

Abstract
Three experiments used postclass formation within-class preference test performances to evaluate the effects of nodal distance on the relatedness of stimuli in equivalence classes. In Experiment 1, two 2-node four-member equivalence classes were established using the simultaneous protocol in which all of the baseline relations were trained together, after which all emergent relations probes were presented together. All training and testing was done using match-to-sample trials that contained two comparisons. After class formation, the effects of nodal distance were evaluated using within-class preference tests that contained samples and both comparisons from the same class. These tests yielded inconsistent performances for most participants. Experiment 2 replicated Experiment 1, but a third null comparison was used on all trials during class formation. Thereafter, virtually all of the within-class probes, for all participants, evoked performances that were consistent with the predicted effects of nodal distance, that is, the selection of comparisons that were nodally closer to the samples. It appears, then, that the establishment of the equivalence classes with a third null comparison induced control by nodal structure of the classes. Experiment 3 demonstrated the generality of these findings with larger classes that contained more nodal separations, that is, three-node five-member classes. Emergent-relations tests conducted immediately after the within-class tests showed the classes to be intact. Thus, the differential relatedness of stimuli in a class or their interchangeability depended on the content of a test trial: within-class probes occasioned responding indicative of differential strength among the stimuli in the class, while cross-class tests occasioned responding indicative of interchangeability of stimuli in the same class.
Keywords: equivalence class, nodal structure, nodal distance, within-class preference, matching-to-sample, college students
An equivalence class contains a finite number of physically disparate stimuli all of which become related to each other after the training of some conditional discriminations among the stimuli. Assuming the stimuli in a potential class are represented by the letters, A, B, C, D and E, a five-member equivalence class can be established by training four baseline relations such as AB, BC, CD, and DE. The formation of such a class would then be documented when the presentation of one stimulus in the set occasions the selection of any other stimulus in the set (Fields & Verhave, 1987). This can be accomplished with the presentation of the untrained probes BA, CB, DC, ED, AC, AD, AE, BD, BE, CE, CA, DA, EA, DB, EB, and EC (Fields & Verhave, 1987; Sidman, 1994; Sidman & Tailby, 1982). If two classes are being established, each probe would contain a sample stimulus from one class and at least two comparison stimuli, one from each class. The selection of the comparsion from the same class as the sample would indicate control of behavior by class membership. That sort of selection by all probes would document the formation of the equivalence classes. Such a pattern of conditional selection would also indicate that the stimuli in the class are functionally interchangeable or substitutable for each other (Sidman, 1994). The substitutability of the stimuli documented by these tests also implies that the members of an equivalence class are equally related to each other (Fields & Verhave, 1987). Of note, these results are obtained from trials that contain stimuli in the same and different classes.
An alternative view is that stimuli in an equivalence class are not equally related to each other (Fields & Moss, 2007). This notion has its roots in the findings described in experiments that have explored the learning of derived lists (Ebbinghaus, 1913), serial learning (Slamecka, 1985), and semantic memory networks (Collins & Loftus, 1975; Collins & Quillian, 1969). In all of these cases, the stimuli in a list or network were separated by a variable number of nodal stimuli and the likelihoods of responding were inversely related to the number of nodes that separated the stimuli in the lists or the networks. In all of these experiments, differential responding was occasioned by relations between stimuli in the same class.
These results led Fields and Verhave (1987) to opine that under some conditions, the relatedness of stimuli in the same equivalence class should also be an inverse function of the nodal distance that separates the stimuli in an equivalence class. In the example above, the training of AB, BC, CD, and DE would lead to the establishment of a five-member equivalence class with a nodal structure represented as A→B→C→D→E (Fields, Adams, & Verhave, 1993a; Fields & Verhave, 1987). In this structure some stimuli are related to other stimuli by training to a common stimulus. A node is defined as any stimulus that links at least two other stimuli in an equivalence class. Furthermore, nodal distance is defined as the number of nodes that link two stimuli within an equivalence class (Fields & Verhave, 1987). Thus, one node (C) separates B and D while three nodes (B, C, D) separate A and E. According to the associative distance view of relatedness of stimuli in equivalence classes, because fewer nodes separate the stimuli in BD than AE, the relational strength of BD should be greater than that of AE, and likewise for other relations with different nodal spreads.
These predictions have been supported using performances obtained under a range of testing conditions. Specifically, when the delayed emergence of equivalence classes was demonstrated, during the initial test block, the degree of class-consistent responding produced by derived-relations probes was an inverse function of the nodal distance that separated the stimuli in each derived relation (Bentall, Jones, & Dickins, 1998; Fields, Adams, Verhave, & Newman, 1990; Kennedy, 1991; Kennedy, Itkonen, & Lindquist, 1994; Sidman, Kirk, & Willson-Morris, 1985; Spencer & Chase, 1996). In addition, the order in which the derived relations reached criterion was a direct function of nodal distance (Fields et al., 1990; Kennedy et al., 1994). Furthermore, latency occasioned by derived relations was a direct function of nodal distance (Bentall et al.,1998; Kennedy et al., 1994; Spencer & Chase, 1996; Tomanari, Sidman, Rubio, & Dube, 2006; Wulfert & Hayes, 1988). In another study, Fields, Adams, Verhave, and Newman (1993b) first established three-node five-member equivalence classes. They then trained a response to one of the class members, and finally presented the remaining class members in repeated test blocks until transfer of responding was complete. For some participants, that response generalized gradually to the other class members. For these participants, on the first test block, the degree of generalization was an inverse function of nodal distance that separated the training stimulus from the other class members. With repeated testing, the accuracy of responding increased to 100% in an order that was an inverse function of nodal distance.
In these studies, the effects of nodal distance that were observed during the delayed emergence disappeared once the classes were fully formed. This could mean that the effects of nodal distance could have been transient, disappearing permanently after the formation of the equivalence classes. Alternatively, the effect of nodal distance could have been permanent, but no longer influenced the performances occasioned by the prevailing test conditions. In such a case, nodal distance should still influence responding in postclass formation tests, a position supported by the results of the following experiments.
Before describing these studies, it is important to note two potential factors that were confounded with nodal distance in the previously mentioned studies. The baseline relations were trained in a serial manner that corresponded to the nodal separation of the stimuli in the classes. In addition, due to serialized training, the number of presentations of each baseline relation was also correlated with nodal distance. Thus, the effects attributed to nodal distance might have been driven by either of these factors alone or in combination. These problems were avoided in the next two experiments by the establishment of equivalence classes using the simultaneous protocol, a procedure in which all baseline relations were trained on a concurrent basis (i.e., in the same blocks), were presented an equal number of times, and all derived-relations probes were introduced on a concurrent basis.
Dual-Option Function Transfer Tests
Fields, Landon-Jimenez, Buffington, and Adams (1995) used the simultaneous protocol (SIM) to establish two 5-member equivalence classes by training AB, BC, CD, and DE. Equivalence classes were formed by only 2 of the 12 participants. After class formation, they were trained to emit different and incompatible responses in the presence of the A stimulus and the E stimulus in the same class. Finally, all class members were then presented separately and without feedback. Under those conditions, the frequency with which the A-based response was occasioned by the B, C, and D stimuli was an inverse function of nodal distance. Similarly, the frequency with which the E-based response was occasioned by the D, C, and B stimuli was an inverse function of nodal distance. This pattern of responding remained stable with test repetition, and thus provided a postclass formation steady-state measure of the effects of nodal distance on responding. These performances could have occurred only if nodal structure was the primary determinant of relational strength among the stimuli in the parent equivalence classes.
Fields and Watanabe-Rose (2008) extended these findings by the establishment of two 4-node six-member equivalence classes through the training of AB, BC, CD, DE, and EF under the simultaneous protocol. After testing for the emergence of all derived relations, different responses were trained to the C and D stimuli of each class using a successive discrimination procedure. Finally, all 12 stimuli in the two classes were presented singly, many times, and in randomized order with no informative feedback. Only 4 of 15 participants in the experiment formed the equivalence classes. For both classes for 2 participants and one class for 1 participant, the response trained to the C stimulus in a class, then, generalized essentially completely to the A and B stimuli but rarely to the D, E, and F stimuli in that class. Likewise, the response trained to the D stimulus generalized essentially completely to the E and F stimuli but rarely to the A, B, and C stimuli. Therefore, the two 6-member equivalence classes were bifurcated into two 3-member equivalence classes each: A1-B1-C1, A2-B2-C2, D1-E1-F1, and D2-E2-F2 as predicted by nodal structure. These results could have occurred only if the stimuli in each equivalence class were related to each other on a permanent basis and the relational strength among the class members was a function of nodal distance. Results for one class for 1 participant and for both classes for another participant, however, did not demonstrate bifurcation by nodal structure. These results indicate that nodal structure was not the only determinant of responding in the dual-option transfer tests.
To summarize, both of these studies established equivalence classes using the simultaneous protocol, which eliminated the confounding of the effects of serial order of presentation and number of presentations of stimuli with nodal distance. On the negative side, however, the use of the simultaneous protocol reduced the percentage of participants who formed classes and were then able to provide data that could be used to evaluate the differential relatedness of stimuli in the equivalence classes.
Within-Class Preference Tests
Within-class preference testing is another method for showing the permanent postclass formation effects of nodal distance. Presented in a matching-to-sample format, these tests permit direct comparisons of the relative strengths of any two relations in an equivalence class that share the same sample stimulus. Fields, Adams, and Verhave (1989; as cited in Fields et al., 1993b) established two 3-node five-member classes by training AB, BC, CD, and DE. The formation of two equivalence classes with a nodal structure represented as A→B→C→D→E was documented by class-consistent responding on the derived relations probes. Thereafter, within-class preference tests were presented in a match-to-sample format in which each trial contained one sample and two comparisons from the same class. For example, in a BDE test for class 1, the B1 stimulus was presented as a sample with the D1 and E1 stimuli as comparisons. The relation between B1 and D1 constituted a one-node transitive relation, while B1 and E1 constituted a two-node transitive relation. The many trials that were presented evaluated different nodal spreads between each comparison and the sample on that trial. Participants virtually always selected the comparison that was nodally closer to the prevailing sample. Thus, in an ABC probe, B was preferred to C, in an ACD probe, C was preferred to D, in an ACE probe, C was preferred to E, etc. These data then demonstrated that preferences were an inverse function of nodal distance that separated the stimuli in the structure of the class; that is, the differential relatedness of stimuli in an equivalence class was determined by nodal structure.
In the above study, the equivalence classes were formed using the simple-to-complex (STC) protocol (Fields, Reeve, Rosen, Varelas, Adams, Belanich, & Hobbie, 1997). This protocol involves the systematic presentation of training and testing trials. Specifically, the baseline relations were established in a serial order that was completely correlated with the nodal distances that separated the stimuli in a class. In addition, the number of training trials presented during class formation was also correlated with nodal distance. Thus, the results of the within-class preference tests, which were attributed to nodal distance, could also have reflected serial order and/or the number of trials used to establish each baseline relation. Alligood and Chase (2007) addressed the issue of unequal number of training trials by the establishment of four-node six-member equivalence classes where the number of training trials was held constant for each of the baseline relations. The postclass formation within-class preference tests typically occasioned the selection of nodally proximal stimuli which supported the view that the relational strengths among stimuli in an equivalence class are inversely related to nodal distance. As with Fields et al. (1989; as cited in Fields et al., 1993b), however, the equivalence classes were established using a serialized training and testing protocol. Thus, the results reported by Alligood and Chase (2007) could still have reflected the effects of serial order rather than nodal distance. To summarize, there is still no measure of the unconfounded effects of nodal distance on the performances evoked by postclass formation within-class preference tests.
EXPERIMENT 1
As noted earlier, the establishment of equivalence classes under the simultaneous protocol eliminates the confounding effects of nodal distance and the serially ordered training of baseline relations. In this experiment, participants attempted to form equivalence classes using the simultaneous protocol before assessing the effects of nodal distance using postclass formation within-class preference tests. When, however, equivalence classes are established using the simultaneous protocol, only a small percentage of participants form classes (Buffington, Fields, & Adams, 1997; Fields et al., 1997; Fields et al., 1995). Thus, any effects of nodal distance might be limited to the small percentage of individuals who can form equivalence classes under the simultaneous protocol. The issue of yield and thus representativeness of outcome was addressed by the incorporation of a yield-enhancing procedure described by Fields et al. (1997). Specifically, they increased the percentage of participants who formed equivalence classes under the simultaneous protocol (yield) by having participants learn other equivalence classes using the simple-to-complex protocol before attempting to learn new equivalence classes under the simultaneous protocol. A similar approach was used in the present experiment to increase the percentage of participants who would form new equivalence classes under the simultaneous protocol. Thus, the effects of nodal distance could be evaluated with a representative sample of participants drawn from the general population.
Method
Participants
The participants were 10 undergraduate students enrolled in a course in Introductory Psychology at Queens College/CUNY. Participation in the experiment fulfilled one of the course requirements. All students read and signed the informed consent statement before participating in the experiment, which was conducted in one 2.5- to 3.5-hr session.
Apparatus
Hardware and software
The experiment was conducted with an IBM-compatible computer that displayed all stimuli on a 15-inch VGA monitor. Responses consisted of touching specific keys on a standard keyboard. The experiment was controlled by custom software that programmed all stimulus presentations and recorded all keyboard responses.
Stimuli
Ten nonsense syllables were used during preliminary training. In class 1, they were BAK (U1), NUC (V1), WEX (W1), HUZ (X1), SIV (Y1); in class 2 the stimuli were REJ (U2), TUD (V2), GIP (W2), DIH (X2), and MEL (Y2). Eight nonsense syllables were the members of the equivalence classes formed under the simultaneous protocol. In class 1, they were QIJ (A1), TUW (B1), COH (C1), MEP (D1); in class 2, they were VIF (A2), KUY (B2), XOL (C2), and GEZ (D2). All stimuli were presented on a computer monitor as black letters centered in white 5-cm × 5-cm squares with black borders.
Procedure
Trial format and contingencies
All trials in the experiment were presented in a simultaneous matching-to-sample format (Cumming & Berryman, 1965). Each trial involved the presentation of a sample and two comparison stimuli. The samples were drawn from one of two sets. The positive comparison (Co+) was from the same set as the sample on that trial and the negative comparison (Co−) was from the other set. The sample was presented on the upper portion of the monitor and was centered horizontally. The positive and negative comparisons were presented below the sample and to the left and right of the sample. The upper edges of the comparisons were below the lower edge of the sample. The right edge of the left comparison was to the left of the left edge of the sample, and the left edge of the right comparison was to the right of the right edge of the sample. The location of the positive comparison was randomly assigned with the stipulation that it was presented the same number of times on the left and on the right in a block of trials.
Trial block organization and feedback reduction
Each phase of the experiment consisted of blocks of trials. In all phases, the trials in a block were presented in a randomized order without replacement. A trial began when “Press ENTER” appeared on the screen. Pressing the ENTER key cleared the screen and a sample stimulus was displayed. Pressing the space bar added the two comparison stimuli to the display. During a trial, the comparisons on the left and the right were selected by pressing the 1 or 2 key, respectively. A comparison selection cleared the screen and immediately displayed a feedback message centered on the screen.
When informative feedback was presented, a “RIGHT” or “WRONG” message appeared on the monitor, and depended on the accuracy of the comparison selection. The message remained on the screen until the participant pressed the R key (R for RIGHT) in the presence of the word “Right” or the W key (W for WRONG) in the presence of the word “Wrong.” These responses demonstrated that the participant discriminated the feedback information. During some training and all testing trials, uninformative feedback was presented after a comparison selection. On these occasions a dashed line that bracketed the letter E (i.e., - - E - -) appeared on the monitor and remained on until the participant pressed the E key, which served as an observing response for the uninformative feedback. The occurrence of an appropriate R, W, or E response cleared the monitor, ended the trial, and enabled the start of the next trial (Fields et al. 1995).
At the start of training, all of the trials in a block resulted in the presentation of informative feedback after each comparison selection, that is, 100% feedback. The block was presented repeatedly with 100% feedback until all trials occasioned the selection of correct comparisons, that is, produced 100% correct responding which was the mastery criterion. Thereafter, the percentage of trials in a block that produced informative feedback was reduced to 75%, 25%, and finally to 0% as long as the mastery criterion was maintained in a block. During feedback reduction, a random generator determined the trials that produced informative feedback. Each block ended with the on-screen message, “Press ENTER to begin the next block.” If 100% correct responding was not achieved within three blocks at a given feedback level during training, the participant was returned to the previous feedback level during the next block.
Experimental Phases
Phase 1: Instructions and keyboard familiarization
The experiment began with the presentation of the following on-screen instructions:
Thank you for volunteering to participate in this experiment. PLEASE DO NOT TOUCH ANY OF THE KEYS ON THE KEYBOARD YET! In this experiment you will be presented with many trials. Each trial contains three or four CUES. These will be familiar and unfamiliar picture images. YOUR TASK IS TO DISCOVER HOW TO RESPOND CORRECTLY TO THE CUES. Initially, there will also be INSTRUCTIONS that tell you how to respond to the cues, and LABELS that will help you to identify the cues on the screen. The labels and the instructions that tell you which KEYS to press will slowly disappear. Your task will be to RESPOND CORRECTLY to the CUES and the INSTRUCTIONS by pressing certain keys on the computer's keyboard. The experiment is conducted in phases. When each phase ends, the screen will sometimes tell you how you did. If you want to take a break at any time, please call the experimenter. PRESS THE SPACEBAR TO CONTINUE.
After pressing the space bar, participants were trained to emit the appropriate keyboard responses to complete a trial. This phase involved the repeated presentation of a block of 16 trials. Trials contained three English words, such as KING, QUEEN, and CAMEL. The semantic relation between the sample word (e.g., KING) and one of the comparisons (e.g., QUEEN) was used to prompt the selection of the correct comparison. The words RIGHT or WRONG followed each comparison selection. Correct responding to the stimuli in a trial was facilitated by the presentation of instructional prompts (e.g., “Make your choice by pressing 1 or 2”, “Press R to continue”, “Press W to continue”, or “Press E to continue”) which was systematically deleted across trials as long as the participant made correct responses. For further details, see Fields et al. (1990) or Fields et al. (1997). Phase 1 ended when the sample and comparison stimuli were presented without prompts, and performance was at 100% accuracy during a single block.
Phase 2: Preliminary training using the simple-to-complex protocol
Preliminary training involved the formation of two 3-node five-member equivalence classes using a variation of a simple-to-complex protocol which is detailed in Table 1. In this protocol, the serial establishment of the baseline relations was interleaved with tests to assess the emergence of the new relations that could be derived from the already documented baseline relations. After the acquisition of each baseline relation, participants were presented with two blocks of overtraining. If responding in a test block was less than 87% accurate, the block was repeated with informative feedback presented on all trials and at the end of the test block, until responding on that block met the above mentioned mastery criterion. If Phase 2 was completed within 2 hr, the participant advanced to Phase 3. Participants who spent more than 2 hr in Phase 2 were excused from the experiment.
Table 1.
Training and testing trials using the simple-to-complex protocol to train two 3-node five-member equivalence classes. Sa = sample; Co+ = positive comparison; Co− = negative comparison.

Phase 3: Equivalence class formation under the simultaneous protocol
The aim of this phase was to establish two new four-member equivalence classes with an A→B→C→D nodal structure. The protocol began with the concurrent training of the baseline conditional discriminations for Class 1 (A1–B1, B1–C1, and C1–D1) and Class 2 (A2–B2, B2–C2, and C2–D2). During each training block, each sample stimulus was presented on two trials (the positive comparison appeared on the left on one trial and on the right on the other trial). The training block was repeated until one block produced 100% accuracy. Thereafter, feedback was reduced until the baseline relations were maintained in the absence of feedback. Once all baseline relations were established and maintained with no informative feedback, the participants were presented with an emergent relations test that contained all of the baseline, symmetrical, one- and two-node transitive and equivalence relations probes. Four trials were presented for each relation; two trials for the relation from Class 1 and two from Class 2 in which the position of the Co+ was on the right for one trial and the left for the other trial. All of the trials that made up the emergent relations test were spread across four test blocks, 12 trials per test block, for a total of 48 trials. Each block consisted of one trial for each relational type so that each of the four trials mentioned above occurred one time across all four blocks. Within a block all trials were presented in a randomized order without replacement. This test (combination of four blocks) was repeated up to four times or until the participant performed with at least 92% accuracy on each of the four blocks.
Phase 4: Within-class preference tests
Once the participant passed the emergent relations test, the effects of nodal distance were assessed. The differential strength of relations in the two-node four-member equivalence classes was measured using within-class preference tests that involved the presentation of trials that contained a sample and two comparisons from the same class. The comparisons differed in terms of the nodal distance that separated each from the sample. Each combination of a specific sample and two comparisons will be called a probe type. Each probe type was designated with three letters, the first of which represents the sample, the second of which designates the comparison that was nodally proximal to the sample, and the third letter designates the comparison that was nodally distal to the sample. For example, a probe type designated as ACD contained A as the sample with C and D as the comparisons, where C and D were one and two nodes removed from the sample stimulus, respectively.
The within-class preference tests consisted of the presentation of four within-class probe blocks. Each probe block consisted of the presentation of two probe types which are referred to as a probe pair. The same probe pair was presented in a block for both Class 1 and Class 2. Trials for each probe type for a given class were presented four times for a total of 16 trials per test block. Trials for all four probe types were presented in a randomized sequence in the same test block.
The four within-class test blocks that were presented to assess the effects of nodal distance on preference among the members of each of the two-node four-member classes are presented in Table 2. This table lists the within-class probe pairs in their order of presentation: ABD/DCA, ACD/ABC, DCB/ABC, and DBA/DCB. Each within-class probe block was presented up to two times. If the participant selected the nodally proximal comparisons on at least 94% of the trials in the first test block, the block was not repeated. Performances that did not meet this criterion resulted in a second presentation of the same test block.
Table 2.
Within-class preference probes for Experiments 1 and 2. The first letter (Sa) depicts the sample in the trial, the second letter (CoP) depicts the nodally proximal stimulus, and the third letter (CoD) depicts the nodally distal stimulus for that probe. Under TYPES the left and right columns depict relations between the sample and nodally proximal comparison and the sample and nodally distal comparison, respectively. The number indicates the nodal distance between the sample and comparison, while the letter indicates the relational type (i.e. BL is baseline, S is symmetry, T is transitivity, and E is equivalence).

Although there were many probe types that could have been used in the within-class preference test, we included those listed in Table 2 for three reasons. First, we were working under a time constraint that limited a participant's involvement in the experiment. Second, we were concerned that the presentation of the totality of the within-class probe types would result in a degrading in performance. Third, some of the probe types evaluated the same nodal spreads as other probe types. With these constraints, we selected the probe types listed in Table 2 because they represented a broad range of nodal spreads.
In each probe pair, one of the probe types was of primary interest and the second probe type was included so that the performances produced by both probe types would be maximally interpretable. Each probe block is described next with a listing of all possible outcomes and interpretations.
The first test block contained ABD and DCA probes. In both of these probes, maximal nodal spreads separated the two comparisons from the sample stimuli. If participants did not respond in accordance with nodal proximity on these probes, it would be unlikely that the participant would respond in accordance with nodal proximity for lesser nodal spreads. This first block contained ABD probes which pitted a 0-node baseline relation (AB) against a two-node transitive relation (AD) and the DCA probes which pitted a 0-node symmetrical relation (DC) against a two-node equivalence relation (DA). The selection of B in the ABD probe and C in the DCA probe would demonstrate conditional control of comparison selection based on nodal proximity.
The second test block contained ACD and ABC probes. The ACD probes pitted a one-node transitive relation (AC) against a two-node transitive relation (AD). The ABC probes pitted a 0-node baseline relation (AB) against a one-node transitive relation (AC). Both probes contained the same sample, while the C comparison acted as the nodally proximal stimulus on one trial and the nodally distal stimulus on the other. The ACD probe was of primary interest because it pitted two relations of the same type (transitive) against each other where the only difference was in terms of nodal spread. The selection of the C stimulus in the ACD probe could reflect control by nodal proximity or possibly an unconditional preference for the C stimulus. The ABC probe was used as a foil to produce comparison selections that would disambiguate the source of control that was the determinant of responding in the ACD probe. In the ABC probe, the C stimulus was nodally distal to the sample. If C was selected in the ABC probe, it would imply that the selection of C in the ACD probe reflected control by simple preference to the C stimulus rather than control by nodal proximity. On the other hand, the selection of B in the ABC probe would imply that C was not simply a preferred stimulus. Thus, the selection of C in the ACD probe would have to be based on nodal proximity.
The third test block consisted of the DCB and ABC probes. The DCB probes pitted a 0-node symmetrical relation (DC) against a one-node equivalence relation (DB). The ABC probes pitted a 0-node baseline relation (AB) against a one-node transitive relation (AC). Both of these contain the same stimuli in the comparison set (B and C). Thus, each comparison is nodally proximal on one probe and is nodally distal on the other probe. The selection of C in the DCB probes and B on the ABC probes would demonstrate conditional control of comparison selection based on nodal proximity, and would rule out control by an unconditional preference for either comparison. In contrast, the uniform selection of the B comparison in both probes would demonstrate a simple unconditional preference for B, and likewise for the uniform selection of the C comparison.
The fourth test block contained DBA and DCB probes. The DBA probes pitted a one-node equivalence relation (DB) against a two-node equivalence relation (DA). The DCB probes pitted a 0-node symmetrical relation (DC) against a one-node equivalence relation (DB). Both probes contained the same sample, while the B comparison acted as the nodally proximal stimulus on one probe and the nodally distal stimulus on the other. The DBA probe was of primary interest because it pitted two relations of the same type (equivalence) against each other where the only difference was in terms of nodal spread. The selection of the B stimulus in the DBA probe could reflect control by nodal proximity or possibly an unconditional preference for the B stimulus. The DCB probe was used as a foil to produce comparison selections that would disambiguate the source of control that was the determinant of responding in the DBA probe. In the DCB probe, the B stimulus was nodally distal to the sample. If B was selected in the DCB probe, it would imply that the selection of B in the DBA probe reflected control by simple preference for the B stimulus rather than control by nodal proximity. On the other hand, the selection of C in the DCB probe would imply that B was not simply a preferred stimulus. Thus, the selection of B in the DBA probe would have to be based on nodal proximity.
Phase 5: Emergent relations test
The maintenance of the two equivalence classes was evaluated with the re-administration of the emergent relations test described in Phase 3.
Results and Discussion
Equivalence Class Formation under the Simple-to-Complex Protocol
Of the 10 participants who began Experiment 1, 8 formed two 3-node five-member equivalence classes during preliminary training under the simple-to-complex (STC) protocol. The 2 remaining participants formed four-member classes during preliminary training but did not have enough time to complete the entire experiment.
Equivalence Class Formation under the Simultaneous Protocol
After forming the two 5-member classes under the STC protocol, the simultaneous protocol was used to establish two new two-node four-member equivalence classes for the 8 remaining participants. All 8 participants acquired the baseline relations, and 7 passed the emergent relations tests which documented the formation of two 2-node four-member equivalence classes.
Within-Class Probes
Figure 1 depicts the data for each of the within-class preference probe types. The probe blocks are listed on the abscissa from left to right in their order of presentation. Although each probe block was presented up to two times, the data in Figure 1 include the performances evoked by the last presentation of each probe block. Each sector depicts the data for one participant. The top and bottom panels in each sector depict data for the probes in Classes 1 and 2, respectively. In addition, the clustered columns within each graph depict data from the two probe types that were presented together in a probe pair. The values on the ordinate represent the percentage of trials that occasioned selection of the nodally proximal stimulus. The particular features of the probe pair that controlled responding can be inferred from the pattern of responding evoked by the two probe types for each class in a probe block.
Fig 1.

Performances on all within-class preference probes for all participants in Experiment 1. Each type of probe is represented by one bar. The two probe types that were included in a given probe block are presented as a clustered pair of bars. The clusters of probes are depicted on the abscissa in their order of presentation. The values on the ordinate are the percentage of trials that occasioned selection of the nodally proximal stimulus (CoP).
Control by Nodal Proximity
Conditional control of responding based on nodal proximity is defined as the selection of the nodally proximal stimulus on at least seven of eight trials for a given probe pair, as illustrated by the ABC–ACD probe pair for Class 2 for Participant 1, among others. Eight within-class probe pairs (indicated by the pairs of bars) were presented to each participant. Conditional selection reflective of control by nodal proximity was evoked by 88% of probe pairs (7 of 8) for 1 participant (1), by 75% of the probe pairs (6 of 8) for 2 participants (2 and 3), by 25% of probe pairs (2 of 8) for 1 participant (6), and by 0% of the probe pairs for 3 participants (4, 5, and 7). As a group, these performances indicate that nodal structure was not a strong determinant of relatedness among the stimuli in multinodal equivalence classes. When viewed by probe type across participants, 38% of the probe pairs evoked performances reflective of control by nodal proximity.
Other Sources of Stimulus Control
Many of the probe pairs occasioned performances indicative of other forms of stimulus control such as nodal distality, preferences for particular comparisons, or avoidance of a particular comparison. These are described next.
Control by nodal distality
Control of responding based on nodal distality, was demonstrated by the selection of the nodally distal stimulus on at least seven of the eight trials for a given probe pair. This is illustrated with the ABD–DCA probes in Classes 1 and 2 for Participant 5. Responding on 16% of probe pairs (9 of 56) produced performances reflective of this form of stimulus control.
Discriminative control by a comparison stimulus: stimulus preference
In many probe blocks, the same stimulus served as the nodally proximal comparison in one probe and the nodally distal comparison in the other probe. The selection of the constant comparison on at least seven of the eight trials in the probe pair would indicate that comparison selection reflected the unconditional discriminative control by, or preference for, a particular stimulus. Preferences for the B comparison are illustrated with the DCB–DBA probes for Classes 1 and 2 for Participant 6. The B and C comparisons were preferred in 12 and 4 of the cases, respectively. In all 16 cases, the same comparison was usually selected in the presence of two different samples. Thus, the performances could not have been determined by a conditional relation between the sample and comparison stimuli. To summarize, responding on 29% (16 of 56) of the probe pairs indicated selection of a particular comparison stimulus which reflected unconditional control by a preferred stimulus.
Avoidance or rejection of a comparison
In some probe blocks, such as ABC–ACD, a participant could select the comparisons that were unique across the two probes; for example, B in the ABC probe and D in the ACD probe. This occurred in the ABC–ACD probe block for Participant 5. A performance such as this can be interpreted in a few ways. First, responding could have been controlled by rejection or avoidance of the constant comparison, the C stimulus. Alternatively, these performances might reflect control by proximity on the ACD probe and distality on the ABC probe. Based on parsimony, we support the former interpretation. Performances on 7% (4 of 56) of the probe pairs indicated responding reflective of control by rejection of a particular stimulus.
Preference or rejection of a comparison
In some probe blocks, such as DCB–ABC, a participant could respond by selecting the same comparison stimulus on all trials. Since both of the comparisons were the same for both of the probe types in this pair (e.g., B and C in the DCB–ABC probe pair), it cannot be determined whether the participant was responding based on a preference for the stimulus selected or rejection of the comparison stimulus that was not selected.
Indeterminate control and equal relatedness
In the remaining 5% (3 of 56) of the probe pairs, the nodally proximal comparisons in the two probes were selected on approximately 50% of the trials, as illustrated by the ABD–DCA probes for Classes 1 and 2 for Participant 7, among others. In these cases, we could not discern a specific source of stimulus control. This finding would imply an equality of strength between each comparison and the sample for these particular probes. If findings such as these were obtained for most of the probes, it would support the view that the stimuli in the equivalence classes were equally related to each other. Since that was not the case, these results provide weak support, at best, for the view that the stimuli in an equivalence class are equally related to each other.
Within-Participant Commonality of Test Performances across Classes
As can be seen in Figure 1, performances on the within-class preference tests varied across and within participants. Nonetheless, each participant demonstrated similar patterns of responding for each probe pair in Classes 1 and 2. This occurred for most participants, suggesting that participants preferred to respond using the same stimulus control topography for relations in the same probe pair across classes. Thus, even if nodal structure might have influenced the selection of a proximal comparison in one class and another source of control influenced comparison selection in the other class, only one of these patterns of responding would have prevailed. This interpretation will be considered in more depth in the Introduction of Experiment 2.
Sources of Stimulus Control by Probe Type
Table 3 enumerates the frequency of occurrence of particular stimulus control topographies that influenced the performances evoked by each of the four within-class probe pairs. Data were aggregated across classes and participants. All four probe pairs occasioned performances controlled by nodal proximity. Three of the probe pairs led to performances controlled by nodal distality. One probe pair each led to performances that were controlled by a preference for the B or C stimuli or rejection of the B or C stimuli. For three of the probe pairs, it was not possible to identify the sources of stimulus control that determined responding. Finally, for some of the presentations of the DCB–ABC probe pairs, which were indicated by *, the predominant selection was the B stimulus. This performance could have been determined either by preference for the B stimuli, since B was present on all trials in the block, or rejection of the C stimuli, since C was present on all trials but was never selected. In general, then, there was no apparent systematic relation between particular probe pairs and the stimulus control topographies that determined test performances.
Table 3.
The number of probe pairs that evoked comparison selections based on one of the listed stimulus control topographies.

Maintenance of Equivalence Classes
As just noted, the within-class tests resulted in substantial levels of variability in responding, which might raise questions about the integrity of the previously established equivalence classes. This possibility was addressed by considering the performances in the cross-class emergent-relation tests presented after the completion of the within-class preference tests. During these cross-class tests, all participants responded in a class-consistent manner on at least 95% of the test trials. These performances demonstrated the maintenance of the two originally established equivalence classes. Thus, the within-class tests did not disrupt the integrity of the underlying equivalence classes, regardless of the responses evoked by the within-class probes.
Summary
Seven of 10 participants formed equivalence classes under the simultaneous protocol. Thereafter, only 1 of the 7 participants showed test performances in the within-class preference tests that reflected control by nodal proximity, and thus the differential relatedness of stimuli in the equivalence classes. When aggregated across probe types, classes, and participants, 38% of the within-class probe pairs evoked the selections of nodally proximal comparisons. The remaining probes occasioned responding controlled by stimulus preferences, rejection of comparisons, or nodal distality. These results, then, did not provide a strong demonstration of the effects of nodal distance on the differential relatedness of stimuli in multinodal equivalence classes. Rather, they indicated that many forms of stimulus control influenced responding and thus precluded drawing definitive conclusions about the differential relatedness of stimuli in equivalence classes. In addition, the notion that the stimuli in the equivalence class were equally related to each other was also not supported because the actual outcomes of the experiment did not match those required by the predictions of equal relatedness.
The literature mentioned in the Introduction indicated that nodal structure influenced test performances in a systematic manner in a number of different testing conditions. In Experiment 1, however, only 38% of the within-class preference probes yielded performances that were controlled by nodal proximity. This disparity in outcome suggests that the expression of the effects of nodal structure using procedures like those specified in Experiment 1 could be enhanced with the addition of some as yet unspecified variable. Experiment 2 was designed to identify such a variable.
EXPERIMENT 2
In Experiment 1, the equivalence classes were formed using trials that contained two comparisons, one from each class. During exit interviews, a large number of participants reported that they learned relations among the stimuli in one class, and also learned to respond to the stimuli in the other class by responding away from the stimuli in the learned class. Specifically, during an emergent-relations test trial, the presentation of a sample from the “unlearned” class led the participant to respond away from the comparison from the “learned” class; thus, responding that appeared to reflect the presence of the other class really involved rejection of the “learned” class. The stimuli in the other “class”, then, were not being treated as members of a class.
The literature cited in the Introduction supports the view that when an equivalence class is formed, participants also learn about the nodal structure of the class. If, however, participants formed only one class (as reported by some of the participants in Experiment 1), as they acquired the baseline relations among the stimuli in the “learned” class, they would also have learned of the nodal structure of that class. In addition, because they did not learn the baseline relations among the stimuli in the other set, they could not have learned of the nodal structure of that set. Thus, nodal structure could not have controlled behavior in a systematic manner for the latter class. As already noted, the participants responded in a similar manner to the probes in both classes, and in general did not respond in accordance with nodal proximity. These results suggest that any control of behavior by nodal proximity for the stimuli in the class that was actually formed was overshadowed by a preference for responding in a similar manner to stimuli in the same probe pair in both classes.
It follows from this argument that a procedure which would guarantee the formation of both classes should also induce control of behavior by nodal structure in each of the classes, and thereby give rise to performances in the preference tests that were correlated with nodal proximity. According to Sidman (1987), to insure that participants form all experimenter-defined equivalence classes, at least three comparisons should be used during the establishment of the baseline relations. This occurs because the inclusion of three comparisons minimizes the selection of class-consistent comparison selection by the rejection of another comparison stimulus, and maximizes selection based on within-class relations for all trained classes. Thus, the use of three comparisons ensures the explicit formation of all specified equivalence classes (Carrigan & Sidman, 1992; Johnson & Sidman, 1993; Sidman, 1987; Sidman, 1994). If all classes are formed using this sort of procedure, it is also possible that control by nodal structure would be induced for all classes and preference tests should occasion selections in accordance with nodal proximity.
Although researchers have implemented this three-comparison strategy by the establishment of three equivalence classes, others have demonstrated the formation of two equivalence classes with the use of three comparisons: one from each class and a null stimulus that was not a member of either class (Dougher, Augustson, Markham, Greenway, & Wulfert, 1994). If two explicit classes are established, the nodal relations among the stimuli in each class should also influence responding. Thus, in Experiment 2, training of the baseline relations and testing for the emergence of derived relations were conducted using trials that contained one comparison from each class along with a third null comparison. If participants respond in a class-indicative manner to the stimuli in both classes, they must be attending to relations among all stimuli in a class and for both classes. It is our hypothesis that equivalence class formation conducted with a null comparison must also establish stimulus control by the nodal structure of equivalence classes and evoke the selection of nodally proximal stimuli during within-class tests.
Method
Participants
Twelve undergraduate students enrolled in a Psychology 101 course at Queens College served as the participants to fulfill a course requirement. All participants read and acknowledged the Informed Consent Statement given to them before the start of the experiment. The experiment lasted from 2.5 to 3.5 hr and was conducted in one session.
Apparatus
Hardware, software, and stimuli were the same as those used in Experiment 1. In addition, the stimuli used for the third comparison were: PIB (A3), DUR (B3), SOF (C3), JEH (D3).
Procedure
Phases 1 and 2. Instructions and keyboard familiarization and preliminary training using the simple-to-complex protocol
Both phases were the same as in Experiment 1.
Phase 3: Equivalence class formation using the simultaneous protocol
This was the same as in Experiment 1 with one exception. Each training and testing trial contained three instead of two comparison stimuli: the positive comparison which was from the same class as the sample, the negative comparison which came from the class not represented by the sample, and a null comparison. During initial training blocks, each sample stimulus was presented on six trials, with each comparison stimulus appearing two times in each of the left, middle, and right positions. Therefore, each training block contained 36 trials. During feedback reduction each sample stimulus was presented on three trials, with each comparison stimulus appearing once in each of the left, middle, and right positions. Once all baseline relations were established and maintained with no informative feedback, the participants were presented with an emergent- relations test that contained all of the baseline, symmetrical, one- and two-node transitive and equivalence-relations probes. Six trials were presented for each relation; three trials for the relation from Class 1 and three from Class 2 in which the position of the Co+ was on the right for one trial, the middle for one trial, and the left for the other trial. All of the tests that made up the emergent-relations test were spread across three test blocks, with one trial for each relation for each class presented in a test block. Therefore, 24 trials were presented in a test block for a total of 72 trials. Within a block all trials were presented in a randomized order without replacement. This test (combination of three blocks) was repeated up to four times or until the participant performed with at least 92% accuracy on each of the three blocks.
Phase 4: Within-class preference tests
Once the participant passed the emergent- relations test, participants were presented with the same within-class preference test trials used in Experiment 1. These trials contained only two comparisons and no null comparison.
Phase 5: Emergent-relations test 2
This phase evaluated the maintenance of the two equivalence classes with the re-administration of the emergent-relations test described in Phase 3.
Results and Discussion
Equivalence Class Formation under the Simple-to-Complex Protocol
During preliminary training, the STC protocol was used to establish two 3-node five-member equivalence classes. These classes were formed by 10 of the 12 participants, who then continued into the main part of Experiment 2. The 2 remaining participants formed four-member classes during exposure to the STC protocol and ran out of time before expanding class size to five members. Thus, these 2 participants did not continue in the present experiment.
Equivalence Class Formation under the Simultaneous Protocol
Of the 10 participants who formed the two 5-member classes under the STC protocol, 7 formed two new 2-node four-member equivalence classes under the simultaneous protocol. Of the 3 participants who did not form the classes, 1 did not acquire the baselines in the allotted time, and 2 acquired the baselines but ran out of time before the administration of the emergent- relations test blocks.
Within-Class Preference Tests
All 7 participants were given the within-class preference tests, the results of which are depicted in Figure 2. All of the within-class probes evoked the selection of the nodally proximal comparison stimuli on at least seven of eight trials in each probe. A comparison of some outcomes showed that a 0-node baseline relation was preferred to a one-node transitive relation, and that was preferred to a two-node transitive relation (0nB > 1nT > 2nT). A comparison of other outcomes showed that a 0-node symmetrical relation was preferred to a one-node equivalence relation, and that was preferred to a two-node equivalence relation (0nS > 1nE > 2nE).
Fig 2.

Performances on all within-class preference probes for all participants in Experiment 2. The format is the same as in Figure 1.
The nearly unanimous selection of nodally proximal comparison stimuli across all of the within-class probes strongly supports the view that the strength of relations among the stimuli in an equivalence class is an inverse function of the nodal distance that separates the stimuli among the trained relations that are the prerequisites of an equivalence class. This result stands in striking contrast to the results obtained in Experiment 1, and is correlated with the use of the null comparison during equivalence class formation in the present experiment and its absence during class formation in Experiment 1. Thus, these data suggest that the inclusion of the null comparison during equivalence class formation was responsible for enhancing sensitivity to the nodal structure of the equivalence classes in all participants.
Maintenance of Equivalence Classes
When reexposed to the cross-class emergent-relation tests, all participants responded in a class-consistent manner on at least 95% of the test trials. Thus, the within-class tests did not disrupt the integrity of the underlying equivalence classes. A necessary corollary of this finding is that the underlying equivalence classes existed during the administration of the within-class preference tests. This matter will be considered in more detail in the General Discussion.
Effect of the Null Comparison
The inclusion of the null comparison in the training and testing trials used to establish and document the formation of equivalence classes did not influence the percentage of participants who formed equivalence classes. It did, however, have a profound effect on the control exerted by nodal structure on the selection of comparisons during virtually all within-class probes. These results suggest that the inclusion of the null comparison during the formation of equivalence classes results in the establishment of two distinct equivalence classes where the nodal structure is learned between the relations in both of the classes. Therefore, the establishment of nodal structure in both classes results in performances indicative of control by nodal structure. The effect of the null comparison was shown in a between-participant preparation. Additional research would be needed to determine whether similar effects would be obtained on a within-subject basis.
EXPERIMENT 3
In Experiment 2, the range of nodal comparisons that could be evaluated was limited by class size. In addition, many of the within-class probes had procedural factors that were correlated with nodal proximity. One such procedural factor that occurred for the DBA and ACE probes was that each sample–comparison relation differed in terms of nodal number as well as the number of functions acquired by each comparison during training. For example, in the DBA probe, the B stimulus served as a sample on the BC trials and as a comparison on AB trials, thus B serves two behavioral functions; that is, the B stimulus served as a node. Conversely, the A stimulus only served as a sample on AB trials, thus A serves only one behavioral function, that is, the A stimulus served as a single. Therefore, the selection of the nodally proximal comparison could have been based on the selection of the comparison that had acquired two functions and not the comparison that acquired one function, that is, the participant could have selected the comparison that served as a node instead of a single. The other procedural factor that was correlated with nodal proximity on the ABC and DCB probes was that each sample–comparison relation differed in terms of relational type. For example, on the ABC probes both comparisons bore different types of relations to the sample stimulus, that is, a 0-node baseline relation for AB and a one-node transitive relation for AC. Thus, the selection of the B comparison could be attributed to a preference for a baseline relation over a transitive relation. Finally, on the ABD and DCA probes, nodal proximity was correlated with both relational type and number of functions acquired by the comparisons. Some of these factors can be eliminated by the establishment of larger equivalence classes with a greater number of nodal stimuli. Thus, Experiment 3 involved the establishment of two 3-node five-member classes, followed by the administration of some within-class probes in which nodal proximity was not confounded with the procedural factors mentioned above.
Method
Participants
Twelve undergraduate students enrolled in a Psychology 101 course at Queens College served as the participants to fulfill a course requirement. All participants read and acknowledged the Informed Consent Statement given to them before the start of the experiment. The experiment lasted from 2.5 to 3.5 hr and was conducted in one session.
Apparatus
The hardware and software were the same as those used in all aspects of Experiments 1 and 2. The stimuli used during preliminary training and during equivalence class training under the simultaneous protocol were the same as in Experiment 1. The stimuli in the two 5-member classes formed under the simultaneous protocol in Experiment 3 were QIJ (A1), TUW (B1), COH (C1), MEP (D1), and RAB (E1); in Class 2, they were: VIF (A2), KUY (B2), XOL (C2), GEZ (D2), and NAS (E2). In addition, the stimuli used for the third comparison were: PIB (A3), DUR (B3), SOF (C3), JEH (D3), and LAV (E3).
Procedure
Phases 1 and 2. Instructions and keyboard familiarization and preliminary training using the simple-to-complex protocol
Both phases were the same as in Experiment 1.
Phase 3: Equivalence class formation under the simultaneous protocol
In this phase equivalence classes were established with stimuli that differed from those used in Phase 2, and by the concurrent training of the AB, BC, CD, and DE baselines for two 3-node five-member equivalence classes that would produce a nodal structure represented by A→B→C→D→E. The baseline relations that were trained between the Class 1 stimuli were A1–B1, B1–C1, C1–D1, and D1–E1. The baseline relations that were trained between the Class 2 stimuli were A2–B2, B2–C2, C2–D2 and D2–E2. Each trial consisted of the presentation of the sample stimulus along with a stimulus from the same class, the stimulus with the same letter designation from the other class, and a third comparison from the third stimulus set with the same letter designation as the other two comparisons.
During initial training blocks, each sample stimulus was presented on six trials, with each comparison stimulus presented two times each on the left, middle, and right positions. Each training block consisted of 48 trials. During the maintenance phase, when block feedback was 75, 25, and 0%, each sample stimulus was only presented three times and each comparison was presented one time in each position, for a total of 24 trials per block.
Thereafter, all of the baseline relations, symmetrical relations, one-, two-, and three-node transitive and equivalence relations were presented in a set of test blocks. Six trials were presented for each relation. Each set of test blocks contained three trials for a given relation from Class 1 and three trials from Class 2, in which the positions of the comparisons changed on each trial so that each stimulus was presented once in each position (i.e., left, middle, right). Although the test consisted of 120 trials, they were presented in a set of three test blocks, each of which contained 40 trials. Each block included one trial for each relational type for Classes 1 and 2. Within a block, all trials were presented in a randomized order without replacement. This set of three blocks was repeated up to four times or until the participant performed with at least 92% accuracy on each of the three blocks in the a set.
Phase 4: Within-class preference tests
Once the participant passed the emergent relations test, the within-class preference test was administered to assess the effects of nodal distance. In the preference tests, each trial contained a sample and two comparisons, all from the same class. The comparisons differed in terms of the nodal distances that separated each from the sample.
Table 4 lists the within-class probes used to assess the effects of nodal distance on preference among the members of each of the three-node five-member classes. Time constraints prevented us from administering all of the possible probes. Instead, we selected a variety of probes to evaluate the effects of many nodal spreads and minimize the effects of possible confounds on test performances. As in Experiment 1, the first block consisted of probes that contained the largest nodal spread between the two comparisons and the sample stimulus.
Table 4.
Within-class preference probes for Experiment 3. See description for Table 2.

Test Block 1 contained ABE and EDA probes. The ABE probe pitted a 0-node baseline relation against a three-node transitive relation. The EDA probe pitted a 0-node symmetrical relation against a three-node equivalence relation. In both probes maximal nodal spreads separated the comparisons from the samples. Test Block 2 contained ACD and EDC probes. The ACD probes pitted one- and two-node transitive relations against each other. The EDC probes pitted a 0-node symmetrical relation against a one-node equivalence relation. Test Block 3 contained ECB and ABC probes. The ECB probes pitted one- and two-node equivalence relations against each other. The ABC probes pitted a 0-node baseline relation against a one-node transitive relation. Test Block 4 contained ACE and ABC probes. The ACE probes pitted one- and three-node transitive relations against each other. The ABC probe pitted a baseline relation against a one-node transitive relation. Test Block 5 contained DBA and DCB probes. The DBA probes pitted one- and two-node equivalence relations against each other. The DCB probe pitted a 0-node symmetrical relation against a one-node equivalence relation.
Phase 5: Emergent-relations test 2
The maintenance of the two equivalence classes was evaluated with the re-administration of the emergent relations test described in Phase 3.
Results and Discussion
Equivalence Class Formation under the Simple-to-Complex Protocol
Ten of the 12 participants formed two 3-node five-member equivalence classes using the STC protocol. The 2 remaining participants formed four-member classes but ran out of time before expanding class size to five members.
Equivalence Class Formation under the Simultaneous Protocol
Of the 10 participants who formed the two 5-member classes under the simple-to-complex protocol, 7 formed two new three-node five-member equivalence classes under the simultaneous protocol. Of the 3 participants who did not form the classes, 1 did not acquire the baselines during the allotted time, and 2 acquired the baselines but ran out of time before the administration of the emergent relations test blocks. Due to time constraints, only 4 of the participants who formed three-node five-member classes under the simultaneous protocol were given the within-class preference tests.
Within-Class Preference Tests
Figure 3 presents the results of the within-class preference tests for the 4 above- mentioned participants. The format of Figure 3 was like that used for Figures 1 and 2 but with different probe types. With one exception, the trials in all of the within-class probes evoked the selection of the nodally proximal comparison stimuli on at least seven of eight trials in each probe. A comparison of some outcomes showed that a 0-node baseline relation was preferred to a one-node transitive relation, and that was preferred to a two-node transitive relation. In addition, a one-node transitive relation was preferred to a three-node transitive relation (0nB > 1nT > 2nT) and (1nT > 3nT).
Fig 3.
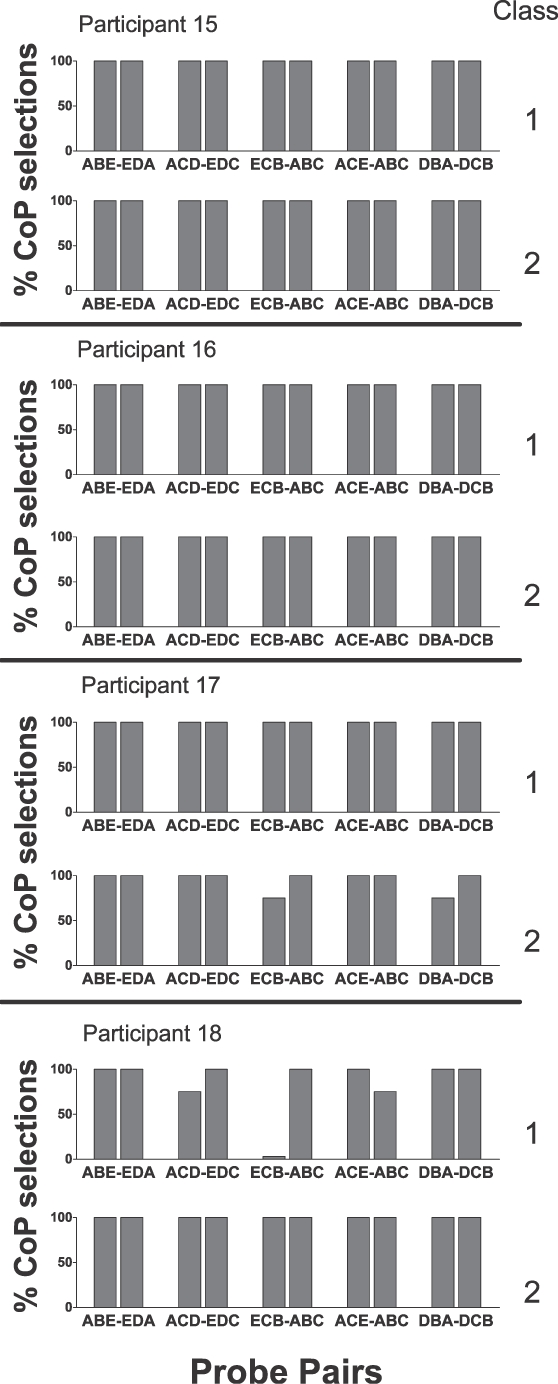
Performances on all within-class preference probes for all participants in Experiment 3. The format is the same as in Figure 1.
A comparison of other outcomes showed that a 0-node symmetrical relation was preferred to a one-node equivalence relation, and that was preferred to a two-node equivalence relation (0nS > 1nE > 2nE). In addition, a 0-node symmetrical relation was preferred to a three-node equivalence relation (0nS > 3nE), and a one-node equivalence relation was preferred to a two-node equivalence relation (1nE > 2nE). The nearly unanimous selection of nodally proximal comparison stimuli across all of the within-class probes strongly supports the view that the strength of relations among the stimuli in an equivalence class is an inverse function of the nodal distance that separates the stimuli among the trained relations that are the prerequisites of an equivalence class. The one exception to this general outcome occurred with the ECB and ABC probes for Class 1 for Participant 18. In these probes, all trials evoked selection of the B stimulus. Therefore, responding on these probes demonstrated control by preference for B or rejection of the C stimulus.
Maintenance of Equivalence Classes
When reexposed to the cross-class emergent-relations tests, all participants responded in a class-consistent manner on at least 95% of the test trials. These performances demonstrated the maintenance of the two originally established equivalence classes. Thus, the presentation of within-class tests between the two sets of cross-class tests did not disrupt the integrity of the underlying equivalence classes. As in Experiment 2, these results support the view that the underlying equivalence classes existed during the administration of the within-class preference tests.
Coexistence of Equal and Graded Relatedness
Responding indicative of class integrity and the equal relatedness of stimuli was expressed with the presentation of cross-class tests. Responding in accordance with nodal distance or graded relatedness was expressed immediately upon the presentation of within-class tests. Both of these performances were expressed immediately in successively presented test blocks, all of which provided no informative feedback for any of the comparisons selected on a trial. These results showed that both between- and within-class sources of stimulus control coexisted but were expressed separately by the presentation of test trials that contained stimuli from different classes or from the same class.
GENERAL DISCUSSION
Effect of Null Comparison on Class Formation and Nodal Distance Effects
Three experiments assessed the strength of relations among the stimuli in equivalence classes by use of post-class formation within-class preference tests. In Experiment 1, 7 of the 10 participants formed two 2-node four-member classes using trials that contained two comparisons each. In post-class formation preference tests, only 1 of the 7 participants responded to all of the probes in a manner indicative of control by nodal proximity. Only 38% of the probe pairs in the within-class preference tests presented to the totality of the participants evoked responding indicative of control by the nodal separation of stimuli in the classes. Although a relatively large percentage of participants formed the equivalence classes, nodal distance exerted a minimal effect on within-class test performances.
In Experiments 2 and 3, a similar percentage of participants formed equivalence classes, but the two classes were formed using trials that contained one comparison from each class and a third null comparison. When the within-class preference tests were administered, virtually all of the test trials presented to all participants evoked responding that reflected control by the nodal proximity of the stimuli in the previously established classes. Thus, the strength of relations among the stimuli in each multinodal equivalence class was an inverse function of the number of nodes that separated the stimuli in the class. This increase in control exerted by nodal structure from Experiment 1 to Experiments 2 and 3 can be attributed to the inclusion of a third null comparison on all trials used to establish the underlying equivalence classes. Although the inclusion of a null comparison increased sensitivity to the effects of nodal proximity, it did not influence the likelihood of forming equivalence classes under the simultaneous protocol.
Generality of Nodal Distance Effects
In two earlier studies (Fields et al., 1995; Fields & Watanabe-Rose, 2008), nodal distance effects were demonstrated by participants who were exposed to tests used to assess the post-class formation effects of nodal distance. The percentage of participants in these studies who showed the effects was very low because only a small number of them formed equivalence classes under the SIM protocol. Thus, the generality of the nodal distance effect in larger populations could be questioned.
In the present experiments, however, much higher percentages of participants formed equivalence classes under the SIM protocol because of the prior establishment of other equivalence classes under the STC protocol. Thus, a relatively large percentage of participants were eligible to engage in the preference tests used to measure the effects of nodal distance. All of the participants in Experiments 2 and 3 showed control of responding by nodal structure during the within-class preference tests. Thus, the control of behavior by nodal structure for a large proportion of participants was achieved by the combination of two procedures: (1) the prior establishment of equivalence classes under the STC protocol, and (2) the subsequent establishment of new equivalence classes under the simultaneous protocol with trials that contained null comparison stimuli. The results of Experiments 2 and 3, then, extend the generality of the effects of nodal distance on the differential relatedness of stimuli in equivalence classes.
Nodality Effects Primed by Initial Probe Pairs?
In Experiments 2 and 3, the first set of probes in the preference test sequence involved the presentation of trials in which the comparison stimuli bore maximally different nodal spreads from the sample. These initial probes, then, might have signaled that the comparison to be selected is the one “most similar” to a trained relation. Since similarity to a trained relation is indexed by nodal separation, the presentation of these initial probes might have primed the selections of the nodally proximal comparisons in the subsequently presented probes. Regardless of the status of the priming function served by the initial trials, responding on virtually all preference tests was a direct function of nodal proximity. Of necessity, such an outcome meant that the participants must have learned the nodal structure among the stimuli in the classes during the formation of the classes. Therefore, the capability of responding to the stimuli in the equivalence class based on differential relatedness was present and could have been activated by the content of the test trials or possibly by combination of the presentation of the initial probe pair and the content of the test trial.
Although the current data cannot be used to determine whether the initial probes primed responding based on nodal structure, that possibility could be evaluated by future research that would replicate the within-class preference tests used in the present experiment but without exposure to the initial probe pairs. The outcome of such an experiment, however, would not invalidate the results obtained in Experiments 2 and 3. At best, it would identify an additional variable that activated the effects of nodal structure on the relatedness of stimuli in equivalence classes.
Scope and Robustness of Nodal Distance Effects
Previous research has shown that nodal structure influenced the delayed emergence of equivalence classes (Bentall et al., 1998; Fields et al., 1990; Kennedy, 1991; Kennedy et al., 1994; Sidman et al., 1985; Spencer & Chase, 1996), response latency and speed during and after class formation (Bentall et al.,1998; Kennedy, 1991; Spencer & Chase, 1996; Tomanari et al., 2006; Wulfert & Hayes, 1988), and the gradual transfer of responding after class formation (Fields et al., 1993b). In addition, nodal distance influenced the bifurcation of previously formed equivalence classes in accordance with nodal structure (Fields et al., 1995; Fields & Watanabe-Rose, 2008). The results of Experiments 2 and 3 demonstrated yet another post-class formation effect of nodal structure: the selection of nodally closer comparisons in within-class preference tests.
In addition to extending the range of conditions that show the effects of nodal distance, the results of Experiments 2 and 3 also address some criticisms that have been raised regarding the robustness of the effects of nodal distance (Sidman, 1994, 2000). These criticisms include: (1) Nodal distance had transient effects during the delayed emergence of equivalence classes and the gradual transfer of responding among the members of equivalence classes. (2) The tests used to assess nodal distance effects forced outcomes indicative of nodal distance. (3) The effects of nodal distance were diminished when more than two classes were established on a concurrent basis. (4) There could have been potential biases, based on different reinforcement histories acquired by stimuli, that were confounded with nodal distance.
None of these criticisms were relevant to the data reported in Experiments 2 and 3. With regard to these four criticisms: (1) the effects of nodal distance were demonstrated in a steady state after the formation of the classes. Thus, the effects of nodal distance were not transient. The same can be said of the results reported by Fields and Watanabe-Rose (2008) and Fields et al. (1995). (2) The design of the within-class preference tests permitted a participant to respond independently of, or in accordance with nodal structure. Therefore, the test performances that matched the predictions based on nodal distance could not be attributed to a forcing of the outcome based on design biases embedded in the test trials themselves. The same can be said of the results reported by Fields and Watanabe-Rose (2008) and Fields et al. (1995). (3) The results of the preference tests were obtained with a wide range of nodal spreads after the formation of two equivalence classes using a third null comparison during training and testing. Rather than a diminution, a maximal effect of nodal distance was demonstrated for the set of two equivalence classes established in each experiment. (4) The use of the simultaneous protocol to establish the equivalence classes minimized differences in stimulus–reinforcer biases that could have been correlated with nodal distance. To summarize, the results of the preference tests in Experiments 2 and 3 could not be accounted for by any of the potential confounding factors mentioned in the previous paragraph. Therefore, the results of the Experiments 2 and 3 support the view that nodal distance exerted a stable and robust effect on the differential relatedness of stimuli in an equivalence class that was not confounded by the factors mentioned above.
Possible Confounds with Nodal Distance in the Preference Tests
The present experiments demonstrated that nodal structure influenced the selection of nodally proximal stimuli as measured using within-class preference tests conducted in a matching-to-sample format. The performances evoked by the ACD and ECB probes in Experiment 3 provided unambiguous and unconfounded demonstrations of the effects of nodal distance on relational preferences after class formation. For other probes in the present experiments, however, nodal proximity was sometimes correlated with relational type and/or the number of functions served by the comparisons. Thus, performances that were correlated with nodal proximity could also have been influenced by these latter variables instead of, or in addition to nodal proximity. These possibilities are illustrated with the ABC, DBA, and EDA probes.
In the case of the ABC probes, different types of relations existed between the sample and each comparison along with differences in nodal spread. Specifically, AB was a 0-node baseline relation while AC was a one-node transitive relation. Thus, the preference for the nodally proximal stimulus might have also reflected control by a trained relation rather than an emergent relation. In the case of the DBA probes, a different number of functions had been acquired by the two comparisons. Specifically, the B stimulus had acquired both a sample function during BC training and a comparison function during AB training, while the A stimulus had acquired only a sample function during AB training. Therefore, DB, the one-node equivalence relation was correlated with a comparison that served two functions while DA, the two-node equivalence relation, was correlated with a comparison that served only one behavioral function. Thus the preference for the nodally proximal comparison might have been controlled by a preference for a comparison that served two instead of one behavioral function. Finally, in the case of the EDA probes, the sample and comparisons differed in terms of relational type (ED being a symmetrical relation and EA being an equivalence relation) and in number of functions served by each comparison (D served as a node and A served as a single). Therefore, the preference for the nodally proximal comparison might have been controlled by a preference for a comparison that served as a node or for a symmetrical relation instead of an equivalence relation.
In summary, the performances evoked by the within-class preference probes, which were correlated with nodal proximity, might have been controlled by other variables. That notion, however, can be ruled out for the following reasons. If these nominally confounded variables were responsible for selection instead of nodal proximity, probes that did not have these confounds (i.e., ACD and ECB) would have produced random selection of comparisons. That, however, did not occur: Instead, virtually all probes occasioned the selection of the nodally proximal comparisons regardless of the presence or absence of these confounds. Since these formally confounded variables were not correlated with test performances, these factors did not account for the performances evoked by the totality of the within-class probes. Nodal proximity was the only factor that was present in all types of probes and was systematically correlated with test performances. Therefore, nodal proximity had to be the variable that was the primary determinant of the relatedness of stimuli in these classes in the present experiments.
Intactness of Equivalence Classes during and after Preference Testing
In all three experiments, after completing the within-class preference test, participants were presented with a final cross-class emergent relations test block. Virtually all of the probes in this block evoked responding indicative of the intactness of the equivalence classes. This occurred regardless of the performances evoked by the within-class preference tests (i.e., in Experiments 1, 2, and 3). Thus, the underlying equivalence classes, which were documented in the emergent-relations tests conducted prior to preference testing must have remained intact during the administration of the within-class preference probes. Of interest, the integrity of the classes was also maintained regardless of the performances occasioned by the within-class preference tests. Similar results were reported by Fields and Watanabe-Rose (2008) who found that equivalence classes remained intact after demonstrations of class bifurcation in dual-option response transfer tests. The results of the latter experiment then demonstrated that the integrity of an equivalence class is maintained following a variety of procedures used to measure differential relatedness of stimuli in the classes.
These results have important implications for Sidman's interpretation of the existence of equivalence classes (Sidman, 1994, 2000). He noted that for the stimuli in a set to be functioning as members of an equivalence class, they must demonstrate the property of interchangeability or substitutability. He went on to state that, “Without [this] common characteristic … the class does not exist.” (1994, p. 543). The data collected during the emergent relations tests conducted after the administration of the within-class preference tests raise questions about this analysis. When the participants in Experiments 2 and 3 were exposed to the emergent relations test after the administration of the preference probes, virtually all of the emergent relations probes evoked responding that demonstrated the intactness of the equivalence classes. According to Sidman's analysis, the classes did not exist—could not be defined—during the previously conducted within-class preference tests (Sidman, 1994, p. 543). The class-consistent responding seen during the final emergent relations tests then could have occurred only with the instant re-creation of the classes without benefit of any training. Because this is implausible, it is far more likely that, once formed, equivalence classes remain intact during the administration of the within-class preference tests. These results then imply that two stimulus control topographies coexist among the stimuli in an equivalence class: one class-based and reflective of equal relatedness among class members, and the other nodally-based and reflective of differential relatedness among the class members. The essential issue, then, is not whether these two stimulus control topographies exist but rather how these topographies come to control responding in different test trials.
Theoretical Integration
Many experiments have shown that a number of stimulus control topographies can gain control of responding when training is conducted using trials presented in a matching-to-sample format (Fields, Garruto, & Watanabe, 2010; Iversen, 1997; Iversen, Sidman, & Carrigan, 1986; McIlvane & Dube, 2003). Whether intended or not, these topographies acquired control because of their planned or adventitious correlation with the contingencies of reinforcement that established the experimenter-defined relations. These findings make plausible the assumption that training conditions designed to ensure the formation of all experimenter-defined equivalence classes also result in the acquisition of control by more than one relation among the stimuli in a class, specifically, class-based and nodally-based stimulus control topographies.
When a set of linked conditional discriminations are established, they define a nodal structure among the stimuli in the relations. When this is done, the relatedness among stimuli in the set will be an inverse function of the number of nodes that separate the stimuli in the trained relations. This assumption is driven by the extensive literature on the relatedness of stimuli in semantic memory networks (Branch, 1994; Collins & Loftus, 1975; Collins & Quillian, 1969; Fields & Verhave, 1987). If established, this differential relatedness is maintained even in the presence of tests devised to assess the formation of equivalence classes or the relatedness of stimuli in the class. In addition, the contingencies embedded in the matching-to-sample trials reinforce differential responding to stimuli in different sets, that is, discrimination between sets, also called class-based stimulus control topographies.
After the establishment of the linked baseline relations, the formation of the equivalence class is evaluated with the presentation of derived relations probes, each of which contains a sample stimulus from one set (Sa), one comparison stimulus from the same set (i.e., the positive comparison or Co+), and at least one other stimulus from other sets (i.e., the negative comparisons or Co-s). The Sa and Co+ are linked by a set of intervening nodes while any Co− is explicitly not linked by training to the sample stimulus of that trial. Thus, the contingencies of reinforcement should result in the selection of comparisons from the same set as the sample rather than stimuli in sets that differ from the sample stimuli. These emergent-relations test performances would show a discrimination between classes and thus document the formation of equivalence classes.
When each stimulus in a class results in the selection of all other stimuli in the class, all of the stimuli would be functioning as stimuli that are substitutable for each other. When any emergent-relations probe is presented, any comparison from a given set is “closer” to the sample stimulus from the same set than any stimulus in another set. Therefore, the selection of a comparison from the same set is most likely, regardless of the nodal separation between the sample and comparison stimuli. On the other hand, a within-class preference probe contains both comparison stimuli from the same class. Because some stimuli are “closer” to each other based on the training structure, participants are more likely to select the comparison that is nodally closer to the sample.
In Experiments 2 and 3, participants were presented with a battery of cross-class emergent relations probes, followed immediately by a battery of within-class preference probes, followed immediately by a second battery of cross-class emergent relations probes. Although they were not accompanied by any instructions or reinforcement for responding, the cross-class probes evoked responding indicative of the presence of two classes of substitutable stimuli where nodal separation of stimuli did not influence test performances. In contrast, the preference probes evoked responding indicative of stimuli that were differentially related in accordance with nodal distance. These essentially instantaneous shifts in performance, which coincided with a change in test type, strongly suggest that the content of the trials were discriminative for the expression of each type of performances.
Indeed, the trials had features that were distinguishable from each other. The cross-class trials contained a sample from one class, a comparison from the same class and another comparison from a different class. The within-class probes contained a sample and two comparisons, all from the same class. Thus, the two types of trials were formally discriminable from each other in terms of the presence or absence of stimuli from the same or different classes. The cross-class probes then functioned as discriminative stimuli for performances controlled by the discriminability of the stimuli between classes, while the within-class probes functioned as discriminative stimuli for performances controlled by nodal proximity. These results support the notion that the content of the probes were discriminative for the occurrence of class-based or nodally-based responding (Fields et al., 1993a; Fields & Moss, 2007; Fields et al., 1995; Fields & Watanabe-Rose, 2008). This analysis is consistent with Sidman's view that different partitions of the stimuli in an equivalence class can be governed by contextual stimuli (Sidman, 1994, pp 548–549).
Although it sounds contradictory for stimuli in a class to be interchangeable and also to be differentially related to each other, the data described in the present experiments attest to such a circumstance. The immediate switches in performance imply that both stimulus control topographies had been established during the training of the baseline relations that linked the stimuli in each set and were maintained after class formation. Although the definition of an equivalence class involves a demonstration of the interchangeability of the stimuli in a set, and by implication their equal relatedness, the procedures used to establish an equivalence class also establish other relations among the stimuli in the set. These relations are indicative of a graded relatedness which is governed by the nodal separation of the stimuli in the set. While these two forms of stimulus control coexist, the expression of each form of stimulus control is determined by the two types of test trials which are discriminable from each other: one being sensitive to cross-class discriminative control and the other being sensitive to the nodal structure of the class. Performances that demonstrate equal relatedness as documented by between-class discriminations do not negate performances that demonstrate differential relatedness as documented by within-class preference tests, and vice versa.
A single set of reinforcement contingencies establishes discrimination between classes and differential relatedness among the stimuli in each class. Performances indicative of each stimulus control repertoire cannot be measured at the same time. Each, however, can be expressed or measured in the presence of a different environmental setting. Each setting is defined by the content of a given test trial. These test trials serve discriminative functions that occasion the expression of the two relations that coexist among the stimuli in a class. The results of Experiments 2 and 3 support this integrated interpretation of the relations that exist among the members of an equivalence class.
Acknowledgments
This research was conducted with support from PSC-CUNY Research Award 61669-00-39 and CUNY Collaborative Research Grant 80209-09-13. Portions of these data appeared in a doctoral dissertation submitted by the first author to The Graduate Center of The City University of New York. We thank Xiqiang Zhu for his assistance in the development of the software used to conduct the experiment.
REFERENCES
- Alligood C, Chase P.N. Choices among stimuli in equivalence classes. 2007. Unpublished doctoral dissertation, Elbery College of Arts and Sciences at West Virginia University.
- Bentall R.P, Jones R.M, Dickins D.W. Errors and response latencies as a function of nodal distance in 5-member equivalence classes. The Psychological Record. 1998;49:93–115. [Google Scholar]
- Branch M. Stimulus generalization, stimulus equivalence, and response hierarchies. In: Hayes S.C, Hayes L.J, Sato M, Ono K, editors. Behavioral Analysis of Language and Cognition. Reno, NV: Context Press; 1994. pp. 51–70. (Eds.) [Google Scholar]
- Buffington D.M, Fields L, Adams B.J. Enhancing equivalence class formation by pretraining of other equivalence classes. The Psychological Record. 1997;47:69–96. [Google Scholar]
- Carrigan P.F, Jr, Sidman M. Conditional discrimination and equivalence relations: A theoretical analysis of control by negative stimuli. Journal of the Experimental Analysis of Behavior. 1992;58:183–204. doi: 10.1901/jeab.1992.58-183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins A.M, Loftus E.F. A spreading activation theory of semantic memory. Psychological Review. 1975;82:407–428. [Google Scholar]
- Collins A.M, Quillian M.R. Retrieval time from semantic memory. Journal of Verbal Learning and Verbal Behavior. 1969;8:240–248. [Google Scholar]
- Cumming W.W, Berryman R. The complex discriminative operant: Studies of matching-to-sample and related problems. In: Mostofsky D.I, editor. Stimulus generalization. Stanford, CA: Stanford University Press; 1965. pp. 284–330. (Ed.) [Google Scholar]
- Dougher M.J, Auguston E, Markham M.R, Greenway D, Wulfert E. The transfer of respondent eliciting and extinction through equivalence classes. Journal of the Experimental Analysis of Behavior. 1994;62:331–351. doi: 10.1901/jeab.1994.62-331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebbinghaus H. On Memory: A contribution to experimental psychology:1885. (H.A. Ruger & C.E. Bussinius, Tr.) New York: Teachers College, Columbia University; 1913. [Google Scholar]
- Fields L, Adams B.J, Verhave T. Relative strength of symmetrical, transitive and equivalence relations after equivalence classes are fully formed. Paper presented in a symposium at the Annual Meeting of the Association for Behavior Analysis, Milwaukee, WI 1989, May.
- Fields L, Adams B.J, Verhave T. The effects of equivalence class structure on test performances. Psychological Record. 1993a;43:697–721. [Google Scholar]
- Fields L, Adams B.J, Verhave T, Newman S. Are stimuli in equivalence classes equally related to each other. The Psychological Record. 1993b;46:85–105. [Google Scholar]
- Fields L, Adams B, Verhave T, Newman S. The effects of nodality on the formation of equivalence classes. Journal of the Experimental Analysis of Behavior. 1990;53:345–358. doi: 10.1901/jeab.1990.53-345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L, Garruto M, Watanabe M. Varieties of stimulus control in match-to-sample: A kernel analysis. The Psychological Record. 2010;60:3–26. [Google Scholar]
- Fields L, Landon-Jimenez D.V, Buffington D.M, Adams B.J. Maintained nodal distance effects after equivalence class formation. Journal of the Experimental Analysis of Behavior. 1995;64:129–146. doi: 10.1901/jeab.1995.64-129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L, Moss P. Stimulus relatedness in equivalence classes: Interaction of nodality and contingency. European Journal of Behavior Analysis. 2007;8((2)):141–159. [Google Scholar]
- Fields L, Reeve K.F, Rosen D, Varelas A, Adams B.J, Belanich J, Hobbie S.A. Using the simultaneous protocol to study equivalence class formation: The facilitating effects of nodal number and size of previously established equivalence classes. Journal of the Experimental Analysis of Behavior. 1997;67:367–389. doi: 10.1901/jeab.1997.67-367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L, Verhave T. The structure of equivalence classes. Journal of the Experimental Analysis of Behavior. 1987;48:317–332. doi: 10.1901/jeab.1987.48-317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L, Watanabe-Rose M. Nodal structure and the partitioning of equivalence classes. Journal of the Experimental Analysis of Behavior. 2008;89:359–381. doi: 10.1901/jeab.2008-89-359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iversen I.H. Stimulus definition in conditional discriminations. Journal of the Experimental Analysis of Behavior. 1997;68:27–45. doi: 10.1901/jeab.1986.45-297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iversen I.H, Sidman M, Carrigan P. Stimulus definition in conditional discriminations. Journal of the Experimental Analysis of Behavior. 1986;45:297–304. doi: 10.1901/jeab.1986.45-297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson C, Sidman M. Conditional discrimination and equivalence relations: Control by negative stimuli. Journal of the Experimental Analysis of Behavior. 1993;29:333–347. doi: 10.1901/jeab.1993.59-333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy C.H. Equivalence class formation influenced by the number of nodes separating stimuli. Behavioural Processes. 1991;24:219–245. doi: 10.1016/0376-6357(91)90077-D. [DOI] [PubMed] [Google Scholar]
- Kennedy C.H, Itkonen T, Lindquist K. Nodality effects during equivalence class formation: An extension to sight-word reading and concept development. Journal of Applied Behavior Analysis. 1994;27:673–683. doi: 10.1901/jaba.1994.27-673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIlvane W.J, Dube W.V. Stimulus control topography coherence theory: Foundations and extensions. The Behavior Analyst. 2003;26:195–213. doi: 10.1007/BF03392076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M. Two choices are not enough. Behavior Analysis. 1987;22((1)):11–18. [Google Scholar]
- Sidman M. Equivalence relations and behavior: A research story. Boston, M.A: Author's Cooperative; 1994. [Google Scholar]
- Sidman M. Equivalence relations and the reinforcement contingency. Journal of the Experimental Analysis of Behavior. 2000;74:127–146. doi: 10.1901/jeab.2000.74-127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M, Kirk B, Willson-Morris M. Six-member stimulus classes generated by conditional-discrimination procedures. Journal of the Experimental Analysis of Behavior. 1985;43:21–42. doi: 10.1901/jeab.1985.43-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M, Tailby W. Conditional discrimination vs. matching-to-sample: An expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior. 1982;37:5–22. doi: 10.1901/jeab.1982.37-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slamecka N.J. Ebbinghaus: Some associations. Journal of Experimental Psychology: Learning, Memory, & Cognition. 1985;11:414–435. [Google Scholar]
- Spencer T.J, Chase P.N. Speed analysis of stimulus equivalence. Journal of the Experimental Analysis of Behavior. 1996;65:643–659. doi: 10.1901/jeab.1996.65-643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomanari G.Y, Sidman M, Rubio A.R, Dube W.V. Equivalence classes with requirements for short latencies. Journal of the Experimental Analysis of Behavior. 2006;85:349–369. doi: 10.1901/jeab.2006.107-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wulfert E, Hayes S.C. Transfer of a conditional ordering response through conditional equivalence classes. Journal of the Experimental Analysis of Behavior. 1988;50:125–144. doi: 10.1901/jeab.1988.50-125. [DOI] [PMC free article] [PubMed] [Google Scholar]