

Abstract
In order to explore the possible role of E-cadherin in familial cancer, 19 familial breast cancer patients, whose tumours demonstrated loss of heterozygosity (LOH) at the E-cadherin locus, were screened for germline mutations. No pathogenic germline alterations were detected in these individuals. However, a somatic mutation was found (49-2A→C) in one of the tumours. This tumour showed a pattern of both ductal and lobular histology. Another 10 families with cases of breast, gastric and colon cancer were also screened for germline mutations, and no mutations were found. A missense mutation in exon 12 of E-cadherin (1774G→A; Ala592Thr) was previously found in one family with diffuse gastric cancer, and colon and breast cancer. An allelic association study was performed to determine whether the Ala592Thr alteration predisposes to breast cancer. In total, we studied 484 familial breast cancer patients, 614 sporadic breast cancer patients and 497 control individuals. The frequencies of this alteration were similar in these groups. However, a correlation between the Ala592Thr alteration and ductal comedo-type tumour was seen. These results, together with previously reported studies, indicate that germline mutations and, more commonly, somatic mutations in E-cadherin may have an influence on the behaviour of the tumours, rather than predispose to breast cancer.
Keywords: breast cancer, ductal comedo-type, E-cadherin, familial, lobular
Synopsis
Introduction:
E-cadherin (CDH1) is expressed on the cell surface in most epithelial tissues [1,2], and evidence is rapidly accumulating that the mammalian E-cadherin gene product plays a role in epithelial tumourigenesis [3]. Germline mutations in E-cadherin have been described in families with early-onset diffuse gastric cancer, and loss of function of this gene has been implicated in the pathogenesis of early-onset colorectal and breast cancers. Despite the fact that E-cadherin expression is decreased in 50% of invasive ductal carcinoma [4], mutations in the coding sequence of E-cadherin have not been observed in this type of breast cancer [5,6]. The role of E-cadherin mutation in development of hereditary gastric cancer has been shown, but its role in predisposing to breast cancer is still unproved. In order to further explore the involvement of E-cadherin in breast cancer, we examined all 16 exons of E-cadherin in 31 familial breast cancer patients, in whom BRCA1 and BRCA2 had been excluded as predisposing genes [7,8,9].
In a previous study of a family with diffuse gastric cancer (Fig. 1a) (Salahshor S, et al, manuscript submitted), we identified an E-cadherin germline mutation that cosegregated with the disease. This missense mutation in exon 12 (Ala592Thr) was also detected in the index patient's mother, who had ductal breast cancer. In an attempt to clarify a possible role for the Ala592Thr alteration in predisposing to breast cancer, we screened for this specific alteration in different series of breast cancer patients and control individuals. In total, 1328 patients with sporadic or familial breast cancer and 497 control individuals were analyzed for this specific alteration.
Figure 1.
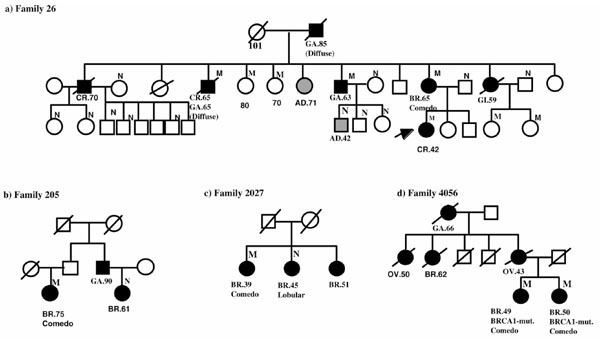
Pedigrees of the four families with Ala592Thr missense alteration. BR, breast cancer; GA, gastric cancer; OV, ovarian cancer; GI, gastrointestinal cancer; AD, colonic adenomas; Endo, endometrial cancer; M, positive for the Ala592Thr variant; N, wild-type E-cadherin.
Materials and method:
Nineteen probands with familial breast cancer who demonstrated LOH at chromosome 16q, and 12 patients from 10 families with breast, gastric and colorectal cancers were screened for germline mutations in the coding sequences of E-cadherin. Nine tumours from the 19 breast cancer cases were also screened for somatic mutations. The frequencies of the 1774G→A variant (Ala592Thr) were determined in DNA extracted from the blood of 358 unrelated probands with familial breast cancer, 214 unrelated early-onset breast cancer patients (age of onset <41 years), 126 unrelated BRCA1 or BRCA2 carriers, and 604 unselected breast cancer patients and 497 control individuals, who were considered to represent the general population in Sweden and Norway.
Two markers D16S7/p79-2-23 and APRT/HUAP15, which both map to 16q24.3, were used to identify LOH, as in a previous study of allelic loss at 16q in familial tumours [10].
Single-stranded conformation polymorphism (SSCP) and denaturing high-performance liquid chromotography (DHPLC; Transgenomic, Santa Clara, CA, USA) were used to screen 16 exons of the E-cadherin gene for the presence of alterations (Table 1). We used the same primers as described by Berx et al [5] and Salahshor et al (manuscript submitted), but with some modification. Samples that exhibited aberrant bands on the SSCP gel or an altered DHPLC pattern were sequenced by reamplifying the corresponding exon. Sequencing was performed either manually using the ThermoSequensase (Amersham Pharmacia Biotech, Uppsala, Sweden) kit, or by 377 ABI automated sequencer using the Taq-Cycle sequencing BigDye Terminator kit (Applied Biosystems Inc ABI, Stockholm, Sweden), according to conditions recommended by the manufacturers. The Ala592Thr variant frequency was detected using specific polymerase chain reaction (PCR)-restriction fragment length polymorphism tests (Salahshor S, et al, manuscript submitted).
Table 1.
PCR and DHPLC conditions for the mutational analysis of E-cadherin
| Annealing | Amplicon | DHPLC | |
| temperature | length | temperature | |
| Exon no | (°C) | (bp) | (°C) |
| 1 | 70 | 285 | 66 |
| 2 | 55 | 377 | 62 |
| 3 | 60 | 352 | 58, 61 |
| 4 | 60 | 351 | 60 |
| 5 | 60 | 398 | 58 |
| 6 | 58 | 246 | 59 |
| 7 | 60 | 329 | 60, 62, 64 |
| 8 | 58 | 225 | 59, 62 |
| 9 | 55 | 252 | 59 |
| 10 | 55 | 309 | 58 |
| 11 | 60 | 243 | 61 |
| 12 | 60 | 326 | 59, 61 |
| 13 | 58 | 297 | 58 |
| 14 | 60 | 206 | 61 |
| 15 | 60 | 244 | 58 |
| 16 | 60 | 318 | 59 |
Results:
No pathogenic mutations were identified in the 12 patients from families with documented breast, gastric or colon cancer. We also searched for germline alterations in 19 individuals with familial breast cancer who showed LOH at the E-cadherin locus in their tumours. Nine tumours from these 19 patients were also tested for somatic alterations in the E-cadherin gene. One somatic mutation (49-2A→C) was found in one of the lobular cancer cases (Fig. 2). One previously reported common polymorphism in E-cadherin [11] was found in exon 13 at codon 692 (GCC→GCT).
Figure 2.
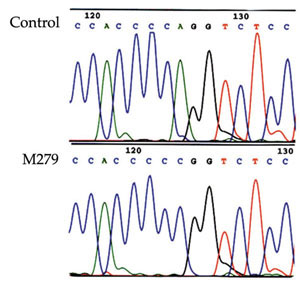
The 3' splice site alteration in the E-cadherin gene (49-2A→C) in tumor M279.
In a previous study in one family with familial diffuse gastric cancer and colon cancer (Fig. 1a) (Salahshor S, et al, manuscript submitted), we found a germline missense mutation in exon 12 (Ala592Thr) of E-cadherin. This alteration cosegregated with diffuse gastric cancer and colorectal cancer in this family, although the penetrance was not complete. The alteration was present in the index patient's mother, who had ductal breast cancer of comedo type at the age of 65 years (Fig. 1a, II:12). In order to investigate whether this alteration was associated with an increased risk for breast cancer, we used a specific restriction enzyme digestion/PCR test to detect the variant, in an allelic association study in familial and sporadic breast cancer cases, as well as control samples that represented the general population. We found the mutation in two out of 358 (0.56%) non-BRCA1/BRCA2 carrier women with familial breast cancer (families 205 and 2027). In family 205 (Fig. 1b), one woman with ductal comedo-type breast cancer at age 75 years had the Ala592Thr variant. The other available case from this family (Figure 1b, II:2) tested negative, and the histology report was not available. In family 2027 (Fig. 1c), one woman with a breast cancer of ductal comedo type at age 37 years showed the Ala592Thr variant. Her two sisters, who had breast cancer at ages 45 and 51 years, did not. One of the sisters had lobular breast cancer, whereas the histopathological information from the third case was unavailable for study. Because the variant did not segregate with the disease, it is not likely that this alteration predisposes to breast cancer in either of these two families.
We also screened 126 BRCA1 or BRCA2 carriers from different families, and found the mutation in one breast cancer patient (family 4056) with a BRCA1 germline mutation (delC2594). It was possible to obtain a sample from her sister who also carried the BRCA1 mutation, and she was found to share the Ala592Thr variant (Fig. 1d). Both tumours were of ductal comedo type. Because both of these tumours were caused by a germline BRCA1 mutation, the E-cadherin variant was not likely to have predisposed to breast cancer. Among the 604 sporadic breast cancer cases, five were found to carry the Ala592Thr alteration (0.83%; Table 2). One of the tumours was of lobular type, and the other four were of ductal type. One of the ductal tumours was of comedo type, and the tumour type of the other three was not known. Four cases with Ala592Thr alteration were also found among the 497 control individuals (0.80%). The alteration was not found among any of the 214 early-onset breast cancer patients.
Table 2.
Frequency of Ala592Thr variant in various populations of breast cancer patients and control population
| No of families/ | Frequency of | |||
| Screened population | no of cases | Ala592Thr variant | Case id | Histopathology |
| Familial breast cancer | ||||
| Non-BRCA1/BRCA2 mutation carrier | 358 | 0.56% | 311-89D | Ductal comedo |
| AL-155 | Ductal comedo | |||
| BRCA1/BRCA2 carrier | 126 | 0.79% | BR-115 | Ductal comedo |
| BR-119 | Ductal comedo | |||
| Sporadic breast cancer | ||||
| Early-onset breast cancer | 214 | - | ||
| Unselected breast cancer | 604 | 0.83% | 17-S | Lobular |
| MB-181 | Ductal comedo | |||
| MB-483 | Ductal | |||
| ULL-17 | Ductal | |||
| ULL-284 | Ductal | |||
| Control population | 495 | 0.80% |
Discussion:
So far, E-cadherin germline mutations have been reported in 17 diffuse gastric cancer families [12,13,14,15,16]. The frequency of E-cadherin germline mutations in breast cancer reported thus far is low [17,18]. Mutations reported in E-cadherin in lobular breast cancer patients are in most cases tumour restricted, and not germline alterations [5,19]. In the present study, we did not find any pathogenic alteration in 10 families that included patients with breast, gastric or colon cancer. We conclude that, although germline E-cadherin mutations are sometimes found in familial gastric and colon cancer, they are not frequently involved in families in which gastric cancer appears to segregate as a part of an inherited predisposition to primary breast cancer.
The frequency of Ala592Thr alteration, which was first identified in one family with familial diffuse gastric cancer plus one case of colon and one case of breast cancer (Fig. 1a), was similar in the various groups studied here (Table 2). This finding does not support an effect of this alteration in predisposing to breast cancer in general. Thus, available data indicate that germline E-cadherin mutations do not constitute a major risk factor for breast cancer. However, other studies have indicated that somatic alterations could have an impact on the phenotype in lobular breast cancer. The fact that many tumours with the alteration (Ala592Thr) in the present study were of the ductal comedo type may indicate a genetic basis for the phenotypic divergence caused by this germline E-cadherin alteration. Although lobular breast carcinoma and ductal breast carcinoma of comedo type are different from each other histologically, neither of these cancers are associated with good prognosis. However, individual prognosis with breast cancer is assessed mainly by grading rather than by histological type [20,21].
In summary, the present study, together with previously reported data, suggests that a germline mutation in E-cadherin is not a major risk factor for breast cancer. However, germline, and more often somatic mutations in this gene could have an impact on phenotypic divergence and prognosis, including growth pattern of the tumours, such as in lobular and perhaps ductal comedo-type breast cancer. In addition, other genetic alterations or epigenetic changes in the E-cadherin gene may have an impact on the metastatic behaviour of the cancer cells, and thereby on the clinical outcome.
Introduction
E-cadherin (CDH1) is expressed on the cell surface in most epithelial tissues [1,2], and evidence is rapidly accumulating that the mammalian E-cadherin gene product plays a role in epithelial tumourigenesis [3]. Loss of function in E-cadherin and/or other collaborating proteins contributes to increased proliferation, invasion and metastasis in many malignant epithelial tumours [22,23]. Mutations in E-cadherin, which encodes a transmembrane protein, have been described in cancers of the endometrium and ovary [11], signet ring cell carcinoma of the stomach [24,25], the diffuse sclerosing variant of papillary thyroid carcinoma [26], invasive lobular breast cancer [5,19,27,28], and diffuse and mixed gastric cancer [24,28,29,30]. E-cadherin germline mutations have been found in early-onset hereditary diffuse gastric cancer [12,14,16], and such mutations have even been implicated as risk factors for early-onset breast and colon cancers [13,15]. Inactivating mutations and decreased expression of E-cadherin have been reported in invasive lobular breast carcinomas [5,19], which demonstrate involvement of E-cadherin alteration in sporadic lobular breast cancer. Lobular carcinoma in situ is known to be a risk factor for breast cancer, but thus far constitutional E-cadherin mutations have not been identified in patients with lobular carcinoma in situ [18]. In sporadic breast cancer, a correlation between loss of E-cadherin expression and metastatic behaviour has been reported [4,31,32,33]. Despite the fact that E-cadherin expression is decreased in 50% of invasive ductal carcinomas [4], mutations in the coding sequence of E-cadherin have not been observed in this type of breast cancer [5,6].
A role of E-cadherin mutation in development of hereditary gastric cancer has been shown, but its role in predisposing to breast cancer is still unproved. In order to further explore the involvement of E-cadherin in breast cancer, we examined all 16 exons of E-cadherin in 31 familial breast cancer patients in whom involvement of BRCA1 and BRCA2 had been excluded [7,8,9]. Twelve of these breast cancer patients (from 10 families) had, besides a family history of breast cancer, a family history of gastric or colorectal cancer. The remaining 19 familial breast cancer patients were identified as having LOH at 16q in their tumour [34]. LOH at 16q occurs frequently in sporadic [35] and in familial breast cancer [10]. It has also been shown to correlate with distant metastasis [34,36]. In the familial breast cancer patients in whom LOH at 16q was identified, E-cadherin was suggested to be a candidate predisposing tumour suppressor gene [34], and the aim of the present study was to elucidate this relation.
In a previous study in a family with diffuse gastric and colon cancer (Salahshor S, et al, manuscript submitted), we found an E-cadherin germline mutation that cosegregated with the disease. This missense mutation in exon 12 (Ala592Thr) was also detected in the index patient's mother, who had ductal breast cancer. In an attempt to clarify a possible role of the Ala592Thr alteration in predisposing to breast cancer, we screened for this specific alteration in different series of breast cancer patients and control individuals. In total, 1328 patients with sporadic or familial breast cancer and 497 control individuals were tested for this specific alteration.
Materials and method
Cases
Cases screened for mutations in the E-cadherin gene
Nineteen women with breast cancer who exhibited LOH at 16q were included in the present study. All of these index patients had a positive family history (more than one first- or second-degree relative with breast cancer). Ten of the 19 cases were of ductal histology, one case was both ductal and lobular, two cases were lobular only and one case was medullar. For five other cases information regarding histology was not available [10,34]. Nine tumours (four ductal, two lobular, one with both ductal and lobular, and two cases with unknown histology) from these 19 patients were also tested for somatic mutations in E-cadherin.
In order to examine whether germline mutations in E-cadherin are involved in familial breast, gastric and colon cancer, 12 patients from 10 such families were also included in the present study. Six of those 12 cases had breast cancer, five had colon cancer, and one was affected with both breast and colon cancer. The histology of the breast and gastric cancers in this group was not available for study.
Individuals genotyped for the Ala592Thr variant
The frequencies of Ala592Thr variant were determined in DNA extracted from blood of 358 probands with familial breast cancer, 214 unrelated early-onset breast cancer patients (age of onset <41 years), 126 unrelated BRCA1 or BRCA2 carriers, and 604 unselected breast cancer patients from consecutive series. In addition 497 control patients were genotyped, who were considered to represent the general populations of Sweden and Norway. The Norwegian control individuals (a total of 244) included 177 healthy female individuals obtained through the Norwegian Population Registry as a population-based series of women aged 20-44 years from the Oslo area, and 67 healthy women (aged 55-79 years) from a mammographic screening programme from Bergen area. The Swedish control individuals (a total of 253) were healthy relatives of individuals who were counseled at the Karolinska hospital.
In the present study, 'familial breast cancer cases' are defined as patients with more than one first- or second-degree relative with breast cancer, regardless of age. All familial cases had already been tested for germline mutations in the BRCA1 and BRCA2 genes.
Determination of loss of heterozygosity
In the previous study of allelic loss at 16q in familial tumours [10] the two markers D16S7/p79-2-23 and APRT/HUAP15, which map to 16q24.3, were used. Both markers are distal to E-cadherin.
Mutation analysis
Polymerase chain reaction/single-stranded conformation polymorphism
SSCP was used to screen 16 exons of the E-cadherin gene for the presence of alterations, as described by Berx et al [5] and Salahshor et al (manuscript submitted), but with some modification.
Denaturing high-performance liquid chromatography
DHPLC was carried out on automated instrumentation equipped with a DNA-separation column (Wave® system; Transgenomic). PCR and Taq Gold™ enzyme (Applied Biosystems Inc ABI) were used to amplify all 16 exons of E-cadherin (96°C, 15 min; 94°C, 30 s; 50-70°C, 30 s; 72°C, 45 s for 30 cycles; 72°C, 2 min). The primers used were as described previously [5] (Salahshor S, et al, manuscript submitted). Heteroduplexes and homoduplexes were formed when the PCR products were hybridized by heating to 95°C (for 5 min) and cooling slowly (by 1.5°C/min, 47 cycles). Five microlitres of each PCR product were then analyzed using the Wave® system (Transgenomic). DNA was eluted with a mixture of bufferA (0.1 mol/l triethylamine acetate) and buffer B (0.1 mol/l triethylamine acetate, 25% acetonitrile) at a flow rate of 0.9 ml/min. The mobile-phase gradients and the temperatures for running each exon were suggested by the Wave-Maker software (Transgenomic), based on the sequence contents. The temperatures at which PCR products were analyzed are given in Table 1. The results were analyzed using HSM7000 software (Transgenomic).
Sequencing
Samples with an aberrant band on the SSCP gel or an altered DHPLC pattern were sequenced by reamplifying the corresponding exon. Sequencing was performed either manually using the ThermoSequensase (Amersham Corp) kit, or with the 377 ABI automated sequencer using the Taq-Cycle sequencing BigDye Terminator kit (Applied Biosystems Inc) according to the standard conditions recommended by the manufacturer.
Restriction enzyme digestion
The Ala592Thr variant was detected using a specific PCR-restriction fragment length polymorphism test. In short, two pairs of primers, E-CAD12F (TGTCTGATGT-GAATGACAGC) and E-CAD12R (TGCTGTGAAGGGAGATGTAT) were used for amplification of nucleotides 1574-1884 of E-cadherin (based on GenBank accession number Z13009). The forward primer generates an in vitro mutation at nucleotide 1772 (A→G), leading to an Hha I restriction site in the PCR product. If the endogenous mutation 1774 (G→A) is present, then the Hha I restriction endonuclease site is lost. Mutated samples are distinguished from normal samples by a 20-bp longer uncleaved exon 12 fragment on agarose gels (Salahshor S, et al, manuscript submitted).
Results
Screening for E-cadherin germline alterations in familial breast cancer
The entire coding region of the E-cadherin gene was studied, including splice sites, using PCR/SSCP and sequencing in 10 families with histories of breast, gastric and colon cancer. No pathogenic germline mutation was identified in any of the individuals screened.
Negative results were also obtained by screening for germline alterations in 19 individuals with familial breast cancer who exhibited LOH at the E-cadherin locus in their tumours. We also searched for somatic alterations in nine of these 19 cases in whom tumours were available for analysis using Wave® (Transgenomic), an HLPC-based nucleic acid fragment analysis system. One somatic mutation in an exon 2 splicing site (49-2A→C) was detected in the tumour that was of both ductal and lobular histology (Fig. 2). Clinical outcomes of these 19 patients were available, with 5-12 years of follow up. The two lobular breast cancer patients had both developed distant metastases, whereas distant metastases had developed in five out of 10 patients with ductal breast cancer. The patient with ductal-lobular histology and the mutation did not have distant metastasis 8 years after surgery. One previously reported common polymorphism in E-cadherin [11] was found in exon13 at codon 692 (GCC→GCT).
The frequency of the Ala592Tyr missense alteration
In a previous study of one family (family 26) with familial colon and gastric cancer (Fig. 1a) (Salahshor S, et al, manuscript submitted), we identified a germline missense alteration in exon12 (Ala592Thr) of E-cadherin. This alteration cosegregated with diffuse gastric cancer and colorectal cancer in this family, although the penetrance was not complete. The alteration was present in the index patient's mother, who had ductal breast cancer of comedo type at age 65 years (Figure 1a, II:12). In order to investigate whether this alteration was associated with an increased breast cancer risk we used a specific restriction enzyme digestion/PCR test to detect the variant, in an allelic association study of familial and sporadic breast cancer patients as well as control individuals who were considered to represent the general population.
We found the mutation in two out of 358 (0.56%) non-BRCA1/BRCA2 carrier women with familial breast cancer (families 205 and 2027). In family 205 (Fig. 1b), one woman with ductal comedo-type breast cancer at age 75 years had the Ala592Thr variant. The other available case from this family (Fig. 1b, II:2) tested negative, and the histology report was not available. In family 2027 (Fig. 1c), one woman with a breast cancer of ductal comedo type at age 37 years exhibited the Ala592Thr variant, whereas her two sisters with breast cancer at ages 45 and 51 years did not. One of the sisters had lobular breast cancer, but histopathological information on the third case was not available. Because the variant did not segregate with the disease, it is not likely that this alteration predisposes to breast cancer in either of these two families.
We also screened 126 BRCA1 or BRCA2 carriers from different families, and identified the mutation in one patient with breast cancer (family 4056) with a BRCA1 germline mutation (2594delC). It was possible to obtain a sample from her sister who also carried the BRCA1 mutation, and she was found to share the Ala592Thr variant also (Fig. 1d). Both tumours were of ductal comedo type. Because both of these tumours were caused by a germline BRCA1 mutation, the E-cadherin variant was not likely to have predisposed to breast cancer.
Among the 604 sporadic breast cancer patients, five were found to carry the Ala592Thr alteration (0.83%; Table 2). One of these was of lobular type and the other four were of ductal type. One of the ductal tumours was of comedo type, but it was not known for the other three whether they were of comedo type. Four cases with Ala592Thr alteration were also found among the 497 control patients (0.80%). The alteration was not found among any of the 214 early-onset breast cancer patients.
Discussion
E-cadherin germline mutations in familial breast cancer
Thus far, E-cadherin germline mutations have been reported in 17 diffuse gastric cancer families [12,13,14,15,16]. The high rate of mutations found in the Maori families probably reflects the criteria used for selection of the families. In these families at least three first-degree relatives were affected by gastric cancer, and the age of onset of at least one case was under 40 years. In other studies, a low frequency of E-cadherin germline mutations in familial gastric cancer has been reported [37,38]. The frequency of E-cadherin germline mutations in breast cancer reported thus far is low [17,18]. Mutations in E-cadherin reported in lobular breast cancer cases are in most cases tumour restricted, and are not germline alterations [5,19]. In the 17 gastric cancer families with germline mutations, five members from three families were also affected by breast cancer [13,17]. Three of these patients from three different families had both breast and gastric cancer. One patient had developed both lobular breast cancer and diffuse-type gastric cancer [17]. In the other cases the gastric cancers were of diffuse type, and the histology of the breast tumours was unknown [13]. In one of these families, two patients with breast cancer only were also found; these cases were early-onset breast cancer.
In the present study, we did not find any pathogenic alterations in 10 families with cases of breast, gastric or colon cancer. We conclude that, although germline E-cadherin mutations are sometimes found in familial gastric and colon cancer, they are not frequently involved in families in which breast cancer appears to segregate as a part of an inherited predisposition for gastric and colon cancer.
Ala592Tyr alteration and breast cancer of ductal comedo type
The frequency of Ala592Thr alteration, which was first identified in one family with familial diffuse gastric, colon and breast cancer (Fig. 1a), was almost the same in the different groups studied here (Table 2). In familial breast cancer cases the frequency was 0.56% in non-BRCA1/BRCA2 cases, and 0.79% in BRCA1 or BRCA2 carriers. In the sporadic cases and in the control population, the frequencies were 0.83 and 0.80%, respectively. This finding does not support an effect of this alteration in predisposing to breast cancer in general. Thus, available data thus far indicate that germline E-cadherin mutations do not constitute a major risk factor for breast cancer. However, it is quite clear that somatic alterations can have an impact on the phenotype in lobular breast cancer. The fact that many breast cancer cases with the alteration (Ala592Thr) in the present study were of the ductal comedo type may indicate a genetic basis for the phenotypic divergence caused by this germline E-cadherin alteration. Although lobular breast carcinoma and ductal breast carcinoma of comedo type differ in many respects histologically, both lack formation of more differentiated mammary gland structures such as tubules. In lobular cancer, the cell characteristically grows in thin single-cell files (so-called Indian files). The comedo cancer is a poorly differentiated ductal cancer, which is rich in mitoses, often with large intraductal masses of cells with central necroses (comedons). Neither of these cancers carry a good prognosis, but individual prognosis with breast cancer is assessed mainly by grading rather than by histological type [20,21].
Correlation between loss of heterozygosity at 16q, tumourigenesis and metastasis in breast cancer
If E-cadherin functions as a classical tumour suppressor gene, then loss or inactivation of the remaining normal E-cadherin allele would be an important event in tumourigenesis in individuals who carry a germline mutation. In tumours with somatic E-cadherin mutations, there is often evidence of deletion of the wild-type E-cadherin allele [19], whereas in cases with germline mutations in the E-cadherin gene this is not usually seen [13,15,17]. We did not find any germline mutations in the 19 familial breast cancer cases with LOH at 16q studied here, which indicates that E-cadherin does not act as a predisposing factor for breast cancer in these families. In order to examine whether somatic alterations in this gene are involved in the carcinogenesis of these tumours, we screened for somatic mutations in nine available tumours from these cases, and we found one somatic mutation in one of the lobular cancer cases. In cases with no mutation in E-cadherin, however, inactivation of this gene could have been achieved by a variety of other mechanisms. Genetic alterations in any component of the cadherin–catenin complex appear to induce loss of adhesion function [39,40]. Furthermore, epigenetic changes such as promoter region hypermethylation, post-transcriptional alteration and aberrant phosphorylation of members of the cadherin-catenin complex can dysregulate E-cadherin function [41,42,43,44]. It is also possible that other gene(s) located at 16q are altered, rather than E-cadherin.
Conclusion
This present study, together with previously reported data, suggests that a germline mutation in E-cadherin is not a major risk factor for breast cancer. However, germline, and more often somatic mutations in this gene probably have an impact on phenotypic divergence and prognosis, including growth patterns of tumours, such as in lobular and perhaps ductal comedo breast cancers. In addition, other genetic alterations or epigenetic events at the E-cadherin gene may have an impact on the metastatic behaviour of the cancer cells, and thereby on the clinical outcome.
Abbrevations
DHPLC = denaturing high-performance liquid chromatography; LOH = loss of heterozygosity; PCR = polymerase chain reaction; SSCP = single-stranded conformation polymorphism.
References
- Shimoyama Y, Hirohashi S, Hirano S, Noguchi M, Shimosato Y, Takeichi M, Abe O. Cadherin cell-adhesion molecules in human epithelial tissues and carcinomas. Cancer Res. 1989;49:2128–2133. [PubMed] [Google Scholar]
- Takeichi M. Cadherins: a molecular family important in selective cell-cell adhesion. Annu Rev Biochem. 1990;59:237–252. doi: 10.1146/annurev.bi.59.070190.001321. [DOI] [PubMed] [Google Scholar]
- Knudsen KA, Frankowski C, Johnson KR, Wheelock MJ. A role for cadherins in cellular signaling and differentiation. J Cell Biochem Suppl. 1998;30-31:168–176. [PubMed] [Google Scholar]
- Moll R, Mitze M, Frixen UH, Birchmeier W. Differential loss of E-cadherin expression in infiltrating ductal and lobular breast carcinomas. Am J Pathol. 1993;143:1731–1742. [PMC free article] [PubMed] [Google Scholar]
- Berx G, Cleton-Jansen AM, Nollet F, de Leeuw WJ, van de Vijver M, Cornelisse C, van Roy F. E-cadherin is a tumour/invasion suppressor gene mutated in human lobular breast cancers. EMBO J. 1995;14:6107–6115. doi: 10.1002/j.1460-2075.1995.tb00301.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashiwaba M, Tamura G, Suzuki Y, Maesawa C, Ogasawara S, Sakata K, Satodate R. Epithelial-cadherin gene is not mutated in ductal carcinomas of the breast. Jpn J Cancer Res. 1995;86:1054–1059. doi: 10.1111/j.1349-7006.1995.tb03020.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zelada-Hedman M, Wasteson Arver B, Claro A, Chen J, Werelius B, Kok H, Sandelin K, Hakansson S, Andersen TI, Borg A, Borresen Dale AL, Lindblom A. A screening for BRCA1 mutations in breast and breast-ovarian cancer families from the Stockholm region. Cancer Res. 1997;57:2474–2477. [PubMed] [Google Scholar]
- Chen J, Hedman MZ, Arver BW, Sigurdsson S, Eyfjord JE, Lindblom A. BRCA2 germline mutations in Swedish breast cancer families. Eur J Hum Genet. 1998;6:134–139. doi: 10.1038/sj.ejhg.5200167. [DOI] [PubMed] [Google Scholar]
- Arver B, Claro A, Langerod A, Borresen-Dale A-L, Lindblom A. BRCA1 screening in patients with a family history of breast or ovarion cancer. Genet Test. 1999;3:223–226. doi: 10.1089/gte.1999.3.223. [DOI] [PubMed] [Google Scholar]
- Lindblom A, Skoog L, Rotstein S, Werelius B, Larsson C, Nordenskjöld M. Loss of heterozygosity in familial breast carcinomas. Cancer Res. 1993;53:4356–4361. [PubMed] [Google Scholar]
- Risinger JI, Berchuck A, Kohler MF, Boyd J. Mutations of the E-cadherin gene in human gynecologic cancers. Nat Genet. 1994;7:98–102. doi: 10.1038/ng0594-98. [DOI] [PubMed] [Google Scholar]
- Guilford P, Hopkins J, Harraway J, McLeod M, McLeod N, Harawira P, Taite H, Scoular R, Miller A, Reeve AE. E-cadherin germline mutations in familial gastric cancer. Nature. 1998;392:402–405. doi: 10.1038/32918. [DOI] [PubMed] [Google Scholar]
- Guilford PJ, Hopkins JBW, Grady WM, Markowitz SD, Willis J, Lynch H, Rajput A, Wiesner GL, Lindor NM, Burgart LJ, Toro TT, Lee D, Limacher JM, Shaw DW, Findlay MPN, Reeve AE. E-cadherin germline mutations define an inherited cancer syndrome dominated by diffuse gastric cancer. Hum Mutat. 1999;14:249–255. doi: 10.1002/(SICI)1098-1004(1999)14:3<249::AID-HUMU8>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- Gayther SA, Gorringe KL, Ramus SJ, Huntsman D, Roviello F, Grehan N, Machado JC, Pinto E, Seruca R, Halling K, MacLeod P, Powell SM, Jackson CE, Ponder BA, Caldas C. Identification of germ-line E-cadherin mutations in gastric cancer families of European origin. Cancer Res. 1998;58:4086–4089. [PubMed] [Google Scholar]
- Richards FM, McKee SA, Rajpar MH, Cole TR, Evans DG, Jankowski JA, McKeown C, Sander DS, Maher ER. Germline E-cadherin gene (CDH1) mutations predispose to familial gastric cancer and colorectal cancer. Hum Mol Genet. 1999;8:607–610. doi: 10.1093/hmg/8.4.607. [DOI] [PubMed] [Google Scholar]
- Yoon K-A, Ku J-L, Yang H-K, Kim WH, Park SY, Park J-G. Germline mutations of E-Cadherin gene in Korean familial gastric cancer patients. J Hum Genet. 1999;44:177–180. doi: 10.1007/s100380050137. [DOI] [PubMed] [Google Scholar]
- Keller G, Vogelsang H, Becker I, Hutter J, Ott K, Candidus S, Grundei T, Becker K, Mueller J, Siewert J, Hofler H. Diffuse type gastric and lobular breast carcinoma in a familial gastric cancer patient with an E-cadherin germline mutation. Am J Pathol. 1999;155:337–342. doi: 10.1016/S0002-9440(10)65129-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahman N, Stone JG, Coleman G, Gusterson B, Seal S, Marossy A, Lakhani SR, Ward A, Nash A, McKinna A, A'Hern R, Stratton MR, Houlston RS. Lobular carcinoma in situ of the breast is not caused by constitutional mutations in the E-cadherin gene. Br J Cancer. 2000;82:568–570. doi: 10.1054/bjoc.1999.0965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berx G, Cleton-Jansen AM, Strumane K, de Leeuw WJ, Nollet F, van Roy F, Cornelisse C. E-cadherin is inactivated in a majority of invasive human lobular breast cancers by truncation mutations throughout its extracellular domain. Oncogene. 1996;13:1919–1925. [PubMed] [Google Scholar]
- Elston CW, Ellis IO. Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: experience from a large study with long-term follow-up. Histopathology. 1991;19:403–410. doi: 10.1111/j.1365-2559.1991.tb00229.x. [DOI] [PubMed] [Google Scholar]
- Garne JP, Aspegren K, Linell F, Rank F, Ranstam J. Primary prognostic factors in invasive breast cancer with special reference to ductal carcinoma and histologic malignancy grade. Cancer. 1994;73:1438–1448. doi: 10.1002/1097-0142(19940301)73:5<1438::aid-cncr2820730519>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- Becker KF, Atkinson MJ, Reich U, Huang HH, Nekarda H, Siewert JR, Hofler H. Exon skipping in the E-cadherin gene transcript in metastatic human gastric carcinomas. Hum Mol Genet. 1993;2:803–804. doi: 10.1093/hmg/2.6.803. [DOI] [PubMed] [Google Scholar]
- Suzuki H, Komiya A, Emi M, Kuramochi H, Shiraishi T, Yatani R, Shimazaki J. Three distinct commonly deleted regions of chromosome arm 16q in human primary and metastatic prostate cancers. Genes Chromosomes Cancer. 1996;17:225–233. doi: 10.1002/(SICI)1098-2264(199612)17:4<225::AID-GCC4>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Tamura G, Sakata K, Nishizuka S, Maesawa C, Suzuki Y, Iwaya T, Terashima M, Saito K, Satodate R. Inactivation of the E-cadherin gene in primary gastric carcinomas and gastric carcinoma cell lines. Jpn J Cancer Res. 1996;87:1153–1159. doi: 10.1111/j.1349-7006.1996.tb03125.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muta H, Noguchi M, Kanai Y, Ochiai A, Nawata H, Hirohashi S. E-cadherin gene mutations in signet ring cell carcinoma of the stomach. Jpn J Cancer Res. 1996;87:843–848. doi: 10.1111/j.1349-7006.1996.tb02109.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soares P, Berx G, van Roy F, Sobrinho-Simoes M. E-cadherin gene alterations are rare events in thyroid tumors. Int J Cancer. 1997;70:32–38. doi: 10.1002/(sici)1097-0215(19970106)70:1<32::aid-ijc5>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- Kanai Y, Oda T, Tsuda H, Ochiai A, Hirohashi S. Point mutation of the E-cadherin gene in invasive lobular carcinoma of the breast. Jpn J Cancer Res. 1994;85:1035–1039. doi: 10.1111/j.1349-7006.1994.tb02902.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker KF, Reich U, Schott C, Becker I, Berx G, van Roy F, Hofler H. Identification of eleven novel tumor-associated E-cadherin mutations. Mutations in brief no. 215. Online. Hum Mutat. 1999;13:171. doi: 10.1002/(SICI)1098-1004(1999)13:2<171::AID-HUMU14>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- Oda T, Kanai Y, Oyama T, Yoshiura K, Shimoyama Y, Birchmeier W, Sugimura T, Hirohashi S. E-cadherin gene mutations in human gastric carcinoma cell lines. Proc Natl Acad Sci USA. 1994;91:1858–1862. doi: 10.1073/pnas.91.5.1858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker KF, Atkinson MJ, Reich U, Becker I, Nekarda H, Siewert JR, Hofler H. E-cadherin gene mutations provide clues to diffuse type gastric carcinomas. Cancer Res. 1994;54:3845–3852. [PubMed] [Google Scholar]
- Siitonen SM, Kononen JT, Helin HJ, Rantala IS, Holli KA, Isola JJ. Reduced E-cadherin expression is associated with invasiveness and unfavorable prognosis in breast cancer. Am J Clin Pathol. 1996;105:394–402. doi: 10.1093/ajcp/105.4.394. [DOI] [PubMed] [Google Scholar]
- Jones JL, Royall JE, Walker RA. E-cadherin relates to EGFR expression and lymph node metastasis in primary breast carcinoma. Br J Cancer. 1996;74:1237–1241. doi: 10.1038/bjc.1996.522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukholm IK, Nesland JM, Karesen R, Jacobsen U, Borresen-Dale AL. E-cadherin and alpha-, beta-, and gamma-catenin protein expression in relation to metastasis in human breast carcinoma. J Pathol. 1998;185:262–266. doi: 10.1002/(SICI)1096-9896(199807)185:3<262::AID-PATH97>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Lindblom A, Rotstein S, Skoog L, Nordenskjöld M, Larsson C. Deletions on chromosome 16 in primary familial breast carcinomas are associated with development of distant metastases. Cancer Res. 1993;53:3707–3711. [PubMed] [Google Scholar]
- Cleton-Jansen AM, Moerland EW, Kuipers-Dijkshoorn NJ, Callen DF, Sutherland GR, Hansen B, Devilee P, Cornelisse CJ. At least two different regions are involved in allelic imbalance on chromosome arm 16q in breast cancer. Genes Chromosomes Cancer. 1994;9:101–107. doi: 10.1002/gcc.2870090205. [DOI] [PubMed] [Google Scholar]
- Caligo MA, Polidoro L, Ghimenti C, Campani D, Cecchetti D, Bevilacqua G. A region on the long arm of chromosome 16 is frequently deleted in metastatic node-negative breast cancer. Int J Oncol. 1998;13:177–182. doi: 10.3892/ijo.13.1.177. [DOI] [PubMed] [Google Scholar]
- Stone J, Bevan S, Cunningham D, Hill A, Rahman N, Peto J, Marossy A, Houlston R. Low frequency of germline E-cadherin mutations in familial and nonfamilial gastric cancer. Br J Cancer. 1999;79:1935–1937. doi: 10.1038/sj.bjc.6690308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iida S, Akiyama Y, Ichikawa W, Yamashita T, Nomizu T, Nihei Z, Sugihara K, Yuasa Y. Infrequent germ-line mutation of the E-cadherin gene in Japanese familial gastric cancer kindreds. Clin Cancer Res. 1999;5:1445–1447. [PubMed] [Google Scholar]
- Morton RA, Ewing CM, Nagafuchi A, Tsukita S, Isaacs WB. Reduction of E-cadherin levels and deletion of the alpha-catenin gene in human prostate cancer cells. Cancer Res. 1993;53:3585–3590. [PubMed] [Google Scholar]
- Oyama T, Kanai Y, Ochiai A, Akimoto S, Oda T, Yanagihara K, Nagafuchi A, Tsukita S, Shibamoto S, Ito F, et al. A truncated beta-catenin disrupts the interaction between E-cadherin and alpha-catenin: a cause of loss of intercellular adhesiveness in human cancer cell lines. Cancer Res. 1994;54:6282–6287. [PubMed] [Google Scholar]
- Hiraguri S, Godfrey T, Nakamura H, Graff J, Collins C, Shayesteh L, Doggett N, Johnson K, Wheelock M, Herman J, Baylin S, Pinkel D, Gray J. Mechanisms of inactivation of E-cadherin in breast cancer cell lines. Cancer Res. 1998;58:1972–1977. [PubMed] [Google Scholar]
- Graff JR, Herman JG, Lapidus RG, Chopra H, Xu R, Jarrard DF, Isaacs WB, Pitha PM, Davidson NE, Baylin SB. E-cadherin expression is silenced by DNA hypermethylation in human breast and prostate carcinomas. Cancer Res. 1995;55:5195–5199. [PubMed] [Google Scholar]
- Yoshiura K, Kanai Y, Ochiai A, Shimoyama Y, Sugimura T, Hirohashi S. Silencing of the E-cadherin invasion-suppressor gene by CpG methylation in human carcinomas. Proc Natl Acad Sci USA. 1995;92:7416–7419. doi: 10.1073/pnas.92.16.7416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata T, Gotoh M, Ochiai A, Hirohashi S. Association of plakoglobin with APC, a tumor suppressor gene product, and its regulation by tyrosine phosphorylation. Biochem Biophys Res Commun. 1994;203:519–522. doi: 10.1006/bbrc.1994.2213. [DOI] [PubMed] [Google Scholar]