

Abstract
BACKGROUND AND PURPOSE
Prostaglandin (PG) D2 has emerged as a key mediator of allergic inflammatory pathologies and, particularly, PGD2 induces leukotriene (LT) C4 secretion from eosinophils. Here, we have characterized how PGD2 signals to induce LTC4 synthesis in eosinophils.
EXPERIMENTAL APPROACH
Antagonists and agonists of DP1 and DP2 prostanoid receptors were used in a model of PGD2-induced eosinophilic inflammation in vivo and with PGD2-stimulated human eosinophils in vitro, to identify PGD2 receptor(s) mediating LTC4 secretion. The signalling pathways involved were also investigated.
KEY RESULTS
In vivo and in vitro assays with receptor antagonists showed that PGD2-triggered cysteinyl-LT (cysLT) secretion depends on the activation of both DP1 and DP2 receptors. DP1 and DP2 receptor agonists elicited cysLTs production only after simultaneous activation of both receptors. In eosinophils, LTC4 synthesis, but not LTC4 transport/export, was activated by PGD2 receptor stimulation, and lipid bodies (lipid droplets) were the intracellular compartments of DP1/DP2 receptor-driven LTC4 synthesis. Although not sufficient to trigger LTC4 synthesis by itself, DP1 receptor activation, signalling through protein kinase A, did activate the biogenesis of eosinophil lipid bodies, a process crucial for PGD2-induced LTC4 synthesis. Similarly, concurrent DP2 receptor activation used Pertussis toxin-sensitive and calcium-dependent signalling pathways to achieve effective PGD2-induced LTC4 synthesis.
CONCLUSIONS AND IMPLICATIONS
Based on pivotal roles of cysLTs in allergic inflammatory pathogenesis and the collaborative interaction between PGD2 receptors described here, our data suggest that both DP1 and DP2 receptor antagonists might be attractive candidates for anti-allergic therapies.
LINKED ARTICLE
This article is commented on by Mackay and Stewart, pp. 1671–1673 of this issue. To view this commentary visit http://dx.doi.org/10.1111/j.1476-5381.2011.01236.x
Keywords: eosinophils, PGD2, LTC4, DP1, DP2, CRTh2, lipid droplets, lipid bodies, allergic inflammation, asthma
Introduction
Eosinophil accumulation and subsequent activation at sites of allergic inflammation control the generation and release of diverse lipid and protein mediators critical to the development and perpetuation of allergic processes (Busse and Lemanske, 2001; Rothenberg and Hogan, 2006). Indeed, eosinophils represent a major source of leukotriene (LT) C4, the intracellular parent of the cysteinyl LTs (cysLTs; LTC4/D4/E4) (Weller et al., 1983; Cowburn et al., 1998; Bandeira-Melo and Weller, 2003). Central to the pathogenesis of allergic diseases, cysLTs cause bronchoconstriction, mucus hypersecretion, increased microvascular permeability, bronchial hyperresponsiveness, eosinophil infiltration and airway remodelling (Drazen and Austen, 1987). LTC4, as the main eosinophil-derived product of the 5-lipoxygenase pathway of arachidonic acid metabolism in allergic diseases, is formed by the conjugation of LTA4 with reduced glutathione by LTC4 synthase. After active transport to the extracellular space, LTC4 is converted to LTD4 and LTE4 through sequential enzymatic removal of glutamic acid and glycine. Even though the eosinophil LTC4-synthesizing enzymatic pathways are well known (Bandeira-Melo et al., 2002a), it remains of particular interest to fully characterize endogenous allergy-relevant stimuli, receptors and signalling pathways, as well as to understand the intracellular compartmentalization mechanisms that control allergen-induced, eosinophil-driven, LTC4 synthesis.
Prostaglandin D2 (PG) D2 is another key lipid mediator of allergic airway inflammation that is produced following allergen exposure in patients with asthma, atopic dermatitis or allergic rhinitis. Similar to cysLTs, PGD2 can mimic a number of important features of allergic processes (Pettipher, 2008). In regard to the sensitization phase of an allergic response, it has been postulated that PGD2 modulates cytokine production by dendritic cells, leading to the polarization of T helper type 2 cell (Th2) cells (Theiner et al., 2006; Hammad et al., 2007). In addition to its immuno-regulatory role, PGD2 is a highly effective trigger of blood flow changes, promoting oedema formation and therefore contributing to the nasal congestion symptom of allergic rhinitis (Doyle et al., 1990; Widdicombe, 1990). PGD2 also mediates the typical cell accumulation of late phase allergic responses by functioning as a selective chemoattractant for Th2 cells, basophils and eosinophils (Hirai et al., 2001; Monneret et al., 2001).
The immuno-modulatory and inflammatory functions of PGD2 are mediated by high-affinity interaction with two receptors, the prostanoid DP1 receptor and the chemoattractant receptor-homologous molecule expressed on Th2 cells receptor, now referred to as the DP2 receptor (receptor nomenclature follows Alexander et al., 2009). Although DP1 and DP2 receptors bind the same ligand, there is very little homology between the two receptors; DP1 receptors being members of the prostanoid receptor family that includes EP1-4, FP, IP and TP, whereas the DP2 receptors are more closely related to other chemotactic receptors such as the LTB4 receptors, BLT1 and BLT2, and the C5a receptor (Pettipher, 2008). Furthermore, while DP1 receptors are coupled to Gαs proteins and signal through elevation of intracellular levels of cAMP, the DP2 receptors are coupled to Gαi and their activation leads to the elevation of intracellular calcium, reduction in cAMP (Sawyer et al., 2002) and downstream activation of phosphatidylinositol-3-kinase (PI3K) (Xue et al., 2007).
Eosinophils co-express the classical DP1 receptors coupled to adenylyl cyclase, as well as the Pertussis toxin (PTX)-sensitive DP2 receptors (Monneret et al., 2001). Although the ability of PGD2 to activate eosinophils while concurrently elevating cAMP levels seems paradoxical, DP1 receptors appear to be able to co-mediate, with DP2 receptors, the mobilization of eosinophils from bone marrow as well as chemotaxis (Schratl et al., 2007). Alternatively, it has also been shown that the final chemotaxis-related response of eosinophils to PGD2 may be, ultimately, determined by a balance between two opposing downstream signalling pathways: the cAMP-dependent, inhibitory pathway activated via DP1 receptors and the prevailing, stimulatory pathway activated via DP2 receptors (Monneret et al., 2001; Kostenis and Ulven, 2006; Sandig et al., 2007).
Recently, we have shown that, in addition to its chemotactic activity towards eosinophils, PGD2 controls allergy-relevant eosinophil activation, particularly the increased LTC4 synthesizing capacity of these cells (Mesquita-Santos et al., 2006). Indeed, other eosinophil chemoattractants, including eotaxin (CCL11), RANTES (CCL5) and platelet activating factor (PAF) are capable of triggering LTC4 synthesis within eosinophils through the activation of their cognate Gαi -coupled chemotactic receptors, such as CCR3 (Bozza et al., 1996; Bandeira-Melo et al., 2001). Therefore, we had initially hypothesized that PGD2-induced LTC4 synthesis could be mediated by the stimulatory activation of DP2 receptors while being counter-balanced by a parallel inhibitory cAMP-dependent DP1 receptor activation. However, here by employing pharmacological strategies, we have uncovered a novel kind of interaction between the PGD2 receptor types expressed on eosinophils. In contrast to the PGD2-driven opposing signalling related to eosinophil chemotactic activities, eosinophil LTC4 synthesis triggered by PGD2 appeared to be controlled by the complementary stimulatory events between DP1 receptor-activated, PKA-driven, lipid bodies and concurrent DP2 receptor signalling. While PGD2 emerges as a potent inflammatory mediator of allergic disorders and as an interesting therapeutic target, because of the mandatory dual activation of DP1 and DP2 receptors for increasing eosinophil LTC4 synthesis, either DP1 or DP2 receptor antagonists might be highly effective candidates as anti-allergic tools to control cysLTs production regulated by the activation of eosinophils at sites of allergic reactions.
Methods
Animals
All animal care and experimental protocols were approved by Oswaldo Cruz Foundation Animal Welfare Committee. For in vivo experiments, male Swiss mice of 16–20 g were obtained from Oswaldo Cruz Foundation breeding unit (Rio de Janeiro, Brazil).
PGD2-induced and allergic pleurisy in sensitized mice
As previously described (Mesquita-Santos et al., 2006), mice were sensitized with an s.c. injection (0.2 mL) of ovalbumin (OVA; 50 µg; Sigma, St. Louis, MO, USA) and Al(OH)3 (5 mg) in 0.9% NaCl solution (saline) at days 1 and 7. Allergic challenge was performed at day 14 by means of an intrapleural injection of OVA (12 µg per cavity; 0.1 mL). Alternatively, sensitized mice were challenged with PGD2 (35 pmol per cavity), BW245C (35 pmol per cavity), DK-PGD2 (35 pmol per cavity) or a combination of the latter two agonists (both at 35 pmol per cavity) (all from Cayman Chemicals, Ann Arbor, MI, USA). All stimuli were diluted in sterile saline immediately before use. Control animals were injected with the same volume (0.1 mL) of vehicle. Mice were killed by CO2 inhalation 24 h after challenge. Pleural fluid was obtained by rinsing cavities with 1 mL of phosphate-buffered saline containing BSA (0.1%). After samples were taken for lipid body analysis, pleural fluid was centrifuged and cell free supernatants were used for the quantification of cysLTs.
Isolation and in vitro stimulation of human eosinophils
Peripheral blood was obtained with informed consent from healthy donors under protocols approved by the ethical review boards of both the Federal University of Rio de Janeiro and the Oswaldo Cruz Foundation (Rio de Janeiro, Brazil). Eosinophils were isolated by negative selection using EasySep™ system (StemCell Technologies Inc., Vancouver, Canada) (purity ∼98%; viability ∼95%) (Bezerra-Santos et al., 2006). Purified eosinophils (2 × 106 cells·mL−1) in Hank's balanced salt solution without Ca2+ and Mg2+ (HBSS−/−) were incubated for 1 h in a water bath (37°C) with PGD2 (25 nM), BW245C (5–625 nM), DK-PGD2 (5–625 nM), a combination of BW245C and DK-PGD2 (both at 25 nM) or PAF (10 µM; from Cayman Chemicals). After samples were taken for lipid body analyses, eosinophils were resuspended in Hank's balanced salt solution with Ca2+ and Mg2+ (HBSS+/+) and stimulated with 0.1 µM A23187 (Sigma) for another 15 min (37°C). Cell-free supernatants were then collected for cysLT quantification. Each in vitro experiment was repeated at least three times, with eosinophils purified from different donors.
Treatments
Using the pleurisy models, animals were pretreated with i.p. injections of the DP1 receptor antagonist BWA868C (1 mg·kg−1; Cayman Chemicals), the dual DP2/TP receptor antagonist ramatroban (also known as Bay-u3405; 1 mg·kg−1; Cayman Chemicals) or the selective DP2 receptor antagonist Cay10471 (1 mg·kg−1; Cayman Chemicals) 30 min before either PGD2 or allergic challenges.
For in vitro studies, eosinophils in HBSS−/− were pretreated for 30 min with the DP1 receptor antagonist BWA868C (200 nM), the dual DP2/TP receptor antagonist ramatroban (200 nM), the selective DP2 antagonist Cay10471 (200 nM), two PKA inhibitors H-89 and PKI (both at 10 µM; Calbiochem, La Jolla, CA, USA); PTX (1 µg·mL−1; Calbiochem) or cell-permeable calcium chelator BAPTA-AM (25 µg·mL−1; Sigma). Of note, these pretreatments did not modify basal lipid body content found within cytoplasm of non-stimulated eosinophils or affected eosinophil viability (∼90%) (data not shown).
Quantification of cysLTs
The amount of cysLTs found in cell-free pleural fluid and eosinophil supernatants was measured by the Cysteinyl Leukotriene EIA kit (catalog number 520501; from Cayman Chemicals), according to the manufacturer's instructions.
EicosaCell for intracellular LTC4 immuno-detection
As previously described (Bandeira-Melo et al., 2001) to localize LTC4 at its sites of synthesis, in vitro stimulated eosinophils were mixed with a solution of 1-ethyl-3-(3-dimethylamino-propyl) carbodiimide (EDAC; 0.1% final concentration with cells in HBSS−/−), used to crosslink eicosanoid carboxyl groups to amines in adjacent proteins. After 15 min incubation at room temperature with EDAC to promote both cell fixation and permeabilization, eosinophils were then washed with HBSS−/−, cytospun onto glass slides and blocked with HBSS−/− containing 1% BSA for 30 min. Cells were incubated with rabbit anti-LTC4 antibodies (Cayman Chemicals) or non-immune rabbit IgG (Jackson ImmunoResearch Inc., West Grove, PA, USA) overnight. Routinely, cells were co-incubated with guinea pig anti-adipose differentiation-related protein (ADRP) antibody (1:300 dilution; Fitzgerald Industries, North Acton, MA, USA) to distinguish cytoplasmic lipid bodies within eosinophils. The cells were washed 3× from 10 min with HBSS−/− containing 1% BSA and incubated with Alexa488-labelled anti-rabbit IgG and Alexa546-labelled anti-guinea pig secondary antibodies for 1 h.
The specificity of the LTC4 immuno-labelling was confirmed by the: (i) lack of immunofluorescence within PGD2-stimulated human eosinophils incubated with irrelevant IgG (data not shown); and (ii) lack of LTC4 immuno-labelling within non-stimulated human eosinophils that were incubated with anti-LTC4 antibody.
Images were obtained using an Olympus BX51 fluorescence microscope at 100× magnification and photographs were taken with the Olympus 72 digital camera (Olympus Optical Co., Tokyo, Japan) in conjunction with CellF Imaging Software for Life Science Microscopy (Olympus Life Science Europa GMBH, Hamburg, Germany). The images were edited using Adobe Photoshop 5.5 software (Adobe Systems, San Jose, CA, USA).
Lipid body staining and enumeration
To enumerate lipid bodies, cells recovered from pleural cavities or human eosinophils were cytocentrifuged (450 rpm, 5 min) onto glass slides. Cells, while still moist, were fixed in 3.7% formaldehyde (in HBSS−/−; pH 7.4), rinsed in 0.1 M cacodylate buffer (pH 7.4), stained with 1.5% OsO4 (Sigma) for 30 min, rinsed in distilled H2O, immersed in 1.0% thiocarbohydrazide for 5 min, rinsed in 0.1 M cacodylate buffer, restained with 1.5% OsO4 for 3 min, rinsed in distilled water, and then air dried and analysed. Cell morphology was observed and lipid bodies were enumerated by light microscopy. Fifty consecutively scanned eosinophils were evaluated by more than one individual and results were expressed as the number of lipid bodies per eosinophil.
Statistical analysis
Results are expressed as mean ± SEM and were analysed statistically by means of analysis of variance followed by the Newman–Keuls Student test, with the level of significance set at P < 0.05.
Results
Both DP1 and DP2 receptors contribute to PGD2-induced enhanced LTC4 secretion from eosinophils
We have reported that PGD2, formerly recognized only as an eosinophil chemoattractant, is also able to trigger eosinophil activation, characterized by enhanced LTC4 synthesis (Mesquita-Santos et al., 2006). Here, by employing a mouse model of PGD2-induced eosinophil activation in vivo (Mesquita-Santos et al., 2006), as well as human-purified eosinophils stimulated in vitro with PGD2, we investigated the molecular and cellular mechanisms involved in PGD2-induced LTC4 synthesis within eosinophils. Our initial goal was to identify the specific PGD2 receptor, either DP1 or DP2, the two well recognized, cloned and eosinophil-expressed receptors for PGD2 that was involved in this response. In vivo, as illustrated in Figure 1, both PGD2 receptors appeared to contribute to the LTC4 secretion from eosinophils, inasmuch as pretreatment with either the DP1 receptor antagonist BWA868C or the dual DP2/TP receptor antagonist ramatroban displayed high levels of inhibition. BWA868C and ramatroban significantly reduced LTC4 secretion levels found increased in sites of PGD2-triggered eosinophilic inflammation by 63 and 81%, respectively (Figure 1A). Similarly, Figure 1B shows that increased amounts of LTC4 detected in the supernatants of in vitro PGD2-stimulated human eosinophils were also inhibited by either the DP1 receptor antagonist BWA868C, the dual DP2/TP receptor antagonist ramatroban or by the selective DP2 antagonist Cay10471 (83, 96 and 89% inhibition, respectively), indicating essential roles for each receptor type, which in turn, perhaps by acting synergistically, evoke PGD2-driven secretion of LTC4 from eosinophils. Of note, by showing that (i) pretreatments with BWA868C and Cay10471 did not interfere with PAF-induced LTC4 synthesis by human eosinophils (Table 1), or (ii) the pretreatment with the selective TP antagonist SQ29548 failed to alter LTC4 synthesis triggered by in vitro stimulation of eosinophils (Figures 1B; 10% inhibition), respectively, we excluded potential non-specific effects of PGD2 receptor antagonists and demonstrated that the inhibitory effects of ramatroban were dependent on DP2, rather than TP receptor antagonism.
Figure 1.
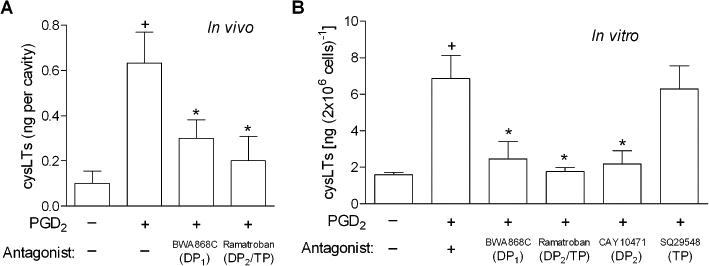
Both DP1 and DP2 receptors control cysLTs production triggered by PGD2. In A, sensitized mice were pretreated with BWA868C (1 mg·kg−1) or ramatroban (1 mg·kg−1) and then stimulated with an i.pl. injection of PGD2 (35 pmol per cavity). Analysis of cysLTs synthesis was performed 24 h after PGD2 administration. Results are expressed as the means ± SEM from at least six animals. †P≤ 0.05 compared with control animals and *P≤ 0.05 compared with PGD2-injected mice. In B, for in vitro analysis of LTC4 synthesis, human eosinophils were pretreated for 30 min with BWA868C (200 nM), ramatroban (200 nM), Cay10471 (200 nM) or SQ29548 (200 nM), and then stimulated for 1 h with PGD2 (25 nM). In vitro results are expressed as the means ± SEM from at least three independent experiments with eosinophils purified from different donors. †P≤ 0.05 compared with control. *P≤ 0.05 compared with PGD2-stimulated eosinophils. cysLT, cysteinyl leukotriene; DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; i.pl., intrapleural; PGD2, prostaglandin D2.
Table 1.
PAF-induced lipid body biogenesis and LTC4 synthesis by human eosinophils are dependent on PTX-sensitive protein Gαi and on intracellular calcium mobilization, but not on activation of DP2 receptors or PKA activity
| Conditions | Treatments | Lipid bodies/eosinophil | cysLTs (ng/2 × 106 cells) |
|---|---|---|---|
| Control | 7.7 ± 0.4 | 1.1 ± 0.6 | |
| PAF | 16.4 ± 1.4† | 4.4 ± 1.4† | |
| +BWA868C | 16.9 ± 1.7 | 4.2 ± 1.3 | |
| +CAY10471 | 16.7 ± 1.6 | 4.5 ± 1.2 | |
| +H-89 | 13.6 ± 2.3 | 4.5 ± 1.6 | |
| +PTX | 10.5 ± 1.4* | 1.6 ± 0.8* | |
| +BAPTA-AM | 8.6 ± 0.5* | 1.5 ± 0.8* |
P≤ 0.05 compared with control group.
P≤ 0.05 compared with PAF-stimulated eosinophils.
Human eosinophils were pretreated for 30 min with BWA868C (200 nM), CAY10471 (200 nM), H-89 (10 µM), Pertussis toxin (PTX; 1 µg·mL−1) or BAPTA-AM (25 µM), and stimulated with PAF (10 µM). Analysis of lipid body biogenesis and LTC4 synthesis were performed 1 h after PAF stimulation. Results were expressed as the means ± SEM from at least three different experiments.
cysLT, cysteinyl leukotriene; DP2, D prostanoid receptor 2; PAF, platelet activating factor.
DP1 and DP2 receptors cooperate to trigger PGD2-driven enhanced LTC4 secretion from eosinophils
To test the hypothesis that a synergistic mechanism of action between the PGD2 receptors, DP1 and DP2, controls PGD2-driven induction of LTC4 secretion from eosinophils, selective agonists of DP1 receptors (BW245C) or of DP2 receptors (DK-PGD2) were applied to in vivo and in vitro systems of eosinophil activation. Figure 2 shows that, alone, neither BW245C nor DK-PGD2 was able to elicit LTC4 release from eosinophils at concentrations that PGD2 by itself is able to trigger LTC4 secretion both in vivo (35 pmol per cavity; Figure 2A) and in vitro (25 nM; Figure 2B). In contrast, by co-stimulating eosinophils with a mixture of DP1 and DP2 agonists, increased cysLTs levels were detected at eosinophilic inflammatory sites (Figure 2A) and human eosinophil-free supernatants (Figure 2B), with comparable magnitude to those found in vivo (Figure 2A) or in vitro (Figure 2B) stimulation with PGD2, reinforcing the possibility of a synergism between DP1 and DP2 receptors.
Figure 2.
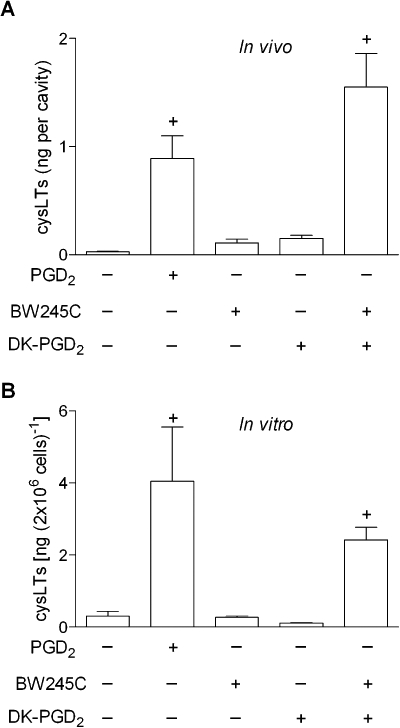
DP1 and DP2 receptors cooperate to trigger cysLTs production. In A, sensitized mice received an i.pl. injection of PGD2 (35 pmol per cavity), BW245C (35 pmol per cavity), DK-PGD2 (35 pmol per cavity) or BW245C plus DK-PGD2 (both at 35 pmol per cavity). Analysis of cysLTs production within pleural fluids was performed 24 h after i.pl. administration. Results are expressed as the means ± SEM from at least six animals. †P≤ 0.05 compared with control animals. In B, for in vitro analyses of LTC4 production in cell-free supernatants, human eosinophils were stimulated for 1 h with PGD2 (25 nM), BW245C (25 nM), DK-PGD2 (25 nM) or with a combination of BW245C plus DK-PGD2 (both at 25 nM). In vitro results are expressed as the means ± SEM from at least three independent experiments with eosinophils purified from different donors. †P≤ 0.05 compared with control. P≤ 0.05 compared with PGD2-stimulated eosinophils. cysLT, cysteinyl leukotriene; DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; i.pl., intrapleural; PGD2, prostaglandin D2.
Cooperation between DP1 and DP2 receptors controls LTC4 synthesis within eosinophil cytoplasmic lipid bodies
Stimulus-dependent extracellular detection of cysLTs depends on sequential intracellular events, including: (i) an initial step of activation and proper compartmentalization of both substrate and enzymatic machinery, culminating in intracellular LTC4 synthesis; (ii) intracellular transport to direct newly synthesized LTC4 to secretory pathways; that is followed by (iii) an active carrier-dependent LTC4 release through the plasma membrane (see Bandeira-Melo et al., 2002a). Therefore, as virtually no cysLTs can be detected in cell-free supernatants of BW245C- or DK-PGD2-stimulated eosinophils, one can argue that, under these stimulatory conditions, the intracellular step of LTC4 synthesis actually occurs, without the subsequent LTC4 transport/release events, which would only follow after the co-stimulation of both PGD2 receptors. By employing EicosaCell technology, a microscopic method that immobilizes and immuno-detects newly synthesized lipid mediators at their sites of synthesis, we have excluded this hypothesis by showing that LTC4 synthesis itself is elicited in vitro within human eosinophils only if both DP1 and DP2 receptors are simultaneously engaged by either PGD2 itself (Figure 3; top panel) or by the combination of both PGD2 receptor agonists BW245C and DK-PGD2 (Figure 3; botton panel). Eosinophils activated only with DP1 or DP2 agonists show no immuno-fluorescent LTC4 (Figure 3; middle panels) in EicosaCell preparations, indicating that the LTC4 synthesizing machinery was indeed not activated under these conditions of stimulation of a single PGD2 receptor.
Figure 3.
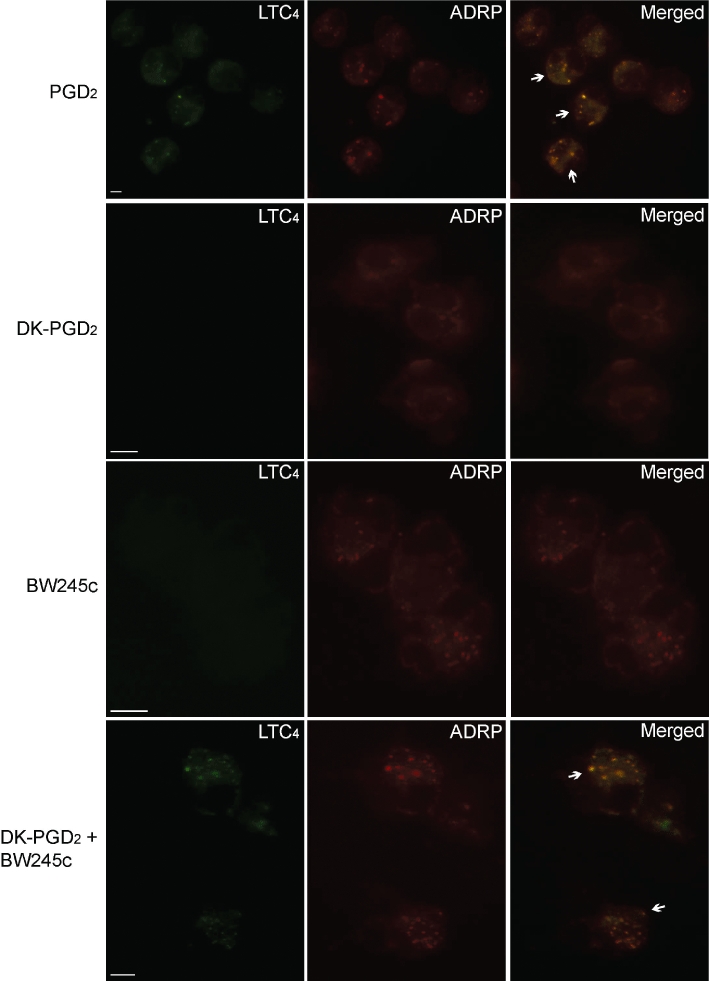
LTC4 synthesis is triggered within eosinophil cytoplasmic lipid bodies by simultaneous activation of DP1 and DP2 receptors by either PGD2 or the combination of BW245C and DK-PGD2 stimulation of human eosinophils in vitro. EicosaCell images illustrate intracellular immuno-detection of newly formed LTC4 (green) and of ADRP (red) in PGD2-stimulated, BW245-stimulated, DK-PGD2-stimulated or BW245C/DK-PGD2 co-stimulated human eosinophils (as indicated). Overlay images of identical fields are shown in the right column. Arrows indicate co-localization of immunolabelled synthesized LTC4 with ADRP-bearing lipid bodies. For EicosaCell analyses, cells were fixed and permeabilized with EDAC and sequentially incubated with anti-LTC4 and anti-ADRP antibodies and Alexa488-labelled anti-rabbit IgG plus Alexa546-labelled anti-guinea pig secondary antibodies. Images are representative of three independent experiments. ADRP, adipose-differentiation-related protein; DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; PGD2, prostaglandin D2.
A more detailed analysis of these preparations revealed that the intracellular LTC4-synthesizing compartment within either PGD2- or BW245C/DK-PGD2-stimulated eosinophils was in a punctate cytoplasmic pattern, proximate to, but separate from the nucleus, and fully consistent in size and form with eosinophil lipid bodies. In fact, the compartmentalization of newly formed LTC4 to eosinophil lipid bodies was confirmed by their co-localization with ADRP (Figure 3; top and bottom panels), a lipid body marker protein. Virtually no LTC4 was immuno-localized within non-stimulated eosinophils (not shown), thus showing that the newly formed lipid bodies of in vitro DP1/DP2-engaged eosinophils are the enzymatically fully equipped organelles responsible for the effective LTC4 synthesis. Of note, eosinophils stimulated solely with BW245C, but not with DK-PGD2, display a lipid body-enriched cytoplasm as detected by the punctate cytoplasmic ADRP immuno-labelling (Figure 3; middle panel as indicated), suggesting that the initially hypothesized synergistic effect at receptor level between DP1 and DP2 does not take place. Instead, PGD2-induced LTC4 synthesis by eosinophils appears to be due to distinct mechanistic roles of DP1 and DP2 receptors with complementary functional features.
DP1 activation, but not that of DP2, evokes biogenesis of eosinophil lipid bodies
To study whether the induction of lipid body assembly driven by DP1-receptor pathways contributed to PGD2-induced LTC4 synthesis, we employed an osmium-based staining methodology that allows the enumeration of these organelles to check for lipid body biogenesis, under either single or combined receptor stimulation. In agreement with EicosaCell images shown previously (Figure 3), Figure 4 shows that selective activation of DP1 receptors by BW245C triggers rapid (within 1 h) assembly of new lipid bodies within human eosinophils in a dose-dependent manner. DP2 receptor activation by DK-PGD2, on the other hand, even when used in high concentrations (Figure 4B) failed to induce eosinophil lipid body biogenesis. No alteration of DP1 receptor-induced lipid body biogenesis was observed when eosinophils were co-stimulated with both DP1 and DP2 receptor agonists (Figure 4A), suggesting that only the DP1 receptor controls the formation of these LTC4-synthesizing organelles.
Figure 4.
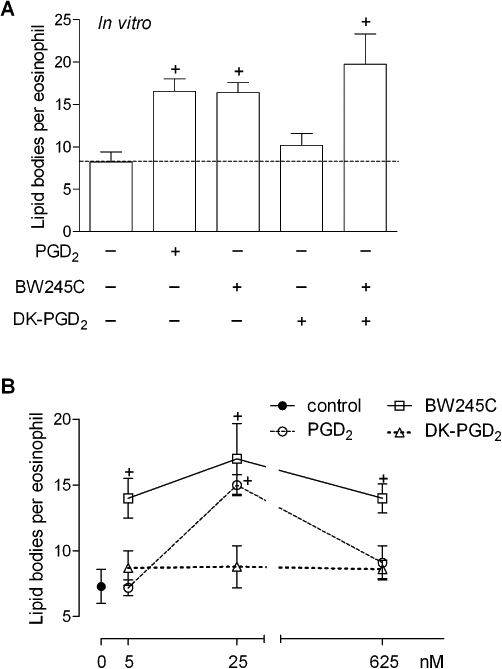
DP1, but not DP2, activation triggers lipid body biogenesis within human eosinophils in vitro. In A, human eosinophils were stimulated with PGD2 (25 nM), BW245C (25 nM), DK-PGD2 (25 nM) or with a combination of BW245C plus DK-PGD2 (both at 25 nM). B shows a dose-response effect of PGD2 (25 nM), BW245C (25 nM) or DK-PGD2 (25 nM) on lipid body biogenesis after stimulation of human eosinophils. Analysis of lipid body biogenesis was performed 1 h after stimulation in osmium-stained cells. Results are expressed as means ± SEM from at least three different experiments with eosinophils purified from distinct donors. †P≤ 0.05 compared with control. DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; PGD2, prostaglandin D2.
To further investigate the role of the DP1 receptor on lipid body biogenesis, we evaluated the participation of each receptor on PGD2-induced lipid body formation (osmium-stained cells) by pretreating PGD2-challenged sensitized mice (Figure 5A) and PGD2-stimulated human eosinophils (Figure 5B) with either DP1 or DP2 receptor antagonists. In vivo, while the pretreatment with the DP2 receptor antagonist ramatroban did not affect the number of cytoplasmic lipid bodies found within infiltrating eosinophils of PGD2-elicited inflammatory reaction, pretreatment with the DP1 antagonist BWA868C nearly abolished this in vivo biogenic process. Similarly, in vitro pre-treatment of human eosinophils with the DP1 receptor antagonist BWA868C significantly inhibited PGD2-induced eosinophil lipid body biogenesis, while two different DP2 receptor antagonists, ramatroban and Cay10471, failed to modify the lipid body assembly triggered by PGD2. Thus, it appears reasonable to postulate that the main role of DP1 receptors in PGD2-induced LTC4 synthesis is to initiate the intracellular signalling pathway that leads to the biogenesis of LTC4 synthesizing organelles in eosinophils.
Figure 5.
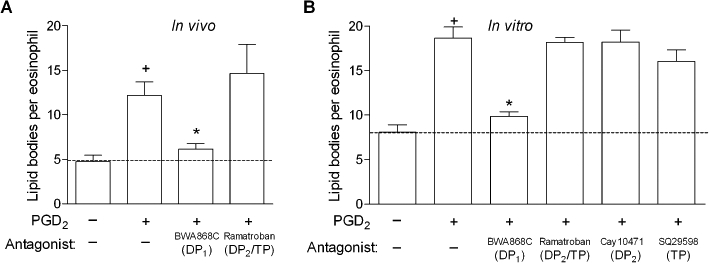
DP1, but not DP2 receptors, control eosinophil lipid body biogenesis triggered by PGD2 either in vivo or in vitro. In A, sensitized mice were pretreated with BW868c (1 mg·kg−1) or ramatroban (1 mg·kg−1), and then stimulated with an i.pl. injection of PGD2 (35 pmol/cavity). Analysis of lipid body biogenesis was performed 24 h after PGD2 administration in osmium-stained cells. Results are expressed as means ± SEM from at least six animals. †P≤ 0.05 compared with control animals and *P≤ 0.05 compared with PGD2-injected mice. In B, for in vitro analysis of lipid body biogenesis, human eosinophils were pretreated for 30 min with BW868c (200 nM), ramatroban (200 nM), Cay10471 (200 nM) or SQ29548 (200 nM), stimulated for 1 h with PGD2 (25 nM) and subsequently stained with osmium. In vitro results are expressed as the means ± SEM from at least three different experiments with eosinophils purified from distinct donors. †P≤ 0.05 compared with control. *P≤ 0.05 compared with PGD2-stimulated eosinophils. DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; i.pl., intrapleural; PGD2, prostaglandin D2.
DP1 receptors signal via PKA activation to trigger eosinophil lipid body biogenesis: a requirement for PGD2-induced LTC4 synthesis
To further evaluate the role of DP1 receptors in PGD2-induced lipid body-driven LTC4 synthesis, we have studied the contribution of DP1 receptor-related cAMP-dependent signalling on PGD2-induced assembly of new lipid bodies within eosinophils. As shown in Figure 6, in vitro lipid body assembly triggered by stimulation with either PGD2 (Figure 6A) or BW245C (Figure 7) was consistent with the central role of DP1 receptors, as pretreatment with H-89 or PKI (10 µM), two non-structurally related inhibitors of PKA activation, decreased the numbers of cytoplasmic lipid bodies found within PGD2- (Figure 6A; bottom panel) and BW245C-stimulated human eosinophils (Figure 7). While the specificity of PKA involvement in DP1 receptor-driven effect was strengthened by the lack of effect of H-89 on PAF-induced eosinophil lipid body biogenesis (Table 1), the ability of forskolin, a well-known activator of adenylate cyclase, to trigger rapid formation of lipid bodies within human eosinophils substantiates the role of cAMP/PKA signalling pathway on the regulation of lipid body biogenic process (C. Bandeira-Melo, unpubl. data).
Figure 6.
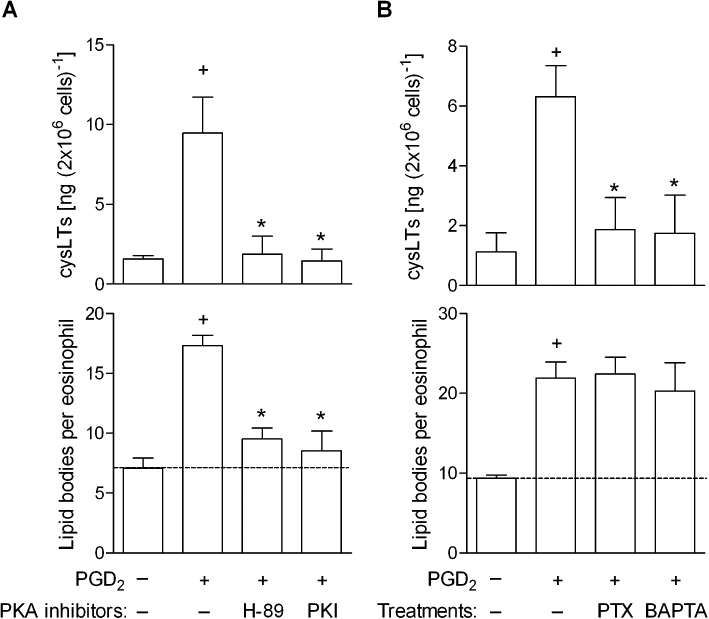
DP1 receptor-driven PKA activation cooperates with DP2-driven Gαi protein activation and calcium influx to mediate lipid body-driven LTC4 sytnhesis within human eosinophils triggered by in vitro PGD2. Human eosinophils were pretreated for 30 min with H-89 (10 µM) and PKI (10 µM) in A, or with PTX (1 µg·mL−1) or BAPTA-AM (25 µg·mL−1) in B and then stimulated with PGD2 (25 nM). In vitro analysis of LTC4 production in cell-free supernatants and lipid body biogenesis were analysed 1 h after PGD2. Results are expressed as the means ± SEM from at least three different experiments with eosinophils purified from different donors. †P≤ 0.05 compared with control group. *P≤ 0.05 compared with PGD2-stimulated eosinophils. DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; PGD2, prostaglandin D2; PTX, Pertussis toxin.
Figure 7.
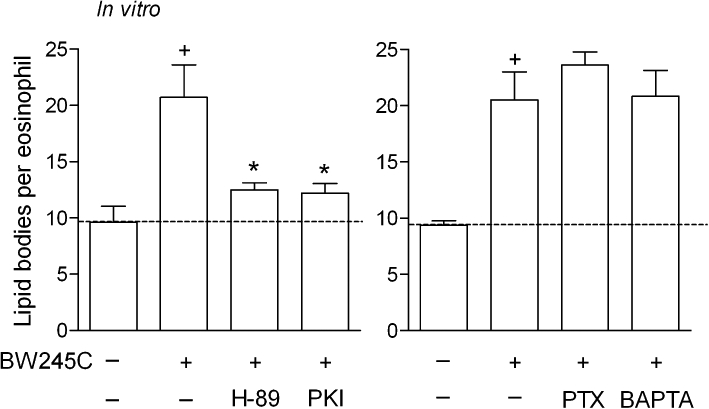
PKA activation, but not Gαi protein and calcium influx, mediates lipid body biogenesis within human eosinophils triggered by BW245C in vitro. Human eosinophils were pretreated for 30 min with PTX (1 µg·mL−1), BAPTA-AM (25 µg·mL−1), H-89 (10 µM) or PKI (10 µM), and then stimulated with BW245C (25 nM). Lipid body biogenesis was analysed 1 h after BW245C stimulation. Results are expressed as the means ± SEM from at least three different experiments with eosinophils purified from different donors. †P≤ 0.05 compared with control group. *P≤ 0.05 compared with PGD2-stimulated eosinophils. PTX, Pertussis toxin.
In agreement with DP1-driven induction of new lipid bodies as a prerequisite to concurrent DP2-elicited signalling required for successful LTC4 synthesis, PKA inhibition by either H-89 or PKI pretreatment also consequently reduced LTC4 synthesis triggered in vitro by the PGD2 stimulation of human eosinophils (Figure 6A; upper panel).
DP2 receptor activation signals via PTX-sensitive Gαi protein and calcium mobilization to prompt DP1 receptor-driven, newly formed lipid bodies to synthesize LTC4
To establish how the concurrent activation of DP2 receptors contributes to PGD2-induced lipid body-driven LTC4 synthesis, we have studied the potential role of Gαi activation and cytoplasmic calcium mobilization elicited by PGD2. Lipid body assembly triggered in vitro by stimulation with either PGD2 (Figure 6B) or BW245C (Figure 7) was not modified by pretreatments with inhibitors of either Gαi activation or cytoplasmic calcium influx, PTX (1 µg·mL−1) and BAPTA-AM (25 µg·mL−1), respectively, ruling out once more the involvement of DP2 receptors in lipid body formation triggered by PGD2. As shown in Table 1, PTX and BAPTA-AM were also able to reduce lipid body biogenesis induced by PAF within human eosinophils.
Although playing no role in PGD2-induced lipid body assembly, the inhibition of Gαi protein and calcium mobilization by, respectively, PTX and BAPTA-AM pretreatment, reduced LTC4 synthesis triggered in vitro by PGD2 stimulation of human eosinophils (Figure 6B; upper panel), indicating that DP2 receptors, by activating calcium-dependent signalling, converted DP1 receptor-induced lipid bodies into enzymatically active organelles capable of LTC4 synthesis.
Eosinophil lipid body-driven LTC4 synthesis elicited in vivo by allergic inflammation is also mediated by a complementary signalling between DP1 and DP2 receptors
To verify whether the cooperative signalling between DP1 and DP2 receptors also operates under allergic inflammatory conditions in vivo, we employed a mouse model of allergic inflammation characterized by eosinophil accumulation and activation. As shown in Figure 8, while the DP1 antagonist BWA868C reduced both eosinophil lipid body biogenesis and increased levels of cysLTs without significantly affecting eosinophil infiltration found in allergic reaction sites, the DP2 receptor antagonists ramatroban and Cay10471 decreased the numbers of recruited eosinophils and inhibited allergic cysLT production but failed to alter eosinophil lipid body biogenesis; thus, yet again, illustrating the mandatory cooperation between the two PGD2 receptors to evoke LTC4 synthesis, as well establishing the relevance of such cooperation to the molecular mechanisms underlying allergy
Figure 8.
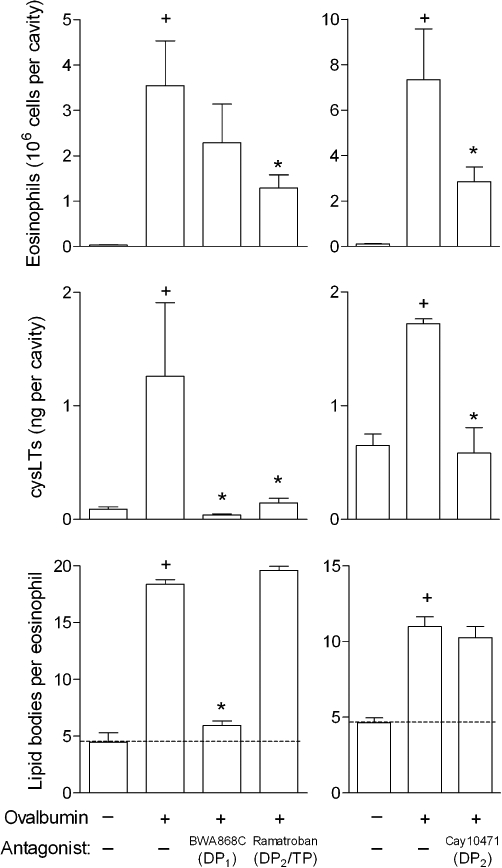
Cooperation between DP1 and DP2 receptors to trigger lipid body-driven LTC4 synthesis within human eosinophils also takes place in allergic inflammatory response in vivo. Sensitized mice were pretreated with BWA868C (1 mg·kg−1), ramatroban (1 mg·kg−1) or Cay10471 (1 mg·kg−1), and then challenged with an i.pl. injection of ovalbumin (12 µg per cavity). Analyses of lipid body biogenesis and cysLTs production were performed 24 h after allergic challenge. Results are expressed as means ± SEM from at least six animals. †P≤ 0.05 compared with saline-challenged mice and *P≤ 0.05 compared with ovalbumin-challenged mice. cysLT, cysteinyl leukotriene; DP1, D prostanoid receptor 1; DP2, D prostanoid receptor 2; i.pl., intrapleural.
Discussion
PGD2 is now emerging as a potential mediator of allergic inflammatory pathologies, because it modulates the polarization of Th2 cells, oedema formation and eosinophil recruitment (Pettipher, 2008). In addition, PGD2 is able to directly activate recruited eosinophils, particularly by eliciting the capacity of eosinophils to synthesize LTC4 at sites of allergic inflammation. While the enzymic pathway by which eosinophils synthesize LTC4 is well characterized, the pathophysiological stimuli and intracellular signalling cascades that govern such activity remain to be fully elucidated. Seeking such characterization, it is important to consider the evolving understanding of the potential roles that some eosinophil chemoattractants, which participate in the recruitment of eosinophils to sites of allergic inflammation, have as priming stimuli on eosinophil LTC4 synthesis.
Our previous studies have demonstrated that, besides PGD2, other eosinophil chemoattractants, such as PAF, CCL11 and CCL5, are secreted in response to allergic challenge and, by acting on their specific receptors expressed on eosinophil membranes, initiate an intracellular cascade leading to enhanced LTC4 synthesis (Bozza et al., 1996; Bandeira-Melo et al., 2001). Our attempts to characterize the intracellular signalling pathways committed to chemoattractant-induced enhanced LTC4 synthesis revealed that diverse stimuli-specific intracellular signalling events control LTC4 synthesis within eosinophils. For instance, while PAF, acting via its PTX-sensitive Gαi-protein-linked receptor appears to induce LTC4 synthesis via a downstream signalling involving PKC and phospholipase C (PLC) activation (Bozza et al., 1996; 1997; 1998;), CCL11 and CCL5, acting via Gαi protein-linked CCR3 receptors, signal via the activation of mitogen-activated proteins kinases and phosphatidylinositide 3-kinase, but not PKC or PLC (Bandeira-Melo et al., 2001). Even though diverse downstream cascades for LTC4 synthesis can be engaged, common upstream steps triggered by the activation of CCR3 and PAF chemotactic receptors share Gαi protein- and calcium influx-regulated cellular activities that, besides culminating in cell polarization/migration, also leads to LTC4 synthesis. Moreover, the well-documented role of PGD2 in eosinophil trafficking as well as a variety of migration-related cellular responses, including actin polymerization and increased expression of adhesion molecules, depends on the activation of Gαi proteins and calcium influx (Monneret et al., 2001; Sawyer et al., 2002). Accordingly, we initially hypothesized that the PGD2 chemotactic DP2 receptor expressed on eosinophils, signalling via the activation of Gαi proteins and calcium influx, would be responsible for PGD2-induced LTC4 synthesis. However, our data showed that specific DP2 receptor stimulation, by itself, was not sufficient to trigger LTC4 synthesis.
Another shared intracellular event triggered by eosinophil chemoattractants that is essential for the successful LTC4 synthesis is the rapid assembly of new cytoplasmic lipid bodies – a biological process recognized as an acute, highly regulated cellular event that is stimulus- and cell-specific (Bozza et al., 2007). As multifunctional organelles, lipid bodies are a hallmark of leukocyte activation and, together with perinuclear envelope (Bandeira-Melo et al., 2001; Tedla et al., 2003) and phagosomes (Balestrieri et al., 2006), represent a potential intracellular domain for LTC4 synthesis. Within eosinophils, compartmentalized LTC4 synthesis triggered by eosinophil chemotactic agents, including PGD2, has been located specifically within lipid bodies, thereby explaining why the biogenesis of lipid bodies critically affects the biosynthesis of LTC4 (Bandeira-Melo et al., 2001; 2002b; Mesquita-Santos et al., 2006). Again, different from the other eosinophil chemotactic receptors, DP2 receptor stimulation alone did not promote PGD2-induced lipid body biogenesis, which was also not dependent on DP2 receptor-related Gαi and calcium signalling. Inasmuch as the lipid body biogenic process is mandatory for LTC4 synthesis, the inability of DP2 receptors to trigger lipid body biogenesis in part explains its inability to promote enhanced LTC4 synthesis within eosinophils.
We found that the discrepancy between DP2 receptors and the other eosinophil chemotactic receptors, in terms of eliciting LTC4 synthesis, relies on the more complex PGD2 receptor system expressed on eosinophils. Besides DP2 receptors, eosinophils also express the non-chemotactic DP1 receptor. By dissecting the specific contributions that each receptor makes to PGD2-induced LTC4 synthesis, we demonstrated that the PGD2-elicited rapid de novo assembly of lipid bodies was insensitive to PTX or a calcium chelator, but was largely dependent on the activation of DP1 receptor-elicited PKA signalling. Of note, the DP1 receptor activation by PGD2 signalling through Gαs proteins leads to increased cAMP and PKA activity – an intracellular signal transducing cascade that is classically related to the inhibition of chemoattractant-induced eosinophil motility (Hirai et al., 2001; Monneret et al., 2001), and consistent with the idea of cAMP elevating agents as powerful anti-inflammatory (Teixeira et al., 1995; Diaz et al., 1996) or pro-resolution (Sousa et al., 2009) agents for the treatment of diseases in which eosinophil accumulation is thought to play a relevant role (Sousa et al., 2009). In this context, it was noticeable that DP1 receptor-driven, PKA-dependent, newly formed lipid bodies were not able to synthesize LTC4, as shown in the EicosaCell preparations. In contrast to PAF- or CCL11-induced LTC4-synthesizing lipid bodies, we demonstrated that under PGD2 stimulation, compartmentalized LTC4 synthesis within DP1 receptor-driven eosinophil lipid bodies, which were assembled under cAMP/PKA regulation, demands concurrent DP2 receptor stimulation, inasmuch as: (i) antagonists for either receptor were equally able to reduce PGD2-induced LTC4 synthesis; (ii) DP1, but not DP2 receptor antagonists, inhibited PGD2-induced lipid body biogenesis; (iii) only by co-stimulating eosinophils with both DP1 and DP2 receptor agonists, was PGD2-induced LTC4 synthesis mimicked; (iv) inhibition of PKA inhibited PGD2-induced DP1 receptor-driven lipid body biogenesis and subsequent lipid body-compartmentalized LTC4 synthesis; and (v) PTX and BAPTA-AM, while failing to interfere with DP1 receptor-dependent lipid body biogenesis, inhibited PGD2-induced lipid body-driven LTC4 synthesis. Therefore, the molecular mechanisms orchestrating how DP2 receptor activation converts DP1 receptor-driven lipid bodies into enzymatically active organelles capable of effective LTC4 synthesis depend on the coordinated Gαi activation and calcium mobilization.
Recently, focusing on eosinophil-driven allergic pathologies, we have found that, alongside CCL11, CCL5, PAF and macrophage migration inhibitory factor (Bandeira-Melo et al., 2001; Vieira-de-Abreu et al., 2005; 2010;), PGD2 is as an endogenous and potent biogenic stimulus of enzymatically active lipid bodies, organelles involved in LTC4 synthesis by eosinophils (Mesquita-Santos et al., 2006). Here, despite the evidence showing that PGD2-driven eosinophil migration-related activities are mediated by a balance of opposing intracellular signalling cascades downstream of DP1 and DP2 receptor activation within eosinophils (Monneret et al., 2001), we uncovered that the intracellular mechanisms of receptor-mediated PGD2-induced LTC4 synthesis rely on the collaborative signalling between both PGD2 receptors. The PGD2-elicited LTC4 synthesis is dependent on the activation of DP1 receptor-elicited PKA-regulated lipid bodies, in addition to an equally important and concomitant DP2 receptor-elicited Gαi/calcium-regulated signaling pathway, which prompts DP1 receptor-driven, newly formed lipid bodies to synthesize LTC4.
Collectively, our findings indicate that PGD2 binding to DP1 receptors triggers PKA-driven biogenesis of cytoplasmic lipid bodies, but is incapable of activating the LTC4-synthesizing machinery, which is switched on by concurrent DP2 receptor activation. Furthermore, by using PGD2 receptor antagonists, we also demonstrated that during allergen-elicited eosinophilic inflammatory reactions, cysLTs production is also regulated by DP1/DP2-orchestrated eosinophil activation, thus indicating that either DP1 or DP2 antagonists might be highly effective at controlling eosinophil activation-regulated LTC4 synthesis at sites of allergic reactions. However, considering the disappointing clinical trial results of the DP1 receptor antagonist laropiprant in asthmatics and allergic rhinitis patients (Philip et al., 2009), therapies based on dual blockade of DP1 and DP2 receptors or PGD2 synthesis inhibition may display increased beneficial outcome.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) and Fundação de Amparo à Pesquisa do Rio de Janeiro (FAPERJ, Brazil).
Glossary
Abbreviations
- 5-LO
5-lipoxygenase
- ADRP
adipose differentiation-related protein
- CRTH2
chemoattractant receptor-homologous molecule expressed on T helper type 2 cell (Th2) cells
- cysLTs
cysteinyl leukotrienes
- DP1
D prostanoid receptor 1
- DP2
D prostanoid receptor 2
- EDAC
1-ethyl-3-(3-dimethylamino-propyl) carbodiimide
- LTC4
leukotreine C4
- OVA
ovalbumin
- PGD2
prostaglandin D2
Conflict of interest
The authors have declared that no competing interests exist.
Supporting Information
Teaching Materials; Figs 1–8 as PowerPoint slide.
References
- Alexander SPH, Mathie A, Peters JA. Guide to Receptors and Channels (GRAC) Br J Pharmacol. (4th edn.) 2009;158(Suppl 1):S1–S254. doi: 10.1111/j.1476-5381.2011.01649_1.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balestrieri B, Hsu VW, Gilbert H, Leslie CC, Han WK, Bonventre JV, et al. Group V secretory phospholipase A2 translocates to the phagosome after Zymosan stimulation of mouse peritoneal macrophages and regulates phagocytosis. J Biol Chem. 2006;281:6691–6698. doi: 10.1074/jbc.M508314200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandeira-Melo C, Weller PF. Eosinophils and cysteinyl leukotrienes. Prostaglandins Leukot Essent Fatty Acids. 2003;69:135–143. doi: 10.1016/s0952-3278(03)00074-7. [DOI] [PubMed] [Google Scholar]
- Bandeira-Melo C, Phoofolo M, Weller PF. Extranuclear lipid bodies, elicited by CCR3-mediated signaling pathways, are the sites of chemokine-enhanced leukotriene C4 production in eosinophils and basophils. J Biol Chem. 2001;276:22779–22787. doi: 10.1074/jbc.M101436200. [DOI] [PubMed] [Google Scholar]
- Bandeira-Melo C, Bozza PT, Weller PF. The cellular biology of eosinophil eicosanoid formation and function. J Allergy Clin Immunol. 2002a;109:393–400. doi: 10.1067/mai.2002.121529. [DOI] [PubMed] [Google Scholar]
- Bandeira-Melo C, Sugiyama K, Woods LJ, Phoofolo M, Center DM, Cruikshank WW, et al. IL-16 promotes leukotriene C(4) and IL-4 release from human eosinophils via CD4- and autocrine CCR3-chemokine-mediated signaling. J Immunol. 2002b;168:4756–4763. doi: 10.4049/jimmunol.168.9.4756. [DOI] [PubMed] [Google Scholar]
- Bezerra-Santos CR, Vieira-de-Abreu A, Barbosa-Filho JM, Bandeira-Melo C, Piuvezam MR, Bozza PT. Anti-allergic properties of Cissampelos sympodialis and its isolated alkaloid warifteine. Int Immunopharmacol. 2006;6:1152–1160. doi: 10.1016/j.intimp.2006.02.007. [DOI] [PubMed] [Google Scholar]
- Bozza PT, Payne JL, Goulet JL, Weller PF. Mechanisms of platelet-activating factor-induced lipid body formation: requisite roles for 5-lipoxygenase and de novo protein synthesis in the compartmentalization of neutrophil lipids. J Exp Med. 1996;183:1515–1525. doi: 10.1084/jem.183.4.1515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bozza PT, Yu W, Penrose JF, Morgan ES, Dvorak AM, Weller PF. Eosinophil lipid bodies: specific, inducible intracellular sites for enhanced eicosanoid formation. J Exp Med. 1997;186:909–920. doi: 10.1084/jem.186.6.909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bozza PT, Yu W, Cassara J, Weller PF. Pathways for eosinophil lipid body induction: differing signal transduction in cells from normal and hypereosinophilic subjects. J Leukoc Biol. 1998;64:563–569. doi: 10.1002/jlb.64.4.563. [DOI] [PubMed] [Google Scholar]
- Bozza PT, Melo RC, Bandeira-Melo C. Leukocyte lipid bodies regulation and function: contribution to allergy and host defense. Pharmacol Ther. 2007;113:30–49. doi: 10.1016/j.pharmthera.2006.06.006. [DOI] [PubMed] [Google Scholar]
- Busse WW, Lemanske J. Asthma. N Eng J Med. 2001;344:350–362. doi: 10.1056/NEJM200102013440507. [DOI] [PubMed] [Google Scholar]
- Cowburn AS, Sladek K, Soja J, Adamek L, Nizankowska E, Szczeklik A, et al. Overexpression of leukotriene C4 synthase in bronchial biopsies from patients with aspirin-intolerant asthma. J Clin Invest. 1998;101:834–846. doi: 10.1172/JCI620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz BL, Serra MF, Alves AC, Cordeiro SB, Martins MA, Silva PM. Local exposure to salbutamol or Bt2 cyclic AMP inhibits pleural exudation and leukocyte influx caused by antigen in rats. Eur J Pharmacol. 1996;296:173–180. doi: 10.1016/0014-2999(95)00694-x. [DOI] [PubMed] [Google Scholar]
- Doyle WJ, Boehm S, Skoner DP. Physiologic responses to intranasal dose-response challenges with histamine, methacholine, bradykinin, and prostaglandin in adult volunteers with and without nasal allergy. J Allergy Clin Immunol. 1990;86:924–935. doi: 10.1016/s0091-6749(05)80156-3. [DOI] [PubMed] [Google Scholar]
- Drazen JM, Austen KF. Leukotrienes and airway responses. Am Rev Respir Dis. 1987;136:985–998. doi: 10.1164/ajrccm/136.4.985. [DOI] [PubMed] [Google Scholar]
- Hammad H, Kool M, Soullie T, Narumiya S, Trottein F, Hoogsteden HC, et al. Activation of the D prostanoid 1 receptor suppresses asthma by modulation of lung dendritic cell function and induction of regulatory T cells. J Exp Med. 2007;204:357–367. doi: 10.1084/jem.20061196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirai H, Tanaka K, Yoshie O, Ogawa K, Kenmotsu K, Takamori Y, et al. Prostaglandin D2 selectively induces chemotaxis in T helper type 2 cells, eosinophils, and basophils via seven-transmembrane receptor CRTH2. J Exp Med. 2001;193:255–261. doi: 10.1084/jem.193.2.255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kostenis E, Ulven T. Emerging roles of DP and CRTH2 in allergic inflammation. Trends Mol Med. 2006;12:148–158. doi: 10.1016/j.molmed.2006.02.005. [DOI] [PubMed] [Google Scholar]
- Mesquita-Santos FP, Vieira-de-Abreu A, Calheiros AS, Figueiredo IH, Castro-Faria-Neto HC, Weller PF, et al. Cutting edge: prostaglandin D2 enhances leukotriene C4 synthesis by eosinophils during allergic inflammation: synergistic in vivo role of endogenous eotaxin. J Immunol. 2006;176:1326–1330. doi: 10.4049/jimmunol.176.3.1326. [DOI] [PubMed] [Google Scholar]
- Monneret G, Gravel S, Diamond M, Rokach J, Powell WS. Prostaglandin D2 is a potent chemoattractant for human eosinophils that acts via a novel DP receptor. Blood. 2001;98:1942–1948. doi: 10.1182/blood.v98.6.1942. [DOI] [PubMed] [Google Scholar]
- Pettipher R. The roles of the prostaglandin D(2) receptors DP(1) and CRTH2 in promoting allergic responses. Br J Pharmacol. 2008;153:S191–S199. doi: 10.1038/sj.bjp.0707488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Philip G, van Adelsberg J, Loeys T, Liu N, Wong P, Lai E, et al. Clinical studies of the DP1 antagonist laropiprant in asthma and allergic rhinitis. J Allergy Clin Immunol. 2009;124:942–948. doi: 10.1016/j.jaci.2009.07.006. [DOI] [PubMed] [Google Scholar]
- Rothenberg ME, Hogan SP. The eosinophil. Annu Rev Immunol. 2006;24:147–174. doi: 10.1146/annurev.immunol.24.021605.090720. [DOI] [PubMed] [Google Scholar]
- Sandig H, Pease JE, Sabroe I. Contrary prostaglandins: the opposing roles of PGD2 and its metabolites in leukocyte function. J Leukoc Biol. 2007;81:372–382. doi: 10.1189/jlb.0706424. [DOI] [PubMed] [Google Scholar]
- Sawyer N, Cauchon E, Chateauneuf A, Cruz RP, Nicholson DW, Metters KM, et al. Molecular pharmacology of the human prostaglandin D2 receptor, CRTH2. Br J Pharmacol. 2002;137:1163–1172. doi: 10.1038/sj.bjp.0704973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schratl P, Royer JF, Kostenis E, Ulven T, Sturm EM, Waldhoer M, et al. The role of the prostaglandin D2 receptor, DP, in eosinophil trafficking. J Immunol. 2007;179:4792–4799. doi: 10.4049/jimmunol.179.7.4792. [DOI] [PubMed] [Google Scholar]
- Sousa LP, Carmo AF, Rezende BM, Lopes F, Silva DM, Alessandri AL, et al. Cyclic AMP enhances resolution of allergic pleurisy by promoting inflammatory cell apoptosis via inhibition of PI3K/Akt and NF-[kappa]B. Biochem Pharmacol. 2009;78:396–405. doi: 10.1016/j.bcp.2009.04.030. [DOI] [PubMed] [Google Scholar]
- Tedla N, Bandeira-Melo C, Tassinari P, Sloane DE, Samplaski M, Cosman D, et al. Activation of human eosinophils through leukocyte immunoglobulin-like receptor 7. Proc Natl Acad Sci USA. 2003;100:1174–1179. doi: 10.1073/pnas.0337567100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teixeira MM, Williams TJ, Hellewell PG. Anti-inflammatory effects of a short-acting and a long-acting beta 2-adrenoceptor agonist in guinea pig skin. Eur J Pharmacol. 1995;272:185–193. doi: 10.1016/0014-2999(94)00643-l. [DOI] [PubMed] [Google Scholar]
- Theiner G, Gessner A, Lutz MB. The mast cell mediator PGD2 suppresses IL-12 release by dendritic cells leading to Th2 polarized immune responses in vivo. Immunobiology. 2006;211:463–472. doi: 10.1016/j.imbio.2006.05.020. [DOI] [PubMed] [Google Scholar]
- Vieira-de-Abreu A, Assis EF, Gomes GS, Castro-Faria-Neto HC, Weller PF, Bandeira-Melo C, et al. Allergic challenge-elicited lipid bodies compartmentalize in vivo leukotriene C4 synthesis within eosinophils. Am J Respir Cell Mol Biol. 2005;33:254–261. doi: 10.1165/rcmb.2005-0145OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vieira-de-Abreu A, Calheiros AS, Mesquita-Santos FP, Magalhaes ES, Mourao-Sa D, Castro-Faria-Neto HC, et al. Crosstalk between MIF and eotaxin in allergic eosinophil activation forms LTC4-synthesizing lipid bodies. Am J Respir Cell Mol Biol. 2010 doi: 10.1165/rcmb.2010-0004OC. doi: 10.1165/rcmb.2010-0004OC. [DOI] [PubMed] [Google Scholar]
- Weller PF, Lee CW, Foster DW, Corey EJ, Austen KF, Lewis RA. Generation and metabolism of 5-lipoxygenase pathway leukotrienes by human eosinophils: predominant production of leukotriene C4. Proc Natl Acad Sci USA. 1983;80:7626–7630. doi: 10.1073/pnas.80.24.7626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widdicombe JG. Nasal pathophysiology. Respir Med. 1990;84:3–9. doi: 10.1016/s0954-6111(08)80001-7. [DOI] [PubMed] [Google Scholar]
- Xue L, Gyles SL, Barrow A, Pettipher R. Inhibition of PI3K and calcineurin suppresses chemoattractant receptor-homologous molecule expressed on Th2 cells (CRTH2)-dependent responses of Th2 lymphocytes to prostaglandin D(2) Biochem Pharmacol. 2007;73:843–853. doi: 10.1016/j.bcp.2006.11.021. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.