

Abstract
The synovial membrane (SM) of affected joints in ankylosing spondylitis (AS) is infiltrated by germinal center-like aggregates (foci) of lymphocytes similar to rheumatoid arthritis (RA). We characterized the rearranged heavy chain variable segment (VH) genes in the SM for gene usage and the mutational pattern to elucidate the B lymphocyte involvement in AS.
Cryosections from an AS-derived SM were stained for B and T lymphocytes. B cells were isolated from different areas of a focus. The rearranged VH genes were amplified by semi-nested polymerase chain reaction (PCR) using oligonucleotides specific for the six different VH families and heavy chain joining segments (JHs). PCR products were cloned and sequenced.
Fifty-nine of 70 different heavy chain gene rearrangements were potentially functional. Most of the rearranged genes were mutated (range, 1–15%). Thirty of 70 products had a mutational pattern typical for antigen selection. Most of the rearranged VH genes belonged to the VH3 family (54%), consistent with data from healthy donors and patients with RA, while VH4 genes, in contrast to RA, were identified less frequently (10%) and VH5 genes were over-represented (11%). In contrast to RA, neither VH6 genes nor the autoimmunity-prone VH4-34 were seen, whereas another autoimmunity-prone gene, V3-23, was predominantly used (11%). One VH1-derived and one VH3-derived B cell clone were expanded. CDR3 were shorter and more variable in length than in RA.
Comparable with RA and reactive arthritis, there is a biased repertoire of selected VH genes, whereas the panel of represented genes is different and less clonal expansion was observed.
Keywords: ankylosing spondylitis, B lymphocyte immunology, heavy chain genes, immunoglobulins, somatic mutations
Introduction
AS is an inflammatory rheumatic disease mainly involving the axial skeleton. Forty percent of AS patients develop periphery mono-/oligoarthritis [1].
The SM of inflamed peripheral joints in AS (Fig. 1) in certain cases resembles RA with intense mononuclear cell infiltration forming foci similar to germinal centers (GC) that contain B and T lymphocytes as well as plasma cells [2,3]. Recent immunohistologic analysis of five AS SMs demonstrated B cell rich infiltrates in two cases, whereas no B cells were detected in the three others [4].
Figure 1.
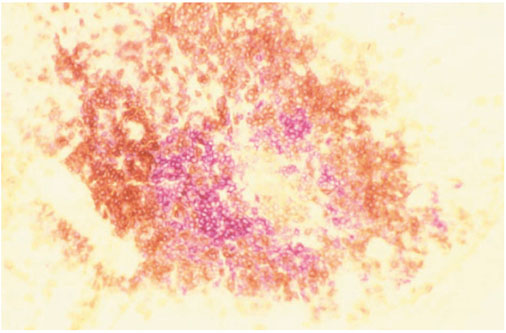
The synovial focus. Double-immunohistological staining (anti-CD20 alkaline phosphatase-anti-alkaline phosphatase and anti-CD3 streptavidin biotin horseradish peroxidase reaction) of 8 μm sections from frozen SM obtained by total joint replacement of an AS patient's right hip (B cells, red staining; T cells, brown staining).
The VH germline repertoire consists of 123 (44 functional) VH segments subdivided into seven VH families (see supplementary material). Antibodies are encoded by one VH segment rearranged to one or several of 25 heavy chain diversity segments (DHs) and one of six JHs [5]. The locus of rearrangement, individual for each B lymphocyte clone, is the CDR3 [6]. Only rearrangements with translatable nucleotide triplets lead to antibody formation, others are nonfunctional. There is a biased use of particular germline VH genes in the VH repertoire. An over-representation of VH4 genes in the autoimmune repertoire has been described [7,8]. A negative selection of VH4 takes place in the peripheral blood lymphocytes from healthy individuals as a means of avoiding autoimmunity [9].
An antigen-driven immune response, characterized by a ratio of mutations leading to amino acid replacement to silent mutations (R:S ratio) >3 [10] within the complementarity determining region (CDR), takes place within the GC of lymphatic organs. Similar clonal development of antibody VH genes has been demonstrated in the SM in RA patients [ 11,12,13, 14,15] and in reactive arthritis (ReA) patients [16]. The B cell subset in AS joints has not yet been characterized. We analyzed the repertoire and distribution of rearranged VH genes to elucidate B lymphocyte involvement in AS, and to investigate similarities to and differences from RA.
Materials and methods
Tissue preparation and staining
A SM of the right hip was obtained from a 57-year-old man, who had suffered from AS for 30 years, requiring total endoprothetic joint replacement. Frozen sections (8 μm) of the immediately snap-frozen tissue sample were screened for B and T cells by double immunohistological staining with anti-CD20 and anti-CD3 monoclonal antibodies using the alkaline phosphatase–anti-alkaline phosphatase and streptavidin biotin horseradish peroxidase technique [17,18]. The stained B lymphocytes from serial sections taken at 50μm intervals were isolated by microdissection, and DNA preparation followed (see supplementary material).
VH PCR
DNA was subjected to a semi-nested PCR for VH genes (see supplementary material). Thirty-five cycles of PCR, using a mix of six oligonucleotides specific for the different VH families as primers at the 5' end and a mix of oligonucleotides corresponding to the known JH overlapping to a part of the intron sequence as primers at the 3' end, were followed by 35 cycles with 1 μl of the first PCR products and individual VH and JH primers (at a more 5' position) (Table 1). A similar PCR without DNA was run as a negative control.
Table 1.
Template generation and sequence analysis
PCR products of the expected 350 base pair (bp) length were detected by standard agarose gel electrophoresis and purified. The purified DNA template was bacterially subcloned followed by plasmid isolation and sequencing. Sequences were analyzed by homology comparison with the EMBL and GenBank gene databases (see supplementary material).
Results
Immunohistological analysis of the inflamed SM revealed foci rich in B and T lymphocytes in a formation similar to GC (Fig. 1) [2]. The B lymphocytes of one focus were isolated from four not directly consecutive sections about 50 μm apart (2c, 3a, 3b, 5c). An estimated number of 150 B lymphocytes could be isolated from each section.
The VH gene usage
Except for the negative control, all VH PCRs led to a product of the expected length (350 bp). All sequences reported in the present paper are accessible on the EMBL database.
Seventy individually rearranged VH genes were characterized by sequence analysis (Table 2): 14 (20%) belonged to the VH1 family, 3 (4%) to VH2, 38 (54%) to VH3, 7 (10%) to VH4, and 8 (11%) belonged to the VH5 family. No sequences corresponded to VH6 (Fig. 2).
Table 2.
The rearranged heavy chain variable segment genes, their functionality, respective germline gene (homology), ratio of mutations leading to amino acid replacement to silent mutations with the number of mutations for the complementarity determining region and the framework region, heavy chain joining segments and CDR3 lengths
| VH | Gene | Functionality | Germline | Homology (%) | R:S ratio CDR | R:S ratio FR | JH | CDR3 (base pairs) |
| 1 | F5-3b6 | nf | DP6 | 97 | 0 | 2/0 | 6 | 26 |
| F5-3a3 | pf | DP7 | 91 | 4/0 | 4/4 | 3 | 24 | |
| F5-2c2 | pf | DP14 | 100 | 0 | 0 | 2 | 30 | |
| F5-2c6 | pf | DP14 | 99 | 0 | 0 | 6 | 30 | |
| 2c-3m4b | pf | DP25 | 89 | 5/6 | 3/10 | 3 | 39 | |
| F5-5c6 | pf | DP25 | 92 | 9/1 | 4/2 | 6 | 39 | |
| F5-2c4* | pf | DP21 | 99 | 0 | 2/1 | 1 or 2 or 4 | 21 | |
| 3a-5m3 | pf | DP21 | 100 | 0 | 1/0 | 5 | 18 | |
| F5-3a6 | pf | DP10 | 98 | 2/0 | 1/0 | 6 | 36 | |
| 5c-4m4 | pf | DP10 | 88 | 9/2 | 11/7 | 4 or 5 | 18 | |
| F5-3a2 | pf | DP75 | 97 | 2/1 | 3/1 | 1 | 27 | |
| F5-5c1 | pf | DP75 | 98 | 0/0 | 2/0 | 1 | 30 | |
| F5-5c2 | nf | DP75 | 96 | 1/2 | 2/0 | Undetermined | 23 | |
| 5c-4m7 | pf | DP75 | 93 | 6/2 | 4/4 | 4 | 30 | |
| 2 | 2c-VH24s | pf | HUMIGVH2e | 97 | 1/1 | 2/0 | 4 | 45 |
| 5c-VH22s | pf | HUMIGVH2e | 99 | 1/1 | 0/0 | 2 | 36 | |
| 5c-VH26s | pf | HUMIGVH2f | 95 | 2/1 | 2/5 | 6 | 15 | |
| 3 | 2c-17m14 | pf | DP54 | 95.5 | 3/2 | 4/2 | 2 or 5 | 30 |
| 2c-19m8 | pf | DP54 | 95.5 | 4/1 | 3/2 | 4 | 12 | |
| 3a-19m6 | pf | DP54 | 95.5 | 3/0 | 5/2 | 4 | 12 | |
| 3b-18m4 | nf | DP54 | 100 | 0/0 | 0/1 | 3 | 23 | |
| 5c-21m1 | nf | DP54 | 96.7 | 5/0 | 1/0 | 5 or 6 | 22 | |
| 2c-18m7 | pf | DP47/V3-23 | 92.4 | 8/2 | 2/4 | 3 | 30 | |
| 2c-21m2 | pf | DP47/V3-23 | 96.9 | 3/0 | 2/2 | 6 | 33 | |
| 2c-21m3 | pf | DP47/V3-23 | 90.1 | 7/6 | 3/3 | 6 | 45 | |
| 3a-19m8 | pf | DP47/V3-23 | 93.3 | 3/2 | 6/4 | 4 | 24 | |
| F5-3a21 | pf | DP47/V3-23 | 92.9 | 2/0 | 4/2 | 5 or 6 | 45 | |
| 5c-19m9 | pf | DP47/V3-23 | 95.5 | 3/3 | 4/0 | 4 | 33 | |
| 5c-21m5 | pf | DP47/V3-23 | 88.4 | 10/1 | 8/6 | 6 | 27 | |
| F5-5c18 | pf | DP47/V3-23 | 91.2 | 6/1 | 3/3 | 3 | 24 | |
| 3b-17m1 | pf | DP46 | 96.9 | 2/1 | 2/1 | 2 | 42 | |
| F5-3b21 | pf | DP46 | 95.9 | 0/1 | 1/1 | 6 | 21 | |
| 2c-20m10 | nf | DP50 | 85.7 | 5/2 | 10/0 | 5 | 26 | |
| 3a-16m4 | pf | DP50 | 95.8 | 4/1 | 0/3 | 1 | 36 | |
| 3b-17m6 | pf | DP50 | 93.3 | 7/2 | 4/2 | 2 or 4 | 36 | |
| 5c-19m7 | nf | DP50 | 87.8 | 7/2 | 5/1 | 4 | 31 | |
| 5c-20m8 | pf | DP50 | 91.1 | 9/3 | 5/3 | 5 | 24 | |
| 2c-17m9 | pf | DP49 | 88.8 | 3/4 | 8/10 | 2 | 6 | |
| 2c-20m9 | pf | DP49 | 93.8 | 5/1 | 6/2 | 5 | 24 | |
| 5c-19m4 | nf | DP49 | 94.0 | 5/3 | 5/0 | 4 | 30 | |
| 2c-20m8 | pf | DP51 | 98.2 | 0/0 | 3/1 | 5 | 27 | |
| 3b-16m4 | pf | DP51 | 93.3 | 6/2 | 4/3 | 1 | 27 | |
| 5c-20m6 | nf | DP51 | 90.2 | 13/2 | 1/6 | 5 | 29 | |
| 2c-19m2 | pf | DP31 | 97.8 | 2/2 | 1/0 | 4 | 18 | |
| F5-3a17 | pf | DP31 | 88 | 5/2 | 2/1 | 2 | 36 | |
| 3a-18m2 | pf | DP31 | 93.6 | 4/0 | 2/1 | 3 | 12 | |
| 3a-19m2 | pf | DP31 | 97.3 | 2/1 | 0/1 | 4 | 30 | |
| 2c-18m3 | pf | DP35 | 100 | 0/0 | 0/0 | 3 | 24 | |
| 2c-19m7 | pf | DP35 | 94.6 | 4/2 | 4/2 | 4 | 33 | |
| 5c-21m8 | nf | DP35 | 94.6 | 3/1 | 0/5 | 6 | 31 | |
| 3a-20m7 | pf | DP77 | 88.3 | 12/2 | 9/4 | 5 | 33 | |
| F5-5c16 | pf | DP77 | 94.3 | 4/0 | 6/2 | 1 | 27 | |
| 2c-21m6 | nf | DP52 | 98.2 | 0/0 | 2/0 | 6 | 25 | |
| 3a-18m5 | pf | DP29 | 89.9 | 6/1 | 9/2 | 3 | 39 | |
| 5c-21m2 | pf | DP53 | 97.3 | 3/0 | 2/1 | 6 | 12 | |
| 4 | 3a-VH43b | pf | HUMIGHCAK | 97 | 1/2 | 4/1 | 3 | 27 |
| 5c-VH44a | pf | HUMIGHCAK | 95 | 8/1 | 3/1 | 6 | 24 | |
| 3b-VH45a | pf | DP63 | 93 | 4/0 | 7/4 | 4 | 23 | |
| 5c-VH44c | pf | DP63 | 99 | 0/1 | 0/0 | 4 | 27 | |
| 5c-VH41a | nf | DP71 | 99 | 0/0 | 3/0 | 1 | 12 | |
| 5c-VH42c | pf | HUMIGHCAG | 98 | 1/0 | 3/0 | 2 | 30 | |
| 5c-VH46b | pf | HUMIGHCAG | > 99 | 1/0 | 0/0 | 6 | 45 | |
| 5 | 2c-VH53b | pf | DP73 | 97 | 3/0 | 4/0 | 3 | 45 |
| 2c-VH53c | pf | DP73 | 95 | 2/1 | 8/0 | 3 | 48 | |
| 2c-VH54c | pf | DP73 | 85 | 12/1 | 16/6 | 4 | 33 | |
| 2c-VH54b | pf | DP73 | 95 | 4/1 | 4/3 | 4 or 6 | 39 | |
| 3a-Vh53s | pf | DP73 | 96 | 5/0 | 3/4 | 3 | 66 | |
| 3a-VH54s | pf | DP73 | 100 | 0/0 | 0/0 | 4 | 36 | |
| 3a-VH56s | nf | DP73 | 90 | 13/1 | 7/0 | 5 or 6 | 31 | |
| 3b-VH53s | pf | DP73 | 95 | 4/1 | 5/2 | 3 or 5 | 12 |
Figure 2.
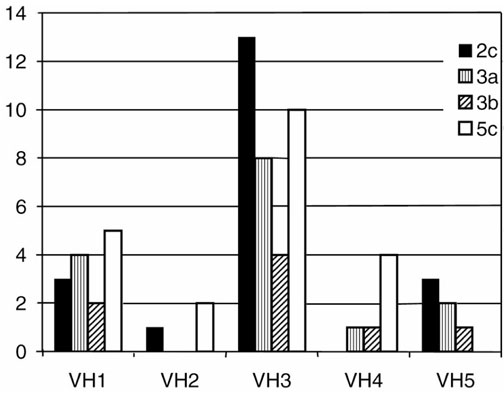
Distribution of the VH families among the PCR products obtained from different slides (2c, 3a, 3b and 5c). Bars for each section represent the respective number of products for each indicated VH family.
Twelve genes in all were nonfunctionally rearranged, with the highest proportion in VH3 (18%), about 14% in VH1, VH4 and VH5, and none in VH2.
VH1 genes
Fourteen individual rearrangements represented VH1 genes. The most often represented VH1 gene was DP75 (four products) followed by DP10, DP14, DP21 and DP25 (two products each). Out of 14 distinct VH1–DH–JH sequences, 2 were nonfunctional (14%). CDR3 lengths varied between 18 and 39 bp (mean, 28.5 bp).
Four of the VH1 products (F5-2c4, F5-3a4, F5-3b4 and 5c-4m6) deriving from different slides had an identically rearranged CDR3 and consecutively represent members of an expanding B lymphocyte clone (Fig. 3a). With 99% homology to DP21, they shared three mutations within the framework region (FR).
Figure 3.

Mutated nucleotide triplets and amino acid sequence of expanded B cell clones (a) VH1 and (b) VH3 in comparison with their respective gl gene (short tables). The amino acid position and the CDR positions are marked. Dashes indicate identity to the corresponding character on top. Amino acid replacements are indicated in the last line. (For whole sequences, see supplementary Figure 1.)
Five productively rearranged genes were highly mutated with a range of 12 (F5-3a3) to 29 mutations to their respective germline (gl) gene. The R:S ratio for the CDR was >3 and lower for the FR, indicating antigen-driven mutation [10].
VH2 genes
Three different VH2 rearrangements were determined. All were potentially functional and > 95% homologous to the respective gl gene. The number of mutations ranged from 2 (5c-VH22s) to 10 (5c-VH26s). R:S ratios were quite low, with a maximum of 2.0 in the CDR (5c-VH26s). CDR3 lengths varied between 15 and 45 bp (mean, 32 bp).
VH3 genes
Thirty-eight different VH3 rearrangements were characterized: eight were most homologous to DP47/V3-23 (21%), five to DP50 (13%), four each to DP54 and DP31 (11%), respectively, and three each to DP49, DP35, DP51 and DP77 (8%), respectively. Seven (18.4%) VH3 genes were nonfunctionally rearranged. CDR3 lengths varied from 6 to 35 bp (mean, 27.2 bp).
One case of clonal expansion within the VH3 genes was observed, represented by the VH3–JH4 rearrangements 3a-19m6 and 2c-19m8 (Fig. 3b ). These genes, with 95.5% homology to DP54, shared four replacement mutations, and six mutations each were localized at different positions.
Twenty-three of the VH3 sequences were highly mutated (10–27 mutations). Except for one case, most of the mutations were localized in the CDR with R:S ratio >3. Three of seven nonfunctionally rearranged genes also revealed a pattern of antigen-induced mutation.
VH4 genes
Seven different VH4 rearrangements could be characterized; six were potentially functional. CDR3 lengths ranged from 12 to 45 bp (mean, 28 bp).
The gl genes HUMIGHCAK, HUMIGHCAG and DP63 were represented twice each, and DP71 once. No homology to DP64/VH4-34/VH4.21 was seen. Five of the sequences had few mutations (one to eight) with a R:S ratio higher in the FR than in the CDR. Two VH4 rearrangements had a R:S ratio >3 in the CDR (5c-VH44a and 3b-VH45a).
VH5 genes
Eight distinct VH5 rearrangements could be determined; seven were potentially functional. CDR3 lengths ranged from 12 to 66 bp (mean, 39.8 bp).
The 3a-VH54s gene was unmutated. There were 7–35 mutations in all other sequences. For five sequences (four were potentially functional), the mutational pattern revealed antigen induction.
Discussion
VH PCR
Analysis of rearranged VH genes from B lymphocyte DNA by PCR and sequencing enabled characterization of the clonal expansion and mutational patterns without bias due to cell isolation procedures. By semi-nested PCR, otherwise used for single-cell analysis [19], one sole VH–DH–JH rearrangement might be identified. Besides the extreme sensitivity, there is a high risk of artificial amplifications because of contamination. A negative control without DNA was run with each PCR as a precaution: no contamination was detected. We cannot exclude the fact that highly mutated genes are amplified less efficiently [20]. However, since the respective data concerning RA and ReA have been established by similar PCR approaches [11,12,13 ,14,15,16 ], a comparison is possible.
The VH gene usage
Within 70 VH–DH–JH rearrangements characterized in this study, all known human VH families except VH6 were represented. The descending order of representation was: VH3 (54%), VH1 (20%), VH5 (11%), VH4 (10%), VH2 (4%). Compared with the germline representation [5], an over-representation of VH5 and an under-representation of VH4 genes were remarkable. The representation of VH1, VH2 and VH3 genes corresponded to their respective germline representation. Our previous studies of the VH gene repertoire in RA SM [13] and studies using a hybridoma approach [8] revealed an over-representation of VH4 and a biased set of represented VH4 genes. A positive selection of VH4 genes, particularly within the plasma cell subset, was recently demonstrated in RA and ReA [16]. The DP64/VH4-34/VH4.21 gene encoding anti-I and anti-i specificity of cold agglutinins [21,22], as well as anti-DNA antibodies in lupus erythmatosus [23], was not detected within the VH4 genes from AS SM.
Another autoimmunity-prone gene, the DP47/V3-23 gene, was also predominant in AS. All of eight V3-23-derived genes were potentially functional rearrangements with >3% mutations to the gl sequence, and four reveal a R:S ratio >3 within the CDR that indicates a selection by antigen. V3-23 was also predominant in RA SM [24]. A comparison of the peripheral repertoire revealed that V3-23 was more often somatically mutated in RA than in healthy controls [9,25 ,26].
Except for VH4, VH5 and VH6, the representation of VH genes was similar to results from the peripheral VH repertoire from healthy donors [ 27,9] as well as from a RA patient [28].
Clonal expansion
Only two cases of clonal expansion characterized by identically rearranged segments were detected on different slides. An accumulation of further point mutations in the course of intraclonal diversification was seen in two offspring of a VH3 clone (both share four mutations and differ in six point mutations). Studies on plasma cells in ReA and RA SMs [16 ] reveal clonal expansion. In RA SMs, clonal B lymphocyte expansion in different areas of the tissue samples [11,15] and recirculation within different joints and blood were reported [14].
The CDR3
The CDR3 lengths of genes from the AS samples were short, with a median of 27.2 nucleotides. A high variability, particularly in the productively rearranged subset and in the VH5 family, was noted. A similarly high variability was shown for polyreactive antibodies and rheumatic factors in RA [29]. Our data from RA SM revealed almost homogeneous CDR3 lengths of intermediate size [13]. CDR3 were much longer in the healthy peripheral repertoire, particularly within nonfunctional rearrangements (mean, 54 nucleotides) [9].
Mutational pattern
Only nine rearrangements were not mutated. R:S ratios >3 within the CDR were observed in 30 genes (two were nonfunctional). Members of the VH1, VH3, VH4 and VH5 families carried more somatic mutations than the VH2 genes.
Mutational pattern and frequency determine about one half of the characterized genes as memory B cells that probably stay in contact with persistent antigens in the micro-environment of the synovial foci [30]. A similar accumulation of memory cells in RA SM has been reported [12,31].
Besides the highly mutated genes already described, one half of the characterized genes showed no evidence of antigen-induced mutations with low R:S ratios within the CDR. There was a bias to silent mutations in four highly mutated genes (2c-3m4b, 2c-21m3, 3a-19m8 and 2c-17m9). This pattern of mutations has been described for cold agglutinins [32], as well as for rheumatic factors, as a mechanism of selection against amino acid replacement to prevent the generation of high-affinity autoantibodies [33].
Nonfunctional rearrangements
Except for VH2 (none) and VH3 (18%), nonfunctional rearrangements had an equal proportion of 13–14% in each family within the genes characterized in the present paper. Except for a bias to VH4, a similar overall proportion of nonfunctional rearrangements was seen in the peripheral blood of healthy donors using single-cell PCR of genomic DNA [9]. In contrast, no bias to elevated CDR3 lengths was seen.
Conclusion
An active immune reaction with GC formation occurs in the inflamed SM in AS. B cell affinity maturation with generation of somatically mutated antibody-coding genes characteristic for memory cells occurs in these GC formations, indicating an antigen-driven response. Comparable with RA and ReA, there is a biased repertoire of selected VH genes, whereas the panel of represented genes is different and clonal expansion was observed less frequently.
Supplementary material
Introduction
By sequence homology of at least 80%, the VH gene segments are classified into seven families. The VH1 family has 14 members (9 genes with open reading frames), VH2 has 4 (3 genes with open reading frames), VH3 has 65 (22 genes with open reading frames), VH4 has 32 (7 genes with open reading frames), VH5 has 2, VH6 has 1, and VH7 has 5 members (1 gene with an open reading frame), respectively [5].
Materials and methods
DNA preparation
After the addition of 40 μl H2O, 0.1 volume of 10 × PCR buffer (mentioned later), 1% Triton X-100 and 0.5 μl Pro-teinase K (100 μg/ml; Merck, Darmstadt, Germany) was added and mixed. The volume was adapted to 80 μl by addition of H2O. The sample was incubated over 45 min at 55°C. After inactivation of proteinase (95°C), 25 μl of the solution was subjected to the first PCR.
Seminested VH PCR
First PCR. An oligonucleotide mix of VH1, VH2, VH3, VH4, VH5, VH6 and an oligonucleotide mix of JH-Intron 1,2-4,5 and JH-Intron 3,6 as primers (final concentration, 0.125 μM each), 200 μM dNTP, 2 mM MgCl2, 1 U Goldstar Taq-polymerase (Eurogentec, Seraing, Belgium), and the manufacturer's reaction buffer. First cycle: 5 min denaturation at 95°C, 3 min annealing at 58°C and 90 second extension at 72°C; cycles 2–35, 80 second denaturation, 30 second annealing, and 90 second extension, with a final extension of 5 min.
Second PCR. One microliter of the product of the first PCR with individual oligonucleotide primers VH1–VH6 and JH1–JH6, under the same conditions as the first PCR except for annealing for VH1, VH2, VH5, VH6 at 58°C, and for VH3 and VH4 at 63°C.
DNA purification and plasmid ligation were performed with commercial kits following the manufacturer's instructions: DNA purification from the agarose-gel with the QUIAquick kit (Quiagen, Hilden, Germany), and bacterial cloning with the TA-cloning kit (Invitrogen, Leek, The Netherlands).
For DNA sequence homology search and sequence comparison, DNASIS software (Hitachi Europe, Olivet, France), and EMBL Nucleotide Sequence Submissions (European Bioinformatics Institute, Wellcome Trust Genome Campus, Cambridge, UK) were used. The GenBank (National Institutes of Health, Bethesda, MD, USA) was also used.
Abbreviations
AS = ankylosing spondylitis; CDR = complementarity determining region; DH = heavy chain diversity segment; FR = framework region; GC = germinal centers; gl = germline; JH = heavy chain joining segment; PCR = polymerase chain reaction; RA = rheumatoid arthritis; ReA = reactive arthritis; SM = synovial membrane; VH = heavy chain variable segment.
Supplementary Figure 1.

Complete nucleotide and amino acid sequences of expanded B cell clones (a) VH1 and (b) VH3 in comparison with their respective gl gene. The amino acid position and the CDR positions are marked. Dashes indicate identity to the corresponding character on top. Amino acid replacements are indicated in the last line.
Acknowledgments
Acknowledgement
This work was supported by Deutsche Forschungsgemeinschaft Ga320, 3-1.
References
- Gran JT, Skomsvoll JF. The outcome of ankylosing spondylitis: a study of 100 patients. Br J Rheumatol. 1997;36:766–771. doi: 10.1093/rheumatology/36.7.766. [DOI] [PubMed] [Google Scholar]
- Zdichavsky M, Schorpp C, Nickels A, Koch B, Pfreundschuh M, Gause A. Analysis of bcl-2+ lymphocyte subpopulations in inflammatory synovial infiltrates by a double-immunostaining technique. Rheumatol Int. 1996;16:151–157. doi: 10.1007/BF01419728. [DOI] [PubMed] [Google Scholar]
- Chang CP, Schumacher HR., Jr Light and electron microscopic observations on the synovitis of ankylosing spondylitis. Semin Arthritis Rheum. 1992;22:54–65. doi: 10.1016/0049-0172(92)90049-j. [DOI] [PubMed] [Google Scholar]
- Cunnane G, Bresnihan B, Fitzgerald O. Immunohistologic analysis of peripheral joint disease in ankylosing spondylitis. Arthritis Rheum. 1998;41:180–182. doi: 10.1002/1529-0131(199801)41:1<180::AID-ART24>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- Matsuda F, Ishii K, Bourvagnet P, Kuma K, Hayashida H, Miyata T, Honjo T. The complete nucleotide sequence of the human immunoglobulin heavy chain variable region locus. J Exp Med. 1998;188:2151–2162. doi: 10.1084/jem.188.11.2151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequences of Proteins of Immunological Interest, edn 5 Bethesda: NIH Publication, 1991.
- Pascual V, Capra JD. VH4-21, a human VH gene segment over-represented in the autoimmune repertoire. Arthritis Rheum. 1992;35:11–18. doi: 10.1002/art.1780350103. [DOI] [PubMed] [Google Scholar]
- Brown CM, Longhurst C, Haynes G, Plater Zyberk C, Malcolm A, Maini RN. Immunoglobulin heavy chain variable region gene utilization by B cell hybridomas derived from rheumatoid synovial tissue. Clin Exp Immunol. 1992;89:230–238. doi: 10.1111/j.1365-2249.1992.tb06937.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brezinschek HP, Brezinschek RI, Lipsky PE. Analysis of the heavy chain repertoire of human peripheral B cells using single-cell polymerase chain reaction. J Immunol. 1995;155:190–202. [PubMed] [Google Scholar]
- Chang B, Casali P. The CDR1 sequences of a major proportion of human germline Ig VH genes are inherently susceptible to amino acid replacement. Immunol Today. 1994;15:367–373. doi: 10.1016/0167-5699(94)90175-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gause A, Gundlach K, Zdichavsky M, Jacobs G, Koch B, Hopf T, Pfreundschuh M. The B lymphocyte in rheumatoid arthritis: analysis of rearranged Vk genes from B cells infiltrating the synovial membrane. Eur J Immunol. 1995;25:2775–2782. doi: 10.1002/eji.1830251010. [DOI] [PubMed] [Google Scholar]
- Gause A, Gundlach K, Carbon G, Daus H, Trümper L, Pfreundschuh M. Analysis of VH gene rearrangements from synovial B cells of patients with rheumatoid arthritis reveals infiltration of the synovial membrane by memory B cells. Rheumatol Int. 1997;17:145–150. doi: 10.1007/s002960050026. [DOI] [PubMed] [Google Scholar]
- Voswinkel J, Trümper L, Carbon G, Hopf T, Pfreundschuh M, Gause A. Evidence for a selected humoral immune response encoded by VH4 family genes in the synovial membrane of a patient with rheumatoid arthritis. Clin Exp Immunol. 1996;106:5–12. doi: 10.1046/j.1365-2249.1996.d01-806.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voswinkel J, Weisgerber K, Pfreundschuh M, Gause A. The B lymphocyte in rheumatoid arthritis: recirculation of B lymphocytes between different joints and blood. Autoimmunity. 1999;31:25–34. doi: 10.3109/08916939908993856. [DOI] [PubMed] [Google Scholar]
- Schröder AE, Greiner A, Seyfert C, Berek C. Differentiation of B cells in the non-lymphoid tisssue of the synovial membrane of patients with rheumatoid arthritis. Proc Natl Acad Sci USA. 1996;93:221–225. doi: 10.1073/pnas.93.1.221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim HJ, Krenn V, Steinhauser G, Berek C. Plasma cell development in synovial germinal centers in patients with rheumatoid and reactive arthritis. J Immunol. 1999;162:3053–3062. [PubMed] [Google Scholar]
- Mason DY, Abdulaziz Z, Falini B, Stein H. Single and double immunoenzymatic techniques for labeling tissue sections with monoclonal antibodies. Ann NY Acad Sci. 1983;420:127–133. doi: 10.1111/j.1749-6632.1983.tb22196.x. [DOI] [PubMed] [Google Scholar]
- Burgess R, Hyde K, Maguire PJ, Kelsey PR, Yin JA, Geary CG. Two-colour immunoenzymatic technique using sequential staining by APAAP to evaluate two cell antigens. J Clin Pathol. 1992;45:206–209. doi: 10.1136/jcp.45.3.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth J, Daus H, Gause A, Trumper L, Pfreundschuh M. Detection of Epstein-Barr virus DNA in Hodgkin and Reed-Sternberg cells by single cell PCR. Leuk Lymphoma. 1994;13:137–142. doi: 10.3109/10428199409051664. [DOI] [PubMed] [Google Scholar]
- Aubin J, Davi F, Nguyen Salomon F, Leboeuf D, Debert C, Taher M, Valensi F, Canioni D, Brousse N, Varet B, et al. Description of a novel FR1 IgH PCR strategy and its comparison with three other strategies for the detection of clonality in B cell malignancies. Leukemia. 1995;9:471–479. [PubMed] [Google Scholar]
- Pascual V, Victor K, Spellerberg M, Hamblin TJ, Stevenson FK, Capra JD. VH restriction among human cold agglutinins. The VH4-21 gene segment is required to encode anti-I and anti-i specificities. J Immunol. 1992;149:2337–2344. [PubMed] [Google Scholar]
- Schutte ME, Van Es JH, Silberstein LE, Logtenberg T. VH4.21-Encoded natural autoantibodies with anti-i specificity mirror those associated with cold hemagglutinin disease. J Immunol. 1993;151:6569–6576. [PubMed] [Google Scholar]
- Stevenson FK, Longhurst C, Chapman CJ, Ehrenstein M, Spellerberg MB, Hamblin TJ, Ravirajan CT, Latchman D, Isenberg D. Utilization of the VH4-21 gene segment by anti-DNA antibodies from patients with systemic lupus erythematosus. J Autoimmun. 1993;6:809–825. doi: 10.1006/jaut.1993.1066. [DOI] [PubMed] [Google Scholar]
- Berek C, Kim HJ. B-Cell activation and development within chronically inflamed synovium in rheumatoid and reactive arthritis. Semin Immunol. 1997;9:261–268. doi: 10.1006/smim.1997.0076. [DOI] [PubMed] [Google Scholar]
- Stewart AK, Huang C, Stollar BD, Schwartz RS. High-frequency representation of a single VH gene in the expressed human B cell repertoire. J Exp Med. 1993;177:1227–1227. doi: 10.1084/jem.177.4.1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki I, Pfister L, Glas A, Nottenburg C, Milner EC. Representation of rearranged VH gene segments in the human adult antibody repertoire. J Immunol. 1995;154:3902–3911. [PubMed] [Google Scholar]
- Huang C, Stewart AK, Schwartz RS, Stollar BD. Immunoglobulin heavy chain gene expression in peripheral blood B lymphocytes. J Clin Invest. 1992;89:1331–1343. doi: 10.1172/JCI115719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang SC, Jiang R, Hufnagle WO, Furst DE, Wilske KR, Milner EC. VH usage and somatic hypermutation in peripheral blood B cells of patients with rheumatoid arthritis (RA). Clin Exp Immunol. 1998;112:516–527. doi: 10.1046/j.1365-2249.1998.00580.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pascual V, Victor K, Randen I, Thompson K, Steinitz M, Forre O, Fu SM, Natvig JB, Capra JD. Nucleotide sequence analysis of rheumatoid factors and polyreactive antibodies derived from patients with rheumatoid arthritis reveals diverse use of VH and VL gene segments and extensive variability in CDR-3. Scand J Immunol. 1992;36:349–362. doi: 10.1111/j.1365-3083.1992.tb03108.x. [DOI] [PubMed] [Google Scholar]
- Berek C. The development of B cells and the B-cell repertoire in the microenvironment of the germinal center. Immunol Rev. 1992;126:5–19. doi: 10.1111/j.1600-065x.1992.tb00628.x. [DOI] [PubMed] [Google Scholar]
- Krenn V, Hensel F, Kim HJ, Souto Carneiro MM, Starostik P, Ristow G, Konig A, Vollmers HP, Muller-Hermelink HK. Molecular IgV(H) analysis demonstrates highly somatic mutated B cells in synovialitis of osteoarthritis: a degenerative disease is associated with a specific, not locally generated immune response. Lab Invest. 1999;79:1377–1384. [PubMed] [Google Scholar]
- Borretzen M, Chapman C, Stevenson FK, Natvig JB, Thompson KM. Structural analysis of VH4-21 encoded human IgM allo- and autoantibodies against red blood cells. Scand J Immunol. 1995;42:90–97. doi: 10.1111/j.1365-3083.1995.tb03630.x. [DOI] [PubMed] [Google Scholar]
- Borretzen M, Randen I, Zdarsky E, Forre O, Natvig JB, Thompson KM. Control of autoantibody affinity by selection against amino acid replacements in the complementarity-determining regions. Proc Natl Acad Sci USA. 1994;91:12917–12921. doi: 10.1073/pnas.91.26.12917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomlinson IM, Walter G, Marks JD, Llewelyn MB, Winter G. The repertoire of human germline VH sequences reveals about fifty groups of VH segments with different hypervariable loops. J Mol Biol. 1992;227:776–798. doi: 10.1016/0022-2836(92)90223-7. [DOI] [PubMed] [Google Scholar]
- van der Maarel S, van Dijk KW, Alexander CM, Sasso EH, Bull A, Milner EC. Chromosomal organization of the human VH4 gene family. Location of individual gene segments. J Immunol. 1993;150:2858–2868. [PubMed] [Google Scholar]
- Andris JS, Brodeur BR, Capra JD. Molecular characterization of human antibodies to bacterial antigens: utilization of the less frequently expressed VH2 and VH6 heavy chain variable region gene families. Mol Immunol. 1993;30:1601–1616. doi: 10.1016/0161-5890(93)90452-h. [DOI] [PubMed] [Google Scholar]