

Abstract
Eating high-fat chow can modify the effects of drugs acting directly or indirectly on dopamine systems and repeated intermittent drug administration can markedly increase sensitivity (i.e., sensitization) to the behavioral effects of indirect-acting dopamine receptor agonists (e.g., methamphetamine). This study examined whether eating high-fat chow alters the sensitivity of male Sprague Dawley rats to the locomotor stimulating effects of acute or repeated administration of methamphetamine. The acute effects of methamphetamine on locomotion were not different between rats (n=6/group) eating high-fat or standard chow for 1 or 4 weeks. Sensitivity to the effects of methamphetamine (0.1–10 mg/kg, i.p.) increased progressively across 4 once per week tests; this sensitization developed more rapidly and to a greater extent in rats eating high-fat chow as compared with rats eating standard chow. Thus, while eating high-fat chow does not appear to alter sensitivity of rats to acutely-administered methamphetamine, it significantly increases the sensitization that develops to repeated intermittent administration of methamphetamine. These data suggest that eating certain foods influences the development of sensitization to drugs acting on dopamine systems.
Keywords: Methamphetamine, high-fat chow, sensitization, rats, locomotor activity
1. Introduction
Changes in food intake (amount and type) can significantly impact the behavioral and neurochemical effects of some drugs, including drugs acting on dopamine systems (Owens et al., 2005; Sevak et al., 2008). For example, eating high-fat food can increase dopamine D2 receptor density, decrease dopamine transporter binding, decrease electrically evoked dopamine release, and attenuate dopamine turnover (Davis et al., 2008; Geiger et al., 2009; South and Huang, 2008; York et al., 2010). Eating high-fat chow also increases the sensitivity of rats to behavioral effects of direct-acting dopamine receptor drugs, including quinpirole (Baladi and France, 2009); however, less is known about the effects of eating high-fat food on sensitivity to indirect-acting dopamine receptor agonists (e.g., methamphetamine).
Stimulants such as methamphetamine act at the dopamine transporter and increase extracellular dopamine that, in turn, acts on all dopamine receptor subtypes. In humans, repeated administration of stimulants can increase sensitivity to their effects and this “sensitization” is thought to be an important component of chronic stimulant abuse (e.g., Ujike and Sato, 2004). In rats, sensitivity to the behavioral effects (e.g., increased locomotion) of stimulants (e.g., methamphetamine) increases after repeated administration (Robinson and Becker, 1986; White and Kalivas, 1998); however, it is not known whether changes in food intake, that have been shown to alter sensitivity to direct- or indirect-acting dopamine receptor agonists, alter the development of sensitization to stimulant drugs.
This study examined whether eating high-fat chow alters the sensitivity of rats to the effects of methamphetamine on locomotion and whether eating high-fat chow modifies the development of sensitization to methamphetamine. Different groups of rats ate standard or high-fat chow and were tested once or once per week for 4 weeks with methamphetamine using a cumulative-dosing procedure. Based on emerging evidence showing increased sensitivity to direct-acting dopamine receptor agonists in rats eating high-fat chow (e.g., quinpirole; Baladi and France, 2009, 2010), it was expected that eating high-fat chow would also increase the sensitivity of rats to methamphetamine and also might increase the development of sensitization to methamphetamine.
2. Materials and Methods
2.1. Subjects
Twenty-four adult male Sprague Dawley rats (Harlan, Indianapolis, IN), 63 days old upon arrival (experiments began one week later when rats were 70 days old), were housed individually in an environmentally controlled room (24 ± 1 °C, 50 ± 10% relative humidity) under a 12/12 h light/dark cycle (lights on at 0700 hr). With the exception of locomotion testing, rats had free access to food and water. Animals were maintained and experiments were conducted in accordance with the Institutional Animal Care and Use Committee, the University of Texas Health Science Center at San Antonio, and the 1996 Guide for Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources on Life Sciences, the National Research Council, and the National Academy of Sciences).
2.2. Apparatus
Locomotion was studied using custom built (Instrumentation Services, University of Texas Health Science Center, San Antonio, Texas, USA) Lexan® polycarbonate (General Electric Structured Products, Mt. Vernon, Indiana, USA) chambers (26×61×23 cm high) equipped with metal floors and located within sound attenuating cubicles (MED Associates Inc., St. Albans, Vermont, USA). Horizontal activity (locomotor activity) was measured with 4 pairs of infrared photo beams (Multi-Varimex, Columbus Instruments, Columbus, Ohio, USA) positioned 4 cm above the floor of the chamber. The photo beams were separated by 15 cm with two of the photo beams located 8 cm from the ends of the chamber.
2.3. Feeding Conditions
After arrival in the laboratory all rats had free access to a standard rat chow for one week prior to the beginning of the study. One group of 12 rats was tested once per week with methamphetamine during 4 weeks of free access to either standard (n=6) or high-fat chow (n=6). A second group of 12 rats was tested once with methamphetamine after 4 weeks of free access to either standard (n=6) or high-fat (n=6) chow; during the first 3 weeks of the study these 12 rats were tested once per week with injections of saline only. The high-fat chow was 34.3% fat (by weight) and 23.5% protein, 5.1 kcal/g (Harlan Teklad #TD.06414; Harlan Laboratories, Madison, Wisconsin, USA). The standard chow was 5.7% fat and 19.9% protein, 4.1 kcal/g (Harlan Teklad #7912). Body weights were measured daily throughout the study.
2.4. Locomotor activity
All 24 rats had free access to standard chow for 7 days; on 3 of those days, rats were placed in the chamber for 105 min periods when baseline locomotion was assessed. In 12 rats (n=6/type of chow) cumulative methamphetamine dose-response curves were determined once per week during 4 weeks of free access to standard or high-fat chow. The remaining 12 rats (n=6/type of chow) were tested one per week with saline for 3 weeks and with methamphetamine in week 4. All rats received the same number of injections (6 per session) and sessions (8 total) in the locomotor chamber, although they differed in the type of chow they ate and the number of times that they received methamphetamine.
For testing, rats were placed in the chamber for an initial 30-min habituation period. Pilot studies indicated that the increase in locomotion that occurs when rats are placed in the chamber is no longer evident after 30 min. Thus, in test sessions and after the 30-min habituation period, rats received an injection (drug or saline) every 15 min. For methamphetamine tests, increasing doses were administered with the cumulative dose increasing by 0.5 log units per injection (i.e., 0.1, 0.32, 1.0, 3.2 and 10 mg/kg, i.p.). Locomotor activity was monitored continuously and data are presented for 5-min periods beginning 10 min after injections (i.e., minutes 11–15 of each cycle).
2.5. Drugs
(+) Methamphetamine hydrochloride was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in sterile 0.9% sodium chloride. Drug and saline were injected i.p. in a volume of 1 ml/kg body weight.
2.6. Data Analysis
Locomotor activity, measured as total number of beam breaks was monitored continuously through the session and averaged over 5-min bins. Activity counts were averaged among rats in a feeding condition and plotted as the average number of counts ± standard error of the mean (S.E.M.) as a function of dose. Statistical analyses were performed using two-way (dose and feeding condition) ANOVA (GraphPad Software Inc., San Diego, California) and Bonferroni post-hoc tests. Locomotion in saline test sessions was compared and found not to vary significantly across cycles within a group or among groups within a session; thus, saline data were pooled and the dashed lines on Figs. 2 and 3 indicate ± 1 S.E.M. of the mean of those pooled data. The maximum increase in locomotion (for all doses in all groups) was also compared across all weekly tests (vehicle or methamphetamine). For all tests, significance was set at P < 0.05.
Fig 2.

Effects of a range of methamphetamine doses (0.1–10 mg/kg) on locomotor activity in rats eating high-fat chow (open symbols; n=6) or standard chow (closed symbols; n=6) during tests conducted once per week for each of the 4 weeks of this study. Abscissa, dose in milligrams per kilogram of body weight; V = vehicle. Ordinates, mean locomotor activity counts/5 min (± S.E.M.). Dashed lines represent ± 1 S.E.M. of the mean of pooled data for all 12 rats for saline test sessions.
Fig 3.
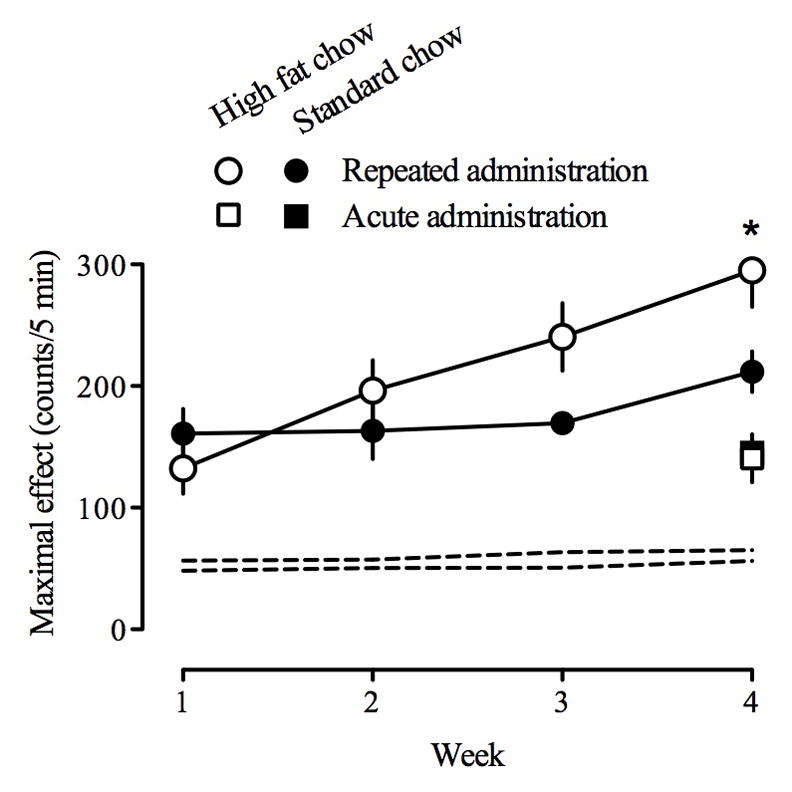
Maximal locomotor effects obtained with methamphetamine across the 4 once per week tests (circles) or in a single test during week 4 (squares). Animals represented by closed symbols ate standard chow and those represented by open symbols ate high-fat chow (n=6/group). Ordinate, mean locomotor activity counts/5 min (± S.E.M.). Abscissa, week; animals represented by circles received methamphetamine once per week for 4 weeks whereas those represented by squares received saline once per week during weeks 1–3 and methamphetamine in week 4. Dashed lines represent ± 1 S.E.M. of the mean of pooled data for all 24 rats for saline test sessions.
3. Results
3.1 Repeated methamphetamine tests
The average body weight (mean ± S.E.M.) of rats that were tested once per week with methamphetamine was not significantly different, at the beginning or at the end of the study, between those eating standard chow and those eating high-fat chow: 310 ± 2 and 305 ± 3 g at the beginning of the study, respectively; 371 ± 4 and 387 ± 8 g after 4 weeks, respectively (circles, Fig. 1). In all rats, small doses of methamphetamine increased while larger doses decreased locomotion, yielding an inverted U-shaped dose-response curve (Fig. 2). After 1 week on their respective feeding condition, the locomotor stimulating effects of methamphetamine were not different between rats eating standard chow and rats eating high-fat chow (upper panel Fig. 1). In both groups, locomotor activity peaked at a dose of 1.0 mg/kg methamphetamine with an average (mean ± S.E.M.) of 143 ± 24 counts for rats eating standard chow and 132 ± 21 counts for rats eating high-fat chow.
Fig 1.

Body weights of rats tested once (squares) or once per week for 4 weeks (circles) with methamphetamine and eating either standard (closed symbols) or high-fat (open symbols) chow (n = 6 per group). Ordinate: body weight in g. Abscissa: weeks of the study.
With repeated weekly drug testing, the ascending limb of the methamphetamine dose-response curve shifted up and to the left (i.e. sensitization) for all rats, regardless of feeding condition (compare all panels, Fig. 2). During week 4, locomotor activity peaked at a dose of 0.32 mg/kg methamphetamine for both groups and the maximum increase in locomotion was significantly greater in rats eating high-fat chow (295 ± 30) as compared with rats eating standard chow (211 ± 17). The maximum effect obtained in each test for both groups is plotted in Fig. 3 (circles). This function indicates the rate at which sensitivity increased to the locomotor stimulating effects of methamphetamine, and the significantly greater slope of the function in rats eating high-fat chow indicates more rapid development of sensitization in those rats (Fig. 3). There was no significant change in locomotion across weekly saline tests in any group of rats (dashed lines, Figs. 2 and 3).
3.2 Single methamphetamine test
The average body weight (mean ± S.E.M.) of rats that were tested three times with saline followed by a single test with methamphetamine was not significantly different, at the beginning or the end of the study, between those eating standard chow and those eating high-fat chow: 277 ± 3 and 276 ± 2 g at the beginning of the study, respectively; 355 ± 9 and 359 ± 8 g after 4 weeks, respectively (squares, Fig. 1). After 4 weeks of free access to either standard or high-fat chow, rats were tested with increasing doses of methamphetamine. In all rats, small doses of methamphetamine increased while larger doses decreased locomotion, yielding an inverted U-shaped dose-response curve (data not shown). The locomotor-stimulating effects of a dose of 0.32 mg/kg methamphetamine were significantly greater in rats eating high-fat chow as compared with rats eating standard chow; the effects of other doses of methamphetamine were not different between groups (data not shown). Moreover, the maximal effect obtained with methamphetamine was not different between the two groups (compare open and closed squares above “4”, Fig. 3). Locomotor activity was maximal after administration of 1.0 mg/kg methamphetamine for both groups: 141 ± 15 counts for rats eating a standard chow and 129 ± 21 counts for rats eating a high-fat chow. The effects of methamphetamine on locomotion also were not different between groups of rats that first were tested with methamphetamine after 1 week or after 4 weeks of eating standard or high-fat chow (compare circles above “1” to squares above “4”, Fig. 3). The rate at which sensitivity increased (i.e., sensitization) to the locomotor stimulating effects of methamphetamine was greater in rats eating high-fat chow as compared with rats eating standard chow (compare open and closed circles, Fig. 4).
4. Discussion
It is well established that changes in food intake (amount and type) can modify the behavioral and neurochemical effects of some drugs (Davis et al., 2008; South and Huang, 2008). For example, eating high-fat food enhances both conditioned (discriminative stimulus; Baladi and France, 2010) and unconditioned (yawning; Baladi and France, 2009) effects of the direct-acting dopamine receptor agonist quinpirole. Several drugs of abuse (e.g., methamphetamine) are indirect-acting dopamine receptor agonists that increase extracellular concentrations of dopamine by actions at the dopamine transporter; however, it is not clear to what extent changes in food intake (e.g., eating high-fat chow), that modify the actions of direct-acting dopamine receptor agonists like quinpirole, also modify the effects of indirect-acting agonists like methamphetamine. The major finding of this study is that eating high-fat chow enhances the development of sensitization to the effects of methamphetamine on locomotion.
Eating high-fat chow for as little as 2 weeks can significantly increase the sensitivity of rats to the behavioral effects of different doses of direct-acting dopamine receptor agonists (Baladi and France, 2009). In contrast, eating high-fat chow for as long as 4 weeks had little effect on the initial sensitivity of rats to methamphetamine-induced locomotion. This differential effect of eating high-fat chow on behavioral effects of direct- and indirect-acting dopamine agonists might result from qualitatively different changes in dopamine receptors and transporters (e.g., South and Huang, 2008) or from quantitatively (e.g., temporally) distinct changes that occur in rats eating high-fat chow. For example, it is possible that sensitivity to methamphetamine increases after longer (i.e., more than 4 weeks) access to high-fat chow. On the other hand, eating high-fat chow alters behavioral effects of direct-acting agonists that are mediated through dopamine D2 or D3 receptors. Thus, other dopamine receptors that are thought to play a role in the effects of methamphetamine on locomotion (e.g., D1) might not be impacted by feeding conditions that alter sensitivity to drugs acting through dopamine D2 or D3 receptors.
Sensitization is a progressive and sometimes permanent enhancement of a drug effect (e.g., methamphetamine-induced locomotion) that develops after repeated intermittent drug administration (Amano et al., 2003). In the current study, sensitization to the effects of methamphetamine on locomotion developed in all rats that were tested repeatedly (weekly) with methamphetamine; however, the rate at which sensitization developed was significantly greater in rats eating high-fat chow as compared to those eating standard chow. Indirect-acting agonists, like methamphetamine, increase extracellular concentrations of dopamine that act on all dopamine receptor subtypes. Among those dopamine receptors, D1 and D2 receptors are thought to be particularly important in the development of sensitization (Kuribara and Uchihashi, 1993; Shuto et al., 2008). For example, the number of dopamine D2 receptors is not changed in animals displaying behavioral sensitization to methamphetamine, although there is a significant increase in the high affinity state of dopamine D2 receptors that parallels the development of sensitization (Amano et al., 2003; Seeman et al., 2002). Moreover, eating high-fat chow increases sensitivity to effects of direct-acting agonists that are mediated by dopamine D2 receptors (e.g. Baladi and France, 2009). Collectively, these results suggest that eating high-fat chow impacts dopamine D2 receptor sensitivity in a manner that both increases sensitivity to direct-acting agonists and facilitates the development of sensitization to methamphetamine.
In summary, this study demonstrates that with repeated administration, sensitization develops to the locomotor effects of methamphetamine regardless of whether animals eat a standard or high-fat chow. In addition, eating high-fat chow increases the rate of sensitization to the locomotor effects of methamphetamine, as compared with the development of sensitization in rats eating a standard chow. That is, after 4 weeks of unlimited access to their respective diets and once per week administration of methamphetamine, the maximal effect for locomotor activity was significantly greater for rats eating high-fat chow. Insofar as sensitization is thought to be an important component of chronic stimulant abuse, these results suggest that the development of sensitization could be significantly influenced by the consumption of certain foods.
Acknowledgments
CPF is supported by Senior Scientist Award from the National Institute on Drug Abuse (DA17918).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Blaine A. McGuire, Email: mcguireb@uthscsa.edu.
Michelle G. Baladi, Email: baladi@uthscsa.edu.
Charles P. France, Email: france@uthscsa.edu.
References
- Amano T, Matsubayashi H, Seki T, Sasa M, Sakai M. Repeated administration of methamphetamine causes hypersensitivity of D2 receptor in rat ventral tegmental area. Neurosci Lett. 2003;347:89–92. doi: 10.1016/s0304-3940(03)00673-6. [DOI] [PubMed] [Google Scholar]
- Baladi MG, France CP. High fat diet and food restriction differentially modify the behavioral effects of quinpirole and raclopride in rats. Euro J Pharm. 2009;610:55–60. doi: 10.1016/j.ejphar.2009.03.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baladi MG, France CP. Eating high-fat chow increases the sensitivity of rats to quinpirole-induced discriminative stimulus effects and yawning. Behav Pharmacol. 2010;21:615–620. doi: 10.1097/FBP.0b013e32833e7e5a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiger BM, Haburcak M, Avena NM, Moyer MC, Hoebel BG, Pathos EN. Deficits of mesolimbic dopamine neurotransmission in rat dietary obesity. Neuroscience. 2009;159:1193–1199. doi: 10.1016/j.neuroscience.2009.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuribara H, Uchihashi Y. Dopamine antagonists can inhibit methamphetamine sensitization, but not cocaine sensitization, when assessed by ambulatory activity in mice. J Pharm Pharmacol. 1993;45:1042–1045. doi: 10.1111/j.2042-7158.1993.tb07177.x. [DOI] [PubMed] [Google Scholar]
- Owens WA, Sevak RJ, Galici A, France CP, Daws LC. Deficits in dopamine clearance and locomotion in hypoinsulinemic rats are restored by repeated administration of amphetamine. J Neurochem. 2005;94:1402–1410. doi: 10.1111/j.1471-4159.2005.03289.x. [DOI] [PubMed] [Google Scholar]
- Robinson TE, Becker JB. Enduring changes in brain and behavior produced by chronic amphetamine administration: A review and evaluation of animal models of amphetamine psychosis. Brain Res Rev. 1996;11:157–198. doi: 10.1016/s0006-8993(86)80193-7. [DOI] [PubMed] [Google Scholar]
- Seeman P, Tallerico T, Ko F, Tenn C, Kapus S. Amphetamine–sensitized animals show a marked increase in dopamine D2 high receptors occupied by endogenous dopamine, even in the absence of acute challenges. Synapse. 2002;46:235–239. doi: 10.1002/syn.10139. [DOI] [PubMed] [Google Scholar]
- Sevak RJ, Daws LC, Koek W, Owens WA, Galli A, France CP. Feeding conditions differentially affect the neurochemical and behavioral effects of dopaminergic drugs in male rats. Eur J Pharmacol. 2008;592:109–115. doi: 10.1016/j.ejphar.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shuto T, Seeman P, Kuroiwa M, Nishi A. Repeated administration of a dopamine D1 receptor agonist reverses the increases proportions of striatal dopamine D1high and D2high receptors in methamphetamine sensitized rats. Eur J Neurosci. 2008;27:2551–2557. doi: 10.1111/j.1460-9568.2008.06221.x. [DOI] [PubMed] [Google Scholar]
- South T, Huang XF. High-fat diet exposure increases dopamine D2 receptor and decreases dopamine transporter receptor binding density in the nucleus accumbens and caudate putamen of mice. Neurochem Res. 2008;33:598–605. doi: 10.1007/s11064-007-9483-x. [DOI] [PubMed] [Google Scholar]
- Ujike H, Sato M. Clinical features of sensitization to methamphetamine observed in patients with methamphetamine dependence and psychosis. Ann N Y Acad Sci. 2004;1025:279–287. doi: 10.1196/annals.1316.035. [DOI] [PubMed] [Google Scholar]
- White FJ, Kalivas PW. Neuroadaptations involved in amphetamine and cocaine addiction. Drug Alcohol Depend. 1998;51:141–153. doi: 10.1016/s0376-8716(98)00072-6. [DOI] [PubMed] [Google Scholar]
- York DA, Teng L, Park-York M. Effects of dietary fat and enterostatin on dopamine and 5-hydroxytryptamine release from rat striatal slices. Brain Res. 2010;1349:48–55. doi: 10.1016/j.brainres.2010.06.036. [DOI] [PubMed] [Google Scholar]