

Abstract
Platelets are the main cellular component in blood responsible for maintaining the integrity of the cardiovascular system via hemostasis. Platelet dysfunction contributes to a wide range of obvious pathologic conditions, such as bleeding or thrombosis, but normal platelet function is also linked to diseases not immediately associated with hemostasis or thrombosis, such as cancer.Since the description of Trousseau’s syndrome in 1865 various experimental and clinical evidences have detailed the interaction of platelets with primary tumors and circulating metastatic tumor cells. Observations have suggested that platelets not only augment the growth of primary tumors via angiogenesis but endow tumor cells physical and mechanical support to evade the immune system and extravasate to secondary organs, the basis of metastatic disease.Many laboratory and animal studies have identified specific targets for anti-platelet therapy which may be advantageous as adjuncts to current cancer treatments. In this review, we summarize important platelet properties which influence tumorigenesis including primary tumor growth and metastasis at the molecular level. The studies provide a link between the well-studied paradigms of platelet hemostasis and tumorigenesis.
Keywords: angiogenesis, experimental metastasis, hemostasis, primary tumor, spontaneous metastasis
Despite major advancements in the basic biology of cancer and new therapeutic interventions, cancer still remains one of the deadliest diseases of the modern age. Over the last few decades, advances in the field of basic and clinical sciences have led to the recognition of hemostatic and coagulation systems in the growth and spread of different cancers in mouse models, as well as in human patients. Various distinct proteins originally described to participate in hemostasis are now found to be involved in different steps of cancer progression (Figure 1). The key mechanisms, whereby hemostatic and coagulation systems cooperate are (a) platelets along with coagulation factors interacting with tumor cells to make platelet-tumor cell emboli aiding tumor cell extravasation to the metastatic niche, (b) a platelet cloak around tumor cells protecting them from natural killer cell cytotoxic activity, and (c) platelets storing various growth factors, proteases, and small molecules which help in tumor growth, invasion, and angiogenesis.In this review, we discuss the role of various platelets factors in tumorigenesis via these mechanisms. We have included thrombin and fibrinogen given their importance to the platelet response but recognize many other coagulation factors not discussed are also important for cancer.
Figure 1.

Interactions of platelets, coagulation, and tumor cells in tumorigenesis. Schematic diagram showing the interplay among various proteins; platelet receptors, coagulation proteins, and tumor cells interacting in the process of tumorigenesis.
Platelet involvement in tumor cell dissemination
Armand Trousseau in 1865 described some patients with unusual migratory thromboses. These patients developed visceral malignancy later.Now Trousseau’s syndrome is explained as a thrombotic event preceding the diagnosis of an occult visceral malignancy and diagnosed from an initial intravascular coagulopathy, platelet-rich microthrombi, microangiopathic hemolytic anemia, or thromboembolic problems.1 For the homeostasis of the vasculature it is crucial to maintain a normal platelet count in the blood.Experimental thrombocytopenia in mice inducedby neuraminidase/anti-platelet serum resulted in a 50% reduction in experimental metastasis and this could be reversed by transfusion of platelet-rich plasma transfusion.2 Nf-E2 knockout mice (SCID background) with extreme thrombocytopenia have a significant reduction (94%) in metastatic burden in experimental metastasis models.3 Others have shown that intra-venous injection of some tumor cells may cause significant thrombocytopenia (50%-70%) in mice.4 Tumor cells which aggregate platelets in vitro produce more lung metastases than tumor cells lacking such ability, illustrating the platelet activating potential of some tumor cells.5,6 Taken together these seminal observations suggest a robust interaction between circulating platelets and tumor cells.
After activation, platelets release small vesicles, called microparticles or microvesicles. Platelet microparticles are small in size (0.05μm – 1μm) with a defined plasma membrane and express selected platelet membrane and cellular proteins.7 Lewis lung carcinoma cells treated with platelet-derived microparticles have increased metastastic potential in syngenic mouse models.8 Platelet microparticles increase invasive potential by increasing adhesion, proliferation, chemotaxis and survival of breast cancer cell lines MDA-231 and BT-549, and the prostate cancer cell line CL-1. In the presence of microparticles a number of cellular events have been documented including upregulation of CXCR4, MAPK p42/44, MMP-2, and MMP-9 along with AKT phosphorylation.9,10 Like the platelet, the platelet microparticle facilitates tumorigenesis.
Platelet receptors and ligands supporting tumor cell growth and survival
Tumor cells contain various membrane receptors which can bind directly to platelets and mediate tumor cell-platelet binding and activate platelets (Figure 2). Flow cytometry, fluorescence microscopy, and intravital microscopy have revealed the presence of platelet-tumor cell aggregates in vitro and in vivo.11,12
Figure 2.
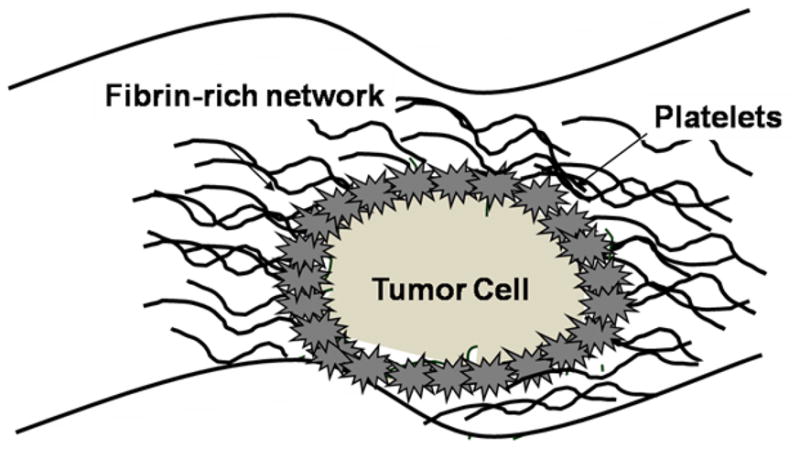
Interaction of platelets, coagulation and tumor cells. Cartoon representation showing some of the molecules implicated for tumor cells and platelets to promote interaction and influence tumor cell growth and survival.
P-selectin is an adhesion receptor found in the α-granules of platelets and Weibel-Palade bodies of endothelial cells.13 After platelet activation P-selectin appears on the platelet surface and aids the recruitment of other circulating platelets and leukocytes. Chondroitin sulfate glycosaminoglycans on the surface of human MDA-MET cells and murine 4T1 cells have been shown to bind selectively P-selectin.14 It has also been suggested that platelet P-selectin recognizes sulfated galactosylceramide SM2, SM3, and SM4 on MC-38 cells and sulfatide removal results in inhibition of in vitro platelet P-selectin binding to MC-38 cells and reduced syngenic experimental metastasis in vivo.15 Experimental metastasis and subcutaneously implanted tumor growth was reported to be reduced in P-selectin deficient mice and in an immunocompetent model with MC-38 colon carcinoma cells and B16 melanoma cells.16,17 Not only is the rate of tumor cell homing to lungs diminished in P-selectin deficient mice, but tumor cells fail to make aggregates with platelets resulting in a decreased number of metastatic nodules in the lungs of P-selectin deficient mice.16
A selectin ligand mimicry peptide, IELLQAR, has been found to have an inhibitory effect on B16 induced experimental metastasis.18 An inhibitor of sialyl Lewis X, such as AcGnG-NM, not only reduces binding of tumor cells to selectin coated surfaces, activated platelets, and TNF-α activated endothelial cells, but also diminishes experimental metastasis in SCID mice.19 Heparin inhibits the binding of P-selectin to its receptors and has been shown to inhibit experimental metastasis in syngenic mouse models.20 Human platelets also express MAPK p38α, aserine threonine kinase, and the expression of MAPK p38α is directly linked to platelet P-selectin expression.Mice lacking MAPK p38α are not viable, but heterozygousp38α+/- mice have reduced experimental metastasis with no effect on primary tumor growth.21 More recently, a role for P-selectin using models of spontaneous tumor metastasis has been presented.22 Thus, through a wide array of studies it can be concluded P-selectin facilitates direct binding to tumor cells and augments tumor metastasis.
The platelet receptor, glycoprotein (GP) Ib-IX supports adhesion of platelets on a compromised vascular wall and, as such, is a key initiator of the platelet paradigm in hemostasis.23 We reported that B16F10 mouse melanoma cell metastasis was reduced 15-fold in GP Ib-IX deficient mouse colonies suggesting an important role for adhesion in a syngenic mouse model.24 However, overexpression of the polyoma middle T antigen in mouse mammary tissue and lung metastasis were not affected by the absence of platelet GP Ib-IX in a model of spontaneous tumor formation (Jain and Ware, unpublished observation).Confounding results have been described with the administration of the anti-GP Ib-IX antibodies and the opposite effect, namely increased colonization of the lung.25 The genetic absence of the platelet collagen receptor, GP VI, is also associated with a 50% reduction in experimental metastasis.26 However, primary tumor growth and angiogenesis was not altered in GP VI deficient mice.26
Although GP Ib-IX is widely considered to be a platelet-specific complex, several studies have suggested the expression of GP Ib-IX subunits by a variety of tumor cells.27–29 In examining the expression of the major subunit of the GP Ib-IX complex, the α-subunit of GP Ib, in lysates of commonly used human tumor cell lines we have been unable to document the presence of GP Ibα (Figure 3) At this time we conclude the expression of GP Ib-IX by cancer cells is not a common mechanism contributing to tumor formation or metastasis.
Figure 3.

Western blot analysis of human tumor cell lines for the presence of human GP Ibα antigen. Also shown are normal mouse and human platelet lysates for representative signals. The blot was reacted with an anti-α-tubulin antibody as a positive control. Human GP Ibα antigen is observed in lysates from purified human platelets (Hu Platelets) and noticeably absent in all other samples (anti-human GPIbα monoclonal antibody, LJ-Ib10, kindly provided by Zaverio Ruggeri, The Scripps Research Institute).mBSS, mouse platelet lysate missing GPIbα; C57, normal mouse platelet lysate; Hu Platelets, human platelet lysate. All other lanes contain lysate from the indicated human tumor cell line.
Von Willebrand factor (VWF) is a key major ligand for the platelet GP Ib-IX complex. Lewis lung carcinoma and B16-B6 mediated experimental metastasis was increased 2-fold and 5-fold, respectively, in VWF deficient mice.30 Lung colonization of tumor cells was increased 1–4 hr post injection tumor cells in VWF deficient mice suggesting VWF may be responsible for tumor cell clearance in the circulation. VWF deficiency did not have any effect on the growth of primary tumors. However, it has also been reported that an anti-VWF antibody protects mice from experimental metastasis in mouse models.31 It is possible that in the absence of VWF, platelet GP Ib-IX availability is increased resulting in increased experimental metastasis. More definitive experimental proof is required to test this possibility.
Integrin αIIbβ3 (GP IIb-IIIa) is the most abundant receptor on the platelet surface.It participates in hemostasis by bridging platelet/platelet interactions via the ligand, fibrinogen.32 GP IIb-IIIa inhibition by the monoclonal antibody 10E5 has been reported to diminish binding of CT26 and HCT28 cells to platelets in vitro.31 Integrin β3−/ − mice show a reduction in B16F10 melanoma induced osteolytic experimental metastasis and reduced osteolytic bone invasion, both reversed by bone marrow transplantation of β3+/+ marrow.33 Antibody inhibition of GP IIb-IIIa diminishes tumor cell adhesion on extracellular matrix under flow conditions suggesting a role for GP IIb-IIIa in platelet–tumor cell emboli extravasation.34 c7E3 (ReoProTM) a mouse-human chimeric antibody for GP IIb-IIIa has anti-angiogenic and anti-tumor properties in mouse models.35 A single treatment of this antibody in a xenograft model of SCID mice reduces experimental metastasis significantly. In addition, c7E3 also inhibits VEGF secretion from platelets in the presence of tumor cells.36 Taken together these studies suggest the major platelet integrin receptor plays a significant role in tumorigenesis at several different mechanistic levels.
The major ligand of GP IIb-IIIa, fibrinogen, is also implicated in metastasis. As a central ligand supporting platelet/platelet interactions and as a key cleavage substrate for thrombin in coagulation, fibrinogen is essential in the well characterized paradigm of hemostasis andthrombosis.37 In the realm of tumor biology, fibrinogen supports the formation of platelet-fibrinogen-tumor cell emboli as tumor cells intravasate.38 Local deposition of fibrin and fibrin products have been found in solid tumors39 and reported to support angiogenesis.40 Experimental metastasis, spontaneous hematogenous metastasis and lymphatic metastasis are significantly diminished in fibrinogen knockout mice.3,41,42 Soluble fibrin monomer infusion facilitates platelet-tumor cell adhesion in vitro and experimental metastasis in vivo.43 Thus, fibrinogen and fibrin participate in a variety of pathways contributing to tumor cell survival and growth.
Thrombin and tumorigenesis
The role of thrombin in normal platelet function and in the pathways of blood coagulation highlight its importance as a central molecule linking the cellular (platelet) and biochemical (coagulation) paradigms of hemostasis.44 Thrombin treatment of platelets facilitates platelet adhesion on tumor cells by 2–4 fold in various cancer cells (HM54, HCT8,CT26, and B16).45 Thrombin activated tumor cells (CT26 and B16F10) show a 10–156 fold increase in experimental metastasis.46 Use of the thrombin agonist TRAP (thrombin receptor activation peptide) on CT26 or B16F10 cells also results in an increase in experimental metastasis.47 Thrombin has been found to break endothelial junctions and aid in VE cadherin and β-catenin mediated angiogenesis and tumor growth.48,49 Thrombin also acts as a mitogenic agent for various mesenchymal tissues and cells by activating growth-stimulatory signals.50 Hirudin, a thrombin antagonist has been found to diminish 4T1 mouse primary tumor growth and spontaneous tumor metastasis in the mouse.51
Thrombin also up-regulates the expression of various growth factors.It induces the secretion/expression of MMP–1, MMP–2, vascular endothelial growth factor (VEGF),angiopoietin-2 (ANG–2), CD31, and receptors KDR and CXCR2 from HUVEC and ANG-1 from platelets.10,52–56 Recently it has been shown that thrombin upregulates secretion of GRO-α from MCF7 and HUVEC and anti-GRO-α antibodies inhibit various angiogenic properties of MCF7 and HUVECs.57 Thrombin was also found to up-regulate expression of TWIST (an angiogenesis and tumor growth promoting transcription factor) in tumor cells and endothelial cells.58 Interestingly, thrombin regulates pro-angiogenic and anti-angiogenic factors differentially.59 In platelets, the PAR–4 agonist (ATPGFK) inhibits VEGF-A secretion while increasing endostatin secretion.The PAR–1 agonist (TFLLR) increases VEGF-A secretion and inhibits endostatin secretion.
Thrombin cleaves the platelet protease-activated receptor-1 (PAR–1), PAR–2, PAR–3 and PAR–4 at their N-terminal end which in turn activates G-protein mediated intracellular signaling.60 PAR1 expression has been directly correlated with the degree of invasiveness in primary breast tissue specimens.61 Overexpression of PAR–1 in B16 cells results in a 5-fold increase in experimental metastasis.62 It has also been reported that MMP–1 cleaves PAR–1 and enhances tumor growth and invasion of MDA-MB–231 cells in vivo.63 PAR–4 deficient mouse colonies (SCID and C57 background) display a significant reduction in B16F10 cell induced experimental pulmonary metastasis.3 In a spontaneous tumor metastasis model, PAR–2−/ − mouse colonies have reduced mammary adenocarcinoma growth and associated spontaneous metastasis.64
Platelets, natural killer cells, and tumorigenesis
As tumor cells intravasate to the circulation from a primary tumor they interact with various components of the circulation system including platelets and immune cells. In mouse models of experimental metastasis it has been found that most tumor cells entering the circulation do not survive with approximately 0.01% colonizing the lung.12 Natural killer (NK) cells are largely responsible for the elimination of cancer cells from the circulation.65,66 Experimental and genetic depletion of NK cells in mice causes a 2–5 fold increase in experimental metastasis.11,12 It has been proposed that platelets make a cloak around tumor cells and protect the tumor cell from NK cells.11 It addition, platelets and fibrinogen are linked to a significant reduction in the cytolytic activity of NK cells in vitro.11 NK cells express Mac–1 (integrin αMβ2 which has been shown to bind to platelet GP Ibα.67 Whether a GP Ibα– Mac–1 mediated platelet-NK cell interaction plays a role in regulating NK cell cytolytic activity for cancer cells, remains to be examined. Taken together, these observations highlight the platelet and coagulation interplay in the NK cell response to tumor cells.
Platelets, angiogenesis and tumorigenesis
In 1971, Judah Folkman proposed that tumor-growth is dependent on angiogenesis.68,69 Platelets store various angiogenesis regulating factors such as VEGF, platelet-derived growth factor (PDGF), fibroblast growth factor (FGF), epidermal growth factor (EGF), hepatocyte growth factor (HGF), insulin-like growth factor (IGF), angiopoietin, lysophophatidic acid, sphingosine 1–phosphate, CD40 ligand, MMP–1, MMP–2, MMP–9, gelatinase A, and heparanase. Most of these angiogenic agents have been shown to participate in angiogenesis for tumor growth directly or indirectly. Platelets also contain anti-angiogenic agents such as angiostatin, thrombospondin–1, platelet factor-4, endostatin, transforming growth factor β (TGF-β), and tissue inhibitor of matrix metalloproteinases (TIMPs).Dissecting the relevance of pro- and anti-angiogenic factors in the milieu of the platelet releasate represents a major challenge for the future.70,71
Recently, it has been found that expression of a negative regulator of angiogenesis, platelet-derived thrombospondin-1, is increased in tumor bearing mice after tumor resection. Primary tumor growth of Lewis lung carcinoma cells was significantly increased in TSP–1 deficient mice suggesting a role for angiogenesis in tumor growth.72 A chemically synthesized COOH-terminal peptide of platelet factor-4 (CXCL4L1) can inhibit angiogenesis and B16 induced melanoma growth in vivo.73 Together these results suggest there is a role for platelet derived anti-angiogenic factors and may represent new directions for future studies.
Conclusions
The role of platelets in hemostasis and thrombosis has been studied for several decades with remarkable molecular insights defining the hemostasis or thrombosis paradigm. Indeed, many of the well-studied platelet receptors and pathways can influence other diseases. Obvious connections to tumor growth, angiogenesis, and metastasis have been described here. Thus, the potential for insights from one discipline to rapidly contribute to new understandings in a different discipline is exciting. Future studies will hopefully contribute to both disciplines ultimately leading to better prevention, diagnosis, and treatment of disease.
Table I.
| Factor | Origin | Primary Tumor | Metastasis |
|---|---|---|---|
| Platelet count | Platelet | ↑ | |
| Platelet microparticles | Platelet | ↑ | |
| P-selectin | Platelet | ↑ | ↑ |
| MAPK p38α | Platelet | No effect | ↑ |
| GP Ibα | Platelet | No effect | ↑ |
| VWF | Platelet, EC | No effect | ↓ |
| GP VI | Platelet | No effect | ↑ |
| GP IIbIIIa | Platelet | ↑ | |
| Fibrinogen | Platelet | ↑ | |
| PAR–1 | Platelet, EC | ↑ | ↑ |
| PAR–2 | EC | ↑ | ↑ |
| PAR–4 | Platelet | ↑ | |
| Thrombin | EC, TC | ↑ | ↑ |
| Chondroitin sulfate glycosaminoglycans | Tumor cells | ↑ | |
| Sialyl Lewis X | Tumor cells | ↑ | |
| TSP-1 | Platelets | ↓ |
EC – Endothelial cells, TC – Tumor cells, ↑- Stimulatory effect, ↓- Inhibitory effect
Footnotes
Disclosure of Conflict of Interests
Authors are supported by grants; NHLBI HL50541 and the Department of Defense Breast Cancer Research Program
References
- 1.Varki A. Trousseau’s syndrome: multiple definitions and multiple mechanisms. Blood. 2007;110:1723–1729. doi: 10.1182/blood-2006-10-053736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gasic GJ, Gasic TB, Stewart CC. Antimetastatic effects associated with platelet reduction. Proc Natl Acad Sci U S A. 1968;61:46–52. doi: 10.1073/pnas.61.1.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Camerer E, Qazi AA, Duong DN, Cornelissen I, Advincula R, Coughlin SR. Platelets, protease-activated receptors, and fibrinogen in hematogenous metastasis. Blood. 2004;104:397–401. doi: 10.1182/blood-2004-02-0434. [DOI] [PubMed] [Google Scholar]
- 4.Karpatkin S, Pearlstein E. Role of platelets in tumor cell metastases. Ann Intern Med. 1981;95:636–641. doi: 10.7326/0003-4819-95-5-636. [DOI] [PubMed] [Google Scholar]
- 5.Gasic GJ, Gasic TB, Galanti N, Johnson T, Murphy S. Platelet-tumor-cell interactions in mice. The role of platelets in the spread of malignant disease. Int J Cancer. 1973;11:704–718. doi: 10.1002/ijc.2910110322. [DOI] [PubMed] [Google Scholar]
- 6.Pearlstein E, Salk PL, Yogeeswaran G, Karpatkin S. Correlation between spontaneous metastatic potential, platelet-aggregating activity of cell surface extracts, and cell surface sialylation in 10 metastatic-variant derivatives of a rat renal sarcoma cell line. Proc Natl Acad Sci U S A. 1980;77:4336–4339. doi: 10.1073/pnas.77.7.4336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Italiano JE, Jr, Mairuhu AT, Flaumenhaft R. Clinical relevance of microparticles from platelets and megakaryocytes. Curr Opin Hematol. 2010 doi: 10.1097/MOH.0b013e32833e77ee. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Janowska-Wieczorek A, Wysoczynski M, Kijowski J, Marquez-Curtis L, Machalinski B, Ratajczak J, Ratajczak MZ. Microvesicles derived from activated platelets induce metastasis and angiogenesis in lung cancer. Int J Cancer. 2005;113:752–760. doi: 10.1002/ijc.20657. [DOI] [PubMed] [Google Scholar]
- 9.Baj-Krzyworzeka M, Majka M, Pratico D, Ratajczak J, Vilaire G, Kijowski J, Reca R, Janowska-Wieczorek A, Ratajczak MZ. Platelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cells. Exp Hematol. 2002;30:450–459. doi: 10.1016/s0301-472x(02)00791-9. [DOI] [PubMed] [Google Scholar]
- 10.Dashevsky O, Varon D, Brill A. Platelet-derived microparticles promote invasiveness of prostate cancer cells via upregulation of MMP–2 production. Int J Cancer. 2009;124:1773–1777. doi: 10.1002/ijc.24016. [DOI] [PubMed] [Google Scholar]
- 11.Nieswandt B, Hafner M, Echtenacher B, Mannel DN. Lysis of tumor cells by natural killer cells in mice is impeded by platelets. Cancer Res. 1999;59:1295–1300. [PubMed] [Google Scholar]
- 12.Palumbo JS, Talmage KE, Massari JV, La Jeunesse CM, Flick MJ, Kombrinck KW, Jirouskova M, Degen JL. Platelets and fibrin(ogen) increase metastatic potential by impeding natural killer cell-mediated elimination of tumor cells. Blood. 2005;105:178–185. doi: 10.1182/blood-2004-06-2272. [DOI] [PubMed] [Google Scholar]
- 13.Wagner DD. New links between inflammation and thrombosis. Arterioscler Thromb Vasc Biol. 2005;25:1321–1324. doi: 10.1161/01.ATV.0000166521.90532.44. [DOI] [PubMed] [Google Scholar]
- 14.Monzavi-Karbassi B, Stanley JS, Hennings L, Jousheghany F, Artaud C, Shaaf S, Kieber-Emmons T. Chondroitin sulfate glycosaminoglycans as major P-selectin ligands on metastatic breast cancer cell lines. Int J Cancer. 2007;120:1179–1191. doi: 10.1002/ijc.22424. [DOI] [PubMed] [Google Scholar]
- 15.Garcia J, Callewaert N, Borsig L. P-selectin mediates metastatic progression through binding to sulfatides on tumor cells. Glycobiology. 2007;17:185–196. doi: 10.1093/glycob/cwl059. [DOI] [PubMed] [Google Scholar]
- 16.Kim YJ, Borsig L, Varki NM, Varki A. P-selectin deficiency attenuates tumor growth and metastasis. Proc Natl Acad Sci U S A. 1998;95:9325–9330. doi: 10.1073/pnas.95.16.9325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Borsig L, Wong R, Hynes RO, Varki NM, Varki A. Synergistic effects of L- and P-selectin in facilitating tumor metastasis can involve non-mucin ligands and implicate leukocytes as enhancers of metastasis. Proc Natl Acad Sci U S A. 2002;99:2193–2198. doi: 10.1073/pnas.261704098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang J, Nakayama J, Ohyama C, Suzuki M, Suzuki A, Fukuda M, Fukuda MN. Sialyl Lewis X-dependent lung colonization of B16 melanoma cells through a selectin-like endothelial receptor distinct from E- or P-selectin. Cancer Res. 2002;62:4194–4198. [PubMed] [Google Scholar]
- 19.Fuster MM, Brown JR, Wang L, Esko JD. A disaccharide precursor of sialyl Lewis X inhibits metastatic potential of tumor cells. Cancer Res. 2003;63:2775–2781. [PubMed] [Google Scholar]
- 20.Borsig L, Wong R, Feramisco J, Nadeau DR, Varki NM, Varki A. Heparin and cancer revisited: mechanistic connections involving platelets, P-selectin, carcinoma mucins, and tumor metastasis. Proc Natl Acad Sci U S A. 2001;98:3352–3357. doi: 10.1073/pnas.061615598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Matsuo Y, Amano S, Furuya M, Namiki K, Sakurai K, Nishiyama M, Sudo T, Tatsumi K, Kuriyama T, Kimura S, Kasuya Y. Involvement of p38alpha mitogen-activated protein kinase in lung metastasis of tumor cells. J Biol Chem. 2006;281:36767–36775. doi: 10.1074/jbc.M604371200. [DOI] [PubMed] [Google Scholar]
- 22.Kohler S, Ullrich S, Richter U, Schumacher U. E-/P-selectins and colon carcinoma metastasis: first in vivo evidence for their crucial role in a clinically relevant model of spontaneous metastasis formation in the lung. Br J Cancer. 2010;102:602–609. doi: 10.1038/sj.bjc.6605492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Andrews RK, Gardiner EE, Shen Y, Whisstock JC, Berndt MC. Glycoprotein Ib-IX-V. Int J Biochem Cell Biol. 2003;35:1170–1174. doi: 10.1016/s1357-2725(02)00280-7. [DOI] [PubMed] [Google Scholar]
- 24.Jain S, Zuka M, Liu J, Russell S, Dent JA, Guerrero JA, Forsyth J, Maruszak B, Gartner TK, Felding-Habermann B, Ware J. Platelet glycoprotein Iba supports experimental lung metastasis. Proc Natl Acad Sci U S A. 2007;104:9024–9028. doi: 10.1073/pnas.0700625104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Erpenbeck L, Nieswandt B, Schon M, Pozgajova M, Schon MP. Inhibition of platelet GPIb alpha and promotion of melanoma metastasis. J Invest Dermatol. 2010;130:576–586. doi: 10.1038/jid.2009.278. [DOI] [PubMed] [Google Scholar]
- 26.Jain S, Russell S, Ware J. Platelet glycoprotein VI facilitates experimental lung metastasis in syngenic mouse models. J Thromb Haemost. 2009;7:1713–1717. doi: 10.1111/j.1538-7836.2009.03559.x. [DOI] [PubMed] [Google Scholar]
- 27.Suter CM, Hogg PJ, Price JT, Chong BH, Ward RL. Identification and characterisation of a platelet GPIb/V/IX–like complex on human breast cancers: implications for the metastatic process. Jpn J Cancer Res. 2001;92:1082–1092. doi: 10.1111/j.1349-7006.2001.tb01063.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li Y, Lu J, Prochownik EV. c-Myc-mediated genomic instability proceeds via a megakaryocytic endomitosis pathway involving Gp1balpha. Proc Natl Acad Sci U S A. 2007;104:3490–3495. doi: 10.1073/pnas.0610163104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li Y, Lu J, Prochownik EV. Modularity of the oncoprotein-like properties of platelet glycoprotein Ibalpha. J Biol Chem. 2009;284:1410–1418. doi: 10.1074/jbc.M806222200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Terraube V, Pendu R, Baruch D, Gebbink MF, Meyer D, Lenting PJ, Denis CV. Increased metastatic potential of tumor cells in von Willebrand factor-deficient mice. J Thromb Haemost. 2006;4:519–526. doi: 10.1111/j.1538-7836.2005.01770.x. [DOI] [PubMed] [Google Scholar]
- 31.Karpatkin S, Pearlstein E, Ambrogis C, Coller BS. Role of adhesive proteins in platelet tumor interaction in vitro and metastasis formation in vivo. J Clin Invest. 1988;81:1012–1019. doi: 10.1172/JCI113411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Coller BS, Shattil SJ. The GPIIb/IIIa (integrin alphaIIbbeta3) odyssey: a technology-driven saga of a receptor with twists, turns, and even a bend. Blood. 2008;112:3011–3025. doi: 10.1182/blood-2008-06-077891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bakewell SJ, Nestor P, Prasad S, Tomasson MH, Dowland N, Mehrotra M, Scarborough R, Kanter J, Abe K, Phillips D, Weilbaecher KN. Platelet and osteoclast beta3 integrins are critical for bone metastasis. Proc Natl Acad Sci U S A. 2003;100:14205–14210. doi: 10.1073/pnas.2234372100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Varon D, Dardik R, Shenkman B, Kotev-Emeth S, Farzame N, Tamarin I, Savion N. A new method for quantitative analysis of whole blood platelet interaction with extracellular matrix under flow conditions. Throm Res. 1997;85:283–294. doi: 10.1016/s0049-3848(97)00014-5. [DOI] [PubMed] [Google Scholar]
- 35.Trikha M, Zhou Z, Timar J, Raso E, Kennel M, Emmell E, Nakada MT. Multiple roles for platelet GPIIb/IIIa and alphavbeta3 integrins in tumor growth, angiogenesis, and metastasis. Cancer Res. 2002;62:2824–2833. [PubMed] [Google Scholar]
- 36.Amirkhosravi A, Amaya M, Siddiqui F, Biggerstaff JP, Meyer TV, Francis JL. Blockade of GpIIb/IIIa inhibits the release of vascular endothelial growth factor (VEGF) from tumor cell-activated platelets and experimental metastasis. Platelets. 1999;10:285–292. doi: 10.1080/09537109975915. [DOI] [PubMed] [Google Scholar]
- 37.Lord ST. Fibrinogen and fibrin: scaffold proteins in hemostasis. Curr Opin Hematol. 2007;14:236–241. doi: 10.1097/MOH.0b013e3280dce58c. [DOI] [PubMed] [Google Scholar]
- 38.Im JH, Fu W, Wang H, Bhatia SK, Hammer DA, Kowalska MA, Muschel RJ. Coagulation facilitates tumor cell spreading in the pulmonary vasculature during early metastatic colony formation. Cancer Res. 2004;64:8613–8619. doi: 10.1158/0008-5472.CAN-04-2078. [DOI] [PubMed] [Google Scholar]
- 39.Brown LF, Asch B, Harvey VS, Buchinski B, Dvorak HF. Fibrinogen influx and accumulation of cross-linked fibrin in mouse carcinomas. Cancer Res. 1988;48:1920–1925. [PubMed] [Google Scholar]
- 40.Thompson WD, Smith EB, Stirk CM, Marshall FI, Stout AJ, Kocchar A. Angiogenic activity of fibrin degradation products is located in fibrin fragment E. J Pathol. 1992;168:47–53. doi: 10.1002/path.1711680109. [DOI] [PubMed] [Google Scholar]
- 41.Palumbo JS, Kombrinck KW, Drew AF, Grimes TS, Kiser JH, Degen JL, Bugge TH. Fibrinogen is an important determinant of the metastatic potential of circulating tumor cells. Blood. 2000;96:3302–3309. [PubMed] [Google Scholar]
- 42.Palumbo JS, Potter JM, Kaplan LS, Talmage K, Jackson DG, Degen JL. Spontaneous hematogenous and lymphatic metastasis, but not primary tumor growth or angiogenesis, is diminished in fibrinogen-deficient mice. Cancer Res. 2002;62:6966–6972. [PubMed] [Google Scholar]
- 43.Biggerstaff JP, Seth N, Amirkhosravi A, Amaya M, Fogarty S, Meyer TV, Siddiqui F, Francis JL. Soluble fibrin augments platelet/tumor cell adherence in vitro and in vivo, and enhances experimental metastasis. Clin Exp Metastasis. 1999;17:723–730. doi: 10.1023/a:1006763827882. [DOI] [PubMed] [Google Scholar]
- 44.Di Cera E. Thrombin as procoagulant and anticoagulant. J Thromb Haemost. 2007;5 (Suppl 1):196–202. doi: 10.1111/j.1538-7836.2007.02485.x. [DOI] [PubMed] [Google Scholar]
- 45.Nierodzik ML, Plotkin A, Kajumo F, Karpatkin S. Thrombin stimulates tumor–platelet adhesion in vitro and metastasis in vivo. J Clin Invest. 1991;87:229–236. doi: 10.1172/JCI114976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nierodzik ML, Kajumo F, Karpatkin S. Effect of thrombin treatment of tumor cells on adhesion of tumor cells to platelets in vitro and tumor metastasis in vivo. Cancer Res. 1992;52:3267–3272. [PubMed] [Google Scholar]
- 47.Nierodzik ML, Bain RM, Liu LX, Shivji M, Takeshita K, Karpatkin S. Presence of the seven transmembrane thrombin receptor on human tumour cells: effect of activation on tumour adhesion to platelets and tumor tyrosine phosphorylation. Br J Haematol. 1996;92:452–457. doi: 10.1046/j.1365-2141.1996.d01-1494.x. [DOI] [PubMed] [Google Scholar]
- 48.Konstantoulaki M, Kouklis P, Malik AB. Protein kinase C modifications of VE–cadherin, p120, and beta–catenin contribute to endothelial barrier dysregulation induced by thrombin. Am J Physiol Lung Cell Mol Physiol. 2003;285:L434–L442. doi: 10.1152/ajplung.00075.2003. [DOI] [PubMed] [Google Scholar]
- 49.Liao F, Li Y, O’Connor W, Zanetta L, Bassi R, Santiago A, Overholser J, Hooper A, Mignatti P, Dejana E, Hicklin DJ, Bohlen P. Monoclonal antibody to vascular endothelial–cadherin is a potent inhibitor of angiogenesis, tumor growth, and metastasis. Cancer Res. 2000;60:6805–6810. [PubMed] [Google Scholar]
- 50.Carney DH, Stiernberg J, Fenton JW. Initiation of proliferative events by human alpha–thrombin requires both receptor binding and enzymic activity. J Cell Biochem. 1984;26:181–195. doi: 10.1002/jcb.240260306. [DOI] [PubMed] [Google Scholar]
- 51.Hu L, Lee M, Campbell W, Perez-Soler R, Karpatkin S. Role of endogenous thrombin in tumor implantation, seeding, and spontaneous metastasis. Blood. 2004;104:2746–2751. doi: 10.1182/blood-2004-03-1047. [DOI] [PubMed] [Google Scholar]
- 52.Mohle R, Green D, Moore MA, Nachman RL, Rafii S. Constitutive production and thrombin-induced release of vascular endothelial growth factor by human megakaryocytes and platelets. Proc Natl Acad Sci U S A. 1997;94:663–668. doi: 10.1073/pnas.94.2.663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tsopanoglou NE, Maragoudakis ME. On the mechanism of thrombin-induced angiogenesis. Potentiation of vascular endothelial growth factor activity on endothelial cells by up-regulation of its receptors. J Biol Chem. 1999;274:23969–23976. doi: 10.1074/jbc.274.34.23969. [DOI] [PubMed] [Google Scholar]
- 54.Ollivier V, Chabbat J, Herbert JM, Hakim J, de Prost D. Vascular endothelial growth factor production by fibroblasts in response to factor VIIa binding to tissue factor involves thrombin and factor Xa. Arterioscler Thromb Vasc Biol. 2000;20:1374–1381. doi: 10.1161/01.atv.20.5.1374. [DOI] [PubMed] [Google Scholar]
- 55.Li JJ, Huang YQ, Basch R, Karpatkin S. Thrombin induces the release of angiopoietin-1 from platelets. Thromb Haemost. 2001;85:204–206. [PubMed] [Google Scholar]
- 56.Huang YQ, Li JJ, Hu L, Lee M, Karpatkin S. Thrombin induces increased expression and secretion of angiopoietin-2 from human umbilical vein endothelial cells. Blood. 2002;99:1646–1650. doi: 10.1182/blood.v99.5.1646. [DOI] [PubMed] [Google Scholar]
- 57.Caunt M, Hu L, Tang T, Brooks PC, Ibrahim S, Karpatkin S. Growth-regulated oncogene is pivotal in thrombin-induced angiogenesis. Cancer Res. 2006;66:4125–4132. doi: 10.1158/0008-5472.CAN-05-2570. [DOI] [PubMed] [Google Scholar]
- 58.Hu L, Roth JM, Brooks P, Ibrahim S, Karpatkin S. Twist is required for thrombin-induced tumor angiogenesis and growth. Cancer Res. 2008;68:4296–4302. doi: 10.1158/0008-5472.CAN-08-0067. [DOI] [PubMed] [Google Scholar]
- 59.Ma L, Perini R, McKnight W, Dicay M, Klein A, Hollenberg MD, Wallace JL. Proteinase-activated receptors 1 and 4 counter-regulate endostatin and VEGF release from human platelets. Proc Natl Acad Sci U S A. 2005;102:216–220. doi: 10.1073/pnas.0406682102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Coughlin SR. Protease-activated receptors in hemostasis, thrombosis and vascular biology. J Thromb Haemost. 2005;3:1800–1814. doi: 10.1111/j.1538-7836.2005.01377.x. [DOI] [PubMed] [Google Scholar]
- 61.Even-Ram S, Uziely B, Cohen P, Grisaru-Granovsky S, Maoz M, Ginzburg Y, Reich R, Vlodavsky I, Bar-Shavit R. Thrombin receptor overexpression in malignant and physiological invasion processes. Nat Med. 1998;4:909–914. doi: 10.1038/nm0898-909. [DOI] [PubMed] [Google Scholar]
- 62.Nierodzik ML, Chen K, Takeshita K, Li JJ, Huang YQ, Feng XS, D’Andrea MR, Andrade-Gordon P, Karpatkin S. Protease-activated receptor 1 (PAR–1) is required and rate-limiting for thrombin-enhanced experimental pulmonary metastasis. Blood. 1998;92:3694–3700. [PubMed] [Google Scholar]
- 63.Boire A, Covic L, Agarwal A, Jacques S, Sherifi S, Kuliopulos A. PAR1 is a matrix metalloprotease-1 receptor that promotes invasion and tumorigenesis of breast cancer cells. Cell. 2005;120:303–313. doi: 10.1016/j.cell.2004.12.018. [DOI] [PubMed] [Google Scholar]
- 64.Versteeg HH, Schaffner F, Kerver M, Ellies LG, Andrade-Gordon P, Mueller BM, Ruf W. Protease-activated receptor (PAR) 2, but not PAR1, signaling promotes the development of mammary adenocarcinoma in polyoma middle T mice. Cancer Res. 2008;68:7219–7227. doi: 10.1158/0008-5472.CAN-08-0419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Gorelik E, Wiltrout RH, Okumura K, Habu S, Herberman RB. Role of NK cells in the control of metastatic spread and growth of tumor cells in mice. Int J Cancer. 1982;30:107–112. doi: 10.1002/ijc.2910300118. [DOI] [PubMed] [Google Scholar]
- 66.Hanna N. The role of natural killer cells in the control of tumor growth and metastasis. Biochim Biophys Acta. 1985;780:213–226. doi: 10.1016/0304-419x(85)90004-6. [DOI] [PubMed] [Google Scholar]
- 67.Simon DI, Chen Z, Xu H, Li CQ, Dong J, McIntire LV, Ballantyne CM, Zhang L, Furman MI, Berndt MC, Lopez JA. Platelet glycoprotein ibalpha is a counterreceptor for the leukocyte integrin Mac–1 (CD11b/CD18) J Exp Med. 2000;192:193–204. doi: 10.1084/jem.192.2.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285:1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 69.Pinedo HM, Verheul HM, D’Amato RJ, Folkman J. Involvement of platelets in tumour angiogenesis? Lancet. 1998;352:1775–1777. doi: 10.1016/s0140-6736(98)05095-8. [DOI] [PubMed] [Google Scholar]
- 70.Italiano JE, Jr, Richardson JL, Patel-Hett S, Battinelli E, Zaslavsky A, Short S, Ryeom S, Folkman J, Klement GL. Angiogenesis is regulated by a novel mechanism: pro- and antiangiogenic proteins are organized into separate platelet alpha granules and differentially released. Blood. 2008;111:1227–1233. doi: 10.1182/blood-2007-09-113837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Peterson JE, Zurakowski D, Italiano JE, Jr, Michel LV, Fox L, Klement GL, Folkman J. Normal ranges of angiogenesis regulatory proteins in human platelets. Am J Hematol. 2010;85:487–493. doi: 10.1002/ajh.21732. [DOI] [PubMed] [Google Scholar]
- 72.Zaslavsky A, Baek KH, Lynch RC, Short S, Grillo J, Folkman J, Italiano JE, Jr, Ryeom S. Platelet-derived thrombospondin-1 is a critical negative regulator and potential biomarker of angiogenesis. Blood. 2010;115:4605–4613. doi: 10.1182/blood-2009-09-242065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Vandercappellen J, Liekens S, Bronckaers A, Noppen S, Ronsse I, Dillen C, Belleri M, Mitola S, Proost P, Presta M, Struyf S, Van Damme J. The COOH-terminal peptide of platelet factor–4 variant (CXCL4L1/PF–4var47–70) strongly inhibits angiogenesis and suppresses B16 melanoma growth in vivo. Mol Cancer Res. 2010;8:322–334. doi: 10.1158/1541-7786.MCR-09-0176. [DOI] [PubMed] [Google Scholar]