

Abstract
While numerous publications have shown that apes can learn some aspects of human language, one frequently cited difference between humans and apes is the relative infrequency of declaratives (comments and statements) as opposed to imperatives (requests) in ape symbol use. This paper describes the use of declaratives in three language-competent apes and two children. The apes produced a lower proportion of spontaneous declaratives than did the children. However, both groups used declaratives to name objects, to interact and negotiate, and to make comments about other individuals. Both apes and children also made comments about past and future events. However, showing/offering/giving, attention getting, and comments on possession were declarative types made by the children but rarely by the apes.
Recent discussions about the evolution of communication have stressed a perceived distinction between humans and our closest evolutionary relatives, the great apes, wherein human nature is described as uniquely cooperative relative to the more competitive great apes (e.g. Moll and Tomasello, 2007; Tomasello, 2007). Some researchers argue that after splitting from the common ancestor, our evolutionary ancestors formed a social environment that supported the desire and drive to communicate and cooperate with others. According to this hypothesis, this socio-communicative environment resulted in a qualitative difference in the cooperative nature of humans relative to the other apes and this cooperative nature drove the evolution of human language and cognition (Moll and Tomasello, 2007). These researchers further suggest that, due to their uncooperative nature, chimpanzees are not fully capable of taking advantage of human social environments that increase cooperative and communicative abilities.
One major argument at the root of this cooperative hypothesis of the origin of language is the relative lack of declarative (strictly communicative) as opposed to imperative (request-driven) communication in apes (Gómez, 2007; Tomasello, 2007). For instance, it has been reported in multiple studies that apes cannot, as a rule, glean information from a human’s declarative cue in a cooperative paradigm (Hare and Tomasello, 2004; and see Lyn, in press for a review of the findings; e.g. Tomasello, 2007), although other, arguably more cooperative species can do so (see Miklosi and Soproni, 2006 for a review). Many of these studies utilize the object choice task where a participant is presented with two containers, one of which holds food, and an experimenter gives a cue such as a gaze, a vocalization, or a pointing gesture to indicate which container to select. Chimpanzees perform correctly, on average, 61% of the time, with only 9 out of 32 chimpanzees performing at above chance levels (Lyn, in press; although see Lyn et al., 2010). However, chimpanzees have performed better when the cues were competitive rather than cooperative – i.e. when the humans reached for the container with the food item (as if to retrieve it for themselves) as opposed to pointing to it (Hare and Tomasello, 2004). These findings have led to the hypothesis that the non-cooperative nature of chimpanzee social behavior and organization is a biological difference from humans that leads to difficulty in comprehending and producing declarative information in a cooperative context.
Not only are there reports that apes have trouble responding to declaratives emitted by others, but several publications report that apes do not use declaratives in their own communications - either in the wild or in experimental studies (Tomasello and Carpenter, 2005). Indeed, one researcher has gone so far as to say that, “Although some apes, especially those with extensive human contact, sometimes point imperatively for humans, no apes point declaratively ever” (Tomasello, 2007 italics added by the authors).
A few studies, while studying more general cognitive or communicative abilities, have attempted to elicit or observe declarative communication in apes. Tomasello and Carpenter, in a study with five chimpanzees, replicated studies with children where novel items are made to move behind an experimenter, but in view of the participant (Tomasello and Carpenter, 2005). None of the chimpanzees in this study pointed to the novel item. Similarly, Rivas (2005), studied 22 hours of videotape of 5 signing chimpanzees and noted types of utterances. Rivas noted that 2% of these apes’ utterances were classified as naming (non-request) (p. 409), but does not investigate further into the declarative or communicative nature of these names (some of which were signed when the chimpanzee was alone). These studies were small-scale investigations into the declarative communication of apes. Due to the low numbers of declaratives reported in the wild and in captivity, a larger sample of communicative behavior may give a better idea of declarative communicative abilities in great apes.
In contrast to the above findings, there has been one report of a likely declarative gesture in one wild bonobo (Veá and Sabater-Pi, 1998); and most reports of sign-language using apes have documented apes gesturing declaratively (e.g. Gardner and Gardner, 1969; Hayes, 1951; Patterson, 1978; Rivas, 2005). Additionally, although most standard-reared apes have trouble making and understanding declarative communication, apes reared in a complex socio-communicative environment have been shown to perform significantly better on declarative comprehension tasks (Lyn, in press; Lyn et al., 2010). However, to date there have been no studies focusing on the use of linguistic declaratives in these language-using apes.
The definition of declarative utterances, according to some researchers, relies on the triadic nature of the utterance (Liszkowski et al., 2004), “It is not enough that there is no clear imperative associated with a point [gesture], it must be social and triadic to be considered declarative”. Triadic interactions are those that include the signaler (the child), the recipient (the adult, usually), and a third object, person, or event. In general, ape communications are considered to be dyadic in nature (just between the signaler and the recipient) (although see Pika, 2008 for examples of triadic ape gestures; Tomasello and Camaioni, 1997)
Declarative utterances can take many forms. One can name concrete items or name items that do not have a concrete basis, for example, feelings or non-visible objects (objects they are looking for or something around the corner). One can also use declarative utterances to interact or negotiate (Child states “No more” when he/she is finished with an activity). Declaratives can also indicate mental time travel by stating an intended action or mentioning an event from the past.
The ability of nonhumans to mentally time-travel, like the ability of nonhumans to make declarations, is a topic of much discussion in the literature (e.g. Hoerl, 2008; Suddendorf and Busby, 2003; Suddendorf and Corballis, 2007b). Mental time travel is the ability to project oneself mentally into the future, to plan for specific events, or to the past to remember specific occurrences. It has been argued that this ability to project oneself out of the current time may be a uniquely human capability and that this ability may be an evolutionary discontinuity between humans and apes (Suddendorf and Corballis, 1997).
In apes, there has been relatively little research into mental time travel, (and see Byrne, 1995 for a review of the findings from the wild; Mulcahy and Call, 2006; but see Osvath and Osvath, 2008; Premack and Premack, 1983; Schwartz et al., 2005). Recently, in an imperative experiment, Menzel (1999) showed that a symbol-using chimpanzee could be shown a food being hidden. Unprompted, up to 16 hours later, she would spontaneously recruit someone to retrieve the food for her, indicating to the person where the food was hidden, and frequently naming that food at her symbol keyboard. Suddendorf and Corballis (2007a) assert that while the chimpanzee might know where the food was and what was hidden such knowledge is not evidence that she remembered the event of hiding itself. They also mention that, to date, the use of symbols by apes in so-called “ape language” studies has not provided evidence for mental time-travel as there are no reports of apes discussing past events or future episodes.
In arguably one of the most successful of the “ape language” studies, the work of Sue Savage-Rumbaugh and colleagues has shown that bonobos (Pan paniscus) and chimpanzees (Pan troglodytes) are capable of many linguistic feats: acquisition of symbolic capacities without explicit training (Greenfield and Savage-Rumbaugh, 1991; Savage-Rumbaugh et al., 1986; Savage-Rumbaugh et al., 1993; Savage-Rumbaugh et al., 1980); using symbols to name objects in double blind studies (Savage-Rumbaugh et al., 1986); associating novel English names with novel objects with very few exposures to both object and word (Lyn and Savage-Rumbaugh, 2000); utilizing imitation in an intentionally communicative context (Greenfield and Savage-Rumbaugh, 1993); making semantically-based combinations across both lexigram and gestural combinations (Greenfield and Lyn, 2007; Greenfield et al., 2008; Greenfield and Savage-Rumbaugh, 1990, 1991; Lyn et al., in press); mentally representing symbols on several levels at the same time (Lyn, 2007); and comprehending English sentences at least at a similar level to a two-and-a-half year old child tested in the same manner as the ape (Savage-Rumbaugh et al., 1993). Several of these papers describe declarative use of symbols by apes (Savage-Rumbaugh, 1986; Savage-Rumbaugh et al., 1986), reporting that declaratives were a relatively low percentage of all ape utterances (approximately 10%). In contrast, in a similar study of early language in children, Greenfield and Smith report that children at the one-word stage use declaratives in as many as 80% of their utterances (Greenfield and Smith, 1976).
However, infrequent phenomena, such as declarative communication in apes, can be significant in evolution because natural selection sometimes utilizes genes for infrequent, but adaptive behavior. As the survival rate of individuals displaying said behavior increases, the behavior becomes more frequent. Similarly, sometimes infrequent behaviors piggy-back on genes for other behaviors. Those infrequent behaviors may not be adaptive at all, until an environmental change occurs and those previously infrequent behaviors become more adaptive, and then more frequent. Given this evolutionary tendency, a behavior, however infrequent (in this case, declarative communication), present in all members of an evolutionary clade (all member species diverging from an evolutionary node (like that which gave rise to humans, bonobos, and chimpanzees) (Byrne, 1995)) would be assumed to have been present in the evolutionary ancestor. The genes that gave rise to that behavior would therefore be excluded as a candidate for a possible biological change in one of the divergent species. We have put forth similar arguments in earlier papers for hierarchical categorization (Lyn, 2007), complex mental representation of symbols (Lyn, 2007), word ordering preferences (Lyn et al., in press) and pretense (Lyn et al., 2006). However, it is possible that, as has been suggested by others (Moll and Tomasello, 2007; Tomasello, 2007), declarative communication in apes is so infrequent that one could dismiss it as an artifact of the data being collected by humans who are sensitive to possible declarations.
Recently, the databases of symbol use from both the Greenfield study of language development in human children and the Savage-Rumbaugh study of symbol use in apes were joined into a combined data set – comprising both ape and child symbol use. This database is unique – composed of over 100,000 “utterances” of apes using a symbolic keyboard as well as over 4000 verbal utterances of children at a similar linguistic stage of development (Gillespie-Lynch et al., in review). Because sets of utterances were recorded according to similar protocols and coded using the same coding scheme regardless of species, the database provides the first truly comparative record of human and ape symbol use. This paper aims to utilize this database to delineate and compare the use of declaratives in apes and children to attempt to answer several questions: Is the symbolic use of declaratives a quantitative or a qualitative difference between apes and humans? Are some of these declaratives triadic in nature? Do the apes use symbols to talk about the past or the future? What are the similarities and differences in declarative use and development in ape and child?
Methods
Participants and their Environments
Apes
Our ape participants were Kanzi, a bonobo (Pan paniscus), Panbanisha, his half sister, and Panpanzee, a chimpanzee (Pan troglodytes) born within six weeks of Panbanisha, and reared alongside her. All three were reared at the Language Research Center, in Atlanta, Georgia in a within- and cross-species communicative environment. This communicative environment consisted of gesture, speech, and written visual symbols (lexigrams) placed on a keyboard. The lexigram keyboard was made available to the apes at all times and some of the available keyboards emitted the sounds of a computer-synthesized English word when the corresponding key was touched. While utilizing any of these keyboards, the caregivers were also instructed to communicate in English. Therefore, English and lexigram use were paired in the communicative environment of the apes.
Emphasis on keyboard use and spoken English began earlier with Panbanisha and Panpanzee than with Kanzi (before they were six weeks old, as opposed to Kanzi’s several months old). Unlike Kanzi, who was initially exposed to a keyboard of 6 symbols during reward-based training sessions with his mother (the first keyboard that he used himself contained 12 symbols), Panbanisha and Panpanzee were initially exposed to a keyboard of more than 256 symbols and were expected to learn the use of these symbols without specific training. Like Kanzi, Panbanisha and Panpanzee were in the company of human caregivers and each other 24 hours a day, seven days a week, although this experience began for them at birth and did not begin for Kanzi until he reached 2.5 years of age (Savage-Rumbaugh et al., 1986). Whereas Kanzi produced his first meaningful lexigram at 30 months of age, Panbanisha and Panpanzee produced theirs around age one, a starting point very similar to that of a human child. (For further details of rearing and linguistic capabilities, see Brakke and Savage-Rumbaugh, 1995, 1996; Savage-Rumbaugh, 1986; Savage-Rumbaugh et al., 1993)
Children
Archived data from two children, NT and MG, were used for comparative purposes. These children were raised in a normal family and home environment, and their language development was tracked by a research assistant through monthly formal observation sessions. Regular observations continued on a monthly basis for MG from 12 to 22 months of age. Regular observations continued on a monthly basis for NT from 18 to 24 months of age.
Data Collection
Apes
Panbanisha and Panpanzee were co-reared for almost five years. Beginning one year into the study, data collection included all uses of the keyboard by Panbanisha and Panpanzee, as well as communicative gestures used in combination with keyboard utterances, which were recorded by hand by the caregivers and input into the computer at the end of the day. This procedure provided the researchers with an exhaustive written record of the apes’ symbol use. The database contains each observed utterance (which could include lexigram(s), gesture(s), or combination(s) of lexigram with gesture), date, record number, ape, researcher, codes as to pragmatic force of the utterance, behavioral concordance notes, and a short contextual note. Behavioral concordance - the relationship between the utterance and the ape’s behavior, particularly his or her behavior subsequent to the utterance, was the main clue as to semantic relations and pragmatic force (Request, statement, comment, etc.). These were similar to observational protocols used in classic studies of child language which provided the comparative foundation for this study (Bowerman, 1973; Brown, 1973; Greenfield and Smith, 1976).
This same procedure was operative for Kanzi over the same five-year period, but the developmental period differed, as Kanzi was 5 years old at the beginning of the co-rearing study. At the end of this study period, the chimpanzee, Panpanzee, was moved out of the language study and no further utterance data was recorded for her. However, opportunistic data were recorded when staffing and scientific priorities allowed for Panbanisha and Kanzi utilizing these same procedures over the next 6 years, yielding a total of 11 years of data for each of the bonobos, although these data are not exhaustive.
Reliability for data collection
As a reliability check, real-time recording was checked against 4.5 hours of videotape. Thirty-seven out of 46 utterances, or 80%, were noted by both the real-time and the video observer (the other 11 were noted only by the video observer) and there was 100% agreement on the lexigram that had been used when both observers noted the utterance. Hence, we conclude that our corpus is highly reliable, but an underestimation of quantity.
Children
Data from the formal observation sessions with the children were included in the present study. During these observations, all verbal utterances were recorded. Contextual notes were included in the child data as were prior utterances and direct responses to the child’s utterance. This data collection procedure occurred prior to and served as the blueprint for the data collection with the apes.
Coding
The data from the children were added to the electronic, searchable database of ape utterances. As each record was added, it was given an unchangeable Record ID number; these record IDs were used to identify particular utterances in the database. The corpus for the apes consisted of 105,629 utterances and the children’s corpus consisted of 4445 verbal utterances. These quantities differed mainly due to data collection procedures – daily for the apes, monthly for the children; over 12 months for the children as opposed to 11 years for some of the apes. Also, the children’s sessions lasted only a few hours, whereas the ape were observed all day. To control for these differences, our analyses use percentages to report on utterance types. Gestures were not noted in the children’s utterances, but any gesture that was combined with a verbal utterance was noted in the contextual notes. This allowed for an analysis of gesture/symbol combinations for both children and apes.
Pragmatic Force
The children’s data were then coded, following the procedure set down for the ape data, for pragmatic force. Utterances were coded as: requests - utterances that served to communicate a desire for an object or activity, comments - utterances that served to refer to current, past, or future objects or events without requesting those items, statements – utterances that served to declare what the child or ape was about to do (only coded if the child or ape immediately performed the activity), structured response - an utterance in response to a caregivers’ request for a specific response, e.g.: “What’s that?”, imitation: any direct repetition of a symbol by a caregiver (e.g. Mother says “Is that a cow?”, child says “cow” or caregiver uses the keyboard to say “APPLE” and the ape responds “APPLE”), response to question – an utterance in response to a more general question e.g. “Where do you want to go?”, responding “yes” or “no” to a question, babbling, emotional vocalizations, sound imitations, and uninterpretable utterances. The apes could also have cross-modal imitations in which a caregiver stated a word in English and they replied by pressing the key glossed as that English word.
These pragmatic force types, coded as above, were then combined into supersets, of which the four most numerous for both the apes and the children were: Requests; Declaratives – combining comments and statements; Responses to a Caregiver – combining structured utterance and responses to questions; Imitation – including imitation and (for the apes) cross-modal imitation.
Reliability for Pragmatic Force Coding
A subset of 407 out of 4445 utterances were coded by an independent coder, with an agreement of 85.5%, Cohen’s kappa .84. This reliability rate is considered “almost perfect” by Landis, J.R. and Koch, G. G. (1977) (Kappa >.80). With 12 possible codes, chance agreement would be 8.3%.
Spontaneous declaratives
An evaluation was made of the spontaneity of both the ape and the child utterances. Removed from the database of spontaneous declarative utterances were: imitations, “structured” utterances (where the ape or child is asked to respond in a specific way – e.g. “Show me the lexigram for milk” or “what is that?”), direct responses to questions (e.g. “Where do you want to go?” or “Can you tell me what you’d like?”, uninterpretable utterances, utterances without proper contextual notes, and babbling (talking or “lexigramming” to oneself). Also eliminated from the declarative ape data were any examples of “testing” situations, in which an ape would be asked to name a photograph, a lexigram, or an English word. Analyses and discussion of these tests can be found in other publications (Brakke and Savage-Rumbaugh, 1995, 1996; Lyn, 2007).
Coding and Reliability for Types of Declarative
Both the apes’ and the children’s utterances were then further coded according to declarative type by the first author. Definitions of specific declarative types, their contexts, and examples are presented in table 1. For the apes, a subset of 599 of the spontaneous declarative utterances were coded by an independent coder, with reliability rate of 83% for the 11 possible codes (Cohen’s kappa .81). For the children, 364 of the declarative utterances were coded for reliability by a second independent coder; the coders agreed on 298 of these, for an agreement of 81.9%, Cohen’s kappa = .80.
Table 1.
| Concrete Item | Labeling a concrete item or behavior (Pure naming). Includes commenting on an activity they currently already engaged in or a food they are already eating. Apes sometimes use the referent to activate the keyboard. | Panpanzee: Record 63713: RAISIN: Panpanzee commented vocally and at the keyboard as she ate her raisins. Kanzi: Record 3764: CARRY: Kanzi was standing over Panbanisha and they were both scooting along in this manner, when Kanzi said CARRY at the keyboard. He seemed to be commenting about his method of “carrying” Panbanisha. MG: Record 108910: “Boy”: Pointed to picture of boy. Mother responded “That’s boy”. |
| Non-concrete Item | When labeling a non-concrete referent. For example a feeling they have, or non-visible referent (e.g. missing (“gone”), something they are looking for, or opening something and guessing what’s inside) | Panbanisha: Record 45517: SCARE: Panbanisha said SCARE and looked over to the generator box. It was indeed making a soft humming noise. We have seen scary things happen at that box before and Panbanisha may have thought it was going to come alive again. Panpanzee: Record 84531: QUIET: On the way to look, Liz and Panbanisha find a turtle. Liz brings it to look. Panpanzee is a little put off by the turtle and makes this comment. She is a little hesitant to touch it whereas Panbanisha picks it up with no problem. Panpanzee also covers her eyes as she makes this comment as if she’s thinking about hiding from the turtle. NT: Record 112079: “Gone”: Opened empty box. Mother responded “gone, it’s all gone” |
| Future plan / statement of intention | Statement of intention (what he/she is going to do). during an activity where they are planning to obtain an object/ perform an activity | Kanzi: Record 2800: ICE: Kanzi touches the ICE symbol, then goes to small refrigerator, opens the door, scrapes ice from freezer and eats. MG: Record 109182: “Bank”: Saw bank, we were going there. |
| Past | Reference to past activity | Panpanzee: Record 29420: JUMP: I had been helping Panpanzee JUMP off the sloped platform in bdrm. one time she stayed up there for quite a while before working up the nerve to jump onto the table next to me. As she landed she lay on her tummy and touched the JUMP lexigram on the faceplate there. She appeared to be thinking about what she had just done. MG: Record 109221: “Hurt hurt”: Lauren had hit him. |
| Interaction / Negotiation | Use of a symbol to distract from a topic, joining into a conversation, or requesting information. | Panbanisha: Record 56265: AUSTIN: After I returned, Panbanisha left Liz and began to climb on the top of the playyard. We told her this was bad and asked her to get down. She did and while we were attempting to discuss about being bad, she repeatedly pounded on the AUSTIN key with her finger as though to distract us away from the topic, which she did not seem to want to hear about. Panbanisha: Record 84886: REFRIGERATOR: After Panbanisha pointed to the refrigerator lexigram, I look questioningly at her as I was unsure as to what she was trying to communicate. She gestured to the butter out on the counter that we had gotten from the refrigerator around ten minutes earlier. Since she seemed to be asking me to show her the name (lexigram) for butter, I told her “Butter” on the keyboard. NT: Record 113454: “bye bye”: Vicky was leaving |
| Possession | Commenting on another’s possession of something | MG: Record 109207: “Mine”: Pointing to papers in Lauren’s pocket NT: Record 112360: “dede” (lady): Picked up a piece of paper that was Vicky’s. Mother responds, “Yes, that’s the lady’s” This Declarative Type was not recorded for the apes |
| Show / offer / give | Shows / offers / or hands the referent to someone else. | Kanzi: Record 13381: SWEET POTATO: After removing the sweet potato from the cooler at the A-frame, Kanzi handed it to me and said SWEET POTATO on the keyboard. Then he went to play with Panpanzee, showing no immediate interest in eating the food. (He did ask to eat it about twenty minutes later) NT: Record 108550: “Cars”: Held up his car. Mother responds, “Cars, yes” |
| Attention getting | Speaking purely for gaining the attention of another | MG: Record 110423: “Mama”: Mother responds, “What?” NT: Record 111393: “watch this one. watch this one.”: Mother responds, “Yes, we’re watching”. This Declarative Type was not recorded for the apes |
| Multi- Declarative (MD) combination | Declarative combinations where the elements belong to multiple types of declarative | Panbanisha: Record 16664: SHIRT ON BAD: Before going out, the experimenter & Panbanisha put shirts on. Panbanisha was hesitant to put hers on. The experimenter encourages her to do so. Panbanisha points to the SHIRT and ON lexigrams, looks at the experimenter, and then points to BAD. Panbanisha seems to express that she doesn’t want her shirt on & feels “bad” about it. Nonconcrete naming, interaction/negotiation MG: Record 109813: “Bye bye door”: Bula was going out the door. Interaction/negotiation, about another |
| About another | Commenting on another person/ape, not coded when involved in the activity themselves. | Panbanisha: Record 38674: TICKLE: I had been tickling Panpanzee for several minutes. At one point, Panpanzee laughed loudly when I tickled her underneath the chin. Panbanisha commented TICKLE and then went back to looking at her new book. NT: Record 109492: “Break”: Mother was breaking eggs. |
| Uninterpretable, | Unclear from the context which code, if any, is applicable. Sometimes overgeneralizations | Panpanzee: Record 25777: DOG: Panpanzee commented dog as I was putting her to sleep. Panbanisha: Record 9905: MILK: Steve had been playing with Panbanisha and then went out the door. Panbanisha watched him go, then ran up to the door, touched MILK on the keyboard, and ran back under the table. MG: Record 108777: “House house”: Digging in sand with the stick. |
Triadic interaction coding
While each utterance was coded within the original database as to overall context, these contextual notes did not, as a rule, include information on eye contact or attention switching – all information that is required to determine the triadic or dyadic nature of the utterance. Some contextual notes did, however, mention either specific behaviors such as eye contact or a more general indication of triadic interaction, such as “Panbanisha was looking at me as she commented”. When such notes were available, the utterances were coded as triadic interactions. Because these notes were ad hoc, with no instructions to the experimenters, no quantitative measures can be applied to triadic interactions. However, qualitative analysis will be used to demonstrate that some declaratives were triadic in nature. There was also no information about triadic interactions systematically recorded within the children’s data, but examples of likely triadic interactions (chosen with the same criteria as the ape examples) will be presented.
Data Analysis
Data were analyzed using SPSS statistical software to perform chi-square analyses on individual differences on utterance types. Because our interest was in both individual and group differences, the SPSS crosstabs analysis was utilized to compute adjusted residuals. These residuals are on a Z-scale, thus each individual’s residual can be compared to the others, and any performance groupings can be noted. Any adjusted residual over the absolute value of 1.96 was considered to deviate from the expected (p<.05).
Results
Corpora
Of the apes’ 105,629 utterances, 78,021 (73.8%) were spontaneous lexigram and gestural utterances by our three participants. Both apes and children made utterances that were coded as declaratives, requests, imitations, and responses to caregivers. The children’s corpus of 4445 verbal utterances had a lower percentage of spontaneous utterances (2399 or 54.0%). The rest were conversational responses of various sorts. Each subject had an individualized pattern of utterance types (X2(12)=11876.5, p<001), however that individual pattern was strongly affected by genus (the apes and the children responded significantly differently) (see Fig 1). Adjusted residuals showed that the genus difference were most clear for requests, with apes showing more requesting utterances (adjusted residuals, requests: Kanzi −4.4; Panbanisha −3.1, Panpanzee −4.3; MG 4.7; NT 6.9). In contrast the apes generally had a much lower proportion of spontaneous declaratives (5654 out of the 105,629 total utterances (5.4%)) than the children (1797 (40.4%)) (adjusted residuals, declaratives: Kanzi −16.1; Panbanisha 0.3 (ns), Panpanzee −29.5; MG 40.4; NT 77.5). The majority of the apes’ utterances were requests, as has been reported elsewhere (Savage-Rumbaugh et al., 1986). In contrast, the results for imitation and response to caregiver were more mixed, but in general, the children had more utterances in these two categories than did the apes (adjusted residuals, imitation: Kanzi −15.1; Panbanisha −7.8, Panpanzee 6.9 MG 8.7; NT 26.0; responses to caregivers: Kanzi 18.3; Panbanisha −0.1 (ns), Panpanzee 0.4 (ns) MG 25.1; NT 15.4) (Fig 1).
Figure 1.
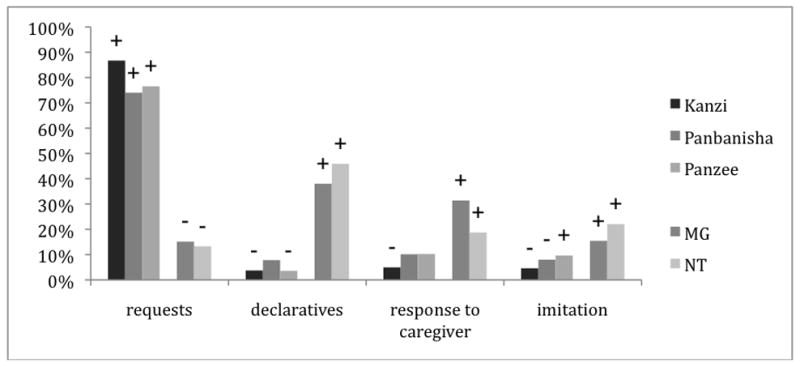
Percentage of utterances across pragmatic types.
− indicates a residual that was significantly lower than expected by chance
+ indicates a residual that was significantly higher than expected by chance
Declaratives
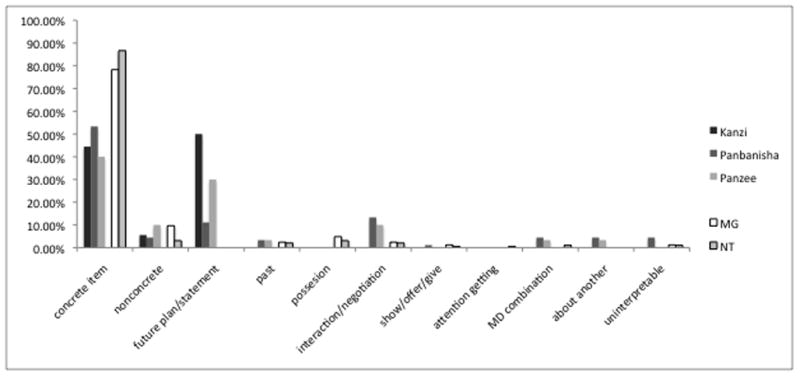
Ape and Child Comparison
All participants made many of the same kinds of declarative utterances (see Figure 2). Again, results indicate that each subject had an individualized pattern of utterance types (X2(40)=1368.5, p<001), but that, in general, apes and children responded differently from each other. These results are best demonstrated by three types of declaratives that were made fairly frequently by the children, but rarely, if ever, by the apes. These types were possession, show/offer/give, and attention getting. The only examples of these declarative types by the apes were 11 instances of show/offer/give.
Figure 2.

Percentage of utterances across declarative types
− indicates a residual that was significantly lower than expected by chance
+ indicates a residual that was significantly higher than expected by chance
Examples of this type of declarative in a child and an ape follow:
MG: Record 110353: “Dirty”: Held up his dirty finger. Mother responds, “Yeah, it’s dirty.” Age 1:5
Panbanisha: Record 5637: “Balloon”: She let all the air out again- held it up and said balloon. Age: 2:0
In addition to the declarative types mentioned above, a greater proportion of the children’s declaratives were concrete and nonconcrete naming. In contrast, the apes had more declaratives describing future plans/statements, interaction/negotiation, declaratives about another, and uninterpretable utterances (see Fig. 2).
The apes had more “future” declaratives, including both utterances describing plans that had already been made (apes, 364 (4.6%), children − 30 (1.7%)) and statements of intentions – spontaneous declaratives about actions the apes or children immediately followed up on (apes − 3742 (47%), children − 67 (3.7%)). Examples:
Panbanisha: Record 4820: “GROOM”: Panbanisha said GROOM and then began to groom her foot. Age: 1:9
MG: Record 109821: “Light”: He’s going to the switch to turn out the light. Age: 1:4
The apes also had higher numbers of interaction and negotiation declaratives, although the adjusted residuals indicate that this difference is almost entirely due to Panbanisha (adjusted residuals, interaction and negotiation: Kanzi −3.2; Panbanisha 10.0, Panpanzee −4.2; MG −3.2; NT −5.4). These codes encompass several subcategories. For example, toward the end of the first year of data collection the apes began to use the lexigram for their current activity to announce a change in activity. For example:
Panbanisha: Record 4623: EGG: As Panbanisha finished her egg, she said EGG and started to get out of her cube. Liz had mentioned earlier that Panbanisha sometimes had been announcing when she was finished with something and ready to do something else. This appeared to be what Panbanisha was doing. Age: 1:9
Kanzi: Record 14101: GRAB: After a long grabbing bout, Kanzi said GRAB at the keyboard and then moved away (no longer in position to continue the game). Age: 6:7
Panpanzee: Record 59577: SURPRISE (meaning a food without a specific lexigram): Panpanzee ate about half of her cookie, then touched SURPRISE and handed it to me. She then instigated play. Age: 3:3
There are no similar examples of the children announcing a change of activity only by naming the current activity. However, the children did announce their preference to change a current activity, most often by using “no”, frequently combined with the non-preferred activity. For example:
NT: Record 112594: “no”: Mother had said, “That’s the record” and put the record on. After NT said “no”, she responded, “No? Should I take it off?” Age: 1:10
MG: Record 109217: “no more”: handed half-eaten pretzel to mother. Age: 1:10
NT: Record 110049: “no penny”: Matthew was taking some of his pennies and NT did not want him to do so. Age: 2:0
Another type of sub-category was within concrete naming - the labeling of an item of which the participant was already in possession (see table for examples). Both the children and the apes made this type of declarative quite frequently (between 14.4% (Panbanisha) and 23.3% (MG). The apes also used referents on the keyboard to make a declarative statement. For example:
Kanzi: Record 1022: BALLOON: After the balloon popped, Kanzi takes the pieces and placed them on top of the balloon lexigram. Age: 5:0
Panbanisha: Record 17896: ICE: Panbanisha was lying on her back with a piece of ice in her hand. She reached up with the ice and put it on the ice lexigram, naming/commenting to herself. Age: 1:10
Past and future events
Both the apes and the children made reference to past events. Kanzi made 38 (10.2%) utterances about past events, Panbanisha made 288 (6.6%), and Panpanzee made 87 (9.3%). This was similar to the frequencies of the children (MG: 30, 6.3%; NT: 80, 6.7%). These past events were most frequently something that had just occurred: Examples:
NT: Record 112042: “drop”: He’s looked at a nut on the floor that he had dropped. Mother responds, “Did one drop? Yes, one dropped.” Age: 1:7
Panbanisha: Record 17186: “TICKLE”: We had been watching Panpanzee and Krista tickle with a pair of children’s scissors. When they had finished and gone to the other side of the room, Panbanisha went over, picked up the scissors, and said TICKLE at Panpanzee’s keyboard. She then examined the scissors. Age: 2:0
All of the children’s “past” utterances were of this type. However, there are also examples of the apes commenting on things that had happened earlier in the day, or even on a previous day. These examples occurred when the apes were at older ages than the children were during the study. Examples:
Kanzi: Record 9277: “M&M”: Kanzi said M&M at the faceplates. Verbally I commented that we had M&MM’s a bit earlier and he vocalized. I commented on several other foods we ate that morning and he vocalized at this, also. He did not indicate at this time that he wanted to eat any of these foods. Age 6:3
Panbanisha: Record 39547: “TURTLE”: When Panbanisha pointed to TURTLE, I asked Laura if they had seen a turtle this afternoon while they were out in the woods. Laura replied that they had seen a squashed turtle. NOTE: Jeannine and I had commented on how gross it looked. It is likely that Panbanisha was referring to the squashed turtle since it was such an unusual occurrence to find a turtle as flat as a pancake. Age 2:8
Panpanzee: Record 39547: “Hide gesture”: Panpanzee put her hand over her eyes in a “hide” gesture as we drove past a place I had hidden yesterday. She appeared to be commenting, remembering our activity. Age 2:11
Both children and apes also made reference to future events. As described above, many of the apes’ future statements were a way of letting experimenters know what they intended to do. However, some utterances described plans that had been made or anticipated future events. Examples:
Kanzi: Record 3783: M&M: Kanzi and I got way ahead of the group as we were chasing to get M&M’s. He stopped to let the others catch up with us at one point. Panbanisha and Panpanzee were walking together, holding onto each other. Kanzi approached them and said M&M, as if to tell them that we were still getting M&M’s and for them to hurry up. Age: 6:7
Panbanisha: Record 31570: MILK LATER: I got a can of milk out of the refrigerator but did not give any to the apes yet, since they both needed baths after playing with the dogs. Panbanisha gesturally requested it several times but I told her she needed a bath first. Finally she made this combination and was more compliant with her bath. She knew she could have her milk later, as soon as she was washed, so she became more cooperative after making this comment. Age: 2:11
Panpanzee: Record 37334: SURPRISE: Panpanzee and I were looking for surprises when we ran into Phil. I asked Panpanzee to tell Phil what we were searching for and she did so and hugged him. He volunteered to look for a surprise and Panpanzee sat and waited. While waiting, Panpanzee made a nest and commented on my faceplate SURPRISE. Age: 2:11
The children also made these kinds of anticipatory utterances. Examples:
MG: Record 108756: “quack quack”: Took piece of bread out. Mother had talked about going to feed the ducks a few minutes earlier. Mother responded, “yea we’ll take it for the duck.” Age: 1:8
NT: Record 109029: “baby”: in anticipation of mom singing a song about a baby. Age: 1:11
Triadic interactions
Both children and apes made triadic utterances (communicating with a caregiver while referring to a third items). Within the apes’ declarative utterances were 1078 utterances in which the written context explicitly suggested a triadic interaction. Examples include:
Panbanisha: Record 54397: STRAWBERRY: Panbanisha was eating one of the fresh strawberries that Rachelle had brought. At one point she approached the large keyboard and commented about this, looking at the experimenter, who agreed that Panbanisha had a strawberry. Age: 2:1 (concrete item)
Panpanzee: Record 34709: Panpanzee was playing in Matata’s cage when she approached the keyboard to say PLAY, making eye contact with me. She then resumed climbing and chasing around. Age 3:10 (concrete naming)
The children had several utterances in which the context explicitly suggested a triadic interaction (many more interactions were implicitly triadic).
NT: Record 112033: “Spoon”: He picks up a spoon and looks at mother. Mother responds, “Yes, that’s a spoon.” Age: 1:7 (concrete item)
MG: Record 111075: “Airplane airplane”: He ran to his mother and then came back and pointed to the airplane in the book. Age: 1:7 (concrete item)
Gestures
While gestures were not explicitly coded in the children’s utterances, contextual notes and previous research (e.g. Greenfield et al., 2008; Greenfield and Smith, 1976; Lyn et al., in press) suggest that this is another point of similarity between the children and the apes. Gestures were used by all of the apes as an element of some declarative utterances − 4.8% of Kanzi’s declaratives included a gesture, as did 2.7% of Panbanisha’s declaratives and 8.8% of Panpanzee’s. Utterances in which gestures were the only element would not have been recorded in the children’s data (so we cannot assess this category in the children), but were present in the apes’ data. Panbanisha had 28 gesture-only utterances and Panpanzee had 52. These utterances were classified as concrete item, nonconcrete item, future plan/statement, past, interaction/negotiation, show/offer/give, about another, and uninterpretable. Some examples follow:
Panpanzee: Record 18365: Show gesture: Panpanzee was playing keep-away with a small piece of paper with Panbanisha. Panbanisha stopped. Panpanzee very carefully took the hand with the paper in it and held it out to show it to Panbanisha. Age: 1:11 (show/offer/give)
Panbanisha: Record 28529: After finding Karen and Panpanzee who had hidden along the trail from hilltop, Panbanisha approached them and put her hands over her eyes in the hide gesture. We all agreed they’d been hiding. Age: 2:7 (past)
Panbanisha: Record 76535: Wave gesture: Panbanisha waved goodbye to John in the observation room. Age: 4:2 (interaction)
In some cases, a gesture was the sole element of the utterance and the contextual notes indicated triadic interaction. panbanisha had 18 of these type of utterances and panpanzee had 14.
Examples include:
Panpanzee: Record 18455: Point gesture: As Panbanisha, Panpanzee and the experimenters were returning from evening car rounds, there was a jet above. As the experimenter walked, Panpanzee then pointed up at the sky to the plane and looked at the experimenter. The experimenter agreed it was a plane and pointed. Age: 1:11
Panbanisha: Record 6259: Point gesture: Panbanisha and Panpanzee both got very frightened at the sound made when the clock above the fridge fell off its mounting. A little while later Panbanisha approached it, then stood bipedaly & piloerect, looked at the experimenter, and gestured up to it. Age: 2:1
Gestures were also combined with symbols (either lexigram symbols or verbal words) by both the children and the apes. The children had more gesture/symbol combinations (278, 15.5%) than did the apes (138, 2.4%). A full examination of the development of symbol and gesture combinations can be found elsewhere (Greenfield, Lyn, and Savage-Rumbaugh, 2008), but when we restrict ourselves to declarative symbol-gesture combinations, we still find many of the same kinds of declarative types in ape and child. Both children and apes used gesture-symbol combinations for concrete naming (Figure 3). These combinations frequently involved the child or ape naming an item and pointing to that item. However, the children showed far more examples of concrete naming (234, 84.2%) than did the apes (68, 49.3%). The children seemed to use concrete naming most frequently during interactions with their caregivers. Specifically, the children frequently sat down with their mothers and pointed to pictures in books, while naming them. The apes did not have these same kinds of interactions (any “sitting down and naming items” was considered a testing situation and therefore was not included in the utterance database).
Figure 3.
Percentage of symbol/gesture combinations across declarative types
In contrast, the apes had more gesture-symbol combinations in the future/plan (28, 20.2%) and the interaction/negotiation (15, 10.9%) types than the children did (kids: future/plan (0, 0%), interaction/negotiation (6, 2.1%). These differences paralleled those seen in utterances that were not symbol/gesture combinations.
Discussion
When exposed to a complex socio-communicative environment, both apes and children made triadic, declarative communications. This finding holds for many definitions of declaratives, including triadic “showing” behaviors, which, while rare, did occur in our sample. Further, both children and apes made many of the same types of declarative utterances. Both named concrete items at a high rate (between 33% and 58% of their declaratives), made declarative statements about others and named nonconcrete items, such as items that were out of visual range or subjective referents, such as feelings. Apes also named items they already possessed, a definitively declarative act as they were unlikely to be requesting an item they already had in their possession. All of these findings suggest that any difference between the use of declaratives in humans and apes is quantitative, rather than qualitative in nature.
Both apes and humans made statements regarding future actions and past events, suggesting some level of mental time travel (Suddendorf and Busby, 2003; Suddendorf and Corballis, 2007b). Apes made many more statements about what they intended to do than do the children. Qualitatively, the apes made past statements about events further in the past than did the children. This is consistent with developmental findings from human children that the ability to communicate about mental time travel increases in preschoolers from 3 years of age to 5 years of age (Atance and Jackson, 2009; Busby and Suddendorf, 2005).
Some of the differences between the apes and the children may be due to data collection differences (where any sort of structured naming in the apes would be considered a testing situation) and even more so to the differences in caregiver expectations and interpretation. Ape caregivers tended to interpret any utterance as a request, unless behaviorally, the ape made it clear that they were not requesting an item (examples of these sorts of interactions can be seen on the fourth author’s website http://kanzi.bvu.edu). This tendency to ignore declaratives may have led the apes to make fewer declaratives.
These kinds of caregiver effects can be seen in the data presented here. The rates and types of utterances are affected in both ape and child by caregiver interactions and expectations. In contrast to early reports that apes imitate symbol use more than do children (e.g. Terrace et al., 1979), the children had larger numbers of imitations and responses to caregivers suggesting either that their caregivers were interacting more with the children, or that the children responded more to those interactions. The children also showed large numbers of concrete declaratives, increased greatly by the many “naming” interactions with their caregivers.
The apes, on the other hand may have responded to social demands of their caregivers, by increasing their already high numbers of future/plans and statements declaratives over the developmental span. This highlights a distinct difference in the way caregivers interacted with apes vs. children. The caregivers exerted more control over the apes’ movements, especially as they got older, while caregivers were more likely to engage in naming (as opposed to testing) interactions with the children. These findings support other findings that show that the social environment in which apes are raised significantly affect their ability to understand declarative communication (Lyn et al., 2010) and together, these findings contradict the implications in recent papers that apes cannot respond to cooperative social environments by producing declarative communications (Moll and Tomasello, 2007).
To date, there is only one record of chimpanzees or bonobos possibly communicating declaratively in the wild (Veá and Sabater-Pi, 1998). While studies of wild apes are not so definitive that one can say that wild apes do not communicate declaratively, it does seem to be rare, if not entirely absent or in a modality that current observation methods do not capture. However, apes when reared in a human social environment do understand and produce declarative communication, showing that, biologically, both chimpanzees and bonobos are capable of utilizing the environment to support declarative communication. As the end result is a distinction of quantity, we must look elsewhere for a qualitative communicative difference between ourselves and our nearest evolutionary relatives.
It is important to note, however, that the quantitative difference found here in declarative production may still indicate an important difference between apes and humans – possibly suggesting mechanisms for the development of human language. In particular, the extreme rarity of these declarative events in wild apes as well as in our sample suggest that while apes have the biological capability to declare, there might be an environmental trigger that must occur to express this ability phenotypically. This is similar to the “Vygotskian hypothesis” (Moll and Tomasello, 2007) that suggests that cooperative, declarative behavior is socially supported in humans, but expands the range of possible social support to our evolutionary relatives, the great apes. For example, a environmentally-regulated gene that triggers declarative communication mechanisms may have become permanently expressed in humans within the evolutionary time frame of the development of human language. Thus a small change switches humans from “capable of declarative communication” to “consistently declarative communicators.”
Acknowledgments
Funding for this study was provided by NIH grant HD-56232 and HD-38105. Database preparation for the child data was supported by a grant from the Leakey Foundation to Patricia Greenfield and Heidi Lyn and by Summer Research Mentorships from UCLA Graduate Division to Kristen Gillespie-Lynch. Child data collection was supported by the Harvard Center for Cognitive Studies. Collection of the original ape data was supported by NICHD grant #06016 to the Language Research Center and National Center for Research Resources #RR-00165 to the Yerkes National Primate Research Center.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Atance CM, Jackson LK. The development and coherence of future-oriented behaviors during the preschool years. Journal of Experimental Child Psychology. 2009;102:379–391. doi: 10.1016/j.jecp.2009.01.001. [DOI] [PubMed] [Google Scholar]
- Bowerman M. Structural relationships in children’s utterances: Syntactic or semantic? In: Moore TE, editor. Cognitive development and the acquisition of language. Academic Press; New York: 1973. pp. 197–213. [Google Scholar]
- Brakke KE, Savage-Rumbaugh ES. The development of language skills in bonobo and chimpanzee - I. Comprehension. Language and Communication. 1995;15:121–148. [Google Scholar]
- Brakke KE, Savage-Rumbaugh ES. The development of language skills in Pan - II. Production. Language and Communication. 1996;16:361–380. [Google Scholar]
- Brown R. A first language. Harvard University Press; Cambridge: 1973. [Google Scholar]
- Busby J, Suddendorf T. Recalling yesterday and predicting tomorrow. Cognitive Development. 2005;20:362–372. [Google Scholar]
- Byrne RW. The thinking ape: evolutionary origins of intelligence. Oxford University Press; Oxford; New York: 1995. [Google Scholar]
- Gardner RA, Gardner BT. Teaching sign language to a chimpanzee. Science. 1969;165:664–672. doi: 10.1126/science.165.3894.664. [DOI] [PubMed] [Google Scholar]
- Gillespie-Lynch K, Greenfield PM, Lyn H, Savage-Rumbaugh S. The Role of Dialogue in the Ontogeny and Phylogeny of Early Word Combinations. in review. [Google Scholar]
- Gómez JC. Pointing Behaviors in Apes and Human Infants: A Balanced Interpretation. Child Development. 2007;78:729–734. doi: 10.1111/j.1467-8624.2007.01027.x. [DOI] [PubMed] [Google Scholar]
- Greenfield PM, Lyn H. Symbol combination in Pan: language, action, and culture. In: Washburn DA, editor. Primate perspectives on behavior and cognition. American Psychological Association; Washington, DC, US: 2007. pp. 255–267. [Google Scholar]
- Greenfield PM, Lyn H, Savage-Rumbaugh ES. Protolanguage in ontogeny and phylogeny: combining deixis and representation. Interaction Studies. 2008;9:34–50. [Google Scholar]
- Greenfield PM, Savage-Rumbaugh ES. Grammatical combination in Pan paniscus: processes of learning and invention in the evolution and development of language. In: Parker ST, Gibson KR, editors. “Language” and intelligence in monkeys and apes: Comparative developmental perspectives. Cambridge University Press; New York, NY, US: 1990. pp. 540–578. [Google Scholar]
- Greenfield PM, Savage-Rumbaugh ES. Imitation, grammatical development, and the invention of protogrammar by an ape. In: Krasnegor NA, Rumbaugh DM, Schiefelbusch RL, Studdert-Kennedy M, editors. Biological and behavioral determinants of language development. Lawrence Erlbaum Associates, Inc; Hillsdale, NJ, US: 1991. pp. 235–258. [Google Scholar]
- Greenfield PM, Savage-Rumbaugh ES. Comparing communicative competence in child and chimp: the pragmatics of repetition. Journal of Child Language. 1993;20:1–26. doi: 10.1017/s0305000900009090. [DOI] [PubMed] [Google Scholar]
- Greenfield PM, Smith JH. The structure of communication in early language development. Academic Press; New York: 1976. [Google Scholar]
- Hare B, Tomasello M. Chimpanzees are more skilful in competitive than in cooperative cognitive tasks. Anim Behav. 2004;68:571–581. [Google Scholar]
- Hayes C. The ape in our house. 1. Gollancz; London: 1951. [Google Scholar]
- Hoerl C. On being stuck in time. Phenomenology and the Cognitive Sciences. 2008;7:485–500. [Google Scholar]
- Liszkowski U, Carpenter M, Henning A, Striano T, Tomasello M. 12-month-olds point to share attention and interest. Developmental Science. 2004;7:297–307. doi: 10.1111/j.1467-7687.2004.00349.x. [DOI] [PubMed] [Google Scholar]
- Lyn H. Mental representation of symbols as revealed by vocabulary errors in two bonobos (Panpaniscus) Anim Cogn. 2007;10:461–475. doi: 10.1007/s10071-007-0086-3. [DOI] [PubMed] [Google Scholar]
- Lyn H. Environment, methodology, and the object choice task in apes: evidence for declarative comprehension and implications for the evolution of language. Journal of Evolutionary Psychology in press. [Google Scholar]
- Lyn H, Greenfield PM, Savage-Rumbaugh ES. The development of representational play in chimpanzees and bonobos: Evolutionary implications, pretense, and the role of interspecies communication. Cognitive Development. 2006;21:199–213. [Google Scholar]
- Lyn H, Greenfield PM, Savage-Rumbaugh ES. Semiotic combinations in Pan: A cross-species comparison of communication in a chimpanzee and a bonobo. First Language in press. [Google Scholar]
- Lyn H, Russell JL, Hopkins WD. The Impact of Environment on the Comprehension of Declarative Communication in Apes. Psychological Science. 2010;21:360–365. doi: 10.1177/0956797610362218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyn H, Savage-Rumbaugh ES. Observational word learning by two bonobos: ostensive and non-ostensive contexts. Language and Communication. 2000;20:255–273. [Google Scholar]
- Menzel CR. Unprompted recall and reporting of hidden objects by a chimpanzee (Pan troglodytes) after extended delays. Journal of Comparative Psychology. 1999;113:426–434. doi: 10.1037/0735-7036.113.4.426. [DOI] [PubMed] [Google Scholar]
- Miklosi A, Soproni K. A comparative analysis of animals’ understanding of the human pointing gesture. Anim Cogn. 2006;9:81–93. doi: 10.1007/s10071-005-0008-1. [DOI] [PubMed] [Google Scholar]
- Moll H, Tomasello M. Cooperation and human cognition: the Vygotskian intelligence hypothesis. Philosophical Transactions of the Royal Society B. 2007;362:639–648. doi: 10.1098/rstb.2006.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulcahy NJ, Call J. Apes save tool for future use. Science. 2006;213:1038–1040. doi: 10.1126/science.1125456. [DOI] [PubMed] [Google Scholar]
- Osvath M, Osvath H. Chimpanzee (Pan troglodytes) and orangutan (Pongo abelii) forethought: Self-control and pre-experience in the face of future tool use. Animal Cognition. 2008;11:661–674. doi: 10.1007/s10071-008-0157-0. [DOI] [PubMed] [Google Scholar]
- Patterson FG. The gestures of a gorilla: language acquisition in another pongid. Brain and Language. 1978;5:72–97. doi: 10.1016/0093-934x(78)90008-1. [DOI] [PubMed] [Google Scholar]
- Pika S. Gestures of apes and pre-linguistic human children: Similar or different? First Language. 2008;28:116–140. [Google Scholar]
- Premack D, Premack AJ. The mind of an ape. 1. Norton; New York: 1983. [Google Scholar]
- Rivas E. Recent Use of Signs by Chimpanzees (Pan Troglodytes) in Interactions With Humans. J Comp Psychol. 2005;119:404–417. doi: 10.1037/0735-7036.119.4.404. [DOI] [PubMed] [Google Scholar]
- Savage-Rumbaugh ES. Ape language: from conditioned response to symbol. Columbia University Press; New York, NY, US: 1986. [Google Scholar]
- Savage-Rumbaugh ES, McDonald K, Sevcik RA, Hopkins WD, Rupert E. Spontaneous symbol acquisition and communicative use by pygmy chimpanzees (Pan paniscus) Journal of Experimental Psychology: General. 1986;115:211–235. doi: 10.1037//0096-3445.115.3.211. [DOI] [PubMed] [Google Scholar]
- Savage-Rumbaugh ES, Murphy J, Sevcik RA, Brakke KE, Williams SL, Rumbaugh DM. Language comprehension in ape and child. Monographs of the Society for Research in Child Development. 1993;58:v–221. [PubMed] [Google Scholar]
- Savage-Rumbaugh ES, Rumbaugh DM, Smith ST, Lawson J. Reference: the linguistic essential. Science. 1980;210:922–925. doi: 10.1126/science.7434008. [DOI] [PubMed] [Google Scholar]
- Schwartz BL, Hoffman ML, Evans S. Episodic-like memory in a gorilla: A review and new findings. Learning and Motivation. 2005;36:226–244. [Google Scholar]
- Suddendorf T, Busby J. Mental time travel in animals? Trends in Cognitive Sciences. 2003;7:391–396. doi: 10.1016/s1364-6613(03)00187-6. [DOI] [PubMed] [Google Scholar]
- Suddendorf T, Corballis MC. Mental time travel and the evolution of the human mind. Genet Soc Gen Psychol Monogr. 1997;123:133–167. [PubMed] [Google Scholar]
- Suddendorf T, Corballis MC. The evolution of foresight: What is mental time travel, and is it unique to humans? Behavioral and Brain Sciences. 2007a;30:299–313. doi: 10.1017/S0140525X07001975. [DOI] [PubMed] [Google Scholar]
- Suddendorf T, Corballis MC. Mental time travel across the disciplines: The future looks bright. Behavioral and Brain Sciences. 2007b;30:335–351. doi: 10.1017/S0140525X07001975. [DOI] [PubMed] [Google Scholar]
- Terrace HS, Petitto LA, Sanders RJ, Bever TG. Can an ape create a sentence? Science. 1979;206:891–902. doi: 10.1126/science.504995. [DOI] [PubMed] [Google Scholar]
- Tomasello M. If They’re So Good at Grammar, Then Why Don’t They Talk? Hints From Apes’ and Humans’ Use of Gestures. Language Learning and Development. 2007;3:133–156. [Google Scholar]
- Tomasello M, Camaioni L. A comparison of the gestural communication of apes and human infants. Human Development. 1997;40:7–24. [Google Scholar]
- Tomasello M, Carpenter M. The Emergence of Social Cognition in Three Young Chimpanzees. Monographs of the Society for Research in Child Development. 2005;70:1–136. doi: 10.1111/j.1540-5834.2005.00324.x. [DOI] [PubMed] [Google Scholar]
- Veá JJ, Sabater-Pi J. Spontaneous pointing behaviour in the wild pygmy chimpanzee (Pan paniscus) Folia Primatologica. 1998;69:289–290. doi: 10.1159/000021640. [DOI] [PubMed] [Google Scholar]