-vaccinated pigs were isolated before and following boosting and treated as described in the legend to Fig. 5, except that only data from Ad5-FMDV-T secondary antigen stimulation are shown. Target cells: ⧫, Ad5-FMDV-T; ▪, mock. Additionally, pigs 864, 865, and 866 were tested on target cells: ▴, Ad5-FMDV-B; •, Ad5-VSV-G. Data from all time points are shown relative to Ad5-Empty. Error bars represent the standard deviation of results for duplicate wells.
PBMCs from pigs boosted with Ad5-FMDV-T and stimulated ex vivo with Ad5-FMDV-T-infected cells were tested for lysis of target cells infected with Ad5-FMDV-T, Ad5-Empty, or mock infected. In contrast to cells from the Ad5-FMDV-B-vaccinated animals, here we observed increased levels of antigen-specific cytolysis 4 days following the boost that waned to preboost levels by day 14 or 28 (Fig. 6). In all three animals, a high level of specific killing of PK(15) cells infected with the Ad5-FMDV-T construct was observed. Pig 925 had little killing activity 8 days after vaccine boost but good activity (equivalent to the day 4 result) when assayed 14 days after boost. This may indicate a technical problem with that day's sample from that animal (day 8, p925). As shown above, this animal (p925) showed no detectable neutralizing antibody either (Fig. 2). With that exception, the Ad5-FMDV-T-vaccinated animals had higher levels of target cell killing than the Ad5-FMDV-B-vaccinated animals through 14 days following the boost.
Animals vaccinated and boosted with Ad5-FMDV-T showed elevated mean killing of Ad5-FMDV-T-infected target cells compared to that of those vaccinated and boosted with Ad5-FMDV-B when cells isolated 4 days after the respective final boost were analyzed. This enhanced killing was statistically significant (P < 0.05; Fig. 7). As described above, the animal that was given PBS as a negative control never showed antigen-specific killing of target cells (see Fig. S1 in the supplemental material).
FIG. 7.
CD6+ CTL activity generated by Ad5-FMDV-T vaccination. Mean CTL cytolysis activity on day 4 following final boost of pigs vaccinated with Ad5-FMDV-B or Ad5-FMDV-T was determined after subtracting percentage of lysis of the highest negative-control target cell group from that of Ad5-FMDV-T target cells for each individual pig. Error bars represent the standard deviations (n = 3), and the asterisk indicates statistical significance (P < 0.05) as determined by an unpaired, one-tailed Student's t test with unequal variances.
CTL cytolytic activity correlates with staining by an FMDV peptide-SLA-1 tetramer.
In order to further characterize the effector cells, we developed pMHC tetramers for the SLA-1 molecule expressed by these animals, SLA-1*0401. The MHC class I peptide binding prediction algorithm NetMHCpan has previously been demonstrated to accurately capture the binding specificity of SLA class I molecules (25). Using this prediction method, we identified a peptide (MTAHITVPY) derived from the P1 capsid precursor of FMDV strain A24 that binds SLA-1*0401 at a low Kd (dissociation constant) (L. E. Pedersen, M. N. Harndahl, K. Lamberth, W. T. Golde, O. Lund, M. Nielsen, and S. Buus, unpublished data). As a negative control, a second SLA-1 tetramer was used that contained a peptide (ATAAATEAY) derived from the Ebola virus VP35 protein, which also binds to SLA-1*0401. For the experiments associated with the second boost (Fig. 6), a portion of the PBMCs isolated for evaluation of CTL killing were set aside for SLA-1 tetramer staining.
Cells were stained with antibodies specific for CD4 and CD8 and with the tetramers. Analysis by flow cytometry was carried out by gating on the CD4−/CD8+ cell population and determining the percentage of tetramer-positive (tetramer+) cells (see Fig. S2 in the supplemental material). For pigs vaccinated with Ad5-FMDV-B, little tetramer staining was detected from samples taken 4 days following the second boost (Fig. 8) or thereafter (data not shown). This result correlated with low levels of cytolytic activity from the same samples (Fig. 6). In contrast, pigs vaccinated with Ad5-FMDV-T had a 2- to 3-fold increase, compared to preboost levels, in tetramer staining of the CD8+ population isolated 4 days following the second boost (Fig. 8). Increased tetramer staining was correlated with peak cytolysis (compare Fig. 6 and Fig. 8) and was no longer detectable by 8 days after the boost. These data show that FMDV peptide MTAHITVPY is presented by SLA-1*0401 molecules in vivo and serve as confirmation of the presence of anti-FMDV, MHC-restricted CTLs.
FIG. 8.

Tetramer staining of CD4−/CD8+ CTLs. For time points shown in Fig. 6, PBMCs were stained with antibodies against CD4 and CD8 and costained with SLA-1*0401 tetramers bound to either an FMDV (red) or Ebola virus (blue) peptide. Histograms of tetramer-positive CTLs from Ad5-FMDV-B (A)- or Ad5-FMDV-T (B)-vaccinated animals are shown. Numbers indicate percent FMDV tetramer-positive cells from the CD8+, CD4− gate and were calculated by subtracting the percentage of cells stained with the negative-control tetramer.
A number of other peptides from the viral capsid precursor protein, P1, were also shown to bind the SLA-1 MHC class I protein. In addition, more peptides were identified that bind SLA-2 of these Yucatan pigs (SLA-2*040201) (L. E. Pedersen, M. Harndahl, M. Nielsen, J. R. Patch, G. Jungersen, S. Buus, and W. T. Golde, unpublished data). We also prepared tetramers from these combinations and tested them on CD8+ T cells from the vaccinated pigs in this study. All of these tetramers were negative for staining cells taken 4 days after boost and all other time points tested. Only the SLA-1*0401/MTAHITVPY tetramer stained cells from the Ad5-FMDV-T vaccinated animals.
DISCUSSION
In the present study, we tested the ability of two nonreplicating adenovirus vectors to generate a CTL cytolytic response in swine to FMDV epitopes. The ability of this vaccine platform to induce the production of neutralizing antibodies to empty FMDV capsids in both swine (30, 31, 35, 43) and cattle (39) is well established. However, its ability to stimulate an antigen-specific CTL response has not been addressed. We hypothesized that the parental adenovirus-based empty capsid vaccine would provide relatively poor stimulation of CTLs because of the efficiency of capsid processing and formation. We reasoned that inhibited processing of FMDV P1 would enhance CTL activation, presumably as a result of enhanced degradation of P1 and subsequent loading of peptides onto class I MHC molecules. Swine were vaccinated with a T cell-directed adenovirus-based vaccine for FMDV (Ad5-FMDV-T), which contained a mutant 3Cpro that processes P1 poorly. The decreased processing of P1 by Ad5-FMDV-T was evident in Western blot analysis, as well as in the titers of virus-neutralizing antibodies in serum.
Whole PBMCs isolated from vaccinated animals demonstrated cytolytic activity against appropriate target cells. However, enriching for CD6+ cells before mixing the effector and target cells improved the analysis of CTL activity. These results are in agreement with those of Denyer and coworkers, who demonstrated improved titration of antigen-specific, MHC-restricted cytolysis by enriching for the CD6+ population of swine T cells (11). The CD6+ cell population has been characterized as consisting of MHC-restricted CTLs and helper T cells, including CD4+/CD8−, CD4−/CD8+, and CD4+/CD8+ populations (8, 11, 41, 42, 49). Thus, an additional advantage of positively selecting for CD6+ cells is the elimination of non-MHC-restricted cytolytic cells such as NK cells and γδ T cells, which do not express CD6 (44).
The MHC restriction of this CTL response was strongly supported by SLA-1 tetramer staining of cells derived from these animals. When peripheral blood cells derived from pigs vaccinated with Ad5-FMDV-T were analyzed, an increase in tetramer+/CD4−/CD8+ cells was detected. The increase in tetramer-positive cells correlated with the peak of cytolytic activity, consistent with MHC-restricted cytolytic activity. We also noted that for pigs vaccinated with Ad5-FMDV-T, the relative tetramer staining of samples taken 4 days after the second boost correlates with the relative level of CTL killing activity. However, given the small number of animals in the group, this correlation is not statistically significant.
The enhanced cytolysis and tetramer staining of cells derived from pigs vaccinated with Ad5-FMDV-T relative to those vaccinated with Ad5-FMDV-B are supportive of our hypothesis that decreased capsid processing enhances CTL activity. However, it is possible that the precursor and mature capsid proteins are cleaved differently by the proteasome, resulting in a lack of MTAHITVPY peptides displayed in MHC molecules of the Ad5-FMDV-B-infected cells. This could explain why few tetramer-positive cells were detected in animals vaccinated with Ad5-FMDV-B. At minimum, this peptide from the C-terminal half of the VP2 region of P1 is not near the cleavage site at the junction of VP2 and VP3. Differential tetramer staining cannot, therefore, be attributed to a loss of this peptide due to precursor cleavage by 3Cpro.
Although CTLs play a central role in the adaptive immune response to intracellular antigens, their role in FMDV immunity has received little attention, in part because of the technical difficulties in establishing appropriate assays with which to measure their activity. In addition to the requirement for genetic matching of MHC class I alleles between effector and target cells, the cytopathic nature of FMDV has hampered development of a cytolytic assay. As a consequence, investigators have measured other biological properties of CD8+ cells, such as proliferation (6, 26, 47, 50) or IFN-γ secretion (22), as proxies for CTL killing activity. The present report describes an alternative strategy for directly assessing CTL killing in response to FMDV antigens and relies on recombinant expression of FMDV proteins. In an earlier report, Li and coworkers described vaccination of swine with an attenuated pseudorabies virus containing a gene for the FMDV P1 capsid precursor that was effective at generating CTL killing activity (29). However, these investigators did not address MHC restriction, characterization of the cells responsible for killing, or antigen specificity of the reported CTL activity. In contrast, we now present data describing the first description of using MHC tetramers to identify antigen-specific CTLs in swine in general and the first time that anti-FMDV CTL killing in any livestock species has been correlated with tetramer staining.
Whether CTLs can play a significant role in FMDV protection and vaccination strategies remains to be determined. Previous work in swine found that a complete lack of capsid processing resulted in a failure to generate neutralizing antibodies, and challenged animals developed clinical signs comparable to those of unvaccinated animals (30, 31). However, the dose of vaccine used was more than a log lower than the dose used here. Reports using higher doses of a P1-based vaccine in swine resulted in partial protection in the absence of FMDV-specific antibodies (50). Similar results were obtained in cattle by the same investigators (52).
In vitro, FMDV inhibition of MHC class I surface expression begins in as little as 30 min postinfection (51). Thus, even under ideal conditions where anti-FMDV CTLs are primed and expanded, control of infection by CTLs depends on host cells infected by wild-type virus in vivo processing and displaying FMDV peptides leading to the CTL response before inhibition of new MHC class I protein expression is blocked by viral proteases. Although neutralizing antibodies are likely to remain the principal focus of FMDV vaccination, CTLs may be able to contribute to cross-serotype and subtype immunity because potential T cell epitopes are not limited to the structural proteins, where most of the immune-driven diversity is located (4). Whereas antibody epitopes are large and three dimensional, the T cell epitopes are linear and extremely small (8 to 12 amino acids) and have the potential to be highly conserved over the different serotypes of FMDV. Further work directed toward refining the induction and measuring of the CTL response, as well as evaluating protection from challenge, is needed to determine if induction of CTL immunity can yield protection across serotypes in diverse breeds of pigs and other species of interest, especially cattle.
Supplementary Material
Acknowledgments
We thank Mary Kenney and Mital Pandya (Plum Island Animal Disease Center, ARS, USDA) for their support and assistance with these studies. We also thank staff of the Animal Resources Unit (Plum Island Animal Disease Center, DHS) for their professional work with the animals used in these experiments.
This work was funded by CRIS grant no. 1940-32000-052-00D from the Agricultural Research Service, USDA (W.T.G.). In addition, this work was partially cofunded by the Danish Council for Independent Research, Technology and Production Sciences grant no. 274-09-0281 (G.J.). L. E. Pedersen was the recipient of a Plum Island Animal Disease Center Research Participation Program fellowship, administered by the Oak Ridge Institute for Science and Education (ORISE) through an interagency agreement between the U.S. Department of Energy (DOE) and the U.S. Department of Agriculture (USDA).
Footnotes
Published ahead of print on 22 December 2010.
Supplemental material for this article may be found at http://cvi.asm.org/.
REFERENCES
- 1.Bautista, E. M., G. S. Ferman, and W. T. Golde. 2003. Induction of lymphopenia and inhibition of T cell function during acute infection of swine with foot and mouth disease virus (FMDV). Vet. Immunol. Immunopathol. 92:61-73. [DOI] [PubMed] [Google Scholar]
- 2.Belsham, G. J., and J. K. Brangwyn. 1990. A region of the 5′ noncoding region of foot-and-mouth disease virus RNA directs efficient internal initiation of protein synthesis within cells: involvement with the role of L protease in translational control. J. Virol. 64:5389-5395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Belsham, G. J., G. M. McInerney, and N. Ross-Smith. 2000. Foot-and-mouth disease virus 3C protease induces cleavage of translation initiation factors eIF4A and eIF4G within infected cells. J. Virol. 74:272-280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Carrillo, C., et al. 2005. Comparative genomics of foot-and-mouth disease virus. J. Virol. 79:6487-6504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chavez-Galan, L., M. C. Arenas-Del Angel, E. Zenteno, R. Chavez, and R. Lascurain. 2009. Cell death mechanisms induced by cytotoxic lymphocytes. Cell Mol. Immunol. 6:15-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Childerstone, A., L. Cedillo-Baron, M. Foster-Cuevas, and R. Parkhouse. 1999. Demonstration of bovine CD8+ T-cell responses to foot-and-mouth disease virus. J. Gen. Virol. 80:663-669. [DOI] [PubMed] [Google Scholar]
- 7.Chinsangaram, J., M. Koster, and M. J. Grubman. 2001. Inhibition of L-deleted foot-and-mouth disease virus replication by alpha/beta interferon involves double-stranded RNA-dependent protein kinase. J. Virol. 75:5498-5503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.de Bruin, T. G., et al. 2000. Discrimination of different subsets of cytolytic cells in pseudorabies virus-immune and naive pigs. J. Gen. Virol. 81:1529-1537. [DOI] [PubMed] [Google Scholar]
- 9.de Los Santos, T., S. de Avila Botton, R. Weiblen, and M. J. Grubman. 2006. The leader proteinase of foot-and-mouth disease virus inhibits the induction of beta interferon mRNA and blocks the host innate immune response. J. Virol. 80:1906-1914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.de Los Santos, T., F. Diaz-San Segundo, and M. J. Grubman. 2007. Degradation of nuclear factor kappa B during foot-and-mouth disease virus infection. J. Virol. 81:12803-12815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Denyer, M. S., T. E. Wileman, C. M. Stirling, B. Zuber, and H. H. Takamatsu. 2006. Perforin expression can define CD8 positive lymphocyte subsets in pigs allowing phenotypic and functional analysis of natural killer, cytotoxic T, natural killer T and MHC un-restricted cytotoxic T-cells. Vet. Immunol. Immunopathol. 110:279-292. [DOI] [PubMed] [Google Scholar]
- 12.Devaney, M. A., V. N. Vakharia, R. E. Lloyd, E. Ehrenfeld, and M. J. Grubman. 1988. Leader protein of foot-and-mouth disease virus is required for cleavage of the p220 component of the cap-binding protein complex. J. Virol. 62:4407-4409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Diaz-San Segundo, F., et al. 2006. Selective lymphocyte depletion during the early stage of the immune response to foot-and-mouth disease virus infection in swine. J. Virol. 80:2369-2379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Doel, T. R., L. Williams, and P. V. Barnett. 1994. Emergency vaccination against foot-and-mouth disease: rate of development of immunity and its implications for the carrier state. Vaccine 12:592-600. [DOI] [PubMed] [Google Scholar]
- 15.Falk, M. M., et al. 1990. Foot-and-mouth disease virus protease 3C induces specific proteolytic cleavage of host cell histone H3. J. Virol. 64:748-756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Golde, W. T., C. K. Nfon, and F. N. Toka. 2008. Immune evasion during foot-and-mouth disease virus infection of swine. Immunol. Rev. 225:85-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Golde, W. T., et al. 2005. Vaccination against foot-and-mouth disease virus confers complete clinical protection in 7 days and partial protection in 4 days: use in emergency outbreak response. Vaccine 23:5775-5782. [DOI] [PubMed] [Google Scholar]
- 18.Grubman, M. J., and B. Baxt. 2004. Foot-and-mouth disease. Clin. Microbiol. Rev. 17:465-493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Grubman, M. J., M. P. Moraes, F. Diaz-San Segundo, L. Pena, and T. de los Santos. 2008. Evading the host immune response: how foot-and-mouth disease virus has become an effective pathogen. FEMS Immunol. Med. Microbiol. 53:8-17. [DOI] [PubMed] [Google Scholar]
- 20.Grubman, M. J., et al. 2010. Adenovirus serotype 5-vectored foot-and-mouth disease subunit vaccines: the first decade. Future Virol. 5:51-64. [Google Scholar]
- 21.Grubman, M. J., M. Zellner, G. Bablanian, P. W. Mason, and M. E. Piccone. 1995. Identification of the active-site residues of the 3C proteinase of foot-and-mouth disease virus. Virology 213:581-589. [DOI] [PubMed] [Google Scholar]
- 22.Guzman, E., G. Taylor, B. Charleston, M. A. Skinner, and S. A. Ellis. 2008. An MHC-restricted CD8+ T-cell response is induced in cattle by foot-and-mouth disease virus (FMDV) infection and also following vaccination with inactivated FMDV. J. Gen. Virol. 89:667-675. [DOI] [PubMed] [Google Scholar]
- 23.Ho, C. S., M. H. Franzo-Romain, Y. J. Lee, J. H. Lee, and D. M. Smith. 2009. Sequence-based characterization of swine leucocyte antigen alleles in commercially available porcine cell lines. Int. J. Immunogenet. 36:231-234. [DOI] [PubMed] [Google Scholar]
- 24.Ho, C. S., et al. 2009. Nomenclature for factors of the SLA system, update 2008. Tissue Antigens 73:307-315. [DOI] [PubMed] [Google Scholar]
- 25.Hoof, I., et al. 2009. NetMHCpan, a method for MHC class I binding prediction beyond humans. Immunogenetics 61:1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Joshi, G., R. Sharma, and N. K. Kakker. 2009. Phenotypic and functional characterization of T-cells and in vitro replication of FMDV serotypes in bovine lymphocytes. Vaccine 27:6656-6661. [DOI] [PubMed] [Google Scholar]
- 27.Kuhn, R., N. Luz, and E. Beck. 1990. Functional analysis of the internal translation initiation site of foot-and-mouth disease virus. J. Virol. 64:4625-4631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Leisner, C., et al. 2008. One-pot, mix-and-read peptide-MHC tetramers. PLoS One 3:e1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li, X., et al. 2008. Induction of protective immunity in swine by immunization with live attenuated recombinant pseudorabies virus expressing the capsid precursor encoding regions of foot-and-mouth disease virus. Vaccine 26:2714-2722. [DOI] [PubMed] [Google Scholar]
- 30.Mayr, G. A., J. Chinsangaram, and M. J. Grubman. 1999. Development of replication-defective adenovirus serotype 5 containing the capsid and 3C protease coding regions of foot-and-mouth disease virus as a vaccine candidate. Virology 263:496-506. [DOI] [PubMed] [Google Scholar]
- 31.Mayr, G. A., V. O'Donnell, J. Chinsangaram, P. W. Mason, and M. J. Grubman. 2001. Immune responses and protection against foot-and-mouth disease virus (FMDV) challenge in swine vaccinated with adenovirus-FMDV constructs. Vaccine 19:2152-2162. [DOI] [PubMed] [Google Scholar]
- 32.McCullough, K. C., et al. 1992. Relationship between the anti-FMD virus antibody reaction as measured by different assays, and protection in vivo against challenge infection. Vet. Microbiol. 30:99-112. [DOI] [PubMed] [Google Scholar]
- 33.Moffat, K., et al. 2005. Effects of foot-and-mouth disease virus nonstructural proteins on the structure and function of the early secretory pathway: 2BC but not 3A blocks endoplasmic reticulum-to-Golgi transport. J. Virol. 79:4382-4395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Moffat, K., et al. 2007. Inhibition of the secretory pathway by foot-and-mouth disease virus 2BC protein is reproduced by coexpression of 2B with 2C, and the site of inhibition is determined by the subcellular location of 2C. J. Virol. 81:1129-1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Moraes, M. P., G. A. Mayr, P. W. Mason, and M. J. Grubman. 2002. Early protection against homologous challenge after a single dose of replication-defective human adenovirus type 5 expressing capsid proteins of foot-and-mouth disease virus (FMDV) strain A24. Vaccine 20:1631-1639. [DOI] [PubMed] [Google Scholar]
- 36.Nfon, C. K., G. S. Ferman, F. N. Toka, D. A. Gregg, and W. T. Golde. 2008. Interferon-alpha production by swine dendritic cells is inhibited during acute infection with foot-and-mouth disease virus. Viral Immunol. 21:68-77. [DOI] [PubMed] [Google Scholar]
- 37.Nfon, C. K., F. N. Toka, M. Kenney, J. M. Pacheco, and W. T. Golde. 2010. Loss of plasmacytoid dendritic cell function coincides with lymphopenia and viremia during foot-and-mouth disease virus infection. Viral Immunol. 23:29-41. [DOI] [PubMed] [Google Scholar]
- 38.Nielsen, M., et al. 2007. NetMHCpan, a method for quantitative predictions of peptide binding to any HLA-A and -B locus protein of known sequence. PLoS One 2:e796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pacheco, J. M., M. C. Brum, M. P. Moraes, W. T. Golde, and M. J. Grubman. 2005. Rapid protection of cattle from direct challenge with foot-and-mouth disease virus (FMDV) by a single inoculation with an adenovirus-vectored FMDV subunit vaccine. Virology 337:205-209. [DOI] [PubMed] [Google Scholar]
- 40.Pacheco, J. M., et al. 2010. IgA antibody response of swine to foot-and-mouth disease virus infection and vaccination. Clin. Vaccine Immunol. 17:550-558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pauly, T., et al. 1995. Classical swine fever virus-specific cytotoxic T lymphocytes and identification of a T cell epitope. J. Gen. Virol. 76(pt. 12):3039-3049. [DOI] [PubMed] [Google Scholar]
- 42.Pauly, T., et al. 1996. Differentiation between MHC-restricted and non-MHC-restricted porcine cytolytic T lymphocytes. Immunology 88:238-246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pena, L., et al. 2008. Delivery of a foot-and-mouth disease virus empty capsid subunit antigen with nonstructural protein 2B improves protection of swine. Vaccine 26:5689-5699. [DOI] [PubMed] [Google Scholar]
- 44.Piriou-Guzylack, L., and H. Salmon. 2008. Membrane markers of the immune cells in swine: an update. Vet. Res. 39:54. [DOI] [PubMed] [Google Scholar]
- 45.Robertson, B. H., D. O. Morgan, and D. M. Moore. 1984. Location of neutralizing epitopes defined by monoclonal antibodies generated against the outer capsid polypeptide, VP1, of foot-and-mouth disease virus A12. Virus Res. 1:489-500. [DOI] [PubMed] [Google Scholar]
- 46.Rock, K. L., D. J. Farfan-Arribas, and L. Shen. 2010. Proteases in MHC class I presentation and cross-presentation. J. Immunol. 184:9-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rodriguez, A., et al. 1996. A porcine CD8+ T cell clone with heterotypic specificity for foot-and-mouth disease virus. J. Gen. Virol. 77(pt. 9):2089-2096. [DOI] [PubMed] [Google Scholar]
- 48.Rodriguez, L. L., and M. J. Grubman. 2009. Foot and mouth disease virus vaccines. Vaccine 27(suppl. 4):D90-D94. [DOI] [PubMed] [Google Scholar]
- 49.Saalmuller, A., T. Pauly, B. J. Hohlich, and E. Pfaff. 1999. Characterization of porcine T lymphocytes and their immune response against viral antigens. J. Biotechnol. 73:223-233. [DOI] [PubMed] [Google Scholar]
- 50.Sanz-Parra, A., et al. 1999. Recombinant viruses expressing the foot-and-mouth disease virus capsid precursor polypeptide (P1) induce cellular but not humoral antiviral immunity and partial protection in pigs. Virology 259:129-134. [DOI] [PubMed] [Google Scholar]
- 51.Sanz-Parra, A., F. Sobrino, and V. Ley. 1998. Infection with foot-and-mouth disease virus results in a rapid reduction of MHC class I surface expression. J. Gen. Virol. 79(pt. 3):433-436. [DOI] [PubMed] [Google Scholar]
- 52.Sanz-Parra, A., et al. 1999. Evidence of partial protection against foot-and-mouth disease in cattle immunized with a recombinant adenovirus vector expressing the precursor polypeptide (P1) of foot-and-mouth disease virus capsid proteins. J. Gen. Virol. 80(pt. 3):671-679. [DOI] [PubMed] [Google Scholar]
- 53.Stanway, G., et al. 2005. Virus taxonomy: classification and nomenclature of viruses: eighth report of the International Committee on the Taxonomy of Viruses. Elsevier Academic Press, San Diego, CA.
- 54.Tesar, M., and O. Marquardt. 1990. Foot-and-mouth disease virus protease 3C inhibits cellular transcription and mediates cleavage of histone H3. Virology 174:364-374. [DOI] [PubMed] [Google Scholar]
- 55.Toka, F. N., C. Nfon, H. Dawson, and W. T. Golde. 2009. Natural killer cell dysfunction during acute infection with foot-and-mouth disease virus. Clin. Vaccine Immunol. 16:1738-1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Toka, F. N., C. K. Nfon, H. Dawson, D. M. Estes, and W. T. Golde. 2009. Activation of porcine natural killer cells and lysis of foot-and-mouth disease virus infected cells. J. Interferon Cytokine Res. 29:179-192. [DOI] [PubMed] [Google Scholar]
- 57.Yang, M., A. Clavijo, R. Suarez-Banmann, and R. Avalo. 2007. Production and characterization of two serotype independent monoclonal antibodies against foot-and-mouth disease virus. Vet. Immunol. Immunopathol. 115:126-134. [DOI] [PubMed] [Google Scholar]
- 58.Yewdell, J. W., and S. M. Haeryfar. 2005. Understanding presentation of viral antigens to CD8+ T cells in vivo: the key to rational vaccine design. Annu. Rev. Immunol. 23:651-682. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.