

Abstract
Cofilin is an important regulator of actin polymerization, cell migration, and chemotaxis. Recent experimental data on mammary carcinoma cells reveal that stimulation by epidermal growth factor (EGF) generates a pool of active cofilin that results in a peak of actin filament barbed ends on the timescale of 1 min. Here, we present results of a mathematical model for the dynamics of cofilin and its transition between several pools in response to EGF stimulation. We describe the interactions of phospholipase C, membrane lipids (PIP2), and cofilin bound to PIP2 and to F-actin, as well as diffusible cofilin in active G-actin-monomer-bound or phosphorylated states. We consider a simplified representation in which the thin cell edge (lamellipod) and the cell interior are represented by two compartments that are linked by diffusion. We demonstrate that a high basal level of active cofilin stored by binding to PIP2, as well as the highly enriched local milieu of F-actin at the cell edge, is essential to capture the EGF-induced barbed-end amplification observed experimentally.
Introduction
Eukaryotic cell motility relies on polymerization of F-actin to generate protrusive forces at the leading edge of the cell. This polymerization requires available fast-growing barbed ends for actin monomer addition. Indeed, in response to stimulation by epidermal growth factor (EGF), a large peak of barbed ends is observed to form in mammary carcinoma cells ∼1 min poststimulus. This peak has been found to be cofilin-dependent (1). Local activation of cofilin has been shown to lead to protrusion initiation and to determine the cell direction (1,2). Further, suppression of cofilin (via knockdown) in the same tumor cell type leads to cells with decreased protrusion velocity and chemotaxis (3,4).
Nucleation of new barbed ends by Arp2/3 is well known, but a similar role for cofilin has recently become apparent (5). Cofilin plays multiple roles, depending on conditions. These include disassembly and/or debranching of the actin dendritic network, as well as de novo F-actin nucleation (6–9). Here, we are concerned only with the role of cofilin in generating the first peak of barbed ends created downstream of EGF stimulation (10).
Cofilin has several states inside the cell. It can bind to F- or G-actin (preferring ADP- rather than ATP-actin (7,11)); it has freely diffusing dephosphorylated (active) and phosphorylated (inactive) forms (12,13). In resting carcinoma cells, van Rheenen et al. (14) found that there is a pool of cofilin bound to the phosphoinositide phosphatidylinositol 4,5-bisphosphate (PIP2), and hence inactive (15), at the cell membrane.
Although both phosphorylated and dephosphorylated cofilin can bind to PIP2 (16), only dephosphorylated cofilin was found to be enriched in the plasma membrane (17). Upon EGF stimulation, phospholipase-C (PLC) is activated and hydrolyzes PIP2 (into inositol trisphosphate and diacylglycerol), so that PIP2 falls to 40–60% of its basal level (1,14). Active cofilin (no longer taken up as rapidly by PIP2) becomes available in the cytoplasmic region adjoining the membrane. Our hypothesis is that this effective flux, together with the high local density of cytoskeleton leads to the rapid barbed-end peak. After severing F-actin, cofilin carries an actin monomer and has to be recharged. Phosphorylation by LIM kinase (LIMK) releases cofilin's bound G-actin, and dephosphorylation by slingshot phosphatase (SSH) allows cofilin to rebind to PIP2, completing its cycle.
It has been suggested that a single cofilin protein can interact with one or more PIP2 molecules (18). Thus, the twofold PIP2 drop would be consistent with a similar drop of PIP2-cofilin or a twofold increase of active cofilin. And yet, the peak of barbed ends is 10- to 15-fold higher than its basal level. This leads to our main question of what creates the observed barbed-end amplification. In addition, we address the following questions:
-
1.
Can cofilin dynamics alone account for the large transient pulse of actin filament barbed ends observed within 1 min of EGF stimulation of carcinoma cells? How does this amplification occur?
-
2.
Based on experimental observations, what are the flows of cofilin between the various pools described above?
-
3.
How much cofilin is in the freely diffusing active form in the resting versus stimulated cell?
-
4.
What are the effects of overexpressing or inhibiting the various agents that control the flows of cofilin between compartments?
To quantify the dynamics and compare with experimental data, we assembled several mathematical models, as described below.
Mathematical Models
Barbed-end amplification
Our preliminary model addresses the large barbed-end amplification. From here on, we define amplification as the ratio of barbed-end peak amplitude to barbed-end steady-state level in the resting state (Bpeak/Bss). Large amplification implies a low basal level of severing and/or a large peak after stimulus. The rapid growth of barbed ends could result from 1), de novo actin nucleation; 2), Arp2/3 mediated branching; or 3), severing of the filaments. Of these possible causes, we rejected the first, since spontaneous nucleation of filaments is slow (19) and limited by profilin (20). As for the second, experiments with PLC inhibitors and cofilin siRNA rule out Arp2/3 in the first peak of barbed ends, and demonstrate its involvement in a later second peak (1). This left the third hypothesis, which is explored further below.
We considered an elementary model for active cofilin (C) and barbed ends (B), with basal rates of production (IB, IC); cofilin inactivation, kp; and barbed-end capping, kcap:
| (1) |
| (2) |
( converts units of C (μM) to units of B (number/μm2).) We assumed that before stimulation, so there is very little severing activity. Poststimulus, , and barbed ends are generated by severing F-actin.
Nonlinear kinetics of severing
We asked what type of severing kinetics, Fsev(C), could account for the observed high amplification for B(t) and explored three possibilities: 1), a linear function, 2), a saturating function, and 3), a Hill function far from saturation. Analysis of Eqs. 1 and 2, shown in the Supporting Material, reveals that the cofilin dependence of the first barbed-end peak requires severing kinetics to be nonlinear and operating far from saturation, i.e., the third possibility. We ascribe this nonlinearity to the cooperative nature of cofilin binding to F-actin (21,22). This preliminary exploration informed our choice of the function Fsev(C) in Eq. 2.
| (3) |
where Crest is the concentration of cofilin in the resting state, n the degree of cooperativity (21), and ksev a constant. Note that the form of Eq. 3 is equivalent to a Hill function commonly used in cooperative kinetics, but in a regime far from saturation.
We asked whether the possible rounds of severing, F-actin polymerization, further severing, etc., could also account for a similar autocatalytic effect. We rejected this possibility for several reasons. First, total F-actin only doubles on the timescale of interest (23), far short of the observed factor of 10. Second, unlike Arp2/3, cofilin preferentially binds to ADP or ADP-Pi-F-actin (5). This would lead to some delay in a further round of severing while filaments lose their phosphate groups. Third, as older filaments recede from the cell edge, they are bound by tropomyosin and protected from further severing by cofilin (23). This limits the ability of self-amplification due to autocatalytic growth, leading us to reject such alternatives.
Cofilin regulation model
We now turn to more detailed models for the cofilin cycle in mammary carcinoma cells (1,14,17). We consider only the events leading up to the first peak of barbed ends post-EGF stimulation, constraining the model to match both basal levels of all intermediates (before stimulation) and the transient after the stimulus. Well-measured intermediates such as PLC and PIP2 are used as inputs, and the size, timing, and shape of the peak of barbed ends are used to appraise the model's predictive ability. The model also predicts cofilin flows between the various pools, both those observed experimentally, and those that are below experimental resolution.
A first attempt to model the cofilin cycle within a single spatial compartment failed to account for certain observations (see the Supporting Material). Here, we present the two-compartment model shown schematically in Fig. 1 (see the Supporting Material for the original and the Appendix for scaled-model equations). The compartments correspond to a small region (∼200 nm in width (23)) of cell edge adjacent to the membrane, and the cell interior with volumes VE and VI (see Fig. 1 enlarged in the Supporting Material). A small cylindrical surface of height l forms the interface between these volumes, through which diffusion can take place, as explained in the Supporting Material. Activation of PLC (denoted PLC(t)) downstream of the EGF signal leads to hydrolysis of PIP2 (P2(t)). We track five cofilin forms: cofilin bound to F-actin (Cf(t)) and to PIP2 on the membrane (C2(t)), both resident at the cell edge; and diffusible forms of cofilin, i.e., active (Ca(t)), actin-monomer-bound (Cm(t)), and inactive/phosphorylated (Cp(t)) cofilin, which can exchange between compartments. CiE and CiI denote diffusible cofilin concentrations in the edge/interior compartments. Basic assumptions include
- Conservation. On the timescale of interest (a few minutes), synthesis/degradation of cofilin is negligible, and the total amount of cofilin is roughly conserved. Thus,
where Ctot, the whole-cell total cofilin concentration, is assumed constant.(4) Severing due to F-actin bound cofilin. We assume that cofilin must be bound to F-actin (CF) to sever it. We adopt Fsev = Fsev(Cf) from Eq. 3.
PIP2 hydrolysis. The basal PIP2 hydrolysis rate was based on PIP2 FRET (14): post-EGF stimulus, PIP2 drops by 40–60%, then recovers to 80% of baseline within 360 s (14). From this, we estimated the recovery rate at dp2 = log 2/360 ∼ 0.002/s.
Binding and unbinding to F-actin. We based the unbinding rate, koff = 0.005/s, on in vitro data (7). We assume that F-actin binding sites for cofilin are not limiting, so that konF is approximately the constant representing the on-rate of cofilin binding to actin.
Phosphorylation and dephosphorylation. We first consider the case where rates of both phosphorylation (kmp) and dephosphorylation (kpm) are constant and similar for the two forms of cofilin, Ca, and Cm.
Actin monomer binding. Although the transition between Cp and Cm is a two-step process, we assume that it is only limited by the phosphorylation and dephosphorylation rates. As a simplification, G-actin binding/unbinding is assumed to be instantaneous and not limited by actin monomer availability. (This is reasonable in view of the ample level of actin monomers in cells, but see (24).)
Rebinding to PIP2. We assumed that poststimulus recovery of C2 follows rebinding of phosphocofilin to PIP2, making the conversion CpE → C2 (rate kp2).
Flux between compartments. Because compartments are of vastly different sizes, our balance equations contain compartment volume factors to preserve mass conservation (Appendix and Supporting Material). We assume that the cofilin flux between compartments is diffusive, and thus proportional to concentration gradients.
Figure 1.
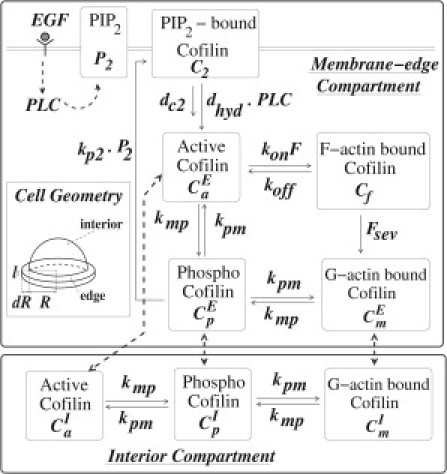
Schematics of the cell geometry and the cofilin regulation pathway used in the model. Cell geometry (also see Supporting Material for enlarged view): interior compartment (volume VI) is approximated as a hemisphere of radius R ≈ 5–10 μm. Edge compartment (volume VE) is shaped like a thin, flat washer (width dR, thickness l, both ∼200 nm) (14,24). Note that the figure is not to scale, as dR << R. Schematics of the cofilin regulation pathway of the ordinary-differential-equation model. The PIP2-bound cofilin, C2, and the F-actin-bound form, Cf, are restricted to the membrane edge compartment, whereas the other forms can diffuse between the two compartments (gray dashed arrows). Other arrows in the figure represent reaction terms in the equations (see Appendix and Supporting Material).
To compare model predictions with experimental observations, we also quantify barbed ends, B(t), using Eq. 2 as before, with Fsev = Fsev(CF), and IB = 0.
Scaling and parameter determination
Because most experimental results quantify levels of variables relative to their basal level, we scale the model accordingly. PLC, P2, and B are divided by their unstimulated steady-state values, and the cofilin forms by the total mean cofilin concentration in the cell, Ctot. Original variables are denoted in capital letters, and the corresponding scaled variables in lower-case letters (see Appendix), so that cjE, cjI are, respectively, the edge and interior scaled concentrations of cofilin form j. We also define the compartment volume fractions vE = VE/(VE + VI) and vI = VI/(VE + VI), so that the conservation statement (Eq. 4) becomes
| (5) |
The total whole-cell fraction of cofilin in form j is then given by vE cjE + vI cjI. We further denote the steady-state fractions as
| (6) |
Equations 5 and 6 then imply that R2 + Ra + Rm + RP + Rf = 1. Song et al. (17) found the fraction of phosphorylated cofilin before EGF stimulation (Rp) to be 20%. Van Rheenen et al. (14) observed that most of the cofilin residing at the edge is bound to either F-actin or to PIP2. They further showed that the ratio between the PIP2- and the F-actin-bound cofilin is 85%:15%, so we similarly take R2/Rf = 0.85/0.15. The fraction of PIP2-bound cofilin, R2, has not been determined experimentally. A lower estimate of 10% has been measured (R. Eddy and J. Condeelis, unpublished). The fraction of diffusible active cofilin, Ra, is difficult to measure experimentally. We used data fitting to quantify both Ra and R2. Finally, Rm can be determined from the conservation condition.
Parameter values (Table 1) were obtained from the literature by imposing steady-state constraints in Eq. 6, and/or by fitting the barbed-end profile from Mouneimne et al. (3) (see Fig. 3) and the time course of phosphocofilin from Song et al. (17) (Fig. 2). All experimental data used for parameter fitting are shown in Fig. 2 and details are discussed in the Supporting Material. Parameter sensitivity analysis was done for parameters determined from data fitting. Briefly, we first determined the 95% confidence level from bootstrapping: thus, if we were to choose a set of parameter values within the intervals, very similar results would be obtained. The results are listed in Table 1. Second, we also performed an error analysis on a broad range of parameter values (Fig. S5 in the Supporting Material) and found a relatively narrow region of parameter values that yields a good agreement between model and data (see Supporting Material for details). The most sensitive parameter is R2, the steady-state level of PIP2-bound cofilin.
Table 1.
List of parameter values
| Parameters | Definition | Values | 95% Interval | Source |
|---|---|---|---|---|
| Cell geometry and diffusion | ||||
| VE | Volume of the membrane-edge compartment | 50 μm3 | Cell geometry (see Supporting Material) | |
| VI | Volume of the interior compartment | 950 μm3 | Cell geometry (see Supporting Material) | |
| D | Diffusion coefficient of cofilin | 10 μm2/s | (36) | |
| l | Thickness of edge compartment | 0.2 μm | (24), (see Supporting Material) | |
| EGF stimulation | ||||
| I0 | Stimulus amplitude | 1.14 | 1.06–1.44 | PLC data fitting |
| ton | Time at which EGF stimulus starts | 25 s | ||
| toff | Time at which EGF stimulus ends | 85 s | ||
| PLC and PIP2 dynamics | ||||
| dplc | Basal PLC degradation rate | 0.026/s | 0.018–0.030 | PLC data fitting |
| dhyd | PLC-induced PIP2 hydrolysis rate | 0.032/s | 0.019–0.034 | Cofilin data fitting |
| dp2 | Basal PIP2 hydrolysis rate | 0.002/s | (14) | |
| Steady-state fractions of cofilin | ||||
| R2 | Fraction bound to PIP2 | 0.62 | 0.49–0.67 | Cofilin data fitting |
| Ra | Fraction of free active form | 0.04 | 0.01–0.11 | Cofilin data fitting |
| Rp | Fraction phosphorylated/inactive | 0.20 | (17) | |
| Rf | Fraction bound to F-actin | 0.11 | R2:Rf ≈ 0.85:0.15 (14) | |
| Rm | Fraction bound to G-actin | 0.03 | Conservation | |
| Cofilin transition rates | ||||
| dc2 | Basal c2 release rate (by PIP2 hydrolysis) | 0.002/s | dc2 = dp2 | |
| koff | Unbinding rate from F-actin | 0.005/s | (7) | |
| konF | Binding rate to F-actin | 0.198/s | Steady-state constraint | |
| kmp | Phosphorylation rate (LIMK) | 0.186/s | 0.19–0.45 | Cofilin data fitting |
| kpm | Dephosphorylation rate (SSH) | 0.03/s | Steady-state constraint | |
| kp2 | Binding rate to PIP2 | 0.11/s | Steady-state constraint | |
| ksev | Severing rate per cofilin molecule | 0.0012/s | Steady-state constraint | |
| n | Degree of cooperativity in severing | 4 | (21) | |
| ϕF | Steady-state value of cf (cf,ss) | 2.2 | Rf/vE | |
| Barbed end | ||||
| kcap | Barbed-end capping rate | 1/s | (24,36,37) | |
| A | Scaling factor for barbed-end generation | 4500 | Set so that b = 1 at rest | |
Values are estimated from literature sources, from steady-state constraints, or from data fitting. The 95% confidence intervals are obtained by bootstrapping 300 data sets (for details, see discussion in the Supporting Material and Fig. S4).
Figure 3.
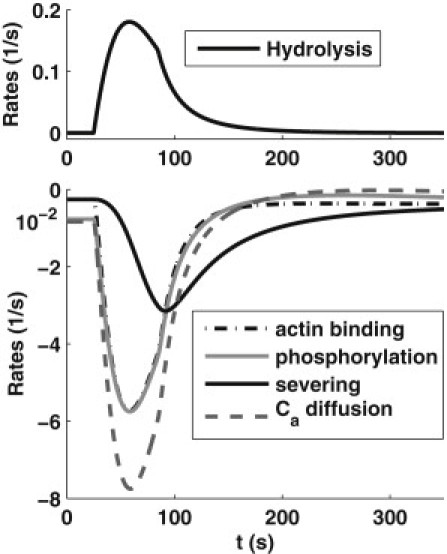
A comparison of rates of processes after stimulation. Shown are the hydrolysis rate, dhyd(plc – 1)c2; the net actin binding rate, −konF × caE + koff cf; the severing rate, fsev(cf); the net phosphorylation rate, −kmpcaE + kpmcpE; and the diffusion rate of active cofilin, ωD/vE(caI – caE).
Figure 2.
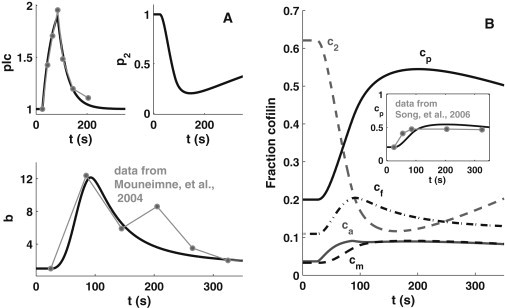
Dynamics of the system in response to EGF stimulation. Model results are obtained by simulation using the parameter values listed in Table 1. Experimental data (line segments connecting dots) are shown for comparison. (A) Time courses of PLC (plc), PIP2 (p2), and barbed-end (b) levels and data from Mouneimne et al. (1). (B) Fraction of total cell cofilin, vEcjE + vIcjI, for PIP2-bound (c2), F-actin-bound (cf), G-actin-bound (cm), phosphorylated (cp), and active (ca) cofilin. (Inset) Comparison of the simulation result of phosphocofilin, cp, and experimental data from Song et al. (17).
Results
Basic behavior
We simulated the model using default parameter values (Table 1), with variables initialized at the resting/steady-state values (Table S3). EGF stimulation (a pulse in Istim) is turned on at 25 s, and off 60 s later. Parameters for the PLC equation are fit to data in Mouneimne et al. (1), and other parameters are found as described above (and in the Supporting Material). The resulting parameter setting represents a control cell. Dynamics of plc, p2, the fraction of total cofilin in each form (vEcjE + vIcjI), and barbed ends are shown in Fig. 2; concentrations within each compartment are shown in Fig. S9. PLC dynamics (Fig. 2 A, upper left) closely match data from Mouneimne et al. (1). The scaled PIP2 (p2), and PIP2-bound cofilin (c2) dynamics agree qualitatively with observed behavior (14). Parameter fitting leads to a barbed-end time course, b(t), that matches the experimentally observed first peak (1) reasonably well. Barbed ends are amplified by an order of magnitude above the basal (resting) value. Only the first barbed-end peak is captured, since this model does not consider Arp2/3. Poststimulus, there is an ∼30 s delay before the sharp increase occurs, so that the peak is attained at ∼90 s. The delay is consistent with a second experimental data set shown in Mouneimne et al. (1), where little barbed-end production was observed within 30 s of EGF stimulation.
The delay in the barbed-end peak stems from the time for binding of cofilin to F-actin (to form cf) and severing of filaments. Although free active cofilin rises immediately poststimulus, F-actin-bound coflilin, cf, increases only ∼10 s later. In turn, G-actin-bound cofilin, cm, generated after severing, starts to grow at ∼20 s. Meanwhile, the level of phosphorylated cofilin, cp, increases more dramatically than cf. Note that the temporal profile of cp, for which we have experimental data (17), is part of the data-fitting process (Fig. 2 B, inset) and increases from 20% at rest to >50% poststimulus. Observe, however, that the rise of cp does not yet match the fast experimental rise, as we so far assumed that the phosphorylation rate is constant. We correct this further on.
In Fig. 3, we compare magnitudes of several poststimulus rates, including the F-actin severing rate (fsev(cf)) (first term of Eq. 12), the hydrolysis rate (dhyd (plc – 1)c2), the net actin-binding rate (konFcaE + koff cf), and the net diffusion of active cofilin out of the edge compartment ((ωD/vE)(caI – caE)). Diffusion dominates after stimulation: the large flux of c2 released into the edge compartment creates a large concentration gradient in active cofilin that rapidly diffuses to the cell interior. Phosphorylation and F-actin binding are slower and less pronounced. Thus, the delay in barbed-end generation stems from competition of cofilin binding to F-actin with cofilin diffusion and phosphorylation. As cf slowly builds up, severing follows. Note that the rate of severing shown in Fig. 3 is lower than other processes. The peak of severing occurs at ∼90 s, consistent with the peak of barbed ends.
Effects of LIMK up-/downregulation
We asked how the up-/downregulation of LIMK (the kinase that phosphorylates cofilin) affects the dynamics. This is of particular interest, since overexpression of LIMK has been reported to both enhance and inhibit cell motility (12,25,26). Moreover, upregulation of LIMK expression is also observed in invasive cancer cells, along with increased cofilin activity (27). To study the effect of LIMK expression in our model, we examined how changing kmp (rate of cofilin phosphorylation) by a factor of 2 affects the dynamics.
Increasing kmp leads to a higher steady-state cp value and thus less severing and fewer barbed ends. However, it also lowers the steady-state barbed-end level, bss, in such a way that the amplification bpeak/bss actually increases (Fig. 4 A). Doubling kmp leads to a >20-fold amplification. The timing of the barbed-end peak is unchanged. Since phosphorylation liberates cofilin from G/F-actin bound forms, cm and cf, increasing kmp also leads to a larger basal c2, i.e., a greater proportion of cofilin stored in a readily recruitable form is available for release and activation upon stimulation.
Figure 4.
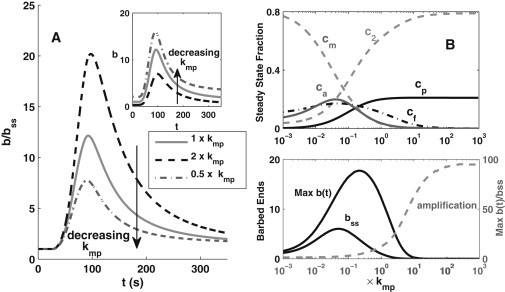
Effects of changing the cofilin phosphorylation rate, kmp, reflecting basal LIMK activity, on barbed-end generation. (A) Increasing (black dashed line) or decreasing (gray dashed line) the value of kmp by a factor of 2 from its original value (solid gray line) affects the dynamics. (Inset) Barbed-end levels relative to baseline (b(t)/bss) and absolute levels (b(t)). (B) Dependence of steady-state and stimulated levels of variables on the phosphorylation rate, kmp, showing the steady-state fractions of various forms of cofilin versus kmp (upper) and the steady-state level (bss), maximum level (bpeak = Max b(t)) (left axis), and amplification (bpeak/bss) (right axis) of barbed ends versus the phosphorylation rate, kmp (lower). The horizontal axis is the fold up-/downregulation of the original phosphorylation rate, kmp, in Table 1.
Analyzing the steady-state dependence, we find that at very low kmp, steady-state cofilin is dominated by cm with very little c2 (Fig. 4 B). As kmp increases, c2 and cp increase and cm decreases monotonically. Further, cf and ca first increase, then decrease with kmp. This nonmonotonicity leads to a nonmonotonic dependence of steady-state barbed ends (bss) and transient peak (bpeak) on kmp (Fig. 4 B, lower). Initially, as kmp is increased, more PIP2-bound cofilin is available to be released so that the barbed-end levels at rest and after stimulation also increase. At the higher range (kmp > konF), phosphorylation robs activated cofilin before it can bind to and sever F-actin. These results imply that regulation of cofilin by the basal level of LIMK plays a role not only in limiting the action of severing, but also in both up- and downregulation of barbed-end amplification.
Dynamics of LIMK
Absent in the model so far are the dynamics of LIMK and SSH poststimulation by EGF. Partly due to that simplification, cp dynamics (Fig. 2, inset) were so far not closely matched to the experimental data, despite the fitting procedure. Song et al. (17) showed that active/phosphorylated LIMK doubles and stays elevated for 360 s poststimulation. The (unmeasured) level of SSH can be inferred to rise since the phosphocofilin level also remained elevated at 360 s after stimulation (17). We asked how such LIMK/SSH dynamics could affect our conclusions.
To avoid significantly increasing the model complexity, we simply fit a time-varying function f(t) to the LIMK data in Song et al. (17) (Fig. 5, upper). We assumed that LIMK = kmp × f(t) and SSH = kpm × f(t), i.e., that LIMK and SSH activities were proportional to f(t).
Figure 5.
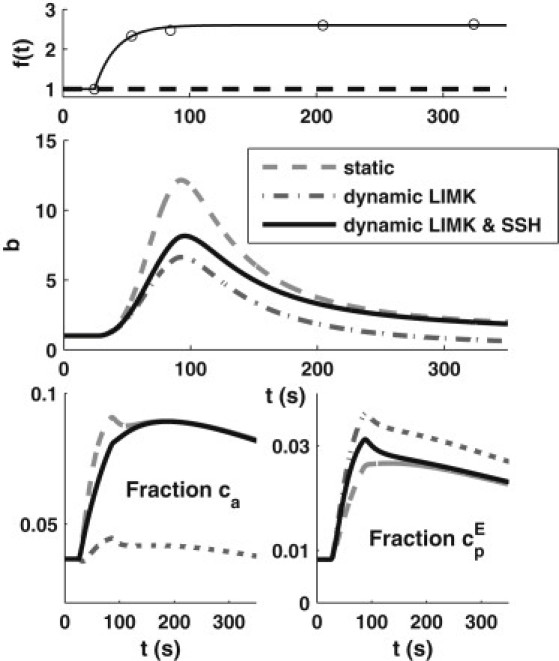
Effects of dynamic LIMK and SSH response to stimulation in model and experimental data (indicated below instead). (Upper) Phosphorylation and dephosphorylation were both assumed to be kmp × f(t) and kpm × f(t), with f(t) = 1 + 1.6(1 – exp(0.06 t)); this function was chosen to fit data (open dots) from Song et al. (17). (Middle) Dynamics of barbed-end profile for the original model/static case (constant kmp and kpm) (dashed gray line), the dynamic LIMK case (kmp × f(t) only) (dash-dotted line), and dynamic LIMK and SSH (kmp × f(t) and kpm × f(t)) (solid line). (Lower) Dynamics of free active (left) and phosphorylated (right) cofilin on the edge compartment.
We compare the output for dynamic LIMK, dynamic LIMK-SSH, and the original model in Fig. 5. In the first case (kmp × f(t), kpm = constant), bpeak is only half of its value in the basic model. This model variant has rampant cofilin phosphorylation due to the poststimulus rise of LIMK activity. Thus, active cofilin, ca, can hardly build up (Fig. 5, lower right), so there is less cofilin binding to F-actin, less severing, and thus a lower bpeak. With both LIMK and SSH dynamics, amplification is ∼8. The dynamics of cofilin forms (Fig. S10), particularly those in the interior compartment, are fairly similar across the three model variants, with the level of ca showing the greatest impact of dynamic LIMK.
We also explored the possibility that the edge compartment is protected from phosphorylation, as suggested by data from Song et al. (17), by setting kmp = kpm = 0 in the edge compartment in the same dynamic LIMK/SSH model variant (Fig. 6). The steady-state value of c2 is slightly lower whereas cf and bss are elevated. Barbed-end amplification is ∼10-fold (Fig. 6, upper). As expected, the build-up of cpE is retarded when LIMK activity is minimal in the edge compartment. Including SSH dynamics leads to cell-interior cofilin dynamics similar to those of the original model (Fig. 6, lower).
Figure 6.
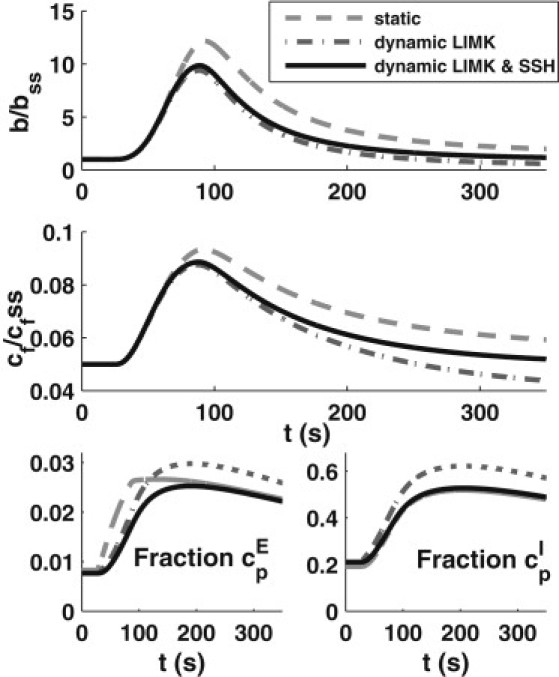
Effects of membrane edge protection where no phosphorylation/dephosphorylation occurs within the edge compartment (kmp = kpm = 0 at edge). In the interior compartment, dynamic LIMK and SSH responses to stimulation are considered as in Fig. 5. (Upper) Barbed ends (b(t)/bss). (Middle) Relative level of cf available for severing (cf scaled by its steady-state level, cf,ss). (Lower) Dynamics of phosphocofilin fractions in the edge and interior compartments.
A comparison with previous results (Fig. 5) shows that with membrane-edge protection, barbed ends peak more quickly (at a rate similar to that of the original model) but then also drop more quickly after peaking. We attribute the decrease in barbed-end amplification here not only to the drop in c2 level ready for release but also to the lack of reactivation occurring within the edge compartment after the first round of severing. This is shown in Fig. 6 (middle), where we plotted the level of cf scaled by its basal/steady-state level. Thus, early barbed-end production is sensitively tuned to events within the small edge compartment of the cell and is relatively insensitive to the cofilin dynamics in the bulk interior of the cell.
Discussion
In addition to recycling old actin filaments, cofilin plays an important role in initiating cell movement by severing filaments and generating new barbed ends. Our two-compartment ordinary-differential-equation model of cofilin dynamics downstream of EGF stimulation in cancer cells reproduces the early transient actin filament barbed-end production observed experimentally, and dissects the cofilin-induced F-actin severing into underlying steps. The model allows us to estimate rates of transition between various forms of cofilin by matching predictions and experimental data for both resting levels and stimulated levels of measurable quantities. Further, the model predicts the levels of variables (such as the fraction of active cofilin) that are currently beyond experimental resolution. The model demonstrates that the response of the system is most strongly affected by 1), the fraction of PIP2-bound cofilin (c2) in the resting cell available for release; and 2), the rate of binding and severing of F-actin relative to competing processes such as cofilin phosphorylation or diffusion. Another important take-home lesson is that upregulating the cofilin regulator LIMK can actually lead to a more amplified response to EGF stimulation, i.e., a greater amplification in barbed-end production poststimulation, even though the primary role of LIMK is to deactivate cofilin.
An essential component of our model is a strong nonlinearity in the severing rate as a function of active cofilin. In fact, we believe that the cofilin-actin cooperative binding kinetics (21,22) is responsible for generating the transient 10- to 15-fold barbed-end amplification (1). This assumption is consistent with previous in vitro and modeling studies on cofilin-actin binding (21,28,29), which show that cofilin binds cooperatively to F-actin with Hill-like kinetics of degree n = 4–10. We explored a range of values of n and found that a fit to the experimental data within the 95% confidence interval could be achieved with values of 4 ≤ n ≤ 10. We chose the value n = 4 as it yielded one of the best fits. Cofilin binding changes the flexibility of actin filaments, allowing more cofilin to bind; the enhanced bending flexibility also promotes severing (22,30,31). It is conjectured, but not yet confirmed, that mechanical tension at the cell edge may also affect cofilin dynamics (E. De La Cruz, Yale University, personal communication, 2011).
In building our model, we used mostly data from mammary carcinoma cells in vivo. Direct in vitro measurement is largely lacking, hindering the estimation of dephosphorylation and phosphorylation rates of cofilin. Having a finer temporal measurement of simultaneous levels of variables would be helpful in data fitting and in resolving the exact temporal dynamics.
From our parameter sensitivity study, we also found a critical parameter, namely the basal amount of PIP2-bound cofilin (c2 ≈ 0.6 at rest) needed to faithfully reproduce the experimental data. This means that ∼60% of total cofilin should be in the PIP2-bound form in the resting cell for dynamics consistent with observations. This is possible given the localization of cofilin with PIP2 observed in carcinoma cells (14). Although a large barbed-end amplification is possible at a lower level of c2, the level of phosphocofilin cannot then increase to the level observed experimentally (17).
Limitations of this model include the very coarse spatial representation (two compartments only) and inclusion of few key intermediates. We did not model here the dynamics of F-actin that takes place once the newly formed barbed ends start to grow and extend. In experiments, F-actin has been found to increase by less than twofold within 60 s after EGF stimulation. A more significant increase occurs later, once Arp2/3 activity is promoted (23) (not currently modeled). The second peak of barbed ends stems from Arp2/3-mediated F-actin branching (1). Tests of the model with a dynamic F-actin variable showed minimal effect beyond that described here and was deemed insignificant relative to other processes on the timescale of 1 min.
Cofilin has other roles that we did not include here. In vitro studies have shown that, depending on conditions, ADF/cofilin (at low concentrations) severs but (at higher levels) depolymerizes F-actin (6–9). The latter helps to recycle G-actin and promote rapid barbed-end growth (10,32,33). Other effects of cofilin include release of the phosphate groups from F-actin, promoting dissociation of Arp2/3 complexes from ADP-F-actin and consequent debranching of filaments (6,34). At a very high concentration (>10 μM), cofilin can also nucleate filaments de novo (6). In the interest of keeping the first models tractable, such effects were not modeled here. These could form extensions of the basic model as a future step.
Our results strongly suggest that the level of barbed-end production is mainly dependent on events occurring within a thin compartment abutting the cell membrane rather than the large interior cytosolic compartment. Comparing the two- versus one-compartment model (where there is no distinction between cell edge and interior), we found that the latter significantly underestimates the barbed-end peak, and the time course of its rising phase (see Supporting Material). Spatial localization is crucial, as cofilin binding to F-actin is restricted to the edge compartment, where filaments are not protected by tropomyosin and where cofilin phosphorylation may be low. If this is true, then, as we have shown, upon release from the membrane, active cofilin strongly targets F-actin rather than being diverted to competing processes. We then attain the same rate of barbed-end production even in the concurrent presence of upregulated LIMK activity. As spatial localization appears to play an important role in determining the response of the system, further modeling based on a spatially extended system that can describe cofilin gradients forms a promising future direction.
Thus far in this model, we have considered only the action of cofilin (and first barbed-end peak) and not the dynamics of Arp2/3 associated with a second peak of barbed ends. Whereas cofilin binds preferentially to ADP-actin rather than ATP-actin monomers (7,11), Arp2/3 has a higher affinity to ATP-actin filaments. Thus, cofilin severing, which allows for new F-actin growth, also promotes subsequent filament branching by Arp2/3 (1–3). Hence, cofilin and Arp2/3 activities work in synergy to create a large increase in actin polymerization (35). Testing the boundaries of this synergy forms another interesting future direction.
Acknowledgments
We are grateful to Raibatak Das, Daniel Coombs, Dan Larson, and Enrique De La Cruz for comments and advice, and to Robert Eddy for unpublished PIP2 data.
This research was supported by a subcontract (to L.E.K.) from the National Institutes of Health (Grant No. R01 GM086882) to Anders Carlsson, Washington University, and by a Natural Sciences and Engineering Research Council discovery grant (to L.E.K.). J.C. is funded by National Institutes of Health grant CA150344. E.P. was supported by an International Graduate Training Center fellowship from the Pacific Institute for the Mathematical Sciences (Canada).
Appendix: List of Equations in Nondimensional Form
- PLC level:
where H(t) is the Heaviside function.(7) - PIP2 level:
(8) - PIP2-bound cofilin:
(9) - Active cofilin in the edge compartment:
(10) - F-actin-bound cofilin in the edge compartment:
(11) - G-actin-bound cofilin in the edge compartment:
(12) - Phosphorylated cofilin in the edge compartment:
(13) - Active cofilin in the interior compartment:
(14) - G-actin-bound cofilin in the interior compartment:
(15) - Phosphorylated cofilin in the interior compartment:
(16) - Barbed-end production:
(17)
Supporting Material
References
- 1.Mouneimne G., Soon L., Condeelis J. Phospholipase C and cofilin are required for carcinoma cell directionality in response to EGF stimulation. J. Cell Biol. 2004;166:697–708. doi: 10.1083/jcb.200405156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ghosh M., Song X., Condeelis J.S. Cofilin promotes actin polymerization and defines the direction of cell motility. Science. 2004;304:743–746. doi: 10.1126/science.1094561. [DOI] [PubMed] [Google Scholar]
- 3.Mouneimne G., DesMarais V., Condeelis J. Spatial and temporal control of cofilin activity is required for directional sensing during chemotaxis. Curr. Biol. 2006;16:2193–2205. doi: 10.1016/j.cub.2006.09.016. [DOI] [PubMed] [Google Scholar]
- 4.Sidani M., Wessels D., Condeelis J. Cofilin determines the migration behavior and turning frequency of metastatic cancer cells. J. Cell Biol. 2007;179:777–791. doi: 10.1083/jcb.200707009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ichetovkin I., Grant W., Condeelis J. Cofilin produces newly polymerized actin filaments that are preferred for dendritic nucleation by the Arp2/3 complex. Curr. Biol. 2002;12:79–84. doi: 10.1016/s0960-9822(01)00629-7. [DOI] [PubMed] [Google Scholar]
- 6.Andrianantoandro E., Pollard T.D. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell. 2006;24:13–23. doi: 10.1016/j.molcel.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 7.Blanchoin L., Pollard T.D. Mechanism of interaction of Acanthamoeba actophorin (ADF/Cofilin) with actin filaments. J. Biol. Chem. 1999;274:15538–15546. doi: 10.1074/jbc.274.22.15538. [DOI] [PubMed] [Google Scholar]
- 8.Carlier M.-F., Laurent V., Pantaloni D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: implication in actin-based motility. J. Cell Biol. 1997;136:1307–1322. doi: 10.1083/jcb.136.6.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pope B.J., Gonsior S.M., Weeds A.G. Uncoupling actin filament fragmentation by cofilin from increased subunit turnover. J. Mol. Biol. 2000;298:649–661. doi: 10.1006/jmbi.2000.3688. [DOI] [PubMed] [Google Scholar]
- 10.van Rheenen J., Condeelis J., Glogauer M. A common cofilin activity cycle in invasive tumor cells and inflammatory cells. J. Cell Sci. 2009;122:305–311. doi: 10.1242/jcs.031146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Blanchoin L., Pollard T.D. Interaction of actin monomers with Acanthamoeba actophorin (ADF/cofilin) and profilin. J. Biol. Chem. 1998;273:25106–25111. doi: 10.1074/jbc.273.39.25106. [DOI] [PubMed] [Google Scholar]
- 12.Oser M., Condeelis J. The cofilin activity cycle in lamellipodia and invadopodia. J. Cell. Biochem. 2009;108:1252–1262. doi: 10.1002/jcb.22372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang W., Eddy R., Condeelis J. The cofilin pathway in breast cancer invasion and metastasis. Nat. Rev. Cancer. 2007;7:429–440. doi: 10.1038/nrc2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.van Rheenen J., Song X., Condeelis J.S. EGF-induced PIP2 hydrolysis releases and activates cofilin locally in carcinoma cells. J. Cell Biol. 2007;179:1247–1259. doi: 10.1083/jcb.200706206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gorbatyuk V.Y., Nosworthy N.J., King G.F. Mapping the phosphoinositide-binding site on chick cofilin explains how PIP2 regulates the cofilin-actin interaction. Mol. Cell. 2006;24:511–522. doi: 10.1016/j.molcel.2006.10.007. [DOI] [PubMed] [Google Scholar]
- 16.Moriyama K., Iida K., Yahara I. Phosphorylation of Ser-3 of cofilin regulates its essential function on actin. Genes Cells. 1996;1:73–86. doi: 10.1046/j.1365-2443.1996.05005.x. [DOI] [PubMed] [Google Scholar]
- 17.Song X., Chen X., Eddy R.J. Initiation of cofilin activity in response to EGF is uncoupled from cofilin phosphorylation and dephosphorylation in carcinoma cells. J. Cell Sci. 2006;119:2871–2881. doi: 10.1242/jcs.03017. [DOI] [PubMed] [Google Scholar]
- 18.Ojala P.J., Paavilainen V., Lappalainen P. Identification of yeast cofilin residues specific for actin monomer and PIP2 binding. Biochemistry. 2001;40:15562–15569. doi: 10.1021/bi0117697. [DOI] [PubMed] [Google Scholar]
- 19.Tobacman L.S., Korn E.D. The kinetics of actin nucleation and polymerization. J. Biol. Chem. 1983;258:3207–3214. [PubMed] [Google Scholar]
- 20.Mullins R.D., Heuser J.A., Pollard T.D. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proc. Natl. Acad. Sci. USA. 1998;95:6181–6186. doi: 10.1073/pnas.95.11.6181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.De La Cruz E.M. Cofilin binding to muscle and non-muscle actin filaments: isoform-dependent cooperative interactions. J. Mol. Biol. 2005;346:557–564. doi: 10.1016/j.jmb.2004.11.065. [DOI] [PubMed] [Google Scholar]
- 22.McGough A., Pope B., Weeds A. Cofilin changes the twist of F-actin: implications for actin filament dynamics and cellular function. J. Cell Biol. 1997;138:771–781. doi: 10.1083/jcb.138.4.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.DesMarais V., Ichetovkin I., Hitchcock-DeGregori S.E. Spatial regulation of actin dynamics: a tropomyosin-free, actin-rich compartment at the leading edge. J. Cell Sci. 2002;115:4649–4660. doi: 10.1242/jcs.00147. [DOI] [PubMed] [Google Scholar]
- 24.Mogilner A., Edelstein-Keshet L. Regulation of actin dynamics in rapidly moving cells: a quantitative analysis. Biophys. J. 2002;83:1237–1258. doi: 10.1016/S0006-3495(02)73897-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bagheri-Yarmand R., Mazumdar A., Kumar R. LIM kinase 1 increases tumor metastasis of human breast cancer cells via regulation of the urokinase-type plasminogen activator system. Int. J. Cancer. 2006;118:2703–2710. doi: 10.1002/ijc.21650. [DOI] [PubMed] [Google Scholar]
- 26.Hotulainen P., Paunola E., Lappalainen P. Actin-depolymerizing factor and cofilin-1 play overlapping roles in promoting rapid F-actin depolymerization in mammalian nonmuscle cells. Mol. Biol. Cell. 2005;16:649–664. doi: 10.1091/mbc.E04-07-0555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang W., Wyckoff J.B., Condeelis J.S. Coordinated regulation of pathways for enhanced cell motility and chemotaxis is conserved in rat and mouse mammary tumors. Cancer Res. 2007;67:3505–3511. doi: 10.1158/0008-5472.CAN-06-3714. [DOI] [PubMed] [Google Scholar]
- 28.Cao W., Goodarzi J.P., De La Cruz E.M. Energetics and kinetics of cooperative cofilin-actin filament interactions. J. Mol. Biol. 2006;361:257–267. doi: 10.1016/j.jmb.2006.06.019. [DOI] [PubMed] [Google Scholar]
- 29.Prochniewicz E., Janson N., De la Cruz E.M. Cofilin increases the torsional flexibility and dynamics of actin filaments. J. Mol. Biol. 2005;353:990–1000. doi: 10.1016/j.jmb.2005.09.021. [DOI] [PubMed] [Google Scholar]
- 30.De La Cruz E.M. How cofilin severs an actin filament. Biophys Rev. 2009;1:51–59. doi: 10.1007/s12551-009-0008-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.McCullough B.R., Blanchoin L., De la Cruz E.M. Cofilin increases the bending flexibility of actin filaments: implications for severing and cell mechanics. J. Mol. Biol. 2008;381:550–558. doi: 10.1016/j.jmb.2008.05.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bamburg J.R. Proteins of the ADF/cofilin family: essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 1999;15:185–230. doi: 10.1146/annurev.cellbio.15.1.185. [DOI] [PubMed] [Google Scholar]
- 33.Pantaloni D., Le Clainche C., Carlier M.-F. Mechanism of actin-based motility. Science. 2001;292:1502–1506. doi: 10.1126/science.1059975. [DOI] [PubMed] [Google Scholar]
- 34.Chan C., Beltzner C.C., Pollard T.D. Cofilin dissociates Arp2/3 complex and branches from actin filaments. Curr. Biol. 2009;19:537–545. doi: 10.1016/j.cub.2009.02.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.DesMarais V., Macaluso F., Bailly M. Synergistic interaction between the Arp2/3 complex and cofilin drives stimulated lamellipod extension. J. Cell Sci. 2004;117:3499–3510. doi: 10.1242/jcs.01211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pollard T.D., Blanchoin L., Mullins R.D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu. Rev. Biophys. Biomol. Struct. 2000;29:545–576. doi: 10.1146/annurev.biophys.29.1.545. [DOI] [PubMed] [Google Scholar]
- 37.Dawes A.T., Bard Ermentrout G., Edelstein-Keshet L. Actin filament branching and protrusion velocity in a simple 1D model of a motile cell. J. Theor. Biol. 2006;242:265–279. doi: 10.1016/j.jtbi.2006.02.017. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.