

Abstract
Animal studies suggest an involvement of CA3 and dentate gyrus (CA3&DG) in memory encoding and early retrieval and an involvement of CA1 in late retrieval, consolidation and recognition. The aim of this study was to test if similar associations could be found between hippocampal subfield volumes measured in vivo using a manual parcellation scheme and selected scores of the California Verbal Learning Test II (CVLTII): Total Immediate Free Recall Discriminability (IFRD), Short Free Recall Discriminability (SFRD), and Delayed Recall Discriminability (DRD). 50 elderly subjects (25 controls, 25 cognitively impaired subjects) had CVLTII and high resolution hippocampal MRI at 4T. Entorhinal cortex, subiculum, CA1, CA1-CA2 transition zone, and CA3&DG were manually marked on five slices in the anterior hippocampal body on the MRI. Pearson correlations followed by stepwise regression analysis were used to test for associations between subfield volumes and CVLTII. IFRD, SFRD, which are measures of encoding/early retrieval, were associated with CA3&DG, and DRD, which measures consolidation/late retrieval, with CA1. These preliminary findings demonstrate that subfield volumetry has the potential to study non invasively subfield specific memory functions.
Keywords: subfield, cornu amonis sector, recall, recognition, high resolution MRI
1. Introduction
Ever since Scoville’s and Milner’s (1957) landmark paper about H.M. who suffered from severe memory disturbances after bilateral temporal lobe resections, there has been general agreement that the hippocampus plays a crucial role in learning and memory. The hippocampus is not a homogeneous structure but consists of several histologically distinct subfields: Subiculum, cornu ammonis sectors (CA) 1–3 and dentate gyrus (DG). (Duvernoy, 2005). Despite their histological specialization, these sectors are tightly interconnected. Currently, at least two major hippocampal pathways are distinguished: 1. The indirect pathway which connects the entorhinal cortex (ERC) with the DG, CA3 and finally CA1 and 2. The direct pathway which connects the ERC directly with CA1 from where it projects back via subiculum to the ERC (Duvernoy, 2005). The existence of different pathways and different histological characteristics suggests that different hippocampal subfields might process different aspects of the memory process, e.g., show a specialization for encoding, consolidation, retrieval etc. This notion is supported by findings of animal models, which used sophisticated memory tasks to tease out different aspects of the learning/memory formation process. These studies suggest that CA3 and DG might be primarily responsible for encoding and early retrieval (Acsady and Kali, 2007; Hasselmo 2005) while CA1 is responsible for consolidation, late retrieval and recognition (Nakazawa et al. 2004, O’Reilly et al. 2001, Hunsaker and Kesner 2008, Rolls and Kesner, 2006, Wan et al 1999, Kesner and Hunsaker, 2010). Evidence that a similar functional specialization might also exist in the human hippocampus comes mostly from case studies of patients with “lesions mostly confined to the hippocampus” or from patients with transient global amnesia (Manns et al., 2003; Bayley et al. 2008; Bird et al. 2008; Kirwan et al. 2009; Bartsch et al. 2006; Quinette et al 2006). However, although these studies demonstrated impairment in encoding, retrieval etc. they did usually not allow to identify how damage to specific subfields contributed to the observed deficits. Only recently fMRI studies using sequences with relative high resolution (ca. 1.5x1.5x1.5 mm) and highly specialized memory tasks succeeded in demonstrating that a specialization of hippocampal subfields similar as that described in animals exists also in healthy and cognitively impaired humans (Eldridge et al. 2005; Bakker et al., 2008; Suthana et al. 2009; Yassa et al. 2010).
Memory impairment is a characteristic early sign of Alzheimer’s Disease (AD) and other forms of dementia. Atrophy of the memory related medial temporal structures, particularly the hippocampal formation, is one of its earliest macroscopical hallmarks and has been consistently reported in autopsy and neuroimaging studies. However, histological studies suggest that not all hippocampal subfields are equally affected by AD. Particularly in the early stages of the disease, neuron loss and tangle accumulation are more prominent in CA1 and subiculum than in other subfields (Bobisnki et al. 1998; Schoenheit et al 2004; Roessler et al 2001, West et al. 1994). Recent advancements with high field MRI (3–4 Tesla) have resulted in improved anatomical images of the brain at sub-millimeter resolution. Although these images do not have the resolution of a histological preparation, they depict details of the internal structure of the hippocampus which together with external arbitrary hippocampal landmarks can be used to further subdivide the hippocampus into “subfields” and thus to obtain “subfield specific“ volumes (Mueller et al. 2007). Preliminary studies have shown that the subfield specific atrophy pattern detected by this method in subjects at risk for AD or diagnosed with AD are in good agreement with the findings described in histopathological studies in these stages (Mueller and Weiner, 2009, Mueller et al. 2010).
Given the evidence for a subfield specialization for different aspects of the memory process and the regional selective hippocampal atrophy in the earliest stages of AD, the overall goal of this study was to test if it is possible to find associations between specific subfield volumes and different aspects of a clinical standard memory test in a population of elderly cognitively intact and mildly impaired subjects at risk for the development of AD. The neuropsychological test chosen for this purpose was the California Verbal Learning Test II (CVLTII, Delis et al. 2000), particularly the variables total immediate free recall discriminability (IFRD), short free recall discriminability (SFRD) and delayed recall discriminability (DRD). IFRD and SFRD were chosen as measures representing encoding (IFRD) and early retrieval (SFRD), and DRD as a measure for delayed retrieval and DRD corrected for IFRD performance (Kramer et al. 2004) as measure for consolidation (please see Neuropsychological testing in the methods section for additional details). In accordance with the findings of animal and fMRI studies, the following specific hypotheses were tested: 1. CA1 atrophy is associated with DRD and DRD corrected for IFRD performance. 2. IFRD and SFRD impairment are associated with volume loss in CA3 and dentate gyrus (DG).
2. Methods
2.1 Study population
The subjects in this study had been recruited for a NIH sponsored project (R01AG010897 PI Dr. M. Weiner). Eligible subjects were over 60 years of age who reported subjective memory problems but were not demented. All subjects underwent a medical examination and laboratory testing. Subjects suffering from medical conditions or taking medications affecting cognitive performance were excluded as were subjects with a history of brain trauma, brain surgery or evidence for stroke (exception: white matter hyperintensities or small lacunes), and skull defects on the MRI. At the time of the preparation of this article, 91 subjects had been recruited into the study. Forty-one of these 91 subjects had to be excluded for various reasons (refused ApoE testing: 7, no CVLTII: 13, pathological 4 T MRI findings, e.g., stroke: 3, no high resolution T2 image: 4, bad image quality: 14). A total of 50 subjects were included in this study (cf. Table 1). Twenty-five subjects were cognitively impaired but not demented (CIND) and had been referred from collaborating Memory Clinics (UCSF, VA Medical Center San Francisco). Fifteen had been diagnosed with amnestic mild cognitive impairment (aMCI) according to the criteria of Petersen (1999). The remaining 10 subjects had been diagnosed as being cognitively impaired (memory deficits: 6; executive deficits: 4) in the clinical examination but did not fulfill the criteria for aMCI. The remaining twenty-five subjects were cognitively intact elderly controls. Cognitive functioning in CIND and controls was assessed by a standardized battery of neuropsychological tests (mini mental state examination, CVLT II, American National Adult Reading Test, Design and Verbal Fluency, Wechsler Adult Intelligence Score (digit symbol, matrix reasoning, vocabulary), Wechsler Memory Scale II (visual reproduction immediate), trails; emotional state and functioning in daily living were assessed with the Geriatric Depression Scale and the Functional Activities Questionnaire (cf. supplementary Table 1). This test battery was part of the routine clinical evaluation for subjective memory problems in CIND and was administered to controls for the purpose of this study, i.e., all subgroups were equally exposed to cognitive testing. All subjects had agreed to undergo testing for the apolipoprotein E epsilon 4 (Apo E4) allele. A previous study from this lab had found significantly reduced CA3 and DG volumes in elderly subjects carrying the Apo E4 allele regardless of their cognitive status (Mueller et al. 2008) and thus the information about Apo E4 carrier state was included in the analysis. Eight subjects (2 controls, 6 CIND) in the current population had also been included in this previous study. Eighteen subjects were Apo E4 positive, i.e., had at least one e4 allele (2/4: 3; 3/4: 12; 4/4: 3), 32 were Apo E4 negative, i.e. had no e4 allele (2/3: 5; 3/3: 27). The study had been approved by the committees of human research at the University of California, San Francisco and Veteran’s Administration Medical Center, San Francisco and written informed consent was obtained from all subjects or their legal representatives.
Table 1.
Characteristics of Study Participants
| Controls ( n = 25) | CIND (n = 25) | |
|---|---|---|
| Demographics | ||
| Age (mean(SD) | 70.5 (8.2) | 72.4 (6.7) |
| Gender (female/male) | 14/11 | 8/17 |
| Years of Education (mean (SD) | 16.5 (2.6) | 15.6 (3.5) |
| Apo E4 carrier state (yes/no) | 7/18 | 11/14 |
| Neuropsychological Scores | ||
| MMSE (mean(SD)) | 29.6 (1.0) (range: 26–30) | 28.2 (1.8)* (range: 24–30) |
| IFRD (mean(SD)) | 2.3 (0.5) (range: 1.3–3.2) | 1.8 (0.5)* (range: 1.2–2.8) |
| SFRD (mean(SD)) | 2.4 (0.8) (range: 0.5–3.7) | 1.7 (0.8)* (range: 0–3.4) |
| DRD (mean(SD)) | 2.4 (0.7) (range: 0.9–3.7) | 1.6 (0.9) (range: −0.4–2.8) |
| Subfield Volumes (mm3) | ||
| ERC (mean(SD)) | 104.8 (25.1) (range: 56.5–151.5) | 94.6 (20.7) (range: 44.2–132.7) |
| Sub (mean(SD)) | 108.5 (21.7) (range: 70.0–144.2) | 108.2 (19.6) (range: 70.6–147.3) |
| CA1 (mean(SD)) | 201.9 (24.7) (range: 141.8–233.6) | 192.3 (39.0) (range: 93.8–258.4) |
| CA1-2 (mean(SD)) | 10.1 (1.4) (range: 8.0–13.5) | 9.6 (1.7) (range: 6.9–13.2) |
| CA3&DG (mean(SD)) | 148.3 (22.5) (range: 141.8–233.6) | 144.3 (26.7) (range: 99.2–212.1) |
p <0.05 lower than controls, SD, standard deviation. MMSE, mini mental state examination, IFRD, immediate free recall discriminability total score, SFRD, short free recall discriminability score; DRD, delayed free recall discriminability score. ERC, entorhinal cortex; Sub, subiculum; CA1, cornu ammonis sector 1; CA1-2, CA1-2 transition (cf. methods for definition); CA3&DG, combined cornu ammonis sector 3 and dentate gyrus. Volumes are corrected for ICV (cf. methods section)
2. 2. Neuropsychological assessment
All subjects were tested with the full version of the CVLT II (Delis et al .2005). The CVLT II presents a list of 16 items over 5 learning trials. Immediately after reading the list to the subject, the subject is asked to recall as many items as possible (immediate recall). The performance during the first trial is thought to be mostly determined by the auditory attention span. The performance during trials 2–5 is thought to reflect the verbal learning or encoding abilities of the examinee. Cognitively intact subjects are expected to learn additional words on the list with each repetition and thus to be able to increase their performance with each trial. The IFRD provides a summary measure of the performance during all five trials. Then a second, different list is introduced which has to be recalled once. This is immediately followed by asking the subject to recall again the items of the first list (SFDR). Due to the short time (ca 5 min) between trial 5 and the new attempt to recall the first list without having heard it again, SFRD is considered to be a measure of short term recall or early retrieval. After an additional delay of about 20 min during which non-related, non-verbal tasks are performed, the subject is again asked to recall the first item list freely and with the help of cues (DRD). A measure of retention or consolidation over time can be obtained from DRD by controlling for initial memory performance, i.e., IFRD (Kramer et al. 2004). Potential effects of age, gender and education on these scores were assessed in the preliminary statistical analysis (cf. Statistical Analysis) and included into the final statistical models if they showed a significant effect in these preliminary analyses.
2.3. MRI acquisition
All imaging was performed on a Bruker MedSpec 4T system controlled by a Siemens TrioTM console and equipped with a USA instruments eight channel array coil that consisted of a separate transmit coil enclosing the eight receiver coils. The following sequences were acquired: 1. For the measurement of hippocampal subfields, a high resolution T2 weighted fast spin echo sequence (TR/TE: 3990/21 ms, 0.4 x 0.4 mm in plane resolution, 2 mm slice thickness, 24 slices covering the anterior three quarters and in some cases the whole hippocampus, acquisition time 5:30 min ). , and 2. For the determination of the intracranial volume (ICV), a T2 weighted turbospin echo sequence (TR/TE 8390/70 ms, 0.9 x 0.9 x 3 mm nominal resolution, 54 slices, acquisition time 3.06 min).
2.4. Hippocampal subfield volumetry
The method used for manual subfield marking including assessment of measurement reliability and its limitations has been described in detail previously (Mueller et al. 2007, Mueller et al. 2008, Mueller et al. 2010). To summarize it briefly: The marking scheme depends on anatomical landmarks, particularly on a hypointense line representing myelinated fibers in the stratum moleculare/lacunosum (Eriksson et al. 2008) which can be reliably visualized on these high resolution images. The distance between this hypointense line and the outer boundary of the hippocampus provides an estimate of the cortical thickness of the hippocampus at this point. Additional external and internal hippocampal landmarks are used to further subdivide the hippocampus into subiculum, CA1, CA1-2 transition zone (CA1-2 transition), CA3 and dentate gyrus. The latter two are lumped together (CA3&DG) because there are no macroscopical landmarks to separate them (cf. Figure 1a). CA1-2 transition is in the dorsal medial region of the hippocampus and consists mostly of CA2. However, due to the landmarks used for labeling it, its volume is influenced by the thickness of the dorsal CA1. To reflect this fact, the sector is called CA1-2 transition rather than CA2. Altogether, hippocampal subfields were marked on a length of 1 cm in the anterior third of the hippocampal body (cf. Figure 1). ICV was determined from the T2 weighted image using the BET program for skull stripping it (FMRIB Image Analysis Group, Oxford University, www.fmrib.ox.ac.uk/fsl).
Figure 1.
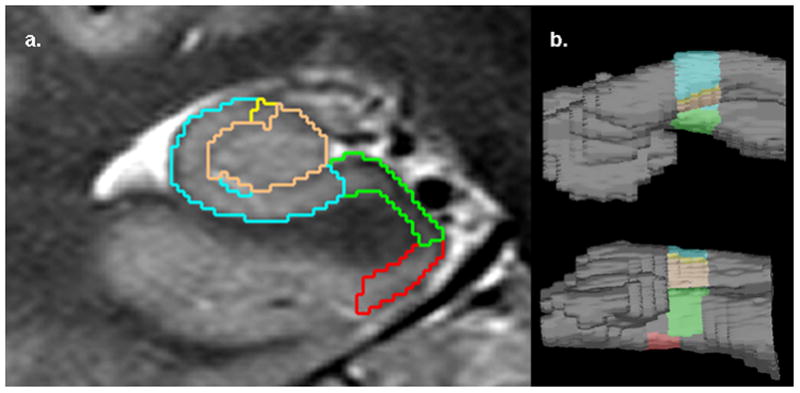
a. Manual hippocampal subfield parcellation scheme and high resolution hippocampal MRI of a 74 years old female aMCI. Red, entorhinal cortex, Green, subiculum, blue; CA1; yellow, CA1-2 transition, i.e. CA2 partially overlapping CA1; maroon, CA3 and dentate gyrus.. b. 3 dimensional representation of the hippocampal section used for subfield parcellation. The number of slices marked in this parcellation scheme is kept constant. Upper panel: Right hippocampus view from above. Lower panel: Right hippocampus view from medial.
2.5. Statistical Analysis
Left and right subfield volumes were combined for the statistical analysis. Multiple linear regression analyses were used to test for effect of group (controls vs CIND) and Apo E4 (Apo E4 pos. vs ApoE4 neg.) on subfield volumes and CVLTII scores. Age (subfields and CVLTII scores), years of education (CVLTII scores only), gender (subfields and CVLTII-scores) and intracranial volume (subfield volumes) were included as independent variables. Correction for head size for the following analyses (correlation, stepwise regression) was done using the formula: corrected volume = raw volume * 1000/ICV. CIND were combined with controls after ensuring that there was an overlap between the two groups by correlation plots to allow for the assessment of the full range of structural–functional associations. In a first step, Pearson correlation analyses were used to test for correlations between subfield volumes and the selected CVLTII scores. However, subfield volumes were also correlated with each other (r: range: 0.22–0.78, cf. correlation matrix in Table 2 of supplementary material) and thus some of these subfield-CVLT correlations could be spurious. In order to exclude such spurious associations and to identify those subfield volumes which were alone or in combination with other subfields significantly associated with a given CVLTII score, stepwise linear regression analyses (forward, probability to enter/leave p = 0.05) with the CVLTII score as dependent and those subfield volumes showing significant correlations with this score in the correlation analysis as independent variables were performed in the next step. Based on the results of the preliminary regression analyses, age and Apo E4 carrier state were forced into these stepwise regression models as independent variables so that only subfield volumes explaining a significant amount of the variation of the memory performance in addition to that explained by these independent variables could be included in the stepwise regression model. To test which subfield volume contributed most to consolidation, a slightly different stepwise regression analysis was performed. In this model DRD was defined as dependent variable, age and IFRD (cf. Introduction) were then forced into the model and Apo E4 and those subfield volumes which were significantly correlated with DRD in the initial correlation analysis were entered in a stepwise fashion. Group was not modeled in any of the stepwise regression analyses because it was intended to test for structural-cognitive correlations over the whole range of memory impairment and subfield volumes represented by the two groups. To exclude that the associations were driven by the two groups forming two separate clusters, the memory scores of the two populations were plotted against the subfield volumes corrected for ICV (c.f. Figure 2.2.). Furthermore, it was established that the slopes of the memory score-subfield volume associations were not different between the two subgroups and thus no additional information was lost by combining the two groups. .Given the a priori hypotheses stated in the Introduction, corrections for multiple comparisons were not performed. All statistical analyses were done in JMP7 (SAS Institute Inc.).
Figure 2.
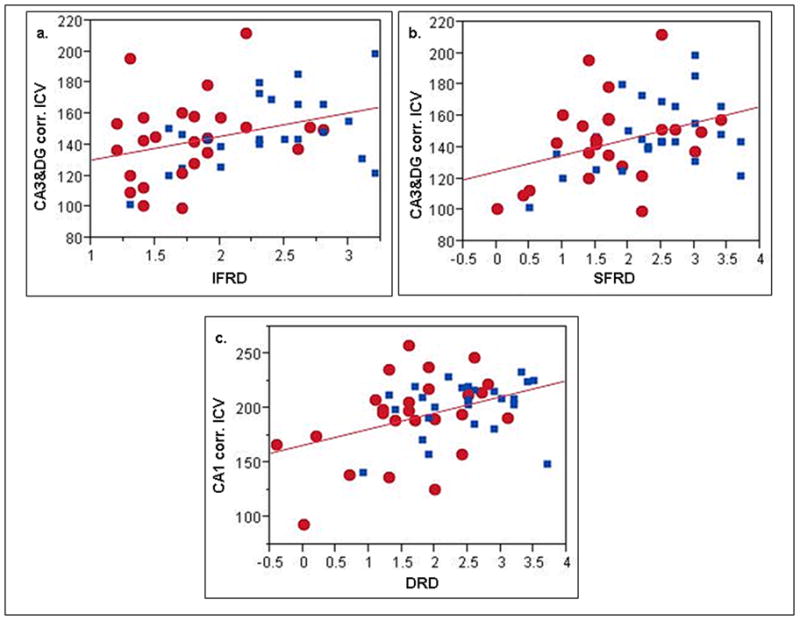
Scatter plots of subfield volumes vs. CVLTII subscores. a. IFRD vs. CA3&DG volumes. B. SFRD vs. CA3&DG volumes and c. DRD vs CA1 volumes. Red dots, CIND subjects, blue squares control subjects; CA3&DG, combined CA3 and dentate gyrus volumes in mm3 corrected for intracranial volume (cf text for formula), CA1, CA1 volumes in mm3 corrected for intracranial volume. IFRD, Total Immediate Free Recall Discriminability; SFRD, Short Free Recall Discriminability and DRD, Delayed Recall Discriminability.
3. Results
3.1. Group comparisons
Please refer to Tables 1 and 2 for an overview of the group comparisons. Although subfield volumes tended to be smaller in CIND than in controls, there was no significant effect for group. There was also no significant effect for gender. In accordance with our previous study, there was a significant effect for Apo E4 carrier state on CA3&DG volume (p = 0.035; beta = 11.1) with Apo E4 carriers having smaller CA3&DG volumes than subjects without it (Apo E4 pos: 136.5 ± 22.4 mm3; Apo E4 neg: 151.7 ± 24.3 mm3, values are corrected for head size). In addition, there were significant effects for age on CA1 (p = 0.001, beta = −2.999) and CA3&DG (p = 0.042, beta = −1.393). Group had a significant effect on all three CVLTII scores with CIND subjects performing worse than control subjects (please also see Table 1, IFRD : p = 0.0022, beta = 0.22; SFRD: p = 0.04, beta = 0.25; DRD : p = 0.005, beta = 0.35). CVLTII scores were negatively influenced by age (IFRD : p = 0.014, beta = −0.025; SFRD: p = 0.011, beta = −0.044, DRD : p = 0.033, beta = −0.036) but not by years of education, gender or Apo E4 carrier state. Excluding subjects with an Apo E2 allele did not change these findings.
Table 2.
Mean and (SD) of Subfield Volumes and CVLTII Scores in Controls and CIND
| Controls | CIND | |||
|---|---|---|---|---|
| Non Apo E4 n = 18 | Apo E4 n = 7 | Non Apo E4 n = 14 | Apo E4 n = 11 | |
| Subfields | ||||
| ERC | 107.6 (27.9) | 97.5 (15.0) | 94.2 (20.8) | 95.1 (21.5) |
| Sub | 109.7 (19.8) | 105.4 (27.4) | 108.9 (18.2) | 107.2 (22.1) |
| CA1 | 208.5 (21.0) | 185.0 (26.9) | 194.6 (32.0) | 189.5 (48.0) |
| CA1-2 trans | 10.0 (1.2) | 10.6 (1.7) | 9.6 (1.5) | 9.7 (2.0) |
| CA3&DG | 151.2 (23.5) | 140.7 (19.6) | 152.4 (26.3) | 133.8 (24.6) |
| CVLTII | ||||
| IFRD | 2.3 (0.5) | 2.4 (0.6) | 1.7 (0.4)* | 1.8 (0.5)# |
| SFRD | 2.4 (0.8) | 2.4 (0.9) | 1.8 (0.8) | 1.7 (1.0) |
| DRD | 2.4 (0.8) | 2.5 (0.7) | 1.7 (0.6)* | 1.5 (1.1) # |
impaired CVLTII performance p<0.05 CIND without Apo E4 compared to controls without Apo E4,
impaired CVLTII performance p<0.05 CIND with Apo E4 compared to controls with Apo E4; ERC, entorhinal cortex, Sub, subiculum; CA1-2, CA1-2 transition zone; CA3&DG, CA3 and dentate gyrus; IFRD, Immediate free discrimination recall total score; SFRD, short free recall discrimination score; DRD, long free recall discrimination score. Subfield volumes are corrected for intracranial head size (cf methods for formula), units are mm3
3.2. Associations between CVLTII scores and hippocampal subfields
Table 3 shows the results of the Pearson correlation analyses. Based on these, ERC, Sub, CA1 and CA3&DG volumes were selected for the stepwise regression analysis. After forcing age and Apo E4 carrier state into the stepwise regression model only CA3&DG contributed significantly to the variability of IFDR (p = 0.0411, beta = 0.0065) and thus was included in the model based on specified inclusion criteria. Similarly, after including age and Apo E4 carrier state into the stepwise model for SFDR only CA3&DG was significantly associated with SFDR (p = 0.03, beta = 0.0112). Finally, only CA1 contributed significantly to the variability of DRD (p = 0.044, beta = 0.008) when age and Apo E4 carrier state were accounted for in the stepwise regression model for DRD. CA1 also explained most of the variation of “consolidation” (DRD controlled for IFRD performance) (p = 0.033, beta = 0.0048). Again, excluding subjects with an Apo E2 allele did not change these findings. Repeating the analysis without forcing age and Apo E4 first into the model had no influence on the subfield volumes selected by the stepwise regression analysis. Please see Figure 2 for the scatter plots of the associations.
Table 3.
Pearson Correlations Subfield Volumes with CVLTII scores
| Score | ERC | Sub | CA1 | CA1-2 | CA3&DG | |
|---|---|---|---|---|---|---|
| IFRD | r p |
0.17 ns |
0.26 0.03 |
0.31 0.01 |
0.12 ns |
0.35 0.006 |
| SFRD | r p |
0.29 0.02 |
0.31 0.01 |
0.41 0.002 |
0.16 ns |
0.38 0.003 |
| DRD | r p |
0.20 ns |
0.22 ns |
0.41 0.001 |
0.15 ns |
0.35 0.006 |
ERC, entorhinal cortex, Sub, subiculum; CA1-2, CA1-2 transition zone; CA3&DG, CA3 and dentate gyrus; IFRD, Immediate free discrimination recall total score; SFRD, short free recall discrimination score; DRD, long free recall discrimination score; r, correlation coefficient of Pearson correlation; p, one sided significance; ns, not significant
4. Discussion
There were two main findings in this study: 1. IFDR and SFDR which assess learning and early retrieval of verbal information were significantly associated with CA3&DG volume. 2. DRD, which measures aspects of consolidation and delayed retrieval of verbal information, was associated with CA1 volume. These findings are in accordance with our a priori hypotheses and demonstrate that subfield volumetry and clinical standard tests may provide similar insights into the functional specialization as it is gained by animal and fMRI studies using sophisticated memory paradigms to tease out specific aspects of the memory process.
The first main finding of this study was that the variability of the IFRD and SFRD performance was significantly determined by the CA3&DG volume even after accounting for effects of confounding variables. Animal and fMRI studies suggest that CA3 and DG are critically involved in pattern separation and pattern completion which play crucial roles in successful memory processing (Treves and Rolls, 1994; Bakker et al. 2008;Yassa et al., 2010). Pattern separation refers to the process which allows the DG to translate the dense, synaptically weak and complex neocortical activity which reaches the DG via the ERC from the different cortical areas active during a specific event, e.g. sitting in a room and listening to the CVLT word list read by the examiner, into different distinct, sparse and synaptically strong DG activations. The sparse signals generated in the dentate granule cells are then forwarded to the CA3 pyramidal cells via the mossy fiber pathway. A prominent feature of the CA3 pyramidal cells is their large number of recurrent collaterals onto other CA3 pyramidal cells (Ishizuka et al. 1990). It has been suggested that these recurrent collaterals allow CA3 to act as an autoassociative network which is able to form arbitrary associations between the different simultaneously incoming DG signals and thus to link information generated by different parts of the neocortex into a very specific hippocampal memory trace for this particular event (Rolls, 1996, 2007, 2010). The generation of this memory trace is associated with modifications of the recurrent collateral CA3 synapses. It not only represents a crucial step for the immediate storage of information (Nakazawa et al 2004, Moser et al.2003, Rolls 2007, Kesner and Hunsacker, 2010) but is also a prerequisite for pattern completion. Pattern completion allows for the restoration of the complete CA3 activation pattern generated during the initial processing from the repeated presentation of just a few of the original elements, i.e. ,from a noisy or partial ERC input activity (Acsady and Kali, 2007;Hasselmo, 2005, Rolls, 1996). Since IFRD and SFRD measure the ability to learn and recall a word list which has repeatedly been read just a few minutes before (about 1–5 Min), it can be expected that both tasks rely on the fast acquisition and retrieval of information and thus on the processes active during pattern separation and completion. Therefore the finding of a significant association between IFRD and SFRD and CA3&DG is in good agreement with the findings of animal and computational studies.
A previous study from this lab found significantly smaller CA3&DG volumes in elderly cognitively intact subjects and AD subjects with the Apo E4 allele compared to subjects without this allele (Mueller et al. 2008). This finding was replicated in this study in so far that regardless of their cognitive status subjects with the Apo E4 allele had a smaller CA3&DG volume than subjects without it. The subgroups, i.e. CIND with and without Apo E4, controls with and without Apo E4, showed the same trends but the findings were not significant due to the small sample sizes. IFRD and SFRD scores however were not different between subjects with and without the Apo E4 allele. The latter finding was unexpected because the Apo E4 allele not only reduces the age of onset in AD but is also associated with an earlier onset of age related memory decline in cognitively intact elderly subjects (Sando et al. 2008; Caselli et al 2009). Based on that, one would have expected Apo E4 carriers to have a more prominent memory deficit than age-matched non Apo E4 carriers. The most obvious explanation for our failure to detect such an Apo E4 effect on IFRD and SFRD (or DRD) is that our study population was too small and too heterogeneous. Another possible explanation however is suggested by fMRI studies which demonstrated a reduced CA3&DG activation during memory tasks in cognitively normal Apo E4 carriers (Suthana et al 2009) but an increased activation during similar tasks in prefrontal and parietal cortical areas (Bookheimer et al, 2000; Burggren et al. 2002). These findings suggests that at least in cognitively intact elderly Apo e4 carriers a potentially negative effect of Apo E4 on CA3&DG function and/or structure can to some degree be successfully compensated by a hyperactivation of cortical brain regions.
The second main finding of this study was that CA1 volume contributed significantly to the variance of DRD performance in a mixed group of cognitively intact and mildly impaired elderly subjects even after effects of confounding variables such as age, Apo E4 carrier state and initial IFRD performance had been accounted for. The synchronized activation patterns generated in the CA3 autoassociative network are forwarded to CA1 via the Schaffer collaterals. It has been suggested that CA1 assembles several of the smaller CA3 information elements into larger compounds and so allows for a more efficient representation of the information (Schultz and Rolls, 1999). In addition to the pre-processed information about the event which CA1 receives from the DG and CA3, it also receives also direct neocortical input about the same event via ERC and the perforant path. It assumed that CA1 is able to associate this ERC generated activity with the activity from CA3 (Hasselmo, 2005; Lee et al. 2005; Hunsaker and Kesner, 2008, Rolls, 2010). During recall, an incomplete cue about the original event can thus be restored in CA3 by pattern completion and be forwarded to CA1 where it not only reconstructs the CA3 associated CA1 activity pattern but also the ERC activity pattern linked to it (Treves and Rolls, 1994; Rolls, 2010). The thus reconstructed CA1 activity reactivates via back-projection neurons in the ERC which on their part re-activate those parts of the cerebral cortex which provided the original signal to the ERC/hippocampus. It has been shown in animal studies that in contrast to CA3 lesions which affect predominantly the retrieval of temporally more recent information, CA1 lesions mostly affect the retrieval of temporally more remote information, suggesting that CA1 is involved in late retrieval and consolidation and also in he temporal ordering of memory traces (Hasselmo 2005; Lee et al. 2005, Hunsaker and Kesner, 2008, Hunsaker et al. 2008, Daumas et al. 2005). DRD is a measure of how well previously encoded words on a list can be recalled after a longer interval (20–25 min) during which new, similar information had to be learned and new unrelated non-verbal tasks had to be performed. That means that DRD tests aspects of memory performance which based on the findings in animal studies can be expected to involve CA1 and thus the association between CA1 and DRD shown in this study is in agreement with the findings of those animal and computational studies.
Previous studies from this lab demonstrated a significant volume loss in the dorso-medial aspect of the hippocampus (CA1-2 transition zone) in aMCI compared to healthy controls (Mueller and Weiner, 2009). There were no significant hippocampal subfield volume losses in CIND compared to controls in this study. This discrepancy can be explained by the more heterogeneous population of cognitively impaired subjects and a reduced power due to the smaller group of healthy controls (25 vs. 53 in previous study) in this study. The failure to demonstrate significant subfield volume losses in CIND compared to controls however contrasts with the significantly worse performance of CIND on all memory scores incl. MMSE even though there were significant associations between some subfield volumes and these scores. There are two possible explanations for this discrepancy. 1. Despite its crucial contribution, the hippocampus is only one element of a widespread cortical and subcortical network of brain structures supporting memory processes which encompass prefrontal and anterior cingulate cortex, lateral temporal lobe, precuneus and parts of the parietal lobes and the thalamus. Functional studies indicate that different structures within these networks process different aspects of the memory process, e.g. are engaged in encoding but not retrieval or monitor the memory process and suppress intrusions (Desgranges et al. 1998). The functional specialization and complexity of these networks however suggest that accumulated damage throughout the network is probably a better predictor of the resulting impairment than the damage to a single structure within it. Although mesial temporal structures are most prominently affected in aMCI, volume losses have also been described in temporal lateral, parietal and frontal areas (Hämäläinen et al. 2007b; Karas et al. 2008), i.e., regions involved in memory processes. Based on these observations, it seems likely that the CIND subjects in this study also have additional extrahippocampal atrophic changes which explain their memory impairment despite their relatively intact hippocampal subfields. 2. Functional MRI studies assessing activation differences during cognitive paradigms in aMCI and controls have provided evidence for a functional compensation in aMCI. Particularly in the earliest stages, aMCI often either show the same but stronger activation pattern than controls, engage larger areas of the same structure, or activate additional structures (Dickerson and Sperling, 2008, Hämäläinen et al. 2007a). A recent fMRI study observed similar compensation mechanisms in ERC and CA3 and dentate in aMCI during memory tasks designed to test pattern separation (Yassa et al 2010). This indicates that subtle structural changes in the hippocampus might be associated with more severe functional hippocampal abnormalities which determine the memory impairment at this stage.
The study has several limitations. 1. The number of study subjects is rather small and the CIND group is heterogeneous regarding type of cognitive impairment which could have obscured structural/cognitive associations. 2. We cannot exclude that the structural–functional associations found in this study were driven by the underlying disease process, i.e. most likely early Alzheimer’s disease, in the cognitively impaired subjects and that different associations would have been found if subjects suffering from cognitive impairment due to a different process, e.g. vascular dementia, would have been chosen instead. 3. Although we used a stepwise regression analysis and thus a relatively conservative statistical approach, it is necessary to keep in mind that a significant association between structure and function does not necessarily also imply a causal relationship. 4. The CVLTII subtests used in this study represent complex memory tasks. Although it is reasonable to assume that they also rely on hippocampus specific functions such as pattern separation etc., their correct execution requires the participation of brain structures beyond the hippocampus. It will therefore be necessary to confirm the associations found in this study with memory tasks which specifically target functions such as pattern separation etc. 5. Functional studies suggest an anterior and posterior specialization of the hippocampus formation for memory processes. (Greicius et al, 2003; Giovanello et al. 2009). In this study subfield measurements were only obtained from the anterior third of the hippocampal body. Manual labeling is time consuming and thus the restricted labeling approach had been chosen for the sake of efficiency. Furthermore, labeling of the whole hippocampus would not have been possible since it was not completely covered in all subjects. The labeling of the anterior hippocampal body has the advantage that this section and the landmarks used for the labeling can be reliably identified in this region. However, the restricted labeling approach prevented us from investigating a possible anterior-posterior specialization. 6. We did not attempt to find evidence for material specific processing of the two hippocampi but rather combined the volumes from both sides. There were several reasons for this decision. a. Exploring such relationships properly requires that the language dominant hemisphere is known. This was not the case in this study. b. Most of the literature showing material-specific processing of the hemisphere comes from lesion studies suggesting that hemispheric material specialization might become most obvious if the network supporting the function has been massively altered. However, there were no systematic volume differences between left and right subfields neither in the study population as a whole nor in any of the subgroups. This raises the chances of finding false positive material specific associations and indeed when both sides were analyzed separately significant CVLT II-subfield correlations were found for the left and right side (cf Table 3 in supplementary material). Based on these observations functional studies seem to be better suited than structural studies to address this particular question in such a population.
In conclusion, in accordance with our a priori hypothesis, this study found an association between CA3&DG and a widely used clinical standard test assessing learning and early retrieval and an association between CA1 and a cognitive task assessing late retrieval and consolidation. This demonstrates that subfield volumetry has the potential to study non invasively and in vivo subfield specific memory functions and thus might provide similar information as animal studies or functional studies in humans. This might ultimately contribute to a better understanding of the hippocampal changes associated with memory decline in normal aging and a better distinction from those associated with the early stages of neurodegenerative diseases.
Supplementary Material
Acknowledgments
The authors would like to express their gratitude to Ms Jennifer Hlavin and Mr. Derek Flenniken who were relentlessly working on collecting all the clinical and neuropsychological data from the different referral sites and transferring it into the main database.
Funding: National Institutes of Health grants: RO1 AG010897 and P01 AG12435 to M.W.W.
Footnotes
Disclosure Statement: None of the authors has a conflict of interest to disclose.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Acsady L, Kali S. Models, structure, function: the transformation of cortical signals in the dentate gyrus. Prog Brain Res. 2007;163:577–599. doi: 10.1016/S0079-6123(07)63031-3. [DOI] [PubMed] [Google Scholar]
- Bakker A, Kirwan CB, Miller M, Stark CEL. Pattern separation in the human hippocampal CA3 and dentate gyrus. Science. 2008;319:1640–1642. doi: 10.1126/science.1152882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartsch T, Alfke K, Wolff Stingele R, Rohr A, Freitag-Wolf S, Jansen O, Deuschl G. Selective affection of hippocampal CA-1 neurons in patients with transient global amnesia without long-term sequelae. Brain. 2006;129:2874–2884. doi: 10.1093/brain/awl248. [DOI] [PubMed] [Google Scholar]
- Bayley PJ, Wixted JT, Hopkins RO, Squire LR. Yes/No recognition, forced-choice recognition and the human hippocampus. J Cogn Neurosci. 2008;20:505–512. doi: 10.1162/jocn.2008.20038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bird CM, Vargha-Khadem F, Burgess N. Impaired memory for scenes but not for faces in developmental hippocampal amnesia: A case study. Neuropsychologia. 2008;46:1050–1059. doi: 10.1016/j.neuropsychologia.2007.11.007. [DOI] [PubMed] [Google Scholar]
- Bobinski M, de Leon MJ, Tarnawski M, Wegiel J, Bobinski M, Reisberg B, Miller DC, Wisniewski HM. Neuronal and volume loss in CA1 of the hippocampal formation uniquely predict duration and severity of Alzheimer’s disease. Brain Res. 1998;805:267–269. doi: 10.1016/s0006-8993(98)00759-8. [DOI] [PubMed] [Google Scholar]
- Bookheimer SY, Stojwas MH, Cohen MS, Saunders AM, Pericak-Vance MA, Mazziotta JC, Small GW. Pattern of brain activation in people at risk for alzheimer’s disease. N Engl J Med. 2000;343:450–456. doi: 10.1056/NEJM200008173430701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burggren AC, Small GW, Sabb FW, Bookheimer SY. Specificity of brain activation patterns in people at genetic risk for Alzheimer Disease. Am J Geriatr Psychiatry. 2002;10:44–51. [PubMed] [Google Scholar]
- Caselli RJ, Dueck AC, Osborne D, Sabbagh MN, Connor DJ, Ahern GL, Baxter LC, Rapcsak SZ, Shi J, Woodruff BK, Locke DEC, Hoffman Snyder C, Alexander GE, Rademakers R, Reiman EM. Longitudinal modeling of age-related memory decline and the Apo ε 4 effect. N Engl J Med. 2009;361:255–263. doi: 10.1056/NEJMoa0809437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daumas S, Halley H, Frances B, Lassalle JM. Encoding, consolidation, and retrieval of contextual memory: differential involvement of dorsal CA3 and CA1 hippocampal subregions. Lern Mem. 2005;12:375–382. doi: 10.1101/lm.81905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delis DC, Kramer JH, Kaplan E, Ober BA. Adult Version Manual. 2. San Antonio, TX: Psychological Cooperation; 2000. California Verbal Learning Test. [Google Scholar]
- Delis DC, Wetter SR, Jacobson MW, Peavy G, Hamilton J, Gongvatana A, Kramer JH, Bondi MW, Corey-Bloom J, Salmon DP. Recall discriminability: Utility of a new CVLT-II measure in the differential diagnosis of dementia. J Int Neuropsychol Soc. 2005;11:708–715. doi: 10.1017/S1355617705050812. [DOI] [PubMed] [Google Scholar]
- Desgranges B, Baron JC, Eustache F. The functional neuroanatomy of episodic memory: The role of the frontal lobes, the hippocampal formation, and other areas. Neuroimage. 1998;8:198–213. doi: 10.1006/nimg.1998.0359. [DOI] [PubMed] [Google Scholar]
- Dickerson BC, Sperling RA. Functional abnormalities of the medial temporal lobe memory system in mild cognitive impairment and Alzheimer’s disease: Insights from functional MRI studies. Neuropsychologia. 2008;46:1624–1635. doi: 10.1016/j.neuropsychologia.2007.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duvernoy HM. Functional anatomy, Vascularization and serial sections with MRI. 3. Berlin, Heidelberg, New York: Springer-Verlag; 2008. The human hippocampus; p. 232. [Google Scholar]
- Eldridge LL, Engel SA, Zeineh MM, Bookheimer SY, Knowlton BJ. A dissociation of encoding and retrieval processes in the human hippocampus. J Neuroscience. 2005;25:3280–3286. doi: 10.1523/JNEUROSCI.3420-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson SH, Thom M, Batlett PA, Symms MR, McEvoy AW, Sisodiya SM, Duncan JS. Propeller MRI visualizes detailed pathology of hippocampal sclerosis. Epilepsia. 2008;49:33–39. doi: 10.1111/j.1528-1167.2007.01277.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giovanello KS, Schnyer D, Verfaellie M. Distinct hippocampal regions make unique contributions to relational memory. Hippocampus. 2009;19:111–117. doi: 10.1002/hipo.20491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius MD, Krasnow B, Boyett-Anderson JM, Eliez S, Schatzberg AF, Reiss AL, Menon V. Regional analysis of hippocampal activation during memory encoding and retrieval: fMRI study. Hippocampus. 2003;13:164–174. doi: 10.1002/hipo.10064. [DOI] [PubMed] [Google Scholar]
- Hämäläinen A, Pihlajamäki M, Tanila H, Hänninen T, Niskanen E, Tervo S, Karjalainen PA, Vanninen RL, Soininen H. Increased fMRI responses during encoding in mild cognitive impairment. Neurobiol Aging. 2007;28:1889–1903. doi: 10.1016/j.neurobiolaging.2006.08.008. [DOI] [PubMed] [Google Scholar]
- Hämäläinen A, Tervo S, Grau-Olivares M, Niskanen E, Pennanen C, Huuskonen J, Kivipelto M, Hänninen T, Tapiola M, Vanhanen M, Vanninen R, Soininen H. Voxel-based morphometry to detect brain atrophy in progressive mild cognitive impairment. Neuroimage. 2007;37:1122–1131. doi: 10.1016/j.neuroimage.2007.06.016. [DOI] [PubMed] [Google Scholar]
- Hasselmo ME. The role of hippocampal CA3 and CA1 in matching entorhinal input with retrieval of associations between objects and context: Theoretical comment on Lee et al. Behav Neurosci. 2005;119:342–345. doi: 10.1037/0735-7044.119.1.342. [DOI] [PubMed] [Google Scholar]
- Hunsaker MR, Kesner RP. Evaluating the differential roles of the dorsal dentate gyrus, dorsal CA3, and dorsal CA1 during a temporal ordering for spatial locations task. Hippocampus. 2008;18:955–964. doi: 10.1002/hipo.20455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunsaker MR, Fieldsted PM, Rosenberg JS, Kesner RP. Dissociating the roles of dorsal and ventral CA1 for the temporal processing of spatial locations, visual objects and odors. Behav Neuroci. 2008;122:643–650. doi: 10.1037/0735-7044.122.3.643. [DOI] [PubMed] [Google Scholar]
- Ishizuka N, Weber J, Amaral DG. Organization of intrahippocampal projections origination from CA3 pyramidal cells in the rat. J Comp Neurol. 1990;295:580–623. doi: 10.1002/cne.902950407. [DOI] [PubMed] [Google Scholar]
- Karas G, Sluimer J, Goekoop R, van der Flier W, Rombouts SARB, Vrenken H, Scheltens P, Fox N, Barkhof F. Amnestic mild cognitive impairment: Structural MR imaging findings predictive of conversion to Alzheimer’s disease. Am J Neuroradiol. 2008;29:944–949. doi: 10.3174/ajnr.A0949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kesner RP, Hunsaker MR. The temporal attributes of episodic memory. Behav Brain Res. 2010;215:299–309. doi: 10.1016/j.bbr.2009.12.029. [DOI] [PubMed] [Google Scholar]
- Kramer JH, Schuff N, Reed B, Mungas D, Du AT, Rosen HJ, Jagust WJ, Miller BL, Weiner MW, Chui H. Hippocampal volume and retention in Alzheimer’s disease. J Int Neurophysiol Soc. 2004;10(4):639–643. doi: 10.1017/S1355617704104050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirwan CB, Wixted JT, Squire LR. A demonstration that the hippocampus supports both recollection and familiarity. Proc Natl acad Sci USA. 2010;107:344–348. doi: 10.1073/pnas.0912543107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee I, Hunsaker MR, Kesner RP. The role of hippocampal subregions in detecting spatial novelty. Behav Neurosci. 2005;119:145–153. doi: 10.1037/0735-7044.119.1.145. [DOI] [PubMed] [Google Scholar]
- Manns JR, Hopkins RO, Reed JM, Kitchener EG, Squire LR. Recognition memory and the human hippocampus. Neuron. 2003;37:171–180. doi: 10.1016/s0896-6273(02)01147-9. [DOI] [PubMed] [Google Scholar]
- Moser EI, Moser MB. One shot memory in hippocampus. Neuron. 2003;38:147–148. doi: 10.1016/s0896-6273(03)00227-7. [DOI] [PubMed] [Google Scholar]
- Mueller SG, Stables L, Du AT, Schuff N, Truran D, Cashdollar N, Weiner MW. Measurements of hippocampal subfields and age related changes with high resolution MRI at 4T. Neurobiol Aging. 2007;28:719–726. doi: 10.1016/j.neurobiolaging.2006.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller SG, Schuff N, Raptentsetsang S, Elman J, Weiner MW. Selective effect of Apo e4 on CA3 and dentate in normal aging and Alzheimer’s disease using high resolution MRI at 4T. Neuroimage. 2008;42:42–48. doi: 10.1016/j.neuroimage.2008.04.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller SG, Weiner MW. Selective effect of age, Apo e4 and Alzheimer’s Disease on hippocampal subfields. Hippocampus. 2009;19:558–564. doi: 10.1002/hipo.20614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller SG, Schuff N, Yaffe K, Madison C, Miller B, Weiner MW. Hippocampal atrophy patterns in mild cognitive impairment and Alzheimer’ Disease. Human Brain Mapping. 2010;31:1339–1347. doi: 10.1002/hbm.20934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakazawa K, McHugh TJ, Wilson MA, Tonegawa S. NMDA receptors , place cells and hippocampal spatial memory. Nature Reviews. 2004;5:361–370. doi: 10.1038/nrn1385. [DOI] [PubMed] [Google Scholar]
- O’Reilly CR, Rudy JW. Conjunctive representation in learning and memory: Principles of cortical and hippocampal function. Psychol Rev. 2001;108:311–345. doi: 10.1037/0033-295x.108.2.311. [DOI] [PubMed] [Google Scholar]
- Petersen RC, Smith GE, Waring SC, Ivnik RJ, Tangalos EG, Kokmen E. Mild cognitive impairment, clinical characterization and outcome. Arch Neurol. 1999;56:303–308. doi: 10.1001/archneur.56.3.303. [DOI] [PubMed] [Google Scholar]
- Quinette P, Guillery-Girard B, Noel A, de la Sayette V, Viader F, Desgranges B, Eustache F. The relationship between working memory and episodic memory disorders in transient global amnesia. Neuropsychologia. 2006;44:2508–2519. doi: 10.1016/j.neuropsychologia.2006.03.031. [DOI] [PubMed] [Google Scholar]
- Rolls ET. A theory of hippocampal function in memory. Hippocampus. 1996;6:601–620. doi: 10.1002/(SICI)1098-1063(1996)6:6<601::AID-HIPO5>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- Rolls ET. An attractor network in the hippocampus: Theory and neurophysiology. Learn Mem. 2007;14:714–731. doi: 10.1101/lm.631207. [DOI] [PubMed] [Google Scholar]
- Rolls ET. A computational theory of episodic memory formation in the hippocampus. Behav Brain Res. 2010;215:180–196. doi: 10.1016/j.bbr.2010.03.027. [DOI] [PubMed] [Google Scholar]
- Rolls ET, Kesner RP. A computational theory of hippocampal function and empirical test of the theory. Prog Neurobiol. 2006;79:1–48. doi: 10.1016/j.pneurobio.2006.04.005. [DOI] [PubMed] [Google Scholar]
- Rössler M, Zarski M, Bohl J, Ohm TG. Stage dependent and sector specific neuronal loss in hippocampus during Alzheimer’s disease. Acta Neuropath. 2002;103:363–369. doi: 10.1007/s00401-001-0475-7. [DOI] [PubMed] [Google Scholar]
- Sando SB, Malquist S, Cannon A, Hutton ML, Sletvold O, Saltverldt I, White LR, Lydersen S, Aasly JO. APOE ε 4 lowers age at onset and is a high risk factor for alzheimer’s disease; A case control study from central Norway. BMC Neurology. 2008;8:9. doi: 10.1186/1471-2377-8-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz SR, Rolls ET. Analysis of information transmission in the Schaffer collaterals. Hippocampus. 1999;9:582–598. doi: 10.1002/(SICI)1098-1063(1999)9:5<582::AID-HIPO12>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- Schoenheit B, Zarski R, Ohm TG. Spatial and temporal relationships between plaques and tangles in alzheimer-pathology. Neurbiol Aging. 2004;25:897–711. doi: 10.1016/j.neurobiolaging.2003.09.009. [DOI] [PubMed] [Google Scholar]
- Scoville WB, Milner B. Loss of recent memory after bilateral hippocampal lesions. J Neurol Neursurg Psychiatry. 1957;20:11–21. doi: 10.1136/jnnp.20.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suthana NA, Ekstrom AD, Moshirvaziri S, Knowlton B, Bookheimer SY. Human hippocampal CA1 involvement during allocentric encoding of spatial information. J Neuroscience. 2009;29:10512–10519. doi: 10.1523/JNEUROSCI.0621-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suthana NA, Krupa A, Donix M, Burggren A, Ekstrom AD, Jones M, Ercoli LM, Miller KJ, Siddarth P, Small GW, Bookheimer SY. Reduces hippocampal CA2, CA3 and dentate gyrus activity in asymptomatic people at genetic risk for Alzheimer’s disease. Neuroimage. 2009 doi: 10.1016/j.neuroimage.2009.12.014. e pub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Treves A, Rolls ET. Computational analysis of the role of the hippocampus in memory. Hippocampus. 1994;4:374–391. doi: 10.1002/hipo.450040319. [DOI] [PubMed] [Google Scholar]
- Wan H, Aggleton JP, Brown MW. Different contributions of the hippocampus and perirhinal cortex to recognition memory. Journal Neurosci. 1999;19:1142–1148. doi: 10.1523/JNEUROSCI.19-03-01142.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West MJ, Coleman PD, Flood DG, Troncoso JC. Differences in the pattern of hippocampal neuronal loss in normal ageing and Alzheimer’s disease. Lancet. 1994;344:769–772. doi: 10.1016/s0140-6736(94)92338-8. [DOI] [PubMed] [Google Scholar]
- Yassa MA, Stark SM, Bakker A, Albert MS, Gallagher M, Stark CEL. High resolution structural and functional MRI of hippocampal CA3 and dentate gyrus in patients with amnestic mild cognitive impairment. Neuroimage. 2010;2010 doi: 10.1016/j.neuroimage.2010.03.040. epub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.