

Abstract
MicroRNAs are a highly conserved class of small RNAs that function in a sequence-specific manner to posttranscriptionally regulate gene expression. Tissue-specific miRNA expression studies have discovered numerous functions for miRNAs in various aspects of embryogenesis, but a role for miRNAs in gonadal development and sex differentiation has not yet been reported. Using the chicken embryo as a model, microarrays were used to profile the expression of chicken miRNAs prior to, during, and after the time of gonadal sex differentiation (Embryonic Day 5.5 [E5.5], E6.5, and E9.5). Sexually dimorphic miRNAs were identified, and the expression patterns of several were subjected to further validation by in situ hybridization and Northern blot analysis. Expression of one chicken miRNA, MIR202*, was observed to be sexually dimorphic, with upregulation in the developing testis from the onset of sexual differentiation. Additional data from deep sequencing of male and female embryonic gonad RNA samples also indicated upregulation of MIR202* in male gonads. These findings provide the first evidence of sexually dimorphic miRNA expression during vertebrate gonadal sex differentiation and suggest that MIR202* may function in regulating testicular development.
Keywords: chicken, embryonic gonad, gene regulation, microRNAs, MIR202*, sex determination, testis
Sexually dimorphic microRNA expression is observed in male and female embryonic gonads, suggesting a role for microRNAs in vertebrate gonadal sex differentiation.
INTRODUCTION
MicroRNAs are highly conserved, small noncoding RNAs that account for approximately 1%–1.5% of all cellular transcripts and may target up to 30% of protein coding genes in animals [1, 2]. The latest release of the miRNA registry [3] (miRBase release 12.0, September 2008; http://microrna.sanger.ac.uk/sequences/) reports a total of 8619 miRNAs across plant and animal genomes [3]. MicroRNAs are expressed either as single transcription units or as polycistronic transcripts from miRNA clusters, encoded within intronic or intergenic regions of the genome [4]. RNA polymerase II drives transcription of miRNAs as inverted repeats embedded in long primary transcripts, which spontaneously fold to form imperfect long hairpins called primary or pri-miRNAs [4–6]. Primary miRNAs are processed into shorter hairpin precursor miRNAs, or pre-miRNAs, in the nucleus by the RNase III enzyme complex of DROSHA(RNASEN)/DGCR8 [7]. Precursor miRNAs are translocated into the cytoplasm by the Ran-guanosine triphosphatase, exportin 5 (XPO5) [8, 9]. In the cytoplasm, the RNase III enzyme DICER1 cleaves the pre-miRNA to form a double-stranded miRNA duplex comprising a mature miRNA (guide strand) and a partially complementary passenger, or “star” (*), strand [6, 10].
The mature miRNA associates with argonaute proteins in the RNA-induced silencing complex (RISC) to direct translational repression by binding to regions of complementarity in 3′ untranslated regions of target mRNAs [11, 12]. RISC loading has been shown to be largely asymmetric, with only a single strand of the miRNA duplex being incorporated to direct silencing [13, 14]. However, some miRNA duplexes encode mature miRNAs on both strands, and recent evidence suggests that strand bias in miRNA expression may be influenced by tissue-specific processing factors [15] or may be specific to individual miRNAs [16].
In plants, miRNA targets perfectly complementary binding sites, which commonly occur within the coding region of the mRNA. This perfect complementarity results in RISC-mediated cleavage of the target mRNA, which is then degraded [17]. Animal miRNA-binding sites, however, show only partial complementarity to the miRNA sequence and are located within 3′ untranslated regions of target mRNAs [18, 19]. Imperfect miRNA-target interaction causes translational repression of target mRNAs. More recently, miRNAs have also been reported to induce translational repression by interacting with imperfect binding sites within coding regions of target mRNAs [20]. Translational repression induced by miRNAs occurs through several possible mechanisms: RISC-mediated recruitment of deadenylation and decapping machinery to destabilize the mRNA; sequestering the mRNA into cytoplasmic P bodies via binding of trinucleotide repeat-containing 6A (TNRC6A; also known as GW182) to RISC components; or disrupting the binding of translation factors to inhibit translation initiation or elongation [18, 21, 22].
Many developmental and cellular differentiation processes are regulated by miRNAs, including limb development [23, 24], skeletal and cardiac myogenesis [25, 26], neuronal differentiation and patterning [27, 28], stem cell pluripotency [29], hematopoiesis [30], and brain development [31]. However, a role for miRNAs in gonadal development has not hitherto been reported.
The gonads are unique among embryonic organs because they follow one of two divergent pathways: ovary or testis formation. The chicken embryo (Gallus gallus) is a particularly good model for studying gonadal development because embryos are readily available and accessible for manipulation. The genome sequence is also available for the chicken, enhancing its use as a model for functional genomics studies in development. The catalog of chicken miRNA expression during embryonic development has also been characterized using deep sequencing [32], providing a strong background of information for analysis of tissue-specific miRNA signatures in chicken embryos.
Although sex in birds and mammals is determined genetically, the mammalian and avian sex chromosomes are not homologous [33]. In the chicken, the male is homogametic (ZZ), and the female is heterogametic (ZW). During gonadal development, genetic pathways are differentially activated in males and females to initiate testis or ovary development [34]. In mammals, the Y-linked gene, sex-determining region Y (SRY in human; Sry in mouse), is expressed from an early stage in male gonads and initiates testis development [35]. A number of other genes, including SRY-box 9 (SOX9/Sox9), encode factors that act downstream of SRY to coordinate male testicular differentiation [36]. Similarly, several female-determining factors, including R-spondin homolog (RSPO1/Rspo1), wingless-type MMTV integration site family, member 4 (WNT4/Wnt4), and forkhead box L2 (FOXL2/Foxl2), mediate ovary development [36–38]. The SRY/Sry homolog is absent from the chicken genome and those of other birds, and no equivalent avian primary sex-determining gene has yet been characterized [39]. Therefore, the mechanism of avian sex determination remains unknown. Gonadogenesis, however, is generally conserved at a morphological level between birds and mammals [40]. This implies that many of the underlying genetic control mechanisms are also likely to be conserved [41, 42].
In the chicken embryo, the gonads form on the ventral surface of the mesonephric kidney around Embryonic Day 3.5 (E3.5; Hamilton-Hamburger stage [HH] 19–20) [43]. The gonads remain bipotential (undifferentiated) in both sexes until E6.5 (HH 29–30), when the onset of sexual differentiation is first observed histologically. From E6.5 onward, seminiferous cords form in the medulla of the developing testes in the male, whereas thickening of the gonadal cortex occurs during ovarian development. In the male, the testes develop bilaterally, whereas the female gonads develop asymmetrically (only the left gonad becomes a functional ovary) [41]. Given the involvement of miRNAs in organogenesis and cellular differentiation [44], it is likely that miRNAs may also play some role in regulating embryonic testis and ovary differentiation.
To date, profiling of miRNA expression in reproductive tissues has been largely associated with characterizing molecular signatures of ovarian, prostate, and testicular germ cell cancers [45–47]. Several groups, however, have recently identified differential miRNA expression in postnatal mouse testes and ovary [48–50] and during spermatogenesis [51]. One recent study of miRNAs in the invertebrate urochordate Oikopleura dioica, the closest living group to vertebrates, has also reported sex-specific miRNA expression in embryonic gonads during sexual differentiation [52]. So far, however, miRNA expression in embryonic gonads at the time of sexual differentiation has not been reported for any vertebrate species.
Here, we report a microarray-based screen for miRNAs expressed in embryonic chicken gonads during sexual differentiation and demonstrate sexually dimorphic upregulation of chicken MIR202* in male gonads, coincident with testis differentiation. These findings provide the first evidence of sexually dimorphic miRNA expression in vertebrate embryonic gonads and suggest a potential role for MIR202* in testicular development.
MATERIALS AND METHODS
Gonad Tissue Collection and Embryo Sexing
All work using chicken embryos was conducted in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes, 7th edition, 2004, and in accordance with institutional animal ethics guidelines (Murdoch Children's Research Institute and Commonwealth Scientific and Industrial Research Organisation [CSIRO] Australian Animal Health Laboratory). Embryonated commercial chicken eggs (Ross 308 and Rhode Island Red/Australop, cross-strains) were obtained from Research poultry farm and Bartter Enterprises Pty. Ltd. and were incubated in a Multiquip E3 egg incubator (Bellsouth Pty. Ltd.). Gonads were dissected from the ventral surface of the mesonephric kidney of chicken embryos at E5.5, E6.5, and E9.5 in Petri dishes (BD Falcon) containing diethylpyrocarbonate-treated PBS (DEPC-PBS). Embryonic Day 9.5 embryos were sexed by gonad morphology, and gonads were pooled directly into 500 μl of miRNA Lysis/Binding buffer (LBB; mirVana miRNA Isolation kit; Ambion) and stored at 4°C for 24 h prior to miRNA extraction. Individual gonad pairs for E5.5 and E6.5 embryos were collected into 20 μl of LBB, vortexed for 10 sec, and lysed on ice or at 4°C for 12–24 h. For sexing, lower limb buds from each embryo were collected into 50 μl of PCR digestion buffer (50 mM KCl; 10 mM Tris-HCl, pH 8.3; 0.1 mg/ml gelatin; 0.45% Nonidet P-40; 0.45% Tween-20; 0.2 mg/ml proteinase K; stock stored at −20°C) at room temperature and digested at 55°C for a minimum of 1 h, then at 95°C for 10 min to release genomic DNA. Sexing was carried out by PCR using the method of Clinton et al. [53]. The PCR mix consisted of 1 μl of digestion mix, 10× RedTaq Reaction buffer (Sigma-Aldrich), MgCl2 to 1.5 mM (Promega), 1 unit of RedTaq DNA polymerase (Sigma-Aldrich), and Milli-Q water (Millipore) to a total volume of 20 μl. Reactions were carried out in Mastercycler S (Eppendorf) PCR machine. Products were run on 1.5% 1× Tris-borate EDTA (TBE) agarose gels.
MicroRNA Nomenclature
All miRNA names used in this paper conform to the Human Genome Organization (HUGO) Gene Nomenclature Committ xee Guidelines for Human Gene Nomenclature and, unless otherwise stated, refer to chicken miRNA sequences.
MicroRNA Extraction and Purification
All RNA extractions were carried out using the mirVana miRNA Isolation Kit (Ambion), using the protocol for fractionation and enrichment for small RNAs. Gonad lysates from E5.5 and E6.5 embryos were pooled according to sex, in 2-ml RNase-free tubes (Ambion) prior to RNA extraction. Both small RNA (miRNA, <200 nucleotides) and high-molecular weight fractions were collected for each sample and eluted in 100 μl of nuclease-free water (NF-H2O) (Ambion) preheated to 95°C. All RNA samples were stored at −20°C. RNA concentrations were determined using a NanoDrop 1000 spectrophotometer (Thermo Scientific).
Labeling of miRNA Samples for Microarray Analysis
For microarray hybridization, gonad miRNA samples were labeled with Cy5 (ULS small RNA labeling kit; Kreatech). Briefly, a 500-ng aliquot of miRNA sample was vacuum dried using SpeedVac (Savant) and was resuspended in 17 μl of NF-H2O (Promega). Labeling reactions containing 17 μl of miRNA sample, 1 μl of ULS-Cy5, and 2 μl of 10× labeling buffer were incubated at 85°C for 15 min, then on ice for 1 min. Excess label was removed by purification using KREApure columns (ULS small RNA labeling kit), according to the manufacturer's instructions. Labeled RNA concentration and Cy5 intensity were measured using a NanoDrop 1000 spectrophotometer.
Microarray Analysis
Custom-designed CustomArray 4X2K microarrays (chip design no. 4166; CombiMatrix) were used to analyze gonad miRNA profiles. Arrays featured 1454 DNA probes, including a nonredundant set of probes complementary to 630 vertebrate miRNAs from human (233); chicken (122); zebrafish (132); mouse (106); and rat (46; miRBase release 6.0 [3]), as well as probes for an additional 18 predicted chicken miRNA sequences (total chicken miRNA sequence probes = 140). Control probes for each miRNA sequence containing 2 nucleotide mutations (designated “2mut controls”) were also included. Positive control probes for chicken U6 small nuclear RNA (snRNA) and negative controls for bacteriophage lambda DNA were also included. The microarray probe identifiers are based on the miRBase nomenclatures, where human miRNAs are denoted by has-miR, mouse by mmu-miR, zebrafish by dre-miR, rat by rno-miR, and chicken by gga-miR. Where probes have been included for chicken miRNAs predicted from existing sequences identified in other species, the sequence is given the prefix “gga_candidate,” followed by the miRNA name from the original species (e.g., gga_candidate-dre-21–1). The “AS” suffix following the miRNA name indicates the probe sequence is antisense to the sequence of the given miRNA.
Solutions and protocols used for microarray hybridization were carried out according to the manufacturer's protocol for miRNA 4X2K microarrays (http://www.combimatrix.com/docs/PTL012_00_4x2K_MicroRNA_Hyb_Imaging.pdf). Cy5-labeled miRNA samples were concentrated to a volume of 5.1 μl or less using Microcon YM-30 columns (Millipore) and were hybridized to arrays at 37°C for 4–6 h.
Microarrays were scanned using an arrayWoRx e microarray scanner (Applied Precision) and arrayWoRx Biochip Reader software (version 2.1.4; Applied Precision). Five exposure-level images were captured for each array at 0.1, 0.2, 0.5, 1.0, and 2.0 sec, respectively. Image analysis and data extraction were carried out using Microarray Imager Software (version 5.7.1; CombiMatrix) and exported to Microsoft Office Excel (2003; Microsoft Corp.). Total array intensities for each array exposure were used to identify data sets with comparable levels of hybridization efficiency. Signal intensities for the selected data sets were then normalized. Background signal level (80%) was determined for each array, and background subtraction was applied to raw signal intensities and then normalized by multiplying by an arbitrary constant of 2 × 107.
Solexa Deep Sequencing of miRNAs in Embryonic Chicken Gonads
Gonads were dissected from E5.5, E6.5, and E9.5 embryos and sexed and pooled as described in Gonad Tissue Collection and Embryo Sexing. Total RNA was prepared from pooled male and female gonad lysates using the mirVana miRNA Isolation kit according to the manufacturer's instructions. Total RNA was eluted in 100 μl of NF-H2O and stored at −20°C. Solexa deep sequencing was carried out at The Leiden Genome Technology Center for Human and Clinical Genetics, Leiden University Medical Centre (Leiden, the Netherlands). Polyacrylamide gel electrophoresis (PAGE) was performed to size select 18- to 40-nucleotide RNAs. Sequencing reactions were run using the Solexa Genome Analysis System (Illumina). Read count data for chicken MIR202 and MIR202* were extracted from all reads featuring 3′ linker sequence tags per sample. Read counts for MIR202 and MIR202* were normalized to the total number of tagged reads per sample. Normalized read count ratios of miRNA reads:total tagged reads were then converted to whole numbers by multiplying by a constant value of 1 × 106 for each sample and were graphed using Microsoft Excel.
Locked Nucleic Acid Probe Design and Labeling
The 5′-digoxigenin (DIG)-labeled, predesigned miRCURY locked nucleic acid (LNA) probes for whole-mount in situ hybridization (WISH) detection of MIR101, MIR193B, MIR449, MIR202, and MIR202*, as well as unlabeled MIRscrambled probe, were obtained from Exiqon. Unlabeled MIR17-5P probe used for WISH was obtained from Sigma-Proligo. Probe sequences (LNA bases in probes are indicated in lowercase; LNA composition not provided for Exiqon probes) were: MIR17-5P, 5′-ACTAcCTgCAcTGtAAgCTcT-3′ (melting temperature [Tm] = 106°C); MIR101, 5′-CTTCAGTTATCACAGTACTGTA-3′ (Tm = 68°C; no. 377011–01); MIR202, 5′-TTTTCCCATGCTCTATGCCTCT-3′ (Tm = 75°C, no. 37111–01); MIR202*, 5′-AAAGAAGTATATGCATAGGAAA-3′ (Tm = 68°C, no. 37110–01); MIR193B 5′-AAAGCGGGACTTTGTGGGCCAGTT-3′ (Tm = 82°C, no. 37109–01); MIR449 5′-ACCAGCTAACATACACTGCCA-3′ (Tm = 76°C, no. 37128–01); and negative control MIRScrambled probe (unlabeled) 5′-TTCACAATGCGTTATCGGATGT-3′ (Tm = 74°C, no. 99001–00). The day before hybridization, probes were 3′ end labeled with DIG (DIG Oligonucleotide 3′-End Labeling Kit, 2nd generation; Roche Applied Sciences) in 100-pmol aliquots, according to the manufacturer's instructions. Labeled probes were stored at −20°C and thawed on ice immediately prior to addition to hybridization solutions.
For Northern blot analysis, additional unlabeled LNA probes for MIR202, MIR202*, and chicken U6 snRNA (cU6 snRNA) were obtained from Sigma-Proligo. Probe sequences were: MIR202, 5′-TTtCCcCATgCTcTATgCCtCt-3′; MIR202*, 5′-AAgAAgTAtATgCAtAGgAAa-3′; cU6 snRNA, 5′-TtTAgTAttATgTGcTG-3′. The LNA probes were labeled for Northern blotting with γ-32P-dATP (PerkinElmer) using Optikinase (USB Corp.). Labeling reactions contained: 1 μl of probe (10 pmol/μl), 2 μl of γ-32P-dATP, 2.5 μl of Optikinase 10× reaction buffer, 1 μl of Optikinase enzyme, and 18.5 μl of NF-H2O (Promega), incubated at 37°C for 30 min and then denatured at 100°C for 2 min.
Gonad WISH
For WISH, three to four embryonic chicken urogenital systems (UGSs) were used for each miRNA at each stage. The UGSs were dissected from E6.5 and E9.5 embryos, sexed by PCR, and fixed overnight in 4% paraformaldehyde (PFA) in DEPC-PBS. Fixed tissues were pooled by sex and washed two times for 10 min each in DEPC-PBS with 1% Triton X-100 (PBTX), dehydrated by a series of 20-min methanol/PBTX (MeOH/PBTX) washes (one time 25%, one time 50%, one time 75%, two times 100%) and stored in 100% MeOH at −20°C for 6–10 days. Tissues were rehydrated through reverse MeOH/PBTX wash series (20 min per wash), washed two times for 10 min each with PBTX, then treated sequentially with proteinase K in PBTX (10 μg/ml) at room temperature for 30 min (E6.5) or 45 min (E9.5), received two 5-min PBTX washes, and then were refixed in 0.2% glutaraldehyde in 4% PFA/PBS for 20 min.
Tissues were prehybridized at probe hybridization temperatures (22°C below probe Tm) for 2 h in 1.5–2 ml of hybridization solution (50% formamide; 5× saline-sodium citrate [SSC]; 2% blocking powder (Boehringer); 0.1% Triton X-100; 0.5% 3-[(cholamidopropyl)dimethylammonio]-1-propanesulfonate [CHAPS; Boehringer]; 1 ml of yeast RNA [no. R6750; Sigma]; 5 mM EDTA; and 50 μg/ml heparin). Prehybridized tissues were either hybridized immediately or stored at −20°C for 24–72 h. Prior to hybridization, stored prehybridized tissues were warmed to hybridization temperature for 2–4 h. DIG-labeled LNA probes were diluted to 2.5 pmol/μl and added to tissues at 2.5 μl per 1 ml of hybridization solution (typically 5 μl per tube of tissues). Hybridizations were carried out overnight at the following temperatures: MIR202*, 46°C; MIR101, 46°C; MIR202, 54°C; MIR193B, 60°C; MIR17-5P, 45°C, in rocking hybridization ovens. After hybridization, tissues were washed three times for 20 min in 2× SSC plus 0.1% CHAPS, and three times for 20 min in 0.2× SSC plus 0.1% CHAPS, followed by two 10-min washes in TBTX (50 mM Tris-HCl, pH 7.5; 150 mM NaCl; 0.1% Triton X-100), and tissues were blocked in TBTX/sheep serum (SS)/bovine serum albumin (BSA; 1× TBTX, 10% sheep serum [Sigma], and 2% BSA) for 2–3 h at room temperature. Anti-DIG-AP, Fab fragments (1:2000; Roche Applied Sciences) were prepared in TBTX/SS/BSA, added to tissues, and incubated overnight at 4°C. Postantibody washes (five times for 1 h and overnight) were performed using TBTX plus 0.1% BSA. For color development, tissues were washed two times, 30 min each, in TBTX, then three times, 10 min each, in fresh NTMT buffer (100 mM NaCl; 100 mM Tris-HCl, pH 9.5; 50 mM MgCl; and 0.1% Tween-20). Color solution was prepared with NTMT buffer and BCIP (175 μg/ml)/NBT (340 μg/ml; 5-bromo-4-chloro-3′-indolyphosphate p-toluidine/nitro blue tetrazolium chloride) chromogen. Tissues were incubated in color solution for 20 min in the dark, with rocking, then transferred to glass embryo dishes in dark boxes for 2–5 h. Images of whole-mount gonads were captured at 1–2 h (MIR202* and MIR202) or 4 h (MIR101, MIR449, MIR193B, MIR17-5P, and MIRscrambled) using a Leica MZ9 stereomicroscope fitted with a DFC290 color camera and IM1000 Image manager software (Leica Microsystems).
The WISH-stained tissues were prepared for cryosectioning by overstaining in NBT/BCIP color solution for 48–72 h, and they were incubated in 30% sucrose in DEPC-PBS at 4°C overnight before being frozen in OCT (TissueTek) on dry ice-ethanol slurry and stored at −80°C. Cryosections were prepared at 14 or 18 μm using a Leica cryostat and were mounted on Superfrost plus microscope slides (Thermo Scientific) with aqueous mounting medium (Aquatex; Merck). Images of sectioned WISH-stained gonads were captured at 200× or 400× magnification as 32-bit RGB images (2048 × 2048 pixels) using an Eclipse 80 inverted microscope (Nikon) fitted with a 10× ocular lens, 20× and 40× objective lenses, and a 14.2 color Mosaic camera (Diagnostic Instruments). Images were captured and scale bars added using SPOT Imaging Software (Advanced v4.6; Diagnostic Instruments).
For publication, whole-mount and sectioned WISH images were resized to 348 × 348 pixels (version 1.36b; Image J; National Institutes of Health; http://rsb.info.nih.gov/ij) and saved in TIFF format. For figure construction, TIFF images were opened in Microsoft Office PowerPoint (2003; Microsoft) and resized to 5.5 × 4.07 cm for whole-mount images and 4.07 × 4.07 cm for section images. Scale bars equal 500 μm for whole-mount images and 100 μm for sectioned images. Zoomed 400× images of sectioned gonads were prepared from full-sized original TIFF images cropped in PowerPoint to show indicated regions. Scale bars were reimposed onto the cropped image and saved as a separate TIFF. Cropped TIFF images were resized to 348 × 348 pixels (version 4.0; IrfanView, Irfan Skiljan) and saved as a new TIFF. Cropped-resized TIFF images opened in PowerPoint for figure construction and were resized to 4.07 × 4.07 cm.
Northern Blot
Gonad miRNA samples were extracted from 12–19 gonad pairs per sex from E5.5, E6.5, and E9.5 embryos, and 1-μg aliquots were concentrated to ≤15-μl volume using Microcon centrifugal filter columns and were combined with gel loading dye (2× RNA loading dye; Fermentas). Samples were separated against RNA Decade Markers (Ambion) on 15% Urea-PAGE gel (4.8 g of urea, 3.9 ml of acrylamide/bis 19:1 [Ambion], 2 ml of 5× TBE, 5 μl of N,N,N′,N′-tetramethylethylenediamine, and 50 μl of 10% ammonium persulfate) at 100 volts (V), 200 farad (F), 200 C, in 1× TBE buffer and transferred to Nylon Hybond N+ membrane (Amersham) using Trans-blot SD Semi-dry transfer cell (Bio-Rad) at 400 V, 500 F, for 1 h. Membrane was ultraviolet cross-linked twice (with 180° rotation) at 122 μJ × 100 (Stratalinker 1800; Stratagene). Membrane was prehybridized in 10 ml of prehybridization solution (50% formamide, 0.5% SDS, 5× sodium chloride, sodium phosphate-EDTA, and 5× Denhart solution plus 10 μl of 100 μg/ml salmon sperm DNA) for 1 h at 42°C in a rolling hybridization oven. Radiolabeled LNA probes for MIR202, MIR202*, or chU6 snRNA were then added to the prehybridization solution and incubated overnight at 42°C. Membranes were exposed to 13 × 18 cm Medical X-ray film (FUJIFILM) for 2–24 h at −80°C. Films were developed by an X-ray processor (FPM-100A; FUJIFILM) and scanned at 600 dpi using a flatbed scanner (PowerLook 1000; UMAX Technologies) and MagiScan Software (version 4.71; UMAX Technologies).
Relative fold changes in miRNA expression levels were calculated from comparison of Northern blot band intensities. Band intensities were calculated using the analyze gels function in ImageJ, and graphs were generated using Excel. Background signal was subtracted from all band intensity values. Negative values following background subtraction were made equal to 0. The U6 band intensities were adjusted to account for variation in amount of RNA loaded between samples by standardizing to highest U6 band intensity (i.e., if highest band intensity is 2000, then a band intensity of 1500 is standardized by: 1500/2000 = 0.75). Band signal intensities were then divided by standardized U6 intensity for each sample and multiplied by constant of 100 to obtain normalized intensity values. All calculations and graphs were generated using Microsoft Excel.
RESULTS
Microarray Profiling of Gonadal miRNA Expression
In the chicken, the onset of sexual differentiation is evident histologically at E6.5 (HH 30) [41]. To identify miRNAs that may regulate testis and ovary differentiation, male and female gonad miRNA samples from three stages (E5.5, E6.5, and E9.5) were analyzed by microarray. The microarray design included probes for a total of 657 chicken, human, mouse, zebrafish, and rat miRNA sequences (see Materials and Methods). Microarray analysis of male and female gonads from E5.5, E6.5, and E9.5 chicken embryos detected expression of 92 of 140 chicken miRNAs (Fig. 1) and 295 of the remaining 517 vertebrate (nonchicken) miRNAs (Supplemental Table S1 available at www.biolreprod.org; data also deposited in the Gene Expression Omnibus under accession no. GSE13154). Given the use of the chicken embryo as a model for gonad development, analysis of sexually dimorphic miRNA expression focused only on chicken miRNA expression data (Fig. 1 and Supplemental Table S2).
FIG. 1.

The miRNAs expressed in embryonic chicken gonads during sexual differentiation. Heat map displays all 92 miRNAs detected with expression in chicken embryonic gonads from E5.5, E6.5, and E9.5. Expression is color coded to reflect average normalized signal intensity per miRNA, per sex and stage. Males are shown on the left and females on the right. Signal intensity values represented by heat map are given in Supplemental Table S1 under the averages columns.
Similar numbers of miRNAs were expressed between the sexes at each time point (Supplemental Table S2). However, a higher number of miRNAs were expressed in both sexes at E9.5 compared with earlier stages (Supplemental Table S2). Sex-specific miRNA expression was observed at each stage, with a total of nine female-specific and seven male-specific miRNAs identified (Table 1). Only three chicken miRNAs showed sex-specific expression at multiple stages: MIR193B and MIR302A in male and MIR101 in female (Table 1). Three of the nine female-specific miRNAs identified were also Z-linked (MIR101, MIR449, and MIR7).
TABLE 1.
Sex-specific miRNA expression detected in chicken embryonic gonads.a

Across the three stages, 50 miRNAs expressed in both sexes showed greater than 2-fold differential expression between male and female gonads (Table 2). The average number of miRNAs differentially expressed between male and female gonads also varied between stages (Fig. 2). At E6.5, male gonads showed a significantly greater number of 2-fold upregulated miRNAs compared with females (P < 0.01). In contrast, by E9.5, female gonads showed greater miRNA upregulation compared with males (P < 0.01). These findings indicate that sexually dimorphic miRNA expression occurs during chicken gonadal sex differentiation.
TABLE 2.
Differentially expressed miRNAs in chicken embryonic gonads showing fold changes.a

FIG. 2.

Average number of miRNAs upregulated greater than 2-fold during gonadal differentiation. Averages are calculated from the number of miRNAs expressed 2-fold higher in either sex for at least three replicate gonad miRNA arrays per sex, per stage. Error bars represent SD. **P < 0.01 between grouped samples. Total number of miRNAs differentially upregulated in embryonic gonads was 50. Five of these miRNAs showed switching in upregulation between male and female at varying stages.
Identification of Sexually Dimorphic miRNA Candidates
Having confirmed differential miRNA expression during gonadal development, we sought to identify individual chicken miRNA candidates that may be involved in regulating sexual differentiation. Analysis of fold-change data for upregulated miRNAs revealed differential upregulation of MIR202 and MIR202* in male and female gonads, respectively (Table 2). This result was of particular interest because these two miRNAs are encoded by a single miRNA precursor (pre-MIR202*/202) [3]. Based on the microarray screening, MIR202 was male enriched across development all stages, whereas MIR202* was female enriched at E9.5 (Table 2).
These results suggested that MIR202 and MIR202* might be involved in gonadal differentiation. To investigate this further, MIR202 and MIR202* were examined using WISH and Northern blot approaches. A deep sequencing data set generated as part of a wider study of vertebrate gonadal miRNA expression (Bannister et al., unpublished results) was also used to extract data on the relative abundance of MIR202 and MIR202* in male and female gonads during sexual differentiation.
Validation of Sexually Dimorphic miRNA Expression During Gonadal Development
In addition to MIR202 and MIR202*, three miRNAs found to be sex specific by microarray (Table 1) were chosen for validation by WISH (Figs. 3 and 4): MIR101 (female specific, multiple-stage expression, Z-linked); MIR449 (female specific, Z-linked); and MIR193B (male specific, multiple-stage expression). For a positive control, MIR17-5P was chosen. This miRNA has been shown previously by in situ hybridization to have widespread expression in chicken embryos [54]. A scrambled miRNA probe was used as a negative control.
FIG. 3.
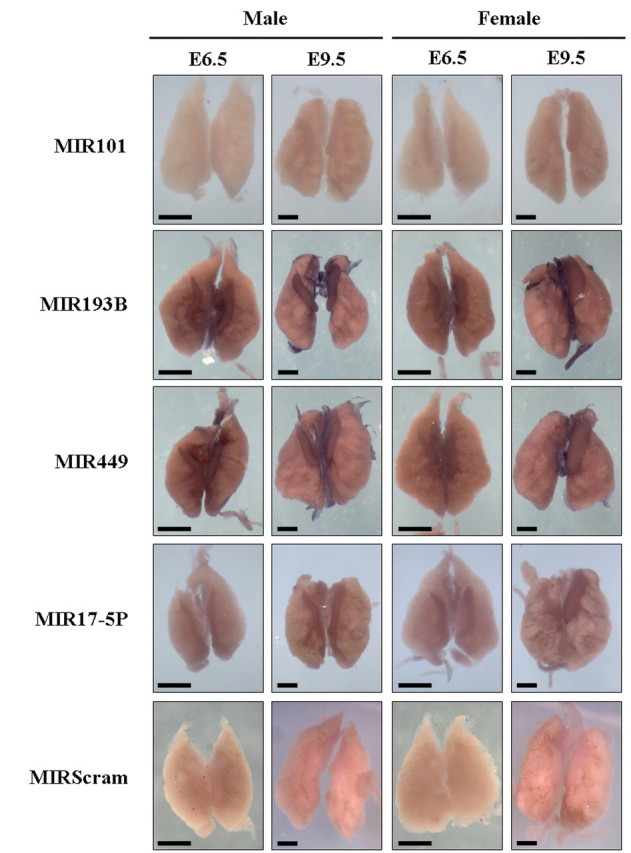
Analysis of MIR101, MIR449, and MIR193B expression by WISH. Whole-mount chicken UGSs from E6.5 and E9.5 embryos probed with LNA probes for MIR101, MIR193B, MIR499, MIR17-5P (positive control), and MIRscrambled (miR-Scram; negative control). Positive detection of miRNA expression is indicated by blue/purple staining. Images were taken approximately 4 h after addition of color solution. Bars = 500 μm.
FIG. 4.
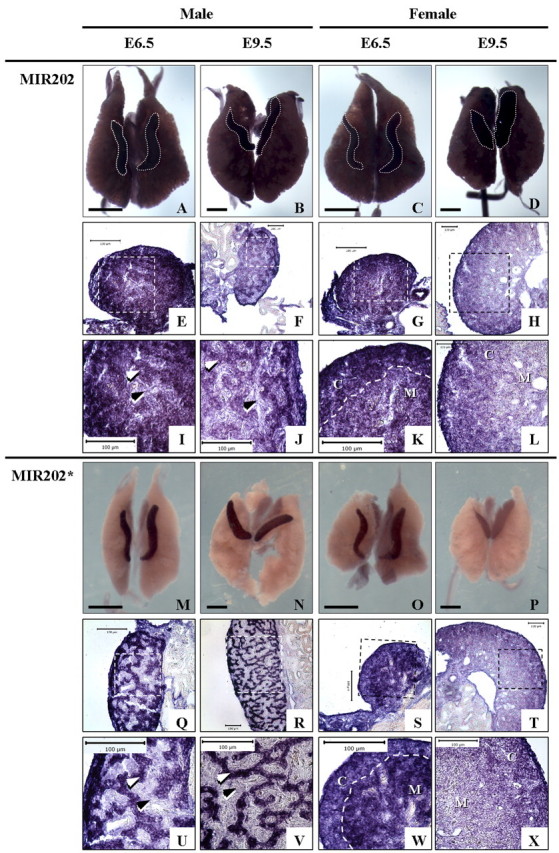
Expression of MIR202 and MIR202* detected by WISH. Whole-mount and transverse-sectioned UGSs showing detection of MIR202 (A–L) and MIR202* (M–X). Gonads in MIR202 whole mount are outlined in gray dashed lines. In female E6.5 panels, white dashed line separates cortex (C) and medulla (M). White arrowheads indicate testis cords, and black arrowheads indicate interstitium. Bars = 500 μm (whole mount) and 100 μm (sections). Area represented in zoomed, high-power (original magnification ×400) images (I–L and U–X) of sectioned gonads is indicated by white/black dashed-line box in low-power/ unzoomed (E6.5, original magnification ×400; E9.5, original magnification ×200) images (E–H, Q–T).
WISH was performed on intact embryonic chicken UGSs (kidneys and gonads) from E6.5 and E9.5 embryos using miRNA-specific LNA probes (Figs. 3 and 4). Expression of MIR17-5P was similar in both male and female gonad and kidney tissues, and no staining was detected in tissues hybridized with the MIRscrambled probe (Fig. 3). In general, all of the miRNAs examined by WISH (except MIR202; see below) showed higher expression in the gonads than in the surrounding embryonic kidneys (Fig. 3). Expression of MIR101 was weak in E9.5 male and female E6.5 gonads and not detectable in male E6.5 gonads.
In general, MIR449 and MIR193B were detected at low levels, but higher levels than MIR101. Expression of both MIR449 and MIR193B also increased from E6.5 to E9.5 (Fig. 3). In contrast to the microarray findings, which suggested sex specificity, both miRNAs were detected in both sexes at both stages. Negligible dimorphic expression was detected in male gonads at E6.5 for MIR449, but no difference between male and female was seen at E9.5. Expression of MIR193B did appear to be slightly higher in male gonads at E9.5 and higher in the left than the right female gonad at E9.5 (Fig. 3). Sectioned gonads for MIR101, MIR193B, and MIR449, however, did not indicate any significant differences in expression between male and female gonads (Supplemental Fig. S1). Taken together, these results suggest that MIR101 may be somewhat female specific at E6.5, and that MIR449 and MIR193B expression does not appear to be sexually dimorphic in embryonic chicken gonads.
Male Upregulation of MIR202*
WISH was also used to further examine MIR202 and MIR202* expression. MIR202 was expressed throughout the gonads and embryonic kidneys in both sexes (Fig. 4, A–D). Overstained and sectioned tissues revealed that MIR202 expression was preferentially localized in the cortex of female gonads (Fig. 4, G, H, K, and L). In male gonads, expression was stronger in the developing seminiferous (testis) cords than interstitium (Fig. 4, E, F, I, and J). In contrast, MIR202* expression was detected specifically in the gonads and not in the surrounding kidneys (Fig. 4, M–P). Expression was clearly higher in male versus female gonads at both stages examined (Fig. 4, compare M and N to O and P). In overstained sections for female gonads, MIR202* was detected throughout the gonad, with stronger expression detected in the medulla at E6.5 (Fig. 4, S and W) but preferentially localized within the cortex at E9.5 (Fig. 4, T and X). In males, MIR202* expression was restricted to the developing testis cords (Fig. 4, Q, R, U, and V). These results further suggest that expression of MIR202 and MIR202* may be subject to specific regulation during gonadal sex differentiation in the chicken embryo.
Analysis of deep sequencing data was used to verify the relative abundance of mature MIR202 and MIR202* sequences expressed between male and female embryonic gonads during sexual differentiation. These data reported similar numbers of normalized sequence reads (abundance) for mature MIR202 in male and female gonads at E5.5 and E6.5 (Fig. 5). At E9.5, read counts for MIR202 were 2.4-fold higher in male gonads (Fig. 5). However, for MIR202*, normalized sequence reads were higher in male versus female gonads at all stages (Fig. 5). By fold-change, MIR202* was upregulated in males 1.1-fold at E5.5, 1.5-fold at E6.5, and 2.6-fold at E9.5. The deep sequencing, therefore, supported the WISH data for MIR202 and MIR202*.
FIG. 5.

Deep sequencing profile of MIR202* and MIR202 expression in chicken embryo gonads. Normalized read counts are shown for MIR202* (black bars) and MIR202 (gray bars) in male and female gonad RNA samples. Normalized read counts were calculated by dividing the number of raw read counts for exact miRBase sequences of MIR202* and MIR202 by the total number of tagged reads obtained, per sample. Ratio of miRNA:total tagged reads was then converted to whole numbers by multiplying by a constant value of 1 × 106.
Expression patterns of MIR202 and MIR202* were further analyzed by Northern blotting (Fig. 6). Small RNA samples from male and female embryonic gonads at E5.5, E6.5, and E9.5 and male and female E9.5 embryonic kidneys were probed using γ-32P-labeled LNA probes to detect mature MIR202 and MIR202*. Chicken U6 snRNA LNA probe was used as a loading control. Analysis of MIR202 by Northern blot detected a major band at 28 nucleotides and a faint band at approximately 24 nucleotides (Fig. 6a). The 24-nucleotide band appeared weakly present in all samples, but no measurable expression in any samples could be resolved after normalization (data not shown). The 28-nucleotide product was detected in gonad and embryonic kidney samples (Fig. 6c). After normalization, the 28-nucleotide product appeared to be expressed in female gonads at all stages but only detectable in male gonads at E9.5. Both the 24- and 28-nucleotide sequences are longer than the mature 22-nucleotide MIR202 sequence defined by miRBase. The major 28-nucleotide product, however, was detected repeatedly in favor of any smaller products; therefore, it was determined that this product was representative of MIR202.
FIG. 6.
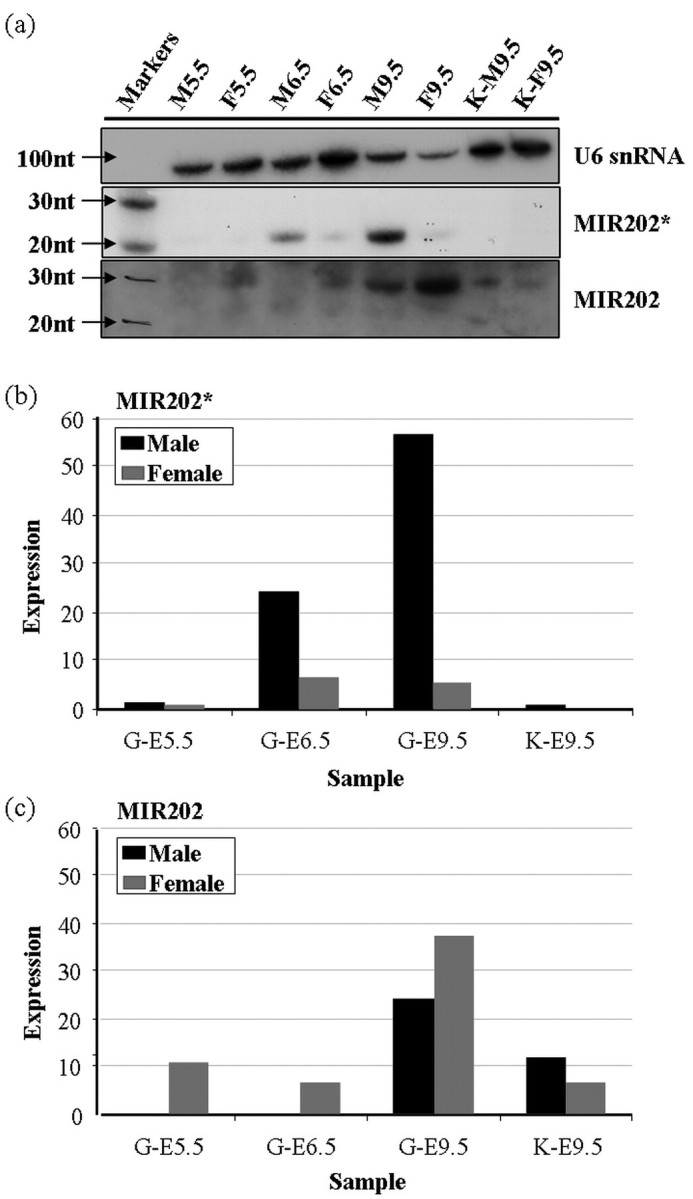
Northern blot detection of MIR202 and MIR202* expression. RNA samples purified for miRNA fractions from male and female chicken embryonic gonads (G) at E5.5, E6.5, and E9.5, and at E9.5 in embryonic kidney (K). a) Northern blot (1 μg RNA/lane) probed with LNA for MIR202* and MIR202, and chicken U6 snRNA probed as loading control. b) Normalized expression of MIR202* in male versus female gonad and kidney samples, as detected by Northern blot, showing male upregulation at E6.5 and E9.5. c) Normalized expression of MIR202 as detected by Northern blot. Normalized values are calculated as a ratio of signal for chicken U6 snRNA loading control per sample and multiplied by a constant of 100 to give whole numbers. nt, nucleotide.
Using the MIR202* LNA probe, a 21-nucleotide band was clearly detected in gonad samples but not in kidney samples. This band corresponded to the size of mature MIR202* defined by miRBase. Expression of 21-nucleotide MIR202* was weak but similar in E5.5 male and female samples, but it was clearly male upregulated at E6.5 and E9.5 (Fig. 6, a and b). Comparison of MIR202* band intensities indicated expression of MIR202* was 3.9-fold higher in males at E6.5 and 11.1-fold higher at E9.5 (Fig. 6b). This result supported the WISH and deep sequencing data, suggesting male upregulation of MIR202* occurs in chicken embryonic gonads from the onset of sexual differentiation at E6.5.
DISCUSSION
Expression profiling of miRNAs in embryonic testis and ovary has not been reported previously for any vertebrate species. Therefore, we sought to characterize male and female miRNA profiles in embryonic gonads using the chicken embryo as a vertebrate model. Our data demonstrate sexually dimorphic miRNA expression in embryonic chicken gonads—the first reported for any vertebrate species. Using microarray analysis, 50 (35.7%) of 140 known chicken miRNAs included on the microarray were observed to be sexually dimorphic (Table 2). We identified 7 (5%) of 140 miRNAs to be male specific and 9 (6.4%) of 140 to be female specific (Table 1).
These proportions of sex-specific miRNA expression in testis are somewhat similar to those reported previously from cloning and RT-PCR analyses of mouse postnatal testis miRNA expression. Ro et al. [49] used RT-PCR to analyze expression of 122 miRNAs cloned from postnatal mouse testis tissue and showed that 5% of these were testis specific. Mishima et al. [50] also reported a comprehensive comparison of adult mouse testis and ovary miRNA profiles derived by miRNA cloning analysis. They obtained 49 testis-only and 48 ovary-only cloned miRNAs and confirmed differential expression of 11 miRNAs between testis and ovary using RT-PCR. Data published by Landgraf et al. [55] also demonstrate differential expression of miRNAs between adult mouse testis and ovary, as do data from human tissue miRNA profiling [56]. Sequencing analysis of miRNAs in platypus has also identified a novel set of testis-expressed miRNAs encoded on the X1 sex chromosome [57]. Sex-specific miRNA expression is reported to be coincident with gonadal sex differentiation and spermatogenesis in the invertebrate urochordate species Oikopleura dioica [52]. Interestingly, urochordates are the closest living invertebrate lineage to vertebrates [52] which, taken together with our data, suggests that sexually dimorphic miRNA expression during gonadal development may have originated in invertebrates and has been retained in higher vertebrates. Given the evidence for sexually dimorphic miRNA expression in postnatal mammalian testis and ovary, it is also likely that sexually dimorphic miRNA expression is present in mammalian gonads during embryogenesis. This, however, has not yet been reported but could be investigated through comparison of miRNA profiles in chicken and mouse embryonic gonads. This may also shed light on the evolution of gene regulatory networks involved in mediating vertebrate sex determination.
Microarray analysis revealed dynamic variation in miRNA expression between male and female embryonic chicken gonads (Fig. 2). A greater number of miRNAs were upregulated in male than female gonads at E6.5, a pattern that was reversed at E9.5. This shift in miRNA upregulation may reflect differences in tissue reorganization occurring in testis and ovary differentiation between E6.5 and E9.5. Evidence from Dicer knockout mice suggests that ovarian miRNA expression is important for normal uterine and oviduct development in the female reproductive system [58, 59]. Similarly, miRNAs are important for proliferation of mouse primordial germ cells and spermatogonia [60]. These and other roles for miRNAs in cellular differentiation and developmental pathways [44], taken together with the observed temporal changes in male and female miRNA expression dynamics, provide support for miRNA involvement in gonadal sex differentiation.
We sought to validate a number of miRNAs identified by microarray as being sex specific or sexually dimorphic. Of those tested further by WISH, however, the majority (gga-MIR101, gga-MIR449, and gga-MIR193B) did not show convincing patterns of sex-specific or sexually dimorphic expression. Expression of MIR101 did appear to be female specific at E6.5, as indicated by microarray. But given the detection in both sexes at E9.5 and the weak level of expression in comparison with negative control tissues, it is unlikely that gga-MIR101 is genuinely sex specific. Similarly, despite observing slightly higher expression of gga-MIR193B in male versus female E9.5 gonads, the difference was not considered to be significant enough to warrant further investigation.
However, MIR202 and MIR202* expression profiles were sexually dimorphic, as assessed by different techniques (WISH, Northern blotting, and/or deep sequencing). Concordant results from deep sequencing, WISH, and Northern blotting showed that chicken MIR202* is expressed in a gonad-specific manner and is upregulated in male gonads from E6.5 (Figs. 4 and 5). Although the deep sequencing, WISH, and Northern blotting indicated male-specific upregulation of MIR202*, this was not seen by microarray analysis. Compared with microarrays, deep sequencing approaches provide a greater depth of coverage for miRNA expression analysis by not being restricted to a designated set of probes. Deep sequencing provides a read count for the relative abundance of individual sequences present within a sample; in this case, miRNAs.
Analysis of miRNA expression by deep sequencing approaches has proven a powerful tool in revealing new miRNAs in defined cells and tissues [57] and in miRNA expression profiling of stem cell differentiation states [61] and stages of chicken embryo development [32]. Since initiation of this study, the number of miRNAs characterized in the chicken has risen from 149 to 475 [3] through application of deep sequencing [32]. We have initiated similar studies using our experimental system, and preliminary analysis of these early data sets for MIR202 and MIR202* support the current observations. Sequencing approaches also limit bias introduced through the need to label miRNA samples for microarray analysis. In our case, the labeling process is biased toward miRNAs with higher guanosine (G) content (Kreatech). This may explain the discrepancy between microarray, WISH, and deep sequencing for MIR202*, because it contains only one G residue, whereas MIR202 contains eight [3]. Thus, the level of MIR202* expression is potentially underrepresented in the microarray data. Collectively, these findings highlight the need to employ multiple approaches for experimental validation of miRNA expression patterns. In this case, further screening using real-time PCR could also be used for verifying sexually dimorphic expression of the remaining candidates identified in this study.
WISH revealed specific localization of MIR202* within the testis cords of male gonads, with lower, more widespread expression in female gonads. Both the MIR202 and MIR202* sequences are highly conserved among human, mouse, rat, zebrafish, and puffer fish (Fugu rubripes) [3], yet the biological functions of MIR202 and MIR202* in any of these species have not been specified. Based on in situ localization of expression, our results indicate that the function of chicken MIR202* may be gonad specific and involved primarily in regulating testicular differentiation. By contrast, MIR202 was expressed throughout the gonad and adjoining mesonephric kidney, suggesting its function may be less specific to gonadal development.
In mouse, expression of CHICK MIR202* homolog in mouse, MIR202-5P, has been analyzed in postnatal gonads, where testis expression was significantly higher than ovary [15]. Similarly, MIR202-5P was among the most abundant miRNAs cloned and sequenced in postnatal mouse testis [50]. Expression of CHICK MIR202 homolog in mouse, MIR202-3P, is also enriched in postnatal mouse testis and ovary but, unlike MIR202-5P, is clearly detected in a range of other tissues [15]. These results compare closely to our own, indicating several aspects of MIR202* expression to be similar between embryonic chicken and postnatal mouse gonads. First, that MIR202* expression within the embryonic urogenital system is highly enriched or is specifically restricted to the gonads. Second, MIR202* is expressed more highly in male versus female gonads. Lastly, that sexually dimorphic expression is more pronounced for MIR202* than for MIR202. These collective observations suggest there may conservation of MIR202* function in avian and mammalian gonadal development. More specifically, male upregulation of MIR202* may play a role in repressing genes involved in female development or maintaining threshold levels of gene expression crucial for male testicular development.
Different spatial expression of MIR202* and MIR202 in sectioned WISH-stained gonads shows that MIR202* expression is more tightly regulated than MIR202. At both E6.5 and E9.5, expression of MIR202 was seen in testis cords and, more weakly, in the interstitium, but MIR202* expression was only detected in testis cords (Fig. 4). Expression of MIR202* is also spatially regulated in female gonads, where expression was preferential in the medulla at E6.5 but stronger in the cortex at E9.5. By comparison, MIR202 expression was preferentially cortical at both stages. Thus, our findings indicate that MIR202* is subject to tighter spatial and sexual regulation than MIR202 during embryonic gonadal development.
The different spatial expression of MIR202 and MIR202* observed in developing testes may be mediated by a selective processing mechanism. The majority of miRNA precursors encode and express only a single mature miRNA, either the 5P or 3P strand of the hairpin duplex. The selected strand is normally that with the lower thermodynamic stability at the 5′ end of the sequence [13, 14]. The MIR202 precursor, and an increasing number of other pre-miRNAs, show more uniform 5′ and 3′ thermodynamic stability and exhibit paired miRNA expression (both 5P and 3P sequences encode miRNAs). For some of these pre-miRNAs, Ro et al. [15] have shown that strand bias in miRNA expression does occur and is regulated in a tissue-dependent manner. In our analysis, MIR202* expression was more restricted than that of MIR202. Therefore, processing of mature MIR202* from pre-MIR202*/202 may be selectively mediated by a gonad-specific factor(s) enriched in male testis cord cells. Alternatively, MIR202* expression may be suppressed in the majority of tissues by some factor and expressed only in tissues where this factor is absent. Consistent with the findings of Ro et al. [15], our data suggest that tissue-specific processing factors are probably involved in spatially and sexually regulating differential expression of paired miRNAs. Therefore, in addition to identifying the function of MIR202* in embryonic gonads, the apparent differences in pre-MIR202*/202 processing we have detected between embryonic gonad and kidney might serve as a useful model for investigating mechanisms of tissue-specific miRNA processing.
Our study constitutes the first analysis of sexually dimorphic miRNA expression in the gonads of a vertebrate embryo. Using the chicken embryo as a model, we have identified differential miRNA upregulation between male and female gonads coinciding with testis and ovary differentiation. We have also validated sexually dimorphic expression of chicken MIR202*, showing that it is upregulated in male gonads from the onset of sexual differentiation. Expression of MIR202* localizes to testis cords in male gonads and has a tighter spatial expression profile compared with MIR202. Although MIR202* upregulation coincides with gonadal sex differentiation, it is not yet clear whether MIR202* expression is involved in driving the progression of sexual differentiation, or whether its upregulation is a consequence of sexual differentiation. Identification of potential gene targets and functional studies are now required to examine the role of MIR202* in gonadal development.
Supplementary Material
Acknowledgments
We acknowledge the work of Dr. Rob Moore and Dr. Evgeny Glazov (CSIRO) in the design of the miRNA microarrays. We also acknowledge assistance provided by Ivano Broz (CSIRO) in extracting expression data for MIR202 and MIR202* from deep sequencing data. Thank you to Dr. Pauline Cottee for assistance with Northern blotting and to Dr. Volker Haring for critical appraisal of the manuscript.
Footnotes
1Supported by funding from CSIRO Emerging Sciences Initiative (M.L.V.T.), and the Australian Research Council Discovery Grant to C.A.S. and A.H.S. Microarray data were deposited in the National Center for Biotechnology Information Gene Expression Omnibus (GEO) database under the accession number GSE13154.
REFERENCES
- Lee RC, Ambros V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001; 294: 862 864 [DOI] [PubMed] [Google Scholar]
- Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005; 120: 15 20 [DOI] [PubMed] [Google Scholar]
- Griffiths-Jones S. The microRNA Registry. Nucleic Acids Res 2004; 32: D109 D111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y, Jeon K, Lee JT, Kim S, Kim VN. MicroRNA maturation: stepwise processing and subcellular localization. EMBO J 2002; 21: 4663 4670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH, Kim VN. MicroRNA genes are transcribed by RNA polymerase II. EMBO J 2004; 23: 4051 4060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutvagner G, McLachlan J, Pasquinelli AE, Balint E, Tuschl T, Zamore PD. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001; 293: 834 838 [DOI] [PubMed] [Google Scholar]
- Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, Lee J, Provost P, Radmark O, Kim S, Kim VN. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003; 425: 415 419 [DOI] [PubMed] [Google Scholar]
- Yi R, Qin Y, Macara IG, Cullen BR. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev 2003; 17: 3011 3016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund E, Guttinger S, Calado A, Dahlberg JE, Kutay U. Nuclear export of microRNA precursors. Science 2004; 303: 95 98 [DOI] [PubMed] [Google Scholar]
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001; 409: 363 366 [DOI] [PubMed] [Google Scholar]
- Hammond SM, Bernstein E, Beach D, Hannon GJ. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 2000; 404: 293 296 [DOI] [PubMed] [Google Scholar]
- Hammond SM, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 2001; 293: 1146 1150 [DOI] [PubMed] [Google Scholar]
- Schwarz DS, Hutvagner G, Du T, Xu Z, Aronin N, Zamore PD. Asymmetry in the assembly of the RNAi enzyme complex. Cell 2003; 115: 199 208 [DOI] [PubMed] [Google Scholar]
- Khvorova A, Reynolds A, Jayasena SD. Functional siRNAs and miRNAs exhibit strand bias. Cell 2003; 115: 209 216 [DOI] [PubMed] [Google Scholar]
- Ro S, Park C, Young D, Sanders KM, Yan W. Tissue-dependent paired expression of miRNAs. Nucleic Acids Res 2007; 35: 5944 5953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guil S, Caceres JF. The multifunctional RNA-binding protein hnRNP A1 is required for processing of MIR18a. Nat Struct Mol Biol 2007; 14: 591 596 [DOI] [PubMed] [Google Scholar]
- Llave C, Xie Z, Kasschau KD, Carrington JC. Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 2002; 297: 2053 2056 [DOI] [PubMed] [Google Scholar]
- Pillai RS, Bhattacharyya SN, Filipowicz W. Repression of protein synthesis by miRNAs: how many mechanisms? Trends Cell Biol 2007; 17: 118 126 [DOI] [PubMed] [Google Scholar]
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993; 75: 855 862 [DOI] [PubMed] [Google Scholar]
- Tay Y, Zhang J, Thomson AM, Lim B, Rigoutsos I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008; 455: 1124 1128 [DOI] [PubMed] [Google Scholar]
- Rehwinkel J, Behm-Ansmant I, Gatfield D, Izaurralde E. A crucial role for GW182 and the DCP1:DCP2 decapping complex in miRNA-mediated gene silencing. RNA 2005; 11: 1640 1647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behm-Ansmant I, Rehwinkel J, Doerks T, Stark A, Bork P, Izaurralde E. mRNA degradation by miRNAs and GW182 requires both CCR4:NOT deadenylase and DCP1:DCP2 decapping complexes. Genes Dev 2006; 20: 1885 1898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hornstein E, Mansfield JH, Yekta S, Hu JK, Harfe BD, McManus MT, Baskerville S, Bartel DP, Tabin CJ. The microRNA MIR196 acts upstream of Hoxb8 and Shh in limb development. Nature 2005; 438: 671 674 [DOI] [PubMed] [Google Scholar]
- Yekta S, Shih IH, Bartel DP. MicroRNA-directed cleavage of HOXB8 mRNA. Science 2004; 304: 594 596 [DOI] [PubMed] [Google Scholar]
- Zhao Y, Samal E, Srivastava D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature 2005; 436: 214 220 [DOI] [PubMed] [Google Scholar]
- Chen JF, Mandel EM, Thomson JM, Wu Q, Callis TE, Hammond SM, Conlon FL, Wang DZ. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat Genet 2006; 38: 228 233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawasaki H, Taira K. Hes1 is a target of microRNA-23 during retinoic-acid-induced neuronal differentiation of NT2 cells. Nature 2003; 423: 838 842 [DOI] [PubMed] [Google Scholar]
- Johnston RJ, Hobert O. A microRNA controlling left/right neuronal asymmetry in Caenorhabditis elegans. Nature 2003; 426: 845 849 [DOI] [PubMed] [Google Scholar]
- Houbaviy HB, Murray MF, Sharp PA. Embryonic stem cell-specific MicroRNAs. Dev Cell 2003; 5: 351 358 [DOI] [PubMed] [Google Scholar]
- Chen CZ, Li L, Lodish HF, Bartel DP. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004; 303: 83 86 [DOI] [PubMed] [Google Scholar]
- Giraldez AJ, Cinalli RM, Glasner ME, Enright AJ, Thomson JM, Baskerville S, Hammond SM, Bartel DP, Schier AF. MicroRNAs regulate brain morphogenesis in zebrafish. Science 2005; 308: 833 838 [DOI] [PubMed] [Google Scholar]
- Glazov EA, Cottee PA, Barris WC, Moore RJ, Dalrymple BP, Tizard ML. A microRNA catalog of the developing chicken embryo identified by a deep sequencing approach. Genome Res 2008; 18: 957 964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridolfsson AK, Cheng H, Copeland NG, Jenkins NA, Liu HC, Raudsepp T, Woodage T, Chowdhary B, Halverson J, Ellegren H. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proc Natl Acad Sci U S A 1998; 95: 8147 8152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merchant-Larios H, Moreno-Mendoza N. Onset of sex differentiation: dialog between genes and cells. Arch Med Res 2001; 32: 553 558 [DOI] [PubMed] [Google Scholar]
- Sinclair AH, Berta P, Palmer MS, Hawkins JR, Griffiths BL, Smith MJ, Foster JW, Frischauf AM, Lovell-Badge R, Goodfellow PN. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990; 346: 240 244 [DOI] [PubMed] [Google Scholar]
- Wilhelm D, Palmer S, Koopman P. Sex determination and gonadal development in mammals. Physiol Rev 2007; 87: 1 28 [DOI] [PubMed] [Google Scholar]
- Parma P, Radi O, Vidal V, Chaboissier MC, Dellambra E, Valentini S, Guerra L, Schedl A, Camerino G. R-spondin1 is essential in sex determination, skin differentiation and malignancy. Nat Genet 2006; 38: 1304 1309 [DOI] [PubMed] [Google Scholar]
- Tomizuka K, Horikoshi K, Kitada R, Sugawara Y, Iba Y, Kojima A, Yoshitome A, Yamawaki K, Amagai M, Inoue A, Oshima T, Kakitani M. R-spondin1 plays an essential role in ovarian development through positively regulating Wnt-4 signaling. Hum Mol Genet 2008; 17: 1278 1291 [DOI] [PubMed] [Google Scholar]
- Smith CA, Roeszler KN, Hudson QJ, Sinclair AH. Avian sex determination: what, when and where? Cytogenet Genome Res 2007; 117: 165 173 [DOI] [PubMed] [Google Scholar]
- Smith CA, Sinclair AH. Sex determination in the chicken embryo. J Exp Zool 2001; 290: 691 699 [DOI] [PubMed] [Google Scholar]
- Smith CA, Sinclair AH. Sex determination: insights from the chicken. Bioessays 2004; 26: 120 132 [DOI] [PubMed] [Google Scholar]
- Morais da Silva S, Hacker A, Harley V, Goodfellow P, Swain A, Lovell-Badge R. Sox9 expression during gonadal development implies a conserved role for the gene in testis differentiation in mammals and birds. Nat Genet 1996; 14: 62 68 [DOI] [PubMed] [Google Scholar]
- Hamburger V, Hamilton HL. A series of normal stages in the development of the chicken embryo. J Morphol 1951; 88: 49 92 [PubMed] [Google Scholar]
- Zhang B, Wang Q, Pan X. MicroRNAs and their regulatory roles in animals and plants. J Cell Physiol 2007; 210: 279 289 [DOI] [PubMed] [Google Scholar]
- Iorio MV, Visone R, Di Leva G, Donati V, Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, Calin GA, Menard S, et al. MicroRNA signatures in human ovarian cancer. Cancer Res 2007; 67: 8699 8707 [DOI] [PubMed] [Google Scholar]
- Ambs S, Prueitt RL, Yi M, Hudson RS, Howe TM, Petrocca F, Wallace TA, Liu CG, Volinia S, Calin GA, Yfantis HG, Stephens RM, et al. Genomic profiling of microRNA and messenger RNA reveals deregulated microRNA expression in prostate cancer. Cancer Res 2008; 68: 6162 6170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voorhoeve PM, le Sage C, Schrier M, Gillis AJ, Stoop H, Nagel R, Liu YP, van Duijse J, Drost J, Griekspoor A, Zlotorynski E, Yabuta N, et al. A genetic screen implicates miRNA-372 and miRNA-373 as oncogenes in testicular germ cell tumors. Cell 2006; 124: 1169 1181 [DOI] [PubMed] [Google Scholar]
- Ro S, Song R, Park C, Zheng H, Sanders KM, Yan W. Cloning and expression profiling of small RNAs expressed in the mouse ovary. RNA 2007; 13: 2366 2380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ro S, Park C, Sanders KM, McCarrey JR, Yan W. Cloning and expression profiling of testis-expressed microRNAs. Dev Biol 2007; 311: 592 602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishima T, Takizawa T, Luo SS, Ishibashi O, Kawahigashi Y, Mizuguchi Y, Ishikawa T, Mori M, Kanda T, Goto T, Takizawa T. MicroRNA (miRNA) cloning analysis reveals sex differences in miRNA expression profiles between adult mouse testis and ovary. Reproduction 2008; 136: 811 822 [DOI] [PubMed] [Google Scholar]
- Yan N, Lu Y, Sun H, Tao D, Zhang S, Liu W, Ma Y. A microarray for microRNA profiling in mouse testis tissues. Reproduction 2007; 134: 73 79 [DOI] [PubMed] [Google Scholar]
- Fu X, Adamski M, Thompson EM. Altered miRNA repertoire in the simplified chordate, Oikopleura dioica. Mol Biol Evol 2008; 25: 1067 1080 [DOI] [PubMed] [Google Scholar]
- Clinton M, Haines L, Belloir B, McBride D. Sexing chick embryos: a rapid and simple protocol. Br Poult Sci 2001; 42: 134 138 [DOI] [PubMed] [Google Scholar]
- Darnell DK, Kaur S, Stanislaw S, Konieczka JH, Yatskievych TA, Antin PB. MicroRNA expression during chick embryo development. Dev Dyn 2006; 235: 3156 3165 [DOI] [PubMed] [Google Scholar]
- Landgraf P, Rusu M, Sheridan R, Sewer A, Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M, Lin C, Socci ND, et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007; 129: 1401 1414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang Y, Ridzon D, Wong L, Chen C. Characterization of microRNA expression profiles in normal human tissues. BMC Genomics 2007; 8: 166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murchison EP, Kheradpour P, Sachidanandam R, Smith C, Hodges E, Xuan Z, Kellis M, Grutzner F, Stark A, Hannon GJ. Conservation of small RNA pathways in platypus. Genome Res 2008; 18: 995 1004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagaraja AK, Andreu-Vieyra C, Franco HL, Ma L, Chen R, Han DY, Zhu H, Agno JE, Gunaratne PH, DeMayo FJ, Matzuk MM. Deletion of Dicer in somatic cells of the female reproductive tract causes sterility. Mol Endocrinol 2008; 22: 2336 2352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong X, Luense LJ, McGinnis LK, Nothnick WB, Christenson LK. Dicer1 is essential for female fertility and normal development of the female reproductive system. Endocrinology 2008; 149: 6207 6212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi K, Chuva de Sousa Lopes SM, Kaneda M, Tang F, Hajkova P, Lao K, O'Carroll D, Das PP, Tarakhovsky A, Miska EA, Surani MA. MicroRNA biogenesis is required for mouse primordial germ cell development and spermatogenesis. PLoS ONE 2008; 3: e1738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin RD, Aksay G, Dolgosheina E, Ebhardt HA, Magrini V, Mardis ER, Sahinalp SC, Unrau PJ. Comparative analysis of the small RNA transcriptomes of Pinus contorta and Oryza sativa. Genome Res 2008; 18: 571 584 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.