

Abstract
Ginkgo biloba extract, EGb 761, a popular and standardized natural extract, contains 24% ginkgo-flavonol glycosides and 6% terpene lactones. EGb 761 is used worldwide to treat many ailments, and while a number of studies have shown its neuroprotective properties, the mechanisms of action have not been elucidated fully. We hypothesize that EGb 761 and some of its bioactive components [Bilobalide (BB), Ginkgolide A (GA), Ginkgolide B (GB), and Terpene Free Material (TFM)] could provide neuroprotection ischemic conditions through heme oxygenase 1 (HO1). Mice were subjected permanent distal middle cerebral artery occlusion (pMCAO) and survived for 7 days. HO1-/- mice showed significantly higher (p<0.05) infarct volume and Neurologic Deficit Scores (NDS) as compared to their wildtype (WT) counterparts. In another cohort, mice subjected to pMCAO and treated at 4 h of pMCAO with 100mg/kg EGb 761 6mg/kg BB, GA, GB, or 10mg/kg TFM showed significantly lower (p<0.05) infarct volumes (BB; 29.0±3.9%, GA; 31.3±4.0%, GB; 32.0±3.8%, TFM; 32.5±3.5%, and 761; 27.4±4.5%) than those in the vehicle-treated mice (46.0±3.7%). Similarly, were lower in BB: 7.1±1.8, GA; 7.4±2.1, GB; 7.9±1.8, TFM; 7.7±1.7, and EGb 6.8±2.0 groups as compared with the vehicle-treated group (13.8±1.5). Interestingly, the protective effect of EGb 761 was essentially lost when HO1 knockout mice were treated with EGb 761. In another cohort, HO1, VEGF and eNOS protein levels in the cortices appeared to be higher in EGb 761 and BB but not in GA, GB and TFM treated groups. Together, these results suggest that HO1 plays, at least in part, an important role in the neuroprotective mechanism of EGb 761 and in delayed ischemia. Targeting this pathway could lead to neuroprotective agents against ischemic stroke.
Keywords: Bioactive, Complementary and alternative medicine, Hemin, Natural supplements, Neurologic disorders
Extracts of Ginkgo biloba leaves have been used for centuries in Asia as a traditional medicine for various diseases, and now the standardized extract of Ginkgo biloba (EGb 761) is being prescribed widely by doctors in Europe for the treatment of a range of conditions including, but not limited to: confusion; depression and anxiety, dizziness and headache, tinnitus, memory and concentration problems (Chan et al., 2007). At present, EGb 761 extract is one of the most renowned and commonly used natural compounds, and it has been used in many preclinical and clinical studies to evaluate its efficacy (Snitz et al., 2009). It contains: 24% ginkgo-flavanol glycosides; 6% terpene lactones such as ginkgolides A, B, C, J, and bilobalide; 5-10% organic acids; and >0.5% proanthocyanidins, defined as flavonoid-based polymers (van Beek, 2002).
Numerous studies have shown neuroprotective properties of EGb 761 and its different constituents, but the mechanism underlying its neuroprotection has not been studied fully (Bastianetto et al., 2000, Lee et al., 2002, Chandrasekaran et al., 2003, Mdzinarishvili et al., 2007, Saleem et al., 2008). We have previously shown that HO1 is a key player for EGb 761 neuroprotection in chronic treatment and transient middle cerebral artery occlusion (MCAO) model (Saleem et al., 2008). HO is the rate-limiting enzyme required for the conversion of deleterious heme into neuroprotective products like bilirubin, biliverdin and carbon monoxide, and it has two isoforms. HO1 is highly inducible, while HO2 is constitutively present in the brain (Ahmad et al., 2006). Under the experimental protocol, no significant difference was observed between WT and HO1-/- mice in a transient MCAO model (1 h occlusion and 23 hrs reperfusion) (Doré et al., 1999). To our knowledge, delayed ischemic effects of HO1 in transient middle cerebral artery occlusion (tMCAO) have not been elucidated yet, as mice do not survive to longer time points with this procedure. Using permanent distal MCAO (pMCAO) ischemia model gives us an advantage as compared to the tMCAO to study the role of protective agents especially on delayed ischemic effects.
Therefore, the present study was designed to evaluate the delayed ischemic response of HO1 by subjecting WT and HO1-/- mice to pMCAO and surviving them up to 7 days. We further questioned whether acute doses of EGb 761 and its bioactive components [Bilobalide (BB); Ginkgolide A (GA); Ginkgolide B (GB); Terpene Free Material (TFM)] may have intrinsic properties and provide protection against pMCAO-induced cortical brain injury. Finally, we investigated the HO1, HO2, VEGF and eNOS protein expression levels in brain cortices of drug and vehicle treated groups.
Experimental Procedures
Animals
All animal protocols were approved by the Johns Hopkins University Institutional Animal Care and Use Committee/University of Toledo Health Science Campus Institutional Animal Care and Use Committee, and the guidelines of the National Institutes of Health were followed throughout the study. C57BL/6 (WT) mice were procured from Charles River Laboratories, Wilmington, MA, and knockout (HO1-/- and HO2-/-) mice were bred in our laboratory. All animals were 5–10 weeks old and weighed about 20–25 grams. Animals were housed at 22±1 °C with a 12 h:12 h light/dark cycle; water and food were available ad libitum.
Permanent distal middle cerebral artery occlusion (pMCAO)
The distal portion of the MCA was occluded as per the previously described method (Saleem et al., 2009, Zeynalov et al., 2009). Mice were anesthetized with halothane (Nicholas Piramal, India), initially with 2% and then maintained at 1% throughout the surgical procedure. Under a surgical microscope, a 1.0-cm vertical skin incision between the right eye and ear was made, the temporal muscle was moved aside, and the underlying temporal bone exposed. Accordingly, a 2.0-mm hole was made with the help of dental drill over the area of distal MCA, visible through the temporal bone. The distal part of MCA was occluded directly with a bipolar coagulator, and complete interruption of blood flow at the occlusion site was confirmed by severance of the MCA and subsequently confirmed by placing the laser-Doppler probe above the temporal ridge to establish that blood flow into the region was terminated. Core body temperature was continuously monitored and maintained at 37.0±0.5 °C during and after the procedure, first with a heating blanket that was attached to the temperature probe for automatic temperature regulation, and then with a temperature-regulated incubator in which the mice recovered from the surgery.
Drug treatment
EGb 761 (100mg/kg), BB (6mg/kg), GA (6mg/kg), GB (6mg/kg), TFM (10mg/kg) were kindly provided by IPSEN/Schwabe Laboratories. Test drugs and vehicle were administered to mice immediately after 4 h of pMCAO. For protein expression experiments, drugs were administered immediately after 1.5 h of pMCAO. The dose concentrations were selected as per the composition of various constituents present in the extract of EGb 761.
Neurologic Deficit Score (NDS)
Neurologic deficits induced by pMCAO were evaluated by a previously optimized 28-point score pattern (Saleem et al., 2009, Zeynalov et al., 2009). After 7 days of pMCAO, NDS were evaluated, and tests included both sensory and motor deficits, such as body symmetry, gait, climbing, circling behavior, front limb symmetry, compulsory circling, and whisker response. Each test of the 28-point scale NDS was graded from 0 to 4; therefore 28 was considered to indicate severe deficit. To perform infarct volume analysis, mice were sacrificed immediately after the NDS evaluation.
Infarction volume analysis
Animals from all the groups were euthanized at 7 days after pMCAO procedure. Brains were dissected out, sliced into five 2-mm-thick coronal sections and incubated in 1% triphenyltetrazolium chloride (Sigma Co, MI, USA) in saline solution for 20 min at 37 °C. Brain sections fixed over night in formaldehyde solution were analyzed for infarct areas with the Image Analysis (SigmaScan pro 5, Systat, Inc., Point Richmond, CA). The infarct area was estimated from five slices of the brain, measuring rostral and caudal sides of each individual slice in combination with the thickness and expressed as a percentage of the volume of the contralateral structure. Infarct volumes were also corrected for brain swelling.
Western blot analysis
Brain cortices of ischemic and non-ischemic mice were dissected out, weighed, and homogenized. Protein concentrations were determined by Bradford reagent (Bio-Rad Laboratories, CA, USA) and samples were analyzed by loading equivalent amounts of total proteins (30 μg) onto 10% SDS-polyacrylamide gels. Proteins were transferred from the gel to PVDF membrane and blocked by 5% dry nonfat milk for 1 h at room temperature followed by overnight incubation at 4 °C with following antibodies: rabbit anti-actin (1:200; Sigma); rabbit anti-HO2 (1:2000; Stressgen, MI, USA); rabbit anti-HO1 (1:1000; Stressgen); rabbit anti-VEGF (1:1000; Santa Cruz Biotechnology, CA, USA); rabbit anti-eNOS (1:750; Thermo Scientific, IL, USA). After washing, membranes were incubated with the secondary antibody, goat anti-rabbit (1:5000; Jackson ImmunoResearch Laboratories, PA, USA). Images were analyzed using Photoshop and Image J software provided by the NIH. The densitometric values were normalized with respect to the values of actin immunoreactivity to correct for any loading and transfer differences between samples.
Statistical analysis
Infarct volumes between WT and knockouts and treatment groups were analyzed by one-way ANOVA with Newman Keuls post-hoc test. Neurologic deficits were analyzed by the non-parametric Kruskal-Wallis test. Data are presented as mean±SEM. A value of p<0.05 was considered to be statistically significant.
Results
Effect of HO1 genetic deletion on infarct volume and NDS, 7 days after pMCAO
HO1-/- mice subjected to pMCAO for 7 d showed significantly (p<0.005) larger hemispheric cortical infarct volumes (57.9±2.4%) as compared to their WT counter parts (44.9±2.9). Similarly, NDS were also observed to be higher (p<0.04) in HO1-/- (19.5±2.1) as compared to the WT (13.6±1.4) mice (Fig. 1).
Fig. 1.
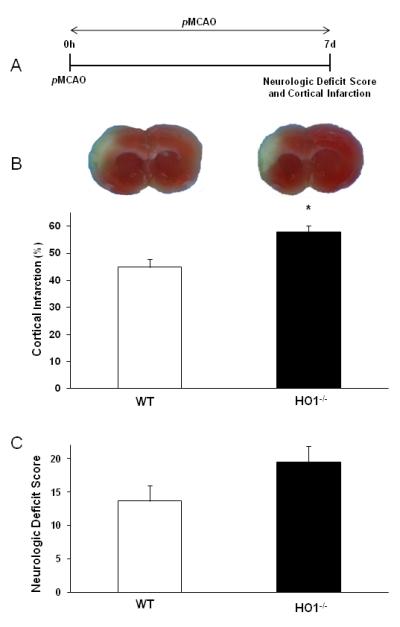
Role of HO1 against 7 d pMCAO. (A) Schematic diagram of experimental protocol. Mice were subjected to pMCAO and survived for 7 d to evaluate NDS and infarct volume. (B and C) Representative TTC stained brain slices and graph show higher infarct volume and NDS in HO1-/- than WT mice. WT (n=8); HO1-/- (n=6). *p<0.05 vs. corresponding control.
Protective effect of EGb 761 and its constituents against 7 days after pMCAO
Next, we evaluated and compared the protective potential of EGb 761 and its constituents (BB, GA, GB and TFM). In another cohort of six groups of animals, test drugs were administered by gavage 4 h after inducing pMCAO, and mice were survived for 7 d. We observed significantly smaller hemispheric cortical infarct volumes in treatment groups of BB (29.0±3.9%, p<0.01), GA (31.3±4.0%, p<0.02), GB (32.0±3.8%, p<0.02), TFM (32.5±3.5, p<0.02) and EGb 761 (27.4±4.5%; p<0.01) as compared to vehicle-treated (46.0±3.7) mice. Neurologic deficit scores were also observed to be significantly lower in BB (7.1±1.8, p<0.02), GA (7.4±2.1, p<0.04), GB (7.9±1.8, p<0.03), TFM (7.7±1.7, p<0.03) and EGb 761 (6.8±2.0; p<0.02) treated groups as compared to vehicle-treated (13.8±1.5) animals (Fig. 2).
Fig. 2.
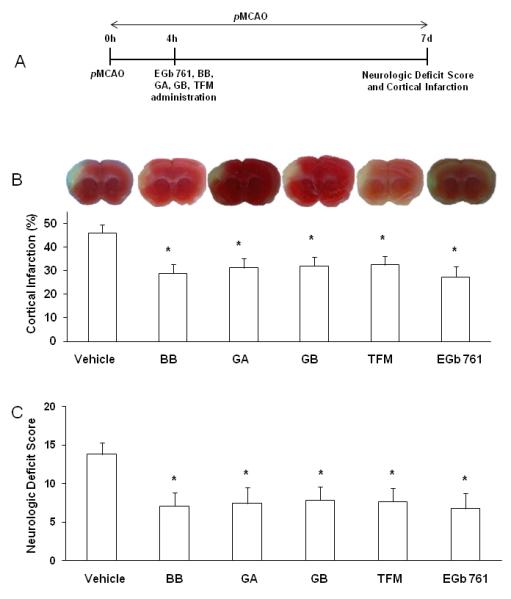
Neuroprotective effect of EGb 761 and its constituents. (A) Schematic diagram of experimental protocol. Vehicle or test drugs were administered after 4 h of pMCAO, and mice survived for 7 d to evaluate NDS and infarct volume. (B and C) Representative TTC stained brain slices and graph show reduced infarct volumes and NDS of WT mice treated with Ginkgo biloba (EGb 761), Bilobalide (BB), Ginkgolide A (GA), Ginkgolide B (GB) and Terpene Free Material (TFM). Vehicle (n=6); BB (n=10); GA (n=9), GB (n=8), TFM (n=9), EGb 761 (n=8). *p<0.05 vs. corresponding vehicle control.
EGb 761 is not protective in HO1-/- mice
To test the hypothesis that HO1 is necessary for EGb 761 neuroprotection, we subjected HO1-/- mice to same experimental protocol. HO1-/- mice treated with EGb 761 at 4 h of pMCAO did not show any beneficial effects on infarct volume as compared with vehicle-treated WT group (Experiment 2). As expected, EGb 761 treated HO1-/- mice showed no differences on neurologic deficits scale than the vehicle-treated WT group in experiment 2 (Fig. 3).
Fig. 3.
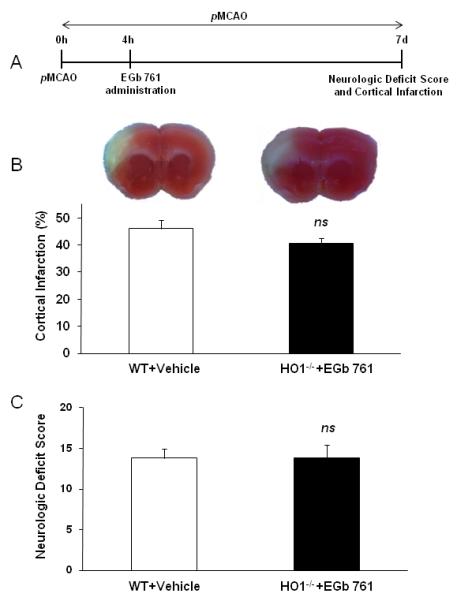
EGb 761 is not protective in HO1-/- mice. (A) Schematic diagram of experimental protocol. Vehicle or EGb 761 was administered after 4 h of pMCAO, and mice survived for 7 d to evaluate NDS and infarct volume. (B and C) Representative TTC stained brain slices and graph show EGb 761 is not protective in HO1-/- mice as compared to vehicle treatment group (WT vehicle treatment group from experiment/Fig. 2). WT and Vehicle (n=6), EGb 761 and HO1-/- (n=7). *p<0.05 vs. Vehicle control; ns, not significant.
EGb 761 is not protective in HO2-/- mice
We further wanted to investigate role of HO2 in EGb 761 protection during pMCAO. Mice with HO2 genetic deletion were subjected to same treatment plan of 4 h post-pMCAO EGb 761 treatment and 7 d survival. We observed that EGb 761 administration was not protective in HO2-/- mice. Infarct volume and neurologic deficits were not different as compared to the vehicle-treated HO2-/- mice (Fig. 4).
Fig. 4.
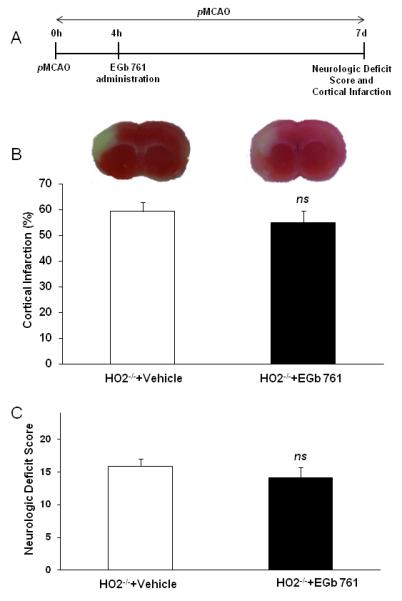
EGb 761 is not protective in HO2-/- mice. (A) Schematic diagram of the experimental protocol. Vehicle or EGb 761 was administered after 4 h of pMCAO, and mice survived for 7 d to evaluate NDS and infarct volume. (B and C) Representative TTC stained brain slices and graph show EGb 761 is not protective in HO2-/- mice as compared to vehicle treatment group. HO2-/- and Vehicle (n=7), EGb 761 and HO2-/- (n=7). *p<0.05 vs. corresponding control; ns, not significant.
HO1, HO2, VEGF and eNOS protein levels in brain cortices
To ascertain whether EGb 761 or its components induce HO1, HO2, VEGF and eNOS protein expression in the brain cortex, WT mice were subjected to pMCAO and post-treated with test drugs (EGb 761, BB, GA, GB, TFM and vehicle) at 1.5 h. Mice brain cortices were dissected out on 7 d for Western blot analysis. HO1 protein expression was significantly increased in EGb 761 and BB treated groups as compared to vehicle treated group. Significant up-regulation was also observed in the EGb 761 drug treated naïve group (no pMCAO). However, as previously reported (Zeynalov et al., 2009a), no differences in HO2 protein expression levels was observed in all treatment groups. VEGF levels were significantly elevated in EGb 761 and showed an increasing trend in BB. Correspondingly, eNOS levels were also observed to be increased in EGb 761 and BB treatment groups. On the other hand, no changes in protein expression levels of HO1, HO2, VEGF and eNOS were observed with the treatment of GA, GB and TFM (Fig. 5).
Fig. 5.
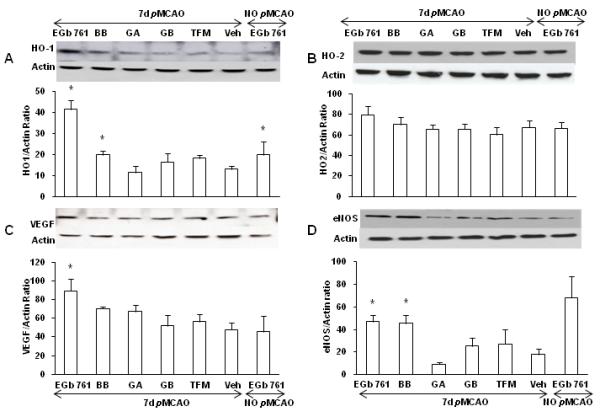
HO1, HO2, VEGF and eNOS protein expression levels in mice brain cortices. Mice were post-treated at 1.5 h of pMCAO with EGb 761 and its constituents, and Western blots were performed to measure the protein expression of HO1, HO2, VEGF and eNOS. The expression of actin was used as a loading control. (A) HO1 expression levels were observed to be significantly increased by the post-treatment of EGb 761 and BB. The expression levels of HO1 were also increased in naïve EGb 761 treated group (non surgery). (B) HO2, known to be constitutively expressed in the brain, was unaffected. (C) VEGF expression levels were significantly increased in EGB 761 and showed an increasing trend in BB treated group. (D) Similarly, eNOS protein expression levels showed significant increase in EGb 761 and BB treated groups. The histograms show the ratio of density captured from HO1, HO2, VEGF, eNOS to that of actin. Values shown are means±SEM from three independent sets of experiments. *p<0.05 vs. corresponding control.
Discussion
In the current study, we have demonstrated that HO1 is neuroprotective in pMCAO-induced ischemic brain injury at 7 d of survival. Secondly, we showed that 4 h post-treatment with Ginkgo biloba (EGb 761) and its constituents [Bilobalide (BB); Ginkgolide A (GA); Ginkgolide B (GB); Terpene Free Material (TFM)] provides neuroprotection against pMCAO. The protective effect of EGb 761 was abrogated in HO1-/- and HO2-/- mice. Furthermore, EGb 761 and BB treated groups showed increased HO1, VEGF and eNOS protein expression levels in the cortical homogenates, while no differences were observed in GA, GB and TFM treated groups.
HO degrades pro-oxidant free heme to iron, carbon monoxide (CO), and biliverdin, which is reduced to bilirubin (BR). It has been shown that CO possesses vasodilatory properties and that biliverdin and BR act as antioxidants. Based on the available literature, it has been reported that HO1-/- and WT mice have no differences in infarct volumes in a transient MCAO reperfusion model at 24 h (Doré et al., 1999), HO1 exacerbates stroke damage in hemorrhage model at 24 and 72 h (Wang and Doré, 2007) and is neuroprotective in NMDA-induced excitotoxicity model. Recently, we determined that HO1-/- mice suffer from higher cortical brain damage as compared to WT mice at 48 h of pMCAO (Zeynalov et al., 2009b). HO1 has been studied in various paradigms, and its role in ischemic (Zeynalov et al., 2009b) and remote organ preconditioning (Lai et al., 2006) has already been established. Cell specificity, origin and nature of damage are possible reasons for variability in the role of HO1 in different disease models. HO2, which is present constitutively, is observed to provide beneficial effects in hemorrhagic stroke (Wang and Doré, 2008) and in transient middle cerebral artery occlusion models (Doré et al., 1999, Namiranian et al., 2005).
In vestibular compensation, EGb 761 has been suggested to have direct effects against necrosis and apoptosis of neurons and improves neural plasticity (Maclennan et al., 2002). In animal models, it was originally suggested that EGb 761 can act as a free-radical scavenger and prevent lipid peroxidation and overall reactive oxygen species at the molecular and the cellular levels (Barth et al., 1991). EGb 761 can regulate the ionic balance in damaged cells and potentially antagonizes the activity of platelet-activating factor (Vogensen et al., 2003). Several investigators have reported the protective effects of Ginkgo extracts in different models of hypoxia and ischemia in various animal species, including gerbils (Lin et al., 2004), rats (Zhang et al., 2000), mice (Lu et al., 2006) and rabbits (Fan et al., 2006). Several controlled clinical studies conducted in Europe and the U.S. have revealed that EGb 761 is an effective therapy for a wide variety of disturbances of cerebral function such as multi-infarct dementia, early or mild cognitive decline, and severe types of senile dementias (Walesiuk and Braszko, 2007). Contrarily, the more recent clinical trial, “Ginkgo Evaluation of Memory (GEM)”, showed that EGB 761 is not effective in treating Alzheimer’s disease (DeKosky et al., 2006). Here, we demonstrated that EGb 761 and some of its active components (BB, GA GB, and TFM) can be protective in permanent stroke model. To our knowledge, this is one of the first studies to conduct experiments involving different components of EGb 761 in ischemic brain injury. We observed variable levels of protection in terms of infarct volumes with different components, i.e. EGb 761 showed highest potential for protection, which was followed by BB, GA, GB and TFM. Our results are consistent with a previous study showing that EGb 761 and its components have variable levels of protection during radiation-induced clastogenic factors in rats (Alaoui-Youssefi et al., 1999). Our conclusion is that the highest protection observed in the EGb 761 may be through its synergistic effects rather than additive, which is consistent with the previous finding. The next possible reason for variability may be the amount of antioxidant properties found in individual components, which warrants further evaluation. We and other investigators have shown that many polyphenolic compounds exert their antioxidant properties via Nrf2 activation, which in-turn binds to ARE’s of antioxidant proteins and provides defense against oxidative stress (Liu et al., 2007, Boettler et al., 2010, Shah et al., 2010). We have showed that the effect of EGb 761 was abrogated in HO1-/- mice, suggesting that HO plays a major role in mediating the mechanism of EGb 761 neuroprotection. Our previous in vitro and in vivo results have demonstrated that EGb 761 upregulates the expression of heme oxygenase (HO), which is thought to be a potent antioxidant (Zhuang et al., 2002, Saleem et al., 2008). Our results are similar to those of Saleem et al. (Saleem et al., 2008), showing the role of HO1 in transient ischemia model at 24 hours reperfusion time point. To further validate and rule out the role of HO2, the constitutively present HO isoform, EGb 761 treatment was not observed to induce protection in HO2-/- mice. The role of HO1 was further confirmed by its increased protein expression in the cortices of the mouse brain treated with EGb 761 and BB.
Discussing further the mechanism of action of EGb 761 and its components, we propose that several extract components may have played a role, individually or collectively, either by increasing blood supply by dilating blood vessels, reducing blood viscosity (Santos et al., 2003), modifying neurotransmitter systems (Davies et al., 2003, Shah et al., 2003), and reducing the concentration of oxygen free radicals (Sastre et al., 1998, Hibatallah et al., 1999, Chandrasekaran et al., 2003). EGb 761 has also been shown to increase CBF in mice (Saleem et al., 2008) and humans (Mashayekh et al.), possibly due to increased HO1 and eNOS levels that lead to vasodilation and increased cerebral blood flow (Wang and Chen, 2005, Koltermann et al., 2009). VEGF has been shown to activate eNOS in endothelial cells, and together they may provide enhanced CBF during injury (Duda et al., 2004). Our results showing increased VEGF and eNOS levels are in consonance with the previous studies and extend further support to the fact that EGb 761 and some of its components increase CBF by upregulating the vasodilatory components such as VEGF and eNOS. Furthermore, extract of EGb 761 and BB were the only candidates that worked through the induction of HO1; other components may have worked through different signaling pathways. Various studies have postulated different signaling pathways for different components. Chandrasekaran et al. (Chandrasekaran et al., 2003) showed that EGb 761 and BB have properties like anti-excitotoxicity, inhibition of free radical generation, and scavenging reactive oxygen species. Another study by Wang et al. (Wang and Chen, 2005) showed GB facilitates glutamate exocytosis via PKA but not by PKC pathway. Saleem et al (Saleem et al., 2008), using primary neuronal cultures, showed that EGb 761 but not BB, or GA/GB pretreatment induced robust HO1 protein expression. All these divergent findings lead us to postulate that EGb 761 and its components work through multiple pathways and warrant further investigation.
To conclude, we have shown the protective role of HO1 in pMCAO model after 7 days of survival. The neuroprotective effect of EGb 761 and its constituents was also evaluated, and it was observed that EGb 761 has a synergistic effect rather than additive. Lastly, we observed the role of HO1, VEGF and eNOS in propagating the protective effect of EGb 761 and some of its components. We believe that the HO1 induced pathway plays a primary role in mediating the neuroprotective potential of EGb 761. Discovering target drug molecules that are upregulated by natural compounds provides an innovative tool for improving treatment for stroke.
Acknowledgements
This work was supported by grants from the National Institutes of Health K99AT004197 (ZAS), and R21AT001836, R21AT005085 (SD). The authors would like to thank Charisse N. Montgomery for her assistance in the manuscript editing.
Abbreviations
- tMCAO
transient middle cerebral artery occlusion
- pMCAO
permanent distal middle cerebral artery occlusion
- HO1
heme oxygenase 1
- WT
wildtype
- HO1-/-
mice
- CBF
Cerebral blood flow
- NDS
neurologic deficit score
Footnotes
Some of the data were previously presented at the 2011 International Stroke Conference.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ahmad AS, Zhuang H, Doré S. Heme oxygenase-1 protects brain from acute excitotoxicity. Neuroscience. 2006;141:1703–1708. doi: 10.1016/j.neuroscience.2006.05.035. [DOI] [PubMed] [Google Scholar]
- Alaoui-Youssefi A, Lamproglou I, Drieu K, Emerit I. Anticlastogenic effects of Ginkgo biloba extract (EGb 761) and some of its constituents in irradiated rats. Mutat Res. 1999;445:99–104. doi: 10.1016/s1383-5718(99)00139-4. [DOI] [PubMed] [Google Scholar]; Barth SA, Inselmann G, Engemann R, Heidemann HT. Influences of Ginkgo biloba on cyclosporin A induced lipid peroxidation in human liver microsomes in comparison to vitamin E, glutathione and N-acetylcysteine. Biochem Pharmacol. 1991;41:1521–1526. doi: 10.1016/0006-2952(91)90570-u. [DOI] [PubMed] [Google Scholar]
- Bastianetto S, Ramassamy C, Doré S, Christen Y, Poirier J, Quirion R. The Ginkgo biloba extract (EGb 761) protects hippocampal neurons against cell death induced by beta-amyloid. Eur J Neurosci. 2000;12:1882–1890. doi: 10.1046/j.1460-9568.2000.00069.x. [DOI] [PubMed] [Google Scholar]
- Boettler U, Sommerfeld K, Volz N, Pahlke G, Teller N, Somoza V, Lang R, Hofmann T, Marko D. Coffee constituents as modulators of Nrf2 nuclear translocation and ARE (EpRE)-dependent gene expression. J Nutr Biochem. 2010 doi: 10.1016/j.jnutbio.2010.03.011. [DOI] [PubMed] [Google Scholar]
- Chan PC, Xia Q, Fu PP. Ginkgo biloba leave extract: biological, medicinal, and toxicological effects. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2007;25:211–244. doi: 10.1080/10590500701569414. [DOI] [PubMed] [Google Scholar]
- Chandrasekaran K, Mehrabian Z, Spinnewyn B, Chinopoulos C, Drieu K, Fiskum G. Neuroprotective effects of bilobalide, a component of Ginkgo biloba extract (EGb 761) in global brain ischemia and in excitotoxicity-induced neuronal death. Pharmacopsychiatry. 2003;36(Suppl 1):S89–94. doi: 10.1055/s-2003-40447. [DOI] [PubMed] [Google Scholar]
- Davies JA, Johns L, Jones FA. Effects of bilobalide on cerebral amino acid neurotransmission. Pharmacopsychiatry. 2003;36(Suppl 1):S84–88. doi: 10.1055/s-2003-40448. [DOI] [PubMed] [Google Scholar]
- DeKosky ST, Fitzpatrick A, Ives DG, Saxton J, Williamson J, Lopez OL, Burke G, Fried L, Kuller LH, Robbins J, Tracy R, Woolard N, Dunn L, Kronmal R, Nahin R, Furberg C. The Ginkgo Evaluation of Memory (GEM) study: design and baseline data of a randomized trial of Ginkgo biloba extract in prevention of dementia. Contemp Clin Trials. 2006;27:238–253. doi: 10.1016/j.cct.2006.02.007. [DOI] [PubMed] [Google Scholar]
- Doré S, Sampei K, Goto S, Alkayed NJ, Guastella D, Blackshaw S, Gallagher M, Traystman RJ, Hurn PD, Koehler RC, Snyder SH. Heme oxygenase-2 is neuroprotective in cerebral ischemia. Mol Med. 1999;5:656–663. [PMC free article] [PubMed] [Google Scholar]
- Duda DG, Fukumura D, Jain RK. Role of eNOS in neovascularization: NO for endothelial progenitor cells. Trends Mol Med. 2004;10:143–145. doi: 10.1016/j.molmed.2004.02.001. [DOI] [PubMed] [Google Scholar]
- Fan LH, Wang KZ, Cheng B. Effects of Ginkgo biloba extract on lipid peroxidation and apoptosis after spinal cord ischemia/reperfusion in rabbits. Chin J Traumatol. 2006;9:77–81. [PubMed] [Google Scholar]
- Hibatallah J, Carduner C, Poelman MC. In-vivo and in-vitro assessment of the free-radical-scavenger activity of Ginkgo flavone glycosides at high concentration. J Pharm Pharmacol. 1999;51:1435–1440. doi: 10.1211/0022357991777083. [DOI] [PubMed] [Google Scholar]
- Koltermann A, Liebl J, Furst R, Ammer H, Vollmar AM, Zahler S. Ginkgo biloba extract EGb 761 exerts anti-angiogenic effects via activation of tyrosine phosphatases. J Cell Mol Med. 2009;13:2122–2130. doi: 10.1111/j.1582-4934.2008.00561.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai IR, Chang KJ, Chen CF, Tsai HW. Transient limb ischemia induces remote preconditioning in liver among rats: the protective role of heme oxygenase-1. Transplantation. 2006;81:1311–1317. doi: 10.1097/01.tp.0000203555.14546.63. [DOI] [PubMed] [Google Scholar]
- Lee EJ, Chen HY, Wu TS, Chen TY, Ayoub IA, Maynard KI. Acute administration of Ginkgo biloba extract (EGb 761) affords neuroprotection against permanent and transient focal cerebral ischemia in Sprague-Dawley rats. J Neurosci Res. 2002;68:636–645. doi: 10.1002/jnr.10251. [DOI] [PubMed] [Google Scholar]
- Lin JY, Cheng FC, Chung SY, Lin MC. Ginkgo biloba extract (EGb761) and FK506 preserve energy metabolites in the striatum during focal cerebral ischemia and reperfusion in gerbils monitored by microdialysis. J Biomed Sci. 2004;11:611–616. doi: 10.1007/BF02256126. [DOI] [PubMed] [Google Scholar]
- Liu XP, Goldring CE, Copple IM, Wang HY, Wei W, Kitteringham NR, Park BK. Extract of Ginkgo biloba induces phase 2 genes through Keap1-Nrf2-ARE signaling pathway. Life Sci. 2007;80:1586–1591. doi: 10.1016/j.lfs.2007.01.034. [DOI] [PubMed] [Google Scholar]
- Lu G, Wu Y, Mak YT, Wai SM, Feng ZT, Rudd JA, Yew DT. Molecular evidence of the neuroprotective effect of Ginkgo biloba (EGb761) using bax/bcl-2 ratio after brain ischemia in senescence-accelerated mice, strain prone-8. Brain Res. 2006;1090:23–28. doi: 10.1016/j.brainres.2006.02.138. [DOI] [PubMed] [Google Scholar]
- Maclennan KM, Darlington CL, Smith PF. The CNS effects of Ginkgo biloba extracts and ginkgolide B. Prog Neurobiol. 2002;67:235–257. doi: 10.1016/s0301-0082(02)00015-1. [DOI] [PubMed] [Google Scholar]
- Mashayekh A, Pham DL, Yousem DM, Dizon M, Barker PB, Lin DD. Effects of Ginkgo biloba on cerebral blood flow assessed by quantitative MR perfusion imaging: a pilot study. Neuroradiology. doi: 10.1007/s00234-010-0790-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mdzinarishvili A, Kiewert C, Kumar V, Hillert M, Klein J. Bilobalide prevents ischemia-induced edema formation in vitro and in vivo. Neuroscience. 2007;144:217–222. doi: 10.1016/j.neuroscience.2006.08.037. [DOI] [PubMed] [Google Scholar]
- Namiranian K, Koehler RC, Sapirstein A, Doré S. Stroke outcomes in mice lacking the genes for neuronal heme oxygenase-2 and nitric oxide synthase. Curr Neurovasc Res. 2005;2:23–27. doi: 10.2174/1567202052773517. [DOI] [PubMed] [Google Scholar]
- Saleem S, Shah ZA, Urade Y, Doré S. Lipocalin-prostaglandin D synthase is a critical beneficial factor in transient and permanent focal cerebral ischemia. Neuroscience. 2009;160:248–254. doi: 10.1016/j.neuroscience.2009.02.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saleem S, Zhuang H, Biswal S, Christen Y, Doré S. Ginkgo biloba extract neuroprotective action is dependent on heme oxygenase 1 in ischemic reperfusion brain injury. Stroke. 2008;39:3389–3396. doi: 10.1161/STROKEAHA.108.523480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos RF, Galduroz JC, Barbieri A, Castiglioni ML, Ytaya LY, Bueno OF. Cognitive performance, SPECT, and blood viscosity in elderly non-demented people using Ginkgo biloba. Pharmacopsychiatry. 2003;36:127–133. doi: 10.1055/s-2003-41197. [DOI] [PubMed] [Google Scholar]
- Sastre J, Millan A, de la Asuncion J Garcia, Pla R, Juan G, Pallardo, O’Connor E, Martin JA, Droy-Lefaix MT, Vina J. A Ginkgo biloba extract (EGb 761) prevents mitochondrial aging by protecting against oxidative stress. Free Radic Biol Med. 1998;24:298–304. doi: 10.1016/s0891-5849(97)00228-1. [DOI] [PubMed] [Google Scholar]
- Shah ZA, Li RC, Ahmad AS, Kensler TW, Yamamoto M, Biswal S, Doré S. The flavanol (-)-epicatechin prevents stroke damage through the Nrf2/HO1 pathway. J Cereb Blood Flow Metab. 2010;30:1951–1961. doi: 10.1038/jcbfm.2010.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah ZA, Sharma P, Vohora SB. Ginkgo biloba normalises stress-elevated alterations in brain catecholamines, serotonin and plasma corticosterone levels. Eur Neuropsychopharmacol. 2003;13:321–325. doi: 10.1016/s0924-977x(03)00005-1. [DOI] [PubMed] [Google Scholar]
- Snitz BE, O’Meara ES, Carlson MC, Arnold AM, Ives DG, Rapp SR, Saxton J, Lopez OL, Dunn LO, Sink KM, DeKosky ST. Ginkgo biloba for preventing cognitive decline in older adults: a randomized trial. JAMA. 2009;302:2663–2670. doi: 10.1001/jama.2009.1913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Beek TA. Chemical analysis of Ginkgo biloba leaves and extracts. J Chromatogr A. 2002;967:21–55. doi: 10.1016/s0021-9673(02)00172-3. [DOI] [PubMed] [Google Scholar]
- Vogensen SB, Stromgaard K, Shindou H, Jaracz S, Suehiro M, Ishii S, Shimizu T, Nakanishi K. Preparation of 7-substituted ginkgolide derivatives: potent platelet activating factor (PAF) receptor antagonists. J Med Chem. 2003;46:601–608. doi: 10.1021/jm0203985. [DOI] [PubMed] [Google Scholar]
- Walesiuk A, Braszko JJ. Preventive action of Ginkgo biloba in stress- and corticosterone-induced impairment of spatial memory in rats. Phytomedicine. 2007 doi: 10.1016/j.phymed.2007.04.012. [DOI] [PubMed] [Google Scholar]
- Wang J, Doré S. Heme oxygenase 2 deficiency increases brain swelling and inflammation after intracerebral hemorrhage. Neuroscience. 2008;155:1133–1141. doi: 10.1016/j.neuroscience.2008.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Doré S. Heme oxygenase-1 exacerbates early brain injury after intracerebral haemorrhage. Brain. 2007;130:1643–1652. doi: 10.1093/brain/awm095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang SJ, Chen HH. Ginkgolide B, a constituent of Ginkgo biloba, facilitates glutamate exocytosis from rat hippocampal nerve terminals. Eur J Pharmacol. 2005;514:141–149. doi: 10.1016/j.ejphar.2005.03.027. [DOI] [PubMed] [Google Scholar]
- Zeynalov E, Shah ZA, Li RC, Doré S. Heme oxygenase 1 is associated with ischemic preconditioning-induced protection against brain ischemia. Neurobiol Dis. 2009;35:264–269. doi: 10.1016/j.nbd.2009.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Fu S, Liu S, Mao T, Xiu R. The therapeutic effect of Ginkgo biloba extract in SHR rats and its possible mechanisms based on cerebral microvascular flow and vasomotion. Clin Hemorheol Microcirc. 2000;23:133–138. [PubMed] [Google Scholar]
- Zhuang H, Pin S, Christen Y, Doré S. Induction of heme oxygenase 1 by Ginkgo biloba in neuronal cultures and potential implications in ischemia. Cell Mol Biol (Noisy-le-grand) 2002;48:647–653. [PubMed] [Google Scholar]