

Abstract
IspH is a 4Fe-4S protein that carries out an essential reduction step in isoprenoid biosynthesis. Using hyperfine sublevel correlation (HYSCORE) spectroscopy, we show that pyridine inhibitors of IspH directly bind to the unique 4th Fe in the 4Fe-4S cluster, opening up new routes to inhibitor design, of interest in the context of both anti-bacterial as well as anti-malarial drug discovery.
Keywords: Isoprenoid, non-mevalonate, IspH (LytB), inhibitors, HYSCORE
Isoprenoid biosynthesis is an important target for drug discovery.1 In most pathogenic bacteria as well as in malaria parasites, the early stages in isoprenoid biosynthesis are carried out by the methylerythritol phosphate pathway.2 This pathway is essential for producing the isoprenoids used in e.g. cell wall biosynthesis in bacteria, and in quinone formation and, since it is not present in humans, the enzymes involved are important targets for the development of new antibiotics.3 The last two enzymes, IspG and IspH, are unusual 4Fe-4S-containing proteins that carry out 2H+/2e- reductions of the substrates MEcPP (2-C-methyl-D-erythritol-2,4-cyclo diphosphate, 1)4-6 and HMBPP (E-1-hydroxy-2-methyl-but-2-enyl-4-diphosphate, 2) to form the C5 isoprenoids IPP (isopentenyl diphosphate, 3) and DMAPP (dimethylallyl diphosphate, 4) in an ∼1:5 ratio,7, 8 as shown in Scheme 1:
Scheme 1.

Reactions catalyzed by the proteins IspG (GcpE) and IspH (LytB).
In recent work we proposed that both the IspG (EC 1.17.7.1, HMBPP synthase, also known as GcpE)9 as well as the IspH (EC 1.17.1.2, HMBPP reductase, also known as LytB) catalyzed reactions involve formation of organometallic species (i.e. containing Fe-C bonds).10 Support for the intermediacy of organometallic species in catalysis comes indirectly from electron paramagnetic resonance (EPR) and electron nuclear double resonance (ENDOR) spectroscopy as well as mechanistic considerations,10 and more directly, from the observation that the Fe-C distances (2.6–2.7 Å) between the apical iron atom in the 4Fe-4S cluster and the allylic species seen crystallographically in IspH are even shorter than the ones observed for bound HMBPP,11 and are far shorter than the 3.6-3.7 Å sum of the Fe, C van der Waals radii.12 We also found that alkynes could be quite potent inhibitors of both IspG and IspH, and that EPR and ENDOR spectra indicated that these alkynes bound at or very close to the unique 4th Fe in the reduced 4Fe-4S cluster. The ability to inhibit IspG or IspH is of interest in the context of the development of anti-infectives, and the ability of a given compound to inhibit both enzymes is of even more interest since, in principle, it will lead to a decrease in drug resistance since both enzymes would be required to mutate.
In addition to alkyne inhibitors, we discovered a second class of IspH inhibitors, pyridine diphosphates,13 but how these bound to the protein was not clear. Here, we report the results of an X-band hyperfine sublevel correlation (HYSCORE) spectroscopic as well as a quantum chemical investigation, which helps clarify how these inhibitors function.
We first investigated a series of pyridine ligands, 5-11, Scheme 2, binding to wild-type IspH from Aquifex aeolicus. The continuous-wave EPR spectrum of IspH + pyridine (5) is the same as that of the unliganded protein (i.e. in the absence of pyridine, Figure S1a), and there is no evidence for any sizeable pyridine-14N hyperfine interaction in the HYSCORE spectrum (Figure S1b), indicating only very weak binding affinity to IspH. The same results are obtained with the more basic (pKa = 6.8 vs. 5.2) species 2-aminopyridine (6, Figure S1c). However, on addition of the inhibitor BPH-293 (7, IC50 = 38 μM), the EPR spectrum changes13 (Figure S1a) and new signals attributable to 14N single and double quantum transitions appear in the (+,-) quadrant of the HYSCORE spectrum (Figure 1a). The 14N hyperfine interaction is quite large, with the hyperfine coupling constant being ∼8 MHz. Reconstituted IspH (Figure 1a) and anaerobically purified IspH (Figure S2) both give the same results. The ortho and para-pyridyl analogs of 7 (compounds 8, IC50 = 1.2 mM and 9, IC50 = 149 μM) show no evidence of any sizeable pyridine-14N hyperfine interaction in their HYSCORE spectra (Figure S1d, e), due presumably to their inability to bind to the 4th Fe, for “steric” reasons. Moreover, chlorine substitution of 7 (compound 10) results in loss of all activity (IC50 > 3 mM), due presumably to loss in donor-ability of the pyridine nitrogen (the computed pKa values of the pyridine fragments in 7 and 10 are 4.7 and 0.7, respectively), consistent with the absence of a pyridine-14N HYSCORE signal (Figure S1f). Addition of one CH2 group to the side-chain of 7 results in a better inhibitor (11, IC50 = 9.1 μM), although there is no significant difference between the HYSCORE spectra of 7 (Figure 1a) and 11 (Figure S1g), indicating that differences in enzyme inhibition are due to differences in the alkyl diphosphate fragment binding in the active site, rather than differences in Fe-pyridine interactions.
Scheme 2.

Aromatic species investigated.
Figure 1.
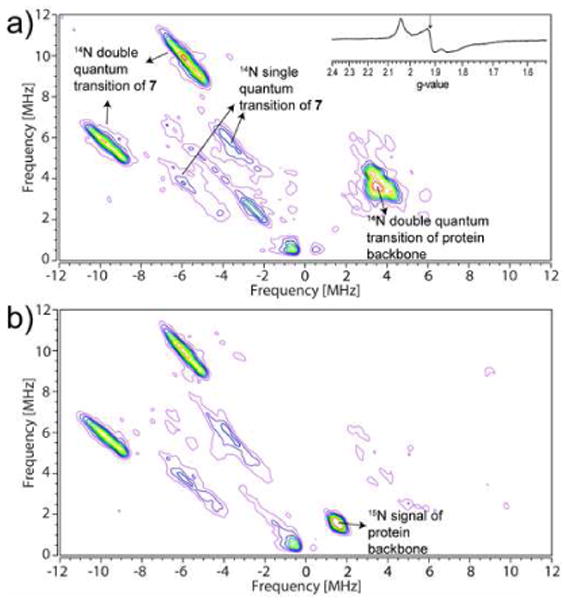
HYSCORE spectra of A. aeolicus IspH + pyridine inhibitor 7. (a) HYSCORE spectra of unlabeled A. aeolicus IspH + 7. The inset shows the CW-EPR spectrum, and the arrow indicates the magnetic field position for collecting the HYSCORE data. (b) HYSCORE spectra of 15N-labeled A. aeolicus IspH + 7. Microwave frequency = 9.66 GHz (a), 9.68 GHz (b); magnetic field was set at g2 = 1.921; τ = 136ns.
These results do not, however, prove that the 14N HYSCORE signals in the (+,-) quadrant (Figure 1a and Figure S1g) arise directly from the inhibitors 7 and 11 since, in principle, inhibitor binding might result in a protein conformational change and binding of a protein ligand to Fe, e.g. the nearby His 42 or 124, which form part of the active site.14 To investigate this possibility, we prepared a sample using uniformly 15N-labeled IspH and inhibitor 7. As can be seen in Figure 1b, the 14N signals centered at ∼3.6 MHz seen in Figure 1 are no longer present, and are replaced by a signal centered at 1.5 MHz, the 15N Larmor frequency. Moreover, the 14N signals in the (+,-) quadrant are essentially identical to those seen in samples prepared using unlabeled IspH (Figure 1a). This strongly suggests that the signals centered at ∼3.6 MHz arise from protein nitrogens near the 4Fe-4S cluster, while the 14N signals in the (+,-) quadrant arise from the bound inhibitor 7, rather than from any protein residues.
To begin to better understand the interaction between the pyridine inhibitor 7 and IspH, we next simulated the HYSCORE spectra of IspH + 7 taken at three different magnetic field strengths (Supporting Information, Figures S3a, b and c) using the EasySpin program15 (Figures S3d, e and f), finding aiso(14N) = 7.4 MHz, Aii(14N) = [6.2 7.6 8.4] MHz for the hyperfine interaction, and e2qQ/h = 3.0 MHz for the nuclear quadrupole coupling constant.
This large aiso(14N) is similar to, or even larger than, that of a number of systems in which nitrogens directly bind to Fe centers. For example, in met-myoglobin the porphyrin nitrogens have aiso = 8.11 MHz and 7.8 MHz, and the histidine Nε has aiso = 9.28 MHz.16 In a model heme complex, FeTPP(4-MeIm)2 (TPP, tetraphenylporphyrin; 4-MeIm, 4-methyl imidazole), the aiso of the porphyrin nitrogens is 5.1 MHz, while that of the coordinated 4-MeIm is 5.7 MHz.17 In Rieske-type 2Fe-2S proteins, aiso(14N) of the coordinated His nitrogens are approximately 5 MHz,18 and in the case of the 4Fe-4S enzyme MoaA (which also has an unique 4th iron), N1 of the substrate guanosine 5′-triphosphate binds to the 4th iron and has aiso (14N) = 3.6 MHz.19 On average, these results give an aiso(14N) ∼ 6 MHz for systems containing Fe-N bonds, suggesting that the IspH + 7 complex also contains an Fe-N bond.
The large 14N hyperfine interaction seen in the IspH + 7 complex might also, at least in principle, indicate that the pyridine fragment is just close-by to the reduced 4Fe-4S cluster, without directly bonding to the 4th iron. For example, the pyridine group might be protonated and interact with e.g. the E126 CO2- group that is close to the cluster; or it could be close-by, but deprotonated. Fortunately, determination of the 14N nuclear quadrupole coupling constant (e2qQ/h) enables an answer to this question, since protonated, neutral and metal-coordinated pyridine ligands have very different e2qQ/h values.20
For pyridine itself, the e2qQ/h is 4.6 MHz, but in species in which there is a formal +1 charge on N, such as the pyridinium ion (12), pyridine-N-oxide (13) and N-methyl pyridinium (14), e2qQ/h values of approximately 1 MHz are observed experimentally.20 In the case of pyridine bonded to Fe in Fe(CO)4(pyr), 15, e2qQ/h is in between these extreme values (e2qQ/h ∼ 2.4 MHz), and for Mo(pyr)2(CO)4 as well as Cr(CO)4(2,2′-bipyridyl), e2qQ/h ∼3.1 MHz. So, when pyr is bonded to Cr, Mo or Fe, the e2qQ/h decreases from the 4.6 MHz seen in free pyridine to ∼ 2.4 – 3.1 MHz, due to metal-ligand bonding, close to the 3.0 MHz value we find from the 14N HYSCORE results.
To see to what extent these e2qQ/h values might be reproduced computationally, we used the Gaussian-09 (Revision A.01) program.21 Results are given in Table S1 and are shown graphically in Figure 2 and Figure S4. Clearly, there is a good correlation (Figure 2a) between theory and experiment (R2 = 0.965; slope = 0.963) for a series of model systems, and when using [Fe4S4(SMe)3(pyr)]2- (16, Figure 2b) as a model, we find e2qQ/h = 2.3 MHz for the pyridine 14N, in quite good accord with experiment.
Figure 2.
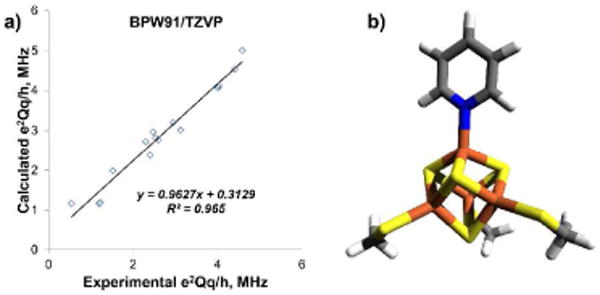
(a) Graph showing correlation between experimental and computed e2qQ/h values for a series of model systems; (b) Model used in quantum mechanical calculation of the 14N e2qQ/h value for IspH+7 complex.
This large decrease in e2qQ/h, from the 4.6 MHz value found for free pyridine to the 2.4 – 3.1 MHz values observed in model systems and the IspH + 7 complex, is also seen in proteins in which imidazole (histidine) ligands bind to Fe. For example, for imidazole (17) the N3 (deprotonated) e2qQ/h is 4.032 MHz,22 in good accord with the 3.894 MHz computed using DFT. The e2qQ/h values for solid imidazole and solid histidine are both smaller and essentially identical (3.27 MHz, Im; 3.36 MHz, His)23 – due presumably to very strong hydrogen bonding in the solid state. But when bound to Fe in metalloproteins, e2qQ/h decreases considerably from the 4 MHz gas phase value (for imidazole).
For example, in myoglobins, e2qQ/h ranges from 2.2 – 2.5 MHz for the directly bonded imidazole nitrogens;24-26 in the (Cys)3 (His)1 – coordinated [2Fe-2S] cluster in the human mito-NEET protein (18), e2qQ/h = (-) 2.47 MHz;27 and in several (Cys)2(His)2-coordinated [2Fe-2S] Rieske-type protein (19), e2qQ/h values have been reported to be in the range ∼2.2-2.9 MHz.28-30 Clearly then, the 14N nuclear quadrupole coupling constant decreases from ∼4 MHz for the free (gas phase) imidazole to ∼2.5 MHz when bound to Fe, similar to the decrease in e2qQ/h we find with pyr bound to Fe in the 4Fe-4S cluster of IspH.
These results all support the idea that the IspH pyridine inhibitors bind to IspH via a Lewis acid/base (Fe4S4 cluster/ligand) mechanism with the donor orbital occupancy (σ) decreasing from 2 (pyridine) to ∼1.73,20 and that the hyperfine coupling seen experimentally is due to this η1-bonding, rather than being due to a neutral pyr, or pyr-H+ ligand just being close to the 4Fe-4S cluster. This, in turn, suggests that stronger Lewis bases (such as imidazole-containing ligands) may be more potent IspH inhibitors. These results also support the idea that other inhibitors, such as alkynes,10, 13 as well as possible reaction intermediates (η3-allyls),10, 11 also act as Lewis bases, when interacting with the 4Fe-4S cluster in IspH.
Overall, these results are of interest for several reasons. First, we find evidence for an 14N HYSCORE signal when the pyridine inhibitor 7 binds to IspH. Based on isotopic labeling, this signal is assigned to the pyridine 14N. Second, the experimental e2qQ/h (from simulations of field-dependent HYSCORE) is 3 MHz. This is between the e2qQ/h = 4.6 MHz found for pyridine itself and e2qQ/h values of ∼1 MHz found in pyridinium salts and pyridine-N-oxide,20 and is in fact within the 2.4 – 3.1 MHz range of values found for pyridines bound to Cr, Mo and Fe carbonyls.23 So, while the ligand may initially bind as the cationic species (to e.g. E126), the η1-complex is the more stable species. Third, we report the results of DFT calculations of the 14N nuclear quadrupole coupling constant (e2qQ/h) in pyridine-containing metal systems finding a good correlation between theory and experiment (R2 = 0.965, slope = 0.963, Figure 2), in addition to predicting a 2.3 MHz e2qQ/h value for a [Fe4S4(SMe)3(pyr)]2- model cluster, in quite good accord with experiment (given that the protein was excluded from the calculation, and the crystallographic structure of the 4Fe-4S/pyridine protein-containing complex is not yet known). When all published experimental results on pyridine and imidazole-containing systems are considered, there is a ∼35 – 40% decrease in the 14N e2qQ/h on metal binding, the same as that found in the IspH + 7 system. This again supports formation of an η1-complex between IspH and 7, an observation of interest in the context of the design of other inhibitors, of interest as anti-infective drug leads.
Supplementary Material
Scheme 3.

Structures discussed in the text.
Acknowledgments
We thank Hassan Jomaa, and Jo-chen Wiesner for providing their IspH plasmid. This work was supported by the United States Public Health Service (NIH grants AI074233 and GM065307). W.W. was supported by a Predoctoral Fellowship from the American Heart Association, Midwest Affiliate (Award 10PRE4430022). EPR instrumentation used in this work was supported by NIH grants S10RR023614 and S10RR025438, NSF CHE-0840501, and NCBC 2009-IDG-1015. Computational research was supported by the National Science Foundation through Teragrid resources provided by NCSA under grant TG-CHE100060.
Footnotes
Supporting Information: Details on protein production and purification, HYSCORE sample preparation, Supporting Information figures, and full citation for reference 21 are reported in the Supporting Information available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Oldfield E. Acc Chem Res. 2010;43:1216–1226. doi: 10.1021/ar100026v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rohmer M. Lipids. 2008;43:1095–1107. doi: 10.1007/s11745-008-3261-7. [DOI] [PubMed] [Google Scholar]
- 3.Rohmer M, Grosdemange-Billiard C, Seemann M, Tritsch D. Curr Opin Investig Drugs. 2004;5:154–162. [PubMed] [Google Scholar]
- 4.Hecht S, Eisenreich W, Adam P, Amslinger S, Kis K, Bacher A, Arigoni D, Rohdich F. Proc Nat Acad Sci USA. 2001;98:14837–14842. doi: 10.1073/pnas.201399298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kollas AK, Duin EC, Eberl M, Altincicek B, Hintz M, Reichenberg A, Henschker D, Henne A, Steinbrecher I, Ostrovsky DN, Hedderich R, Beck E, Jomaa H, Wiesner J. FEBS Lett. 2002;532:432–436. doi: 10.1016/s0014-5793(02)03725-0. [DOI] [PubMed] [Google Scholar]
- 6.Seemann M, Bui BTS, Wolff M, Tritsch D, Campos N, Boronat A, Marquet A, Rohmer M. Angew Chem Int Ed Engl. 2002;41:4337–4339. doi: 10.1002/1521-3773(20021115)41:22<4337::AID-ANIE4337>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 7.Altincicek B, Duin EC, Reichenberg A, Hedderich R, Kollas AK, Hintz M, Wagner S, Wiesner J, Beck E, Jomaa H. FEBS Lett. 2002;532:437–440. doi: 10.1016/s0014-5793(02)03726-2. [DOI] [PubMed] [Google Scholar]
- 8.Wolff M, Seemann M, Bui BTS, Frapart Y, Tritsch D, Garcia Estrabot A, Rodriguez-Concepcion M, Boronat A, Marquet A, Rohmer M. FEBS Lett. 2003;541:115–120. doi: 10.1016/s0014-5793(03)00317-x. [DOI] [PubMed] [Google Scholar]
- 9.Wang W, Li J, Wang K, Huang C, Zhang Y, Oldfield E. Proc Natl Acad Sci USA. 2010;107:11189–11193. doi: 10.1073/pnas.1000264107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang W, Wang K, Liu YL, No JH, Nilges MJ, Oldfield E. Proc Natl Acad Sci USA. 2010;107:4522–4527. doi: 10.1073/pnas.0911087107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Grawert T, Span I, Eisenreich W, Rohdich F, Eppinger J, Bacher A, Groll M. Proc Natl Acad Sci USA. 2010;107:1077–1081. doi: 10.1073/pnas.0913045107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Batsanov S. Inorg Mater. 2001;37:871–885. [Google Scholar]
- 13.Wang K, Wang W, No JH, Zhang Y, Zhang Y, Oldfield E. J Am Chem Soc. 2010;132:6719–6727. doi: 10.1021/ja909664j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rekittke I, Wiesner J, Rohrich R, Demmer U, Warkentin E, Xu W, Troschke K, Hintz M, No JH, Duin EC, Oldfield E, Jomaa H, Ermler U. J Am Chem Soc. 2008;130:17206–17207. doi: 10.1021/ja806668q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Stoll S, Schweiger A. J Magn Reson. 2006;178:42–55. doi: 10.1016/j.jmr.2005.08.013. [DOI] [PubMed] [Google Scholar]
- 16.Fittipaldi M, Garcia-Rubio I, Trandafir F, Gromov I, Schweiger A, Bouwen A, Van Doorslaer S. J Phys Chem B. 2008;112:3859–3870. doi: 10.1021/jp709854x. [DOI] [PubMed] [Google Scholar]
- 17.Vinck E, Van Doorslaer S. Phys Chem Chem Phys. 2004;6:5324–5330. doi: 10.1039/b701568b. [DOI] [PubMed] [Google Scholar]
- 18.Dikanov SA, Shubin AA, Kounosu A, Iwasaki T, Samoilova RI. J Biol Inorg Chem. 2004;9:753–767. doi: 10.1007/s00775-004-0571-y. [DOI] [PubMed] [Google Scholar]
- 19.Lees NS, Hanzelmann P, Hernandez HL, Subramanian S, Schindelin H, Johnson MK, Hoffman BM. J Am Chem Soc. 2009;131:9184–9185. doi: 10.1021/ja903978u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Brown TL. Inorg Chem. 1980;19:392–398. [Google Scholar]
- 21.Frisch MJ, et al. Gaussian 09 (Revision A.01) see Supporting Information. [Google Scholar]
- 22.Palmer MH, Stephenson D, Smith JAS. Chem Phys. 1985;97:103–111. [Google Scholar]
- 23.Ashby CIH, Cheng CP, Brown TL. J Am Chem Soc. 1978;100:6057–6063. [Google Scholar]
- 24.Scholes CP, Lapidot A, Mascarenhas R, Inubushi T, Isaacson RA, Feher G. J Am Chem Soc. 1982;104:2724–2735. [Google Scholar]
- 25.Magliozzo RS, Peisach J. Biochemistry. 1992;31:189–99. doi: 10.1021/bi00116a028. [DOI] [PubMed] [Google Scholar]
- 26.Magliozzo RS, Peisach J. Biochemistry. 1993;32:8446–56. doi: 10.1021/bi00084a009. [DOI] [PubMed] [Google Scholar]
- 27.Dicus MM, Conlan A, Nechushtai R, Jennings PA, Paddock ML, Britt RD, Stoll S. J Am Chem Soc. 2010;132:2037–2049. doi: 10.1021/ja909359g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Britt RD, Sauer K, Klein MP, Knaff DB, Kriauciunas A, Yu CA, Yu L, Malkin R. Biochemistry. 1991;30:1892–1901. doi: 10.1021/bi00221a023. [DOI] [PubMed] [Google Scholar]
- 29.Shergill JK, Joannou CL, Mason JR, Cammack R. Biochemistry. 1995;34:16533–16542. doi: 10.1021/bi00051a001. [DOI] [PubMed] [Google Scholar]
- 30.Gurbiel RJ, Batie CJ, Sivaraja M, True AE, Fee JA, Hoffman BM, Ballou DP. Biochemistry. 1989;28:4861–7481. doi: 10.1021/bi00437a051. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.