

Abstract
A native immunoisolation procedure has been used to investigate the role of clathrin-coated vesicles (CCVs) in the transport of vacuolar proteins between the trans-Golgi network (TGN) and the prevacuolar/endosome compartments in the yeast Saccharomyces cerevisiae. We find that Apl2p, one large subunit of the adaptor protein-1 complex, and Vps10p, the carboxypeptidase Y vacuolar protein receptor, are associated with clathrin molecules. Vps10p packaging in CCVs is reduced in pep12Δ and vps34Δ, two mutants that block Vps10p transport from the TGN to the endosome. However, Vps10p sorting is independent of Apl2p. Interestingly, a Vps10CtΔp mutant lacking its C-terminal cytoplasmic domain, the portion of the receptor responsible for carboxypeptidase Y sorting, is also coimmunoprecipitated with clathrin. Our results suggest that CCVs mediate Vps10p transport from the TGN to the endosome independent of direct interactions between Vps10p and clathrin coats. The Vps10p C-terminal domain appears to play a principal role in retrieval of Vps10p from the prevacuolar compartment rather than in sorting from the TGN.
INTRODUCTION
The sorting and delivery of vacuolar hydrolases in the yeast Saccharomyces cerevisiae is similar to the transport of lysosomal proteins in mammalian cells (Klionsky and Emr, 1990). Like all proteins entering the secretory pathway, hydrolytic enzymes destined to the vacuole are first translocated from the cytoplasm into the endoplasmic reticulum lumen, and then travel to the Golgi complex by vesicular transport. At the trans face of the Golgi complex, vacuolar hydrolases are sorted away from the secretory pathway to be transported to the vacuole via a prevacuolar/late endosome compartment.
In mammalian cells, the mannose 6-phosphate receptor (M6PR) mediates lysosomal enzyme transport from the trans-Golgi network (TGN) to the endosome (for review, see Hille-Rehfeld, 1995). Clathrin has been directly linked to the process of M6PR sorting at the TGN. Assembly protomers (triskelions), consisting of three heavy (CHC) and three light (CLC) chains, polymerize to form a polyhedral lattice creating a vesicle carrying lysosomal hydrolases with a mannose 6-phosphate residue (Schmid, 1997). The ligand–receptor complex is then transported in clathrin-coated vesicles (CCVs) to a prelysosomal/late endosome compartment. After ligand–receptor dissociation, lysosomal hydrolases are delivered to the lysosome, whereas the M6PR is recycled back to the TGN for another round of transport. Previous work showed that M6PR recruitment into CCVs at the TGN is directed by specific signals (Johnson and Kornfeld, 1992). A dileucine and a tyrosine motif are important for the efficient transport of M6PR from the TGN to the prelysosome. These sequences may represent the binding site for the Golgi-specific clathrin adaptor protein-1 (AP-1) complex (Glickman et al., 1989; Honing et al., 1997; Le Borgne and Hoflack, 1997; Klumperman et al., 1998). Tyrosine and dileucine motifs are also present in many cell surface proteins and interact with the μ2 subunit of the AP-2 complex to mediate internalization into endocytic CCVs (Kirchhausen et al., 1997; Marks et al., 1997; for review, see Bonifacino and Dell'Angelica, 1999; Heilker et al., 1999). The sequence YXXØ (where X is any amino acid and Ø is amino acid with a bulky hydrophobic group) represents the consensus signal for interaction with the μ2 subunit. Recently, a crystal structure revealed that the sorting signal is in an extended conformation when bound, and that the tyrosine and Ø residues reside in hydrophobic pockets (Owen and Evans, 1998).
Two Golgi-to-vacuole protein pathways have been identified in S. cerevisiae. One pathway was discovered by the isolation of mutants that missort carboxypeptidase Y (CPY), a soluble vacuolar protein (Bankaitis et al., 1986; Rothman and Stevens, 1986; Robinson et al., 1988; Rothman et al., 1989). This pathway traverses a prevacuolar/endosomal compartment before reaching the vacuole. An alternative pathway has been identified for alkaline phosphatase (ALP) that bypasses the endosomal compartment to reach the vacuole (Cowles et al., 1997a; Piper et al., 1997).
The CPY-sorting receptor was identified as the product of the VPS10 gene (Marcusson et al., 1994). Although CPY is sorted by recognition of a peptide rather than a carbohydrate signal, Vps10p function is otherwise similar to that of mammalian M6PR in providing a means of CPY delivery to the endosomal compartment. Recycling of Vps10p to the TGN is mediated by a tyrosine-based signal located in its cytoplasmic C-terminal tail. Mutation of this tyrosine leads to a rapid degradation of the receptor in the vacuole and to missorting of CPY (Cereghino et al., 1995; Cooper and Stevens, 1996).
Clathrin probably participates at some point in this sorting because a brief restrictive incubation of a temperature-sensitive mutant (chc1-521) results in rapid secretion of CPY (Seeger and Payne, 1992a). However, a backup process has been proposed to compensate the clathrin defect because a longer restrictive incubation of this mutant or the use of other clathrin mutants restores a normal rate of pro-CPY maturation. The role of the AP complexes is less clear. Three distinct AP complexes have been identified in yeast. Each complex contains two large chains (Apl), one medium chain (Apm) and one small chain (Aps). Null alleles of the different AP-1 subunit genes (APL2, APL4, APM4, and APS1) display genetic interactions with clathrin missense mutations, consistent with a role for the AP-1–related subunits in some clathrin-dependent pathway(s) (Phan et al., 1994; Rad et al., 1995; Stepp et al., 1995; Yeung et al., 1999). The two other complexes, AP-2R and AP-3, do not interact with clathrin (Yeung et al., 1999). AP-2R displays the highest protein sequence similarity to mammalian AP-2. Although the mammalian AP-2 associates with endocytic CCVs, yeast AP-2R mutants are not defective in endocytosis. Thus, the role of yeast AP-2R remains uncertain. AP3 is required for transport of ALP from the TGN to the vacuole and is clathrin-independent (Cowles et al., 1997b; Vowels and Payne, 1998). Surprisingly, disruptions of the four AP-1 subunits, or subunit disruptions that result in cells with no functional heterotetrameric AP complexes, do not affect clathrin formation and protein transport (Huang et al., 1999; Yeung et al., 1999). Thus, either clathrin alone is sufficient to execute roles in coat assembly and cargo selection or other proteins in CCVs are functionally redundant with the APs.
In this study we have developed a procedure to immunoisolate CCVs to determine the membrane proteins associated with clathrin molecules. We found that CCVs transport wild-type and sorting-deficient mutant Vps10p from the TGN to the endosome. This suggests that cargo proteins do not require a specific sorting signal in their cytoplasmic domain to be packaged into CCVs from the TGN.
MATERIALS AND METHODS
Strains, Growth Conditions, and Reagents
Yeast strains and plasmids used in this study are listed in Tables 1 and 2, respectively, and their construction is described below. Yeast cells were grown in synthetic or rich media (Sherman, 1991) at 30°C, or at 24°C as indicated. DNA manipulations in Escherichia coli were performed as described (Ausubel et al., 1987–1995). Yeast transformation was accomplished by using standard methods (Ausubel et al., 1987–1995).
Table 1.
Yeast strains used in this study
| Strain | Genotype | Source |
|---|---|---|
| EHY191 | MATa pep4::TRP1 TPI::SUC2::HIS ade2-1 his3-11, 15 leu2-3, 112 trpl-Δ1 ura3-1 | E. Harsay, unpublished |
| LSY93-2A | MATa leu2-3, 112 ura3-52 his3-Δ200 trpl-Δ901 suc2-Δ9 CLC1 | Silveira et al., 1990 |
| LSY93-5B | MATa leu2-3, 112 ura3-52 his3-Δ200 trpl-Δ901 suc2-Δ9 clc1::HIS3 | Silveira et al., 1990 |
| EHY202 | MATα chc1-521(ts) ura3-52 | E. Harsay, unpublished |
| GPY409 | MATa leu2-3, 112 ura3-52 his4 and/or his6 trpl-289 chc1-521(ts) sst1-3 pep::LEU2 | Rad et al., 1995 |
| PHY102 | MATa suc2-Δ9 ura3-52 leu2-3 his3-Δ200 trp1Δ911 lys 2-801 vps34Δ::TRP1 | Herman and Emr, 1990 |
| GPY1783-10A | MATα ura3-52 leu2-3 his3-200 trp1-901 lys2-801 suc1-9 GAL mel apl2::TRP1 | Yeung et al., 1999 |
| RSY1306 | MATα ura3-52 leu2,3-113 his3-Δ200 trp1-Δ901 lys2-801 pep12Δ::HIS3 | Becherer et al., 1996 |
| CCV120 | MATα vps45Δ2::HIS3 his3-Δ200 leu2-3, 112 lys2-801 trp1-Δ901 ura3-52 suc2-Δ9 | Burd et al., 1997 |
| EHY62 | MATα sec6-4 vps10Δ::TRP1 his3-Δ200 leu2-3, 112 lys2-801 trp1-1 ura3-52 | E. Harsay, unpublished |
| ODY117 | EHY202::KEX2HA::URA3 | This study |
| ODY47 | LSY93-2A::KEX2HA::URA3 | This study |
| ODY50 | LSY93-2A::PEP1HA::URA3 | This study |
| ODY129 | EHY202::VPS10HA::URA3 | This study |
| ODY56 | LSY93-5B::VPS10 HA::URA3 | This study |
| ODY62 | GPY1783-10A::VPS10 HA::URA3 | This study |
| ODY63 | RSY1306::VPS10 HA::URA3 | This study |
| ODY54 | PHY102::VPS10 HA::URA3 | This study |
| ODY80 | LSY93-2A/pRS426vps10G1423stop-HA | This study |
| ODY112 | CCY120/pRS426vps10G1423stop-HA | This study |
| ODY114 | RSY1306/pRS426vps10G1423stop-HA | This study |
| ODY140 | GPY1783-10A/pRS426vps10G1423stop-HA | This study |
| ODY141 | EHY62/pRS426vps10G1423stop-HA | This study |
| ODY142 | PHY102/pRS426vps10G1423stop-HA | This study |
| ODY143 | EHY191/pRS426vps10G1423stop-HA | This study |
| ODY144 | GPY409/pRS426vps10G1423stop-HA | This study |
Table 2.
Plasmids used in this study
| Plasmid | Description | Source |
|---|---|---|
| pGEX-2TGSTCLC1 | CLC1 (ORF) was fused to GST gene of pGEX-2T vector | This study |
| pBAD24CLC1HIS6 | CLC1 (ORF) containing 6 HIS at the C terminus was cloned into the pBAD24 vector | This study |
| pRS306KEX2HA | pRS306 containing KEX2 gene with three copies of the HA epitope inserted at the C terminus of the ORF | Nothwehr et al., 1995 |
| pRS306VPS10HA | pRS306 containing VPS10 gene with three copies of the HA epitope inserted at the C terminus of the ORF | This study |
| pRS426vps10HAG1423stop | pRS426 containing VPS10 gene with three copies of the HA epitope followed by two stop codons inserted at the codon 1423 of the ORF | This study |
| pGTEP1 | pBluescript II SK (+/−) containing three HA copies | RS strain collection |
Enzymes for the manipulation of DNA were purchased from New England Biolabs (Beverly, MA) or Boehringer-Mannheim Biochemicals (Indianapolis, IN). A plasmid containing KEX2HA (Nothwehr et al., 1995) was a gift from S.T. Nothwehr (University of Missouri, Columbia, MO). Monoclonal Chc1p antibody has been published (Lemmon et al., 1988) and antisera against Apl2p, Apl1p, Vps10p, and CPY have been described (Feldheim et al., 1993; Cooper and Stevens, 1996; Yeung et al., 1999). Mouse12CA5 anti-hemagglutinin (HA) antibodies were purchased from Berkeley Antibody (Richmond, CA). Donkey anti-rabbit and sheep anti-mouse secondary antibodies coupled to horseradish peroxidase were obtained from Amersham (Arlington Heights, IL). Zymolyase 100T was obtained from United States Biological (Swampscott, MA). [35S]Promix was purchased from Amersham. Other chemicals were purchased from Sigma Chemical Co. (St. Louis, MO), unless indicated.
Strains and Plasmid Construction
The pGEX-2TCLC1GST plasmid was constructed by using one oligonucleotide primer containing a BamHI site and a second primer containing an EcoRI site corresponding to the upstream and the downstream region of the CLC1 open reading frame (ORF), respectively. The CLC1 gene was amplified by PCR and fused into the BamHI and EcoRI sites of pGEX-2T containing a gluthathione S-transferase (GST) gene. The CLC1 (ORF/697 bp) PCR product containing a C-terminal 6 HIS encoding sequence was cloned into EcoRI and XbaI sites of the pBAD24 vector (Guzman et al., 1995), yielding the pBAD24CLC1HIS6 plasmid. The influenza HA epitope-tagged VPS10 allele was constructed by first inserting a NotI site by PCR just upstream of the VPS10 ORF stop codon in the pRS315VPS10 plasmid (Piper et al., 1995). A NotI fragment containing three copies of the HA epitope (YPYDVPDYA) digested from pGTEP1 plasmid was then ligated into the VPS10 gene containing a NotI site, resulting in plasmid pRS315VPS10HA. The VPS10HA fragment from pRS315VPS10HA was subcloned into the SacI and SalI sites of the pRS306 vector. pRS426vps10G1423stop-HA was constructed as described for pRS306VPS10HA by inserting three copies of the HA epitope followed by two stop codons just upstream of the glycine 1423 residue of the open reading frame of the VPS10 gene.
We constructed strains containing a single copy of KEX2-HA or VPS10-HA. For the integration of KEX2-HA, we digested pRS306KEX2-HA with BglII (which cuts in KEX2) and inserted it into EHY202 and LSY93-2A by transformation, resulting in ODY117 and ODY47, respectively. For the integration of VPS10-HA, we digested pRS306VPS10-HA with AflII (which cuts in VPS10) and introduced it into LSY93-2A, EHY202, LSY93-5B, GPY1783-10A, RSY1306, and PHY102 by transformation, resulting in ODY50, ODY129, ODY55, ODY62, ODY63, and ODY54, respectively.
Antibody Generation
The pGEX-2TGST, pGEX-2TGSTCLC1, and pBAD24CLC1HIS6 plasmids were propagated by transformation into BL21 E. coli cells. Expression of the Clc1p-HIS6 protein was induced with 0.1% arabinose and purified by using a nickel-nitrilotetraacetic acid agarose column. Expression of the GST and Clc1p-GST proteins was induced by isopropyl β-d-thiogalactopyranoside and then purified by using gluthathione-agarose chromatography. The purified GST and Clc1p-GST fusion proteins were injected into rabbits (Northview Pacific Laboratories, Berkeley, CA). GST and Clc1p rabbit antibodies were affinity-purified on GST and Clc1p proteins covalently attached to ReactiGel (6X) carbonydiimidazole-activated agarose (Pierce Chemical, Rockford, IL), respectively, as described (Chuang and Schekman, 1996).
Native Immunoisolation
Cells were grown in YPD at 24°C to an OD600 < 1. Forty OD600 units of cells were sedimented by centrifugation and washed once with 20 mM cold NaN3. Cells were resuspended and incubated in 0.8 ml (50 OD600/ml) of 0.1 M Tris-HCl, pH 9.4, 10 mM dithiothreitol (DTT), 20 mM NaN3 for 10 min at room temperature. Cells were then converted to spheroplasts by treatment with Zymolyaze T100 (7.5 μg/OD) in 0.8 ml of spheroplast buffer (10 mM Tris-HCl, pH 7.5, 0.8 M sorbitol, 20 mM NaN3, 1 mM DTT) for 15 min at 30°C. Spheroplasts were sedimented by centrifugation for 2 min at 3000 × g and washed once with spheroplast buffer and once with 2-(N-morpholino)ethanesulfonic acid (MES)/0.8 M sorbitol buffer (100 mM MES pH 6.5, 0.8 M sorbitol, 0.5 mM MgCl2, 1 mM EGTA, 0.2 mM DTT) at 4°C. Spheroplasts were then hypotonically lysed in 0.4 ml (100 OD600/ml) of MES/0.2 M sorbitol buffer (100 mM MES, pH 6.5, 0.2 M sorbitol, 0.5 mM MgCl2, 1 mM EGTA, 0.2 mM DTT, 1 mM phenylmethylsulfonyl fluoride, 1 mM benzamidine, pepstatin A [1 μg/ml], and leupeptin [1 μg/ml]) at 4°C and homogenized in a glass Dounce homogenizer (1 ml) by using a tight-fitting pestle. The yeast lysate was centrifuged for 30 min, 4°C at 21,000 × g. The pellet was discarded and the supernatant centrifuged for 60 min at 4°C at 100,000 × g in a TLS 55 rotor (Beckman Instruments, Palo Alto, CA) onto a 200-μl cushion of 80% Percoll. The supernatant was discarded and the material layered on the Percoll pellet was resuspended in 500 μl of MES/0.2 M sorbitol buffer and divided into two Eppendorf tubes. Both preparations were diluted to 1 ml with MES/0.2 M sorbitol buffer containing 1% bovine serum albumin (BSA) and incubated with 1.5 μg of affinity-purified Clc1p or GST antibodies, respectively, for 2 h or overnight at 4°C. The immunoabsorbed complexes were isolated by addition of 30 μl of protein A-Sepharose (20% in MES/0.2 M sorbitol buffer containing 1% BSA) and incubated for an additional hour at 4°C. Nonabsorbed materials were removed by centrifuging and the immunoabsorbate-bound protein A-Sepharose was resuspended and washed once in 1 ml of MES/0.2 M sorbitol buffer containing 0.1% BSA and three times in 1 ml of MES/0.2 M sorbitol buffer without BSA. The immunoabsorbates were eluted in 20 μl of Laemmli buffer (Laemmli, 1970) and then heated at 95°C for 5 min. Protein A-Sepharose was removed by centrifugation and the supernatant fractions were resolved on 8% SDS-PAGE gels. Proteins were visualized by immunoblot by using the ECL+ kit (Amersham Pharmacia Biotech, Piscataway, NJ). Protein quantitation was performed by using a PhosphorImager (Molecular Dynamics, Sunnyvale, CA) according to manufacturer's recommendations.
Radiolabeling, Immunoprecipitation, and Immunoblot Analysis
Vps10p, CPY, and HA-tagged proteins were immunoprecipitated under denaturing conditions from radiolabeled extracts by using a procedure described previously (Nothwehr et al., 1995; Bryant et al., 1998), with the appropriate polyclonal or monoclonal antibodies.
Enzyme Assays
GDPase assays were performed essentially as described (Abeijon et al., 1989; Yanagisawa et al., 1990). A portion (0.00125% of total) of the high-speed membrane pellet (HSP) fraction and all the anti-GST and anti-Clc1p immunoabsorbates were added to 100 μl of reaction buffer (20 mM imidazole-HCl, pH 7.4, 2 mM CaCl2, 0.1% Triton X-100, and 9 mM GDP). Reactions were incubated at 30°C for 15 min and stopped by adding 150 μl of 2% SDS. GDPase activity was measured based on absorbance at 820 nm.
Invertase activity was assayed by the method of Goldstein and Lampen (1975). An aliquot (0.05% of total) of the HSP fraction and all of the anti-GST and anti-Clc1p immunoabsorbates were added to the enzyme reaction buffer containing 0.1% Triton X-100. Invertase activity was measured based on absorbance at 540 nm.
Blanks were measured in reactions conducted without membrane fractions. Error bars were calculated from two independent experiments
RESULTS
Clathrin Associates with Apl2p in a Membrane Fraction
To isolate yeast CCVs, we developed a procedure with anti-clathrin antibodies for the affinity separation of CCVs from a cell extract. Previously, affinity-purified anti-Clc1p antibody was successfully used to immunoprecipitate the intact clathrin trimeric complex (Pishvaee et al., 1997). We reasoned that a similar approach would allow us to immunoisolate intact coated-vesicles from a crude, slowly sedimenting membrane fraction. Whole-cell extracts were prepared by gentle osmotic lysis followed by a differential centrifugation to obtain a HSP fraction. A native immunoprecipitation was then performed by using the HSP fraction combined with affinity-purified antibodies against Clc1p followed by protein A-Sepharose. The resulting immunoprecipitate was analyzed by SDS-PAGE and immunoblotted. Because the AP-1 complex has been shown to interact with clathrin, we first evaluated coimmunoprecipitation of Apl2p, the large subunit of AP-1. We found that 60% of Chc1p and 6% of Apl2p of the total HSP fraction were coprecipitated with Clc1p (Figure 1 and Table 3). As a control, no coprecipitation (<0.5%) was observed when antibodies against GST were used. In contrast, Apl1p, the large subunit of AP-2 was not detected. This result is in good agreement with previous work showing that of the three AP complexes, only AP-1 interacts with clathrin (Yeung et al., 1999). Clathrin and AP are soluble proteins that are recruited on membrane for the formation of vesicles. Thus, we tested whether Alp2p could be coprecipitated with clathrin from the high-speed soluble (HSS) fraction. As expected, Chc1p was coprecipitated with Clc1p because clathrin is present as soluble triskelions in the cytosol (Pishvaee et al., 1997). In contrast, Apl2p was not detected, suggesting that interaction between clathrin and Apl2p requires additional components from the membrane fraction. It is possible that lipids or/and membrane proteins induce a conformational change in AP-1 to increase its affinity for clathrin.
Figure 1.

Clathrin interacts with Apl2p. Native immunoprecipitations were performed from a lysate of strain LSY93-2A by using affinity-purified Clc1p and GST antibodies. Lanes 1 and 4 correspond to 1/20 of the HSP and HSS fractions used, respectively, for the immunoprecipitations. Lanes 2 and 3 correspond to the immunoprecipitates of an HSP fraction by using GST and Clc1p antibodies, respectively. Lanes 5 and 6 correspond to the immunoprecipitates from an HSS fraction by using GST and Clc1p antibodies, respectively. The HSP/HSS fractions and the immunoprecipitates were subjected to SDS-PAGE and transferred to a nitrocellulose filter. The nitrocellulose was cut into strips and immunoblotted with Clc1p, Chc1p, Apl2p, and Apl1p antibodies.
Table 3.
Protein enrichments by immunoprecipitations
| Protein enrichments (%)
|
||||
|---|---|---|---|---|
| WT
(%)
|
chc1 (%)
|
|||
| αGST | αClc1p | αGST | αClc1p | |
| Clc1p | <0.5 | >60 | ND | ND |
| Chc1p | <0.5 | 60 | ND | ND |
| Apl1p | <0.5 | <0.5 | <0.5 | <0.5 |
| Apl2p | <0.5 | 6 | <0.5 | <0.5 |
| Vps10p-HA | <0.5 | 6 | <0.5 | 1.5 |
Each value represents protein immunoprecipitation with αGST and αClc1p antibodies, respectively, in percentage of the total HSP fraction with a standard error of ∼20%. Quantification was performed from three independent immunoprecipitation experiments and determined by using Phosphorimager (Molecular Dynamics).
ND, not determined.
Vps10p Is Coimmunoprecipitated with Clathrin
To determine whether our immunoisolation provides intact CCVs, we explored the enrichment of specific cargo proteins associated with clathrin. First, we evaluated Kex2p because this Golgi membrane protein, which is required for the maturation of α-factor precursor, is mislocalized to the cell surface in clathrin mutant strains and coelutes with clathrin in CCVs resolved by gel filtration (Payne and Schekman, 1989; Chu et al., 1996). We performed a similar native immunoprecipitation experiment as described in Figure 1. To improve the detection of Kex2p, we used a construct containing a functional version of Kex2p tagged with the influenza HA epitope integrated at the chromosomal KEX2 locus. We found that Kex2p-HA was coimmunoprecipitated with Clc1p, Chc1p, and Apl2p (Figure 2A).
Figure 2.

Detection of Vps10p-HA by coimmunoprecipitation with clathrin antibodies. Native immunoprecipitations were performed as in Figure 1. (A) Lanes 1–3 correspond to native immunoprecipitation from WT/ODY50. Lanes 4–6 correspond to native immunoprecipitation from chc1ts/ODY129. Native immunoprecipitations for Kex2p-HA enrichment were performed from WT/ODY47 (lane 1–3) and from chc1ts/ODY117 (lane 4–6). Kex2p-HA and Vps10p-HA were detected by using monoclonal 12CA5 anti-HA antibody. GDPase (B) and invertase (C) activities were performed on HSP and immunoprecipitation fractions from WT/EHY191.
In the next experiment, we examined the role of clathrin in the sorting of vacuolar proteins. Prior work showed that a rapid inactivation of clathrin results in a severe defect in the sorting of the vacuolar soluble protein CPY (Seeger and Payne, 1992a). The precursor form of CPY (p2CPY) interacts with Vps10p in the Golgi complex and is believed to be transported to the endosome by CCVs (see INTRODUCTION). We examined the association of Vps10p with clathrin and found that 6% of a functional HA-tagged Vps10p was coimmunoprecipitated (Figure 2A and Table 3). To further demonstrate the specificity of cargo detection, we used a chc1-521 mutant in which the stability of the clathrin trimeric complex is decreased (Pishvaee et al., 1997). We found that clathrin remains cytosolic (no Clc1p in HSP; Figure 2A, lane 4) in a chc1-521 cell lysate prepared from cells grown at a permissive temperature, conditions where Kex2p and Vps10p trafficking is not affected. In the absence of clathrin in the HSP fraction, Apl2p (<0.5%) was not detected and very little contamination of Kex2p-HA and Vps10p-HA (<1.5%) was observed (Figure 2A and Table 3), confirming a specific protein enrichment.
These results do not exclude the possibility that Kex2-HA and Vps10p-HA are part of clathrin-coated TGN membranes in our native immunoprecipitation. Therefore, we examined the precipitation of proteins that are excluded from CCVs. We used GDPase and invertase as Golgi and secreted protein markers, respectively. Figure 2, B and C, show that no GDPase and invertase activities above background were coimmunoprecipitated with clathrin antibodies, whereas a significant level of both activities was detected in the HSP fraction. Together these results provide reasonable evidence that transport of Kex2p and Vps10p is mediated by CCVs.
Vps10p Is Degraded in the chc1 Mutant
Previous studies showed that Kex2p is rapidly degraded in a chc1 mutant (Redding et al., 1996). We examined the stability of Vps10p in a chc1 mutant by a pulse-chase experiment. Vps10p is an extremely stable protein and no degradation was observed after a chase of 90 min at 37°C in wild-type (WT) cells (Cereghino et al., 1995; Cooper and Stevens, 1996) (Figure 3). However, in the chc1 mutant, Vps10p was slowly degraded (Figure 3). Degradation was blocked in a protease-deficient pep4 mutant strain (Figure 3A, lane 9). A similar result was obtained with a clc1Δ strain (Figure 4A). Clearly, Kex2p and Vps10p are degraded at distinct rates in the clathrin mutant, possibly reflecting the fact that Vps10p continues to sort CPY by a clathrin-independent process.
Figure 3.

Vps10p is degraded in chc1ts cells. WT/LSY93–2A and chc1ts/EHY202 strains were radiolabeled for 10 min with [35S]methionine and cysteine directly at 24°C (lanes 1–4) or were incubated for 45 min (lanes 5–9) at 37°C before labeling at 37°C. After the indicated chase times, aliquots of cells were removed and subjected to immunoprecipitation with antibodies to Vps10p. Lane 9 corresponds to Vps10p immunoprecipitation after a 90-min chase at 37°C in a pep4 background (WT/EHY191 and chc1ts/GPY409). The PEP4-dependent cleavage product is indicated (∗). Samples were analyzed by SDS-PAGE and autoradiography.
Figure 4.

Vps10p stability and CPY maturation in WT, apl2Δ, clc1Δ, pep12Δ, and vps34Δ strains. WT/LSY93-2A, apl2Δ/GPY1783-10A, clc1Δ/LSY93-5B, pep12Δ/RSY1306, and vps34Δ/ PHY102 strains were radiolabeled for 10 min with [35S]methionine and cysteine directly at 30°C. After a chase for the indicated times (0, 1, and 2 h), aliquots of cells were removed and subjected to immunoprecipitation with antibodies to Vps10p (A) or CPY (B). The PEP4-dependent cleavage product is indicated (∗).
Coimmunoprecipitation of Vps10p Is Reduced in pep12Δ, and vps34Δ, but not in apl2Δ Mutants
To further characterize Vps10p transport by CCVs, we examined mutants that affect clathrin or protein transport from the TGN to the endosome. However, to be able to assess coimmunoprecipitation of Vps10p with clathrin, we restricted our analysis to mutants in which Vps10p was not degraded (t1/2 < 1 h) and was retained in an HSP sedimentable fraction as in wild-type cells. Apl2p interacts with clathrin, but disruption of APL2 displays no discernable phenotypes (Rad et al., 1995). In agreement with these results, a pulse-chase analysis showed that Vps10p stability and CPY maturation were not affected in the apl2Δ mutant (Figure 4). Pep12p is a t-SNARE component required for the fusion of vesicles with the endosome (Becherer et al., 1996; Burd et al., 1997). Vps10p was stable in pep12Δ cells (Figure 4A), most likely because it accumulated in proteolytically inactive vesicles and was protected from degradation, as proposed for other mutants (vps45 and vps21) that also block vesicle fusion with endosome (Bryant et al., 1998; Gerrard et al., 2000). As a result, >90% of CPY is then secreted because Vps10p cannot be recycled back to the TGN for another round of transport of CPY (Figure 4B). A similar result was obtained with vps34Δ. Vps34p is a phosphoinositide 3-kinase required for protein trafficking (Schu et al., 1993; Stack and Emr, 1994; Stack et al., 1995). Mutations in VPS34 block protein transport from the TGN to the endosome. As a consequence, Vps10p fails to return to the TGN, resulting in secretion of CPY (Figure 4B; Herman and Emr, 1990). We found that Vps10p was degraded to some extent (Figure 4A), but still accumulated in an HSP fraction.
We next performed immunoprecipitations as described in Figure 1 from WT, apl2Δ, pep12Δ, and vps34Δ cells. Interestingly, in apl2Δ, no decrease of clathrin (Chc1p and Clc1p) in the HSP was observed (Figure 5), indicating that the AP-1 complex is not essential for the recruitment of clathrin onto the membrane. Furthermore, Vps10p-HA was detected with WT efficiency. Thus, Apl2p is totally dispensable for Vps10p transport. In contrast, in pep12Δ and vps34Δ cells in which CPY sorting from the TGN is blocked, Vps10p-HA enrichment was clearly reduced (Figure 5). This result can be explained by the formation of new CCVs that are depleted of Vps10p because of deficient receptor recycling to the TGN.
Figure 5.

Coimmunoprecipitation of Vps10p-HA with clathrin is reduced in pep12Δ and vps34Δ but not in apl2Δ/mutants. Native immunoprecipitations were performed from ODY50 (WT/lane 1–3), ODY62 (apl2Δ/lane 4–6), ODY63 (pep12Δ/lane 7–9), and ODY54 (vps34Δ/lane 10–12) as described in Figure 1.
Vps10p Does Not Interact with Clathrin via its C-Terminal Sorting Domain
Early studies showed that Vps10p contains a retrieval signal in the C-terminal cytoplasmic domain that is essential for its recycling from the endosome to the TGN (Cereghino et al., 1995; Cooper and Stevens, 1996). A Vps10p mutant lacking the C-terminal cytoplasmic domain (Vps10CtΔp) is rapidly degraded in the vacuole with a half-time of ∼25 min in WT cells. The degradation of Vps10CtΔp is blocked in vps45Δ, an observation that has been explained by entrapment of Vps10CtΔp inside proteolytically inactive vesicles that are unable to fuse with a post-Golgi/endosome compartment (Bryant et al., 1998). We examined the fate of Vps10CtΔp in pep12Δ and vps34Δ, by using a construct of Vps10p containing an HA epitope in place of its C-terminal cytoplasmic tail (Vps10CtΔp-HA). From a pulse-chase analysis, we found that Vps10CtΔp-HA was processed with a t1/2 < 30 min in WT cells; proteolysis was blocked in pep12Δ, vps45Δ, and vps34Δ mutants (Figure 6). This result supports a model in which both Vps10p-HA and Vps10CtΔp-HA are transported in the same Golgi-derived vesicles that require Pep12p, Vps45p, and Vps34p to reach the endosome. We next examined whether Vps10CtΔp-HA was contained in CCVs. To compensate for the lack of recycling of Vps10CtΔp-HA, we overproduced the protein by using a 2-μ plasmid (pRS426 vector). A degradation product of Vps10CtΔp-HA was detected in the HSP (Figure 7A). However, only the full-length Vps10CtΔp-HA was coimmunoprecipitated with clathrin. A similar result was obtained by using pep4 cells in which Vps10CtΔp-HA is stable (WT, Figure 7B). Coprecipitation was dependent on functional clathrin because little Vps10CtΔp-HA was detected in Clc1p precipitates from pep4 cells lacking clathrin heavy chain (chc1, Figure 7B). Vps10CtΔp-HA association with clathrin was not affected in the apl2Δ mutant. Detection of plasmid-encoded Vps10CtΔp-HA in CCVs isolated from cells missing the chromosomal VPS10 locus (vps10, Figure 7A) rules out the possibility that Vps10CtΔp-HA is recruited into CCVs by interaction with endogenous full-length Vps10p. Together, these data indicate that CCVs mediate anterograde transport of proteins from the TGN to the endosome and that the cytoplasmic C-terminal tail of Vps10p is not essential for its recruitment into CCVs.
Figure 6.
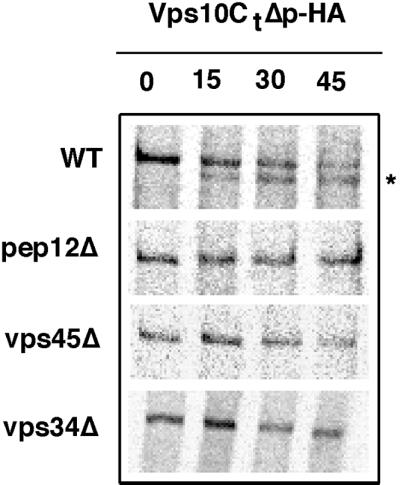
pep12Δ, vps45Δ, and vps34Δ mutants block Vps10CtΔp-HA degradation. WT/ODY80, pep12Δ/ODY114, vps45Δ/ODY112, and vps34Δ/ODY142 strains containing plasmid encoding Vps10CtΔp-HA were subjected to pulse-chase immunoprecipitation and analyzed as described in Figure 4. Except that monoclonal 12CA5 anti-HA antibody was used to immunoprecipitate Vps10CtΔp-HA from the cell extracts. The PEP4-dependent cleavage product is indicated (∗).
Figure 7.

Detection of Vps10CtΔp-HA by coimmunoprecipitation with clathrin antibodies. Native immunoprecipitations were performed as in Figure 1. (A) pRS426vps10G1423stop-HA plasmid was introduced into WT (ODY80), apl2Δ (ODY140), and vps10Δ (ODY141) strains. (B) pRS426vps10G1423stop-HA was introduced into WT (ODY143) and chc1ts (ODY144) strains. Vps10CtΔp-HA was detected by using monoclonal 12CA5 anti-HA antibody.
DISCUSSION
Clathrin-coated vesicles are required for sorting and retention of proteins in a late-Golgi compartment in yeast. In clathrin mutants, Golgi membrane proteins such as the dipeptidyl-aminopeptidase A (DPAP A) and Kex2p, two endoproteases required for the maturation of α-factor precursor, are mislocalized to the cell surface (Seeger and Payne, 1992b). These results suggested that Golgi proteins may accompany anterograde cargo proteins in CCVs leaving a late Golgi membrane, and then be efficiently recycled from the endosome by virtue of a retrieval mechanism. Thus, inactivation of clathrin would block Golgi-membrane proteins from reaching the endosome and thereby prevent their retrieval to the TGN. The sorting receptor Vps10p, which also contains a retrieval sorting signal, cycles between the TGN and the endosome in a manner similar to Kex2p and DPAP A to mediate the anterograde transport of CPY (Conibear and Stevens, 1998). Our study provides direct evidence that CCVs transport both Kex2p and Vps10p. Furthermore, Vps10p sorting into CCVs is independent of sorting signals in the cytoplasmic domain.
The observation that Vps10p is associated with clathrin by coimmunoprecipitation correlates with previous results showing that a rapid inactivation of temperature sensitive clathrin results in mislocalization of CPY (Seeger and Payne, 1992a). Both Kex2p and Vps10p contain a tyrosine-sorting motif in their C-terminal domains, which allows them to be recycled back to the TGN. Substitution of this motif or deletion of the entire domain leads to rapid degradation of these proteins in the vacuole. Kex2p and Vps10p C-terminal mutant proteins have been shown to transit through the endosome to reach the vacuole for degradation (Brickner and Fuller, 1997; Bryant and Stevens, 1997). In this regard, vps45Δ, pep12Δ, and vps34Δ mutants that affect the anterograde transport of Vps10p to the endosome also block the transport of Vps10CtΔp. This suggests that both Vps10p and Vps10CtΔp are transported by the same population of vesicles. Because we have been able to coimmunoprecipitate Vps10CtΔp-HA with clathrin, we propose that CCVs represent this population of vesicle and therefore, mediate the anterograde transport of proteins from the TGN to the endosome. Support for this view comes from the observation that clathrin genetically interacts with vps1, a dynamin that is believed to be required for the formation of vesicles at the TGN, and pep12, a t-SNARE involved in the fusion of post-Golgi vesicles with the endosome (Bensen et al., 2000). Consistently, we failed to coimmunoprecipitate Vps10p with clathrin antibodies in these two mutants (Figure 5; our unpublished results). Although we cannot exclude the possibility that other types of vesicle mediate transport from the TGN to the endosome, the observation that Golgi membrane proteins such as Kex2p and DPAP A are rerouted to the cell surface in clathrin mutants affirms this view (Seeger and Payne, 1992b; Redding et al., 1996). In this regard, we have observed that both Vps10p and Vps10CtΔp are rerouted to the cell surface in a chc1 mutant (unpublished data).
The cytoplasmic domain of membrane cargo proteins may serve directly to facilitate the recruitment of coat proteins. Previous studies showed that a specific recognition of tyrosine or dileucine motifs by APs is essential for the packaging of cargo proteins into CCVs in mammalian cells (Kirchhausen et al., 1997; Marks et al., 1997; for review, see Bonifacino and Dell'Angelica, 1999; Heilker et al., 1999). Cargo recruitment into yeast CCVs does not depend solely on any of the three identified AP complexes (Huang et al., 1999; Yeung et al., 1999). Mutants containing multiple deletions of APs did not display any phenotypes associated with clathrin deficiency, including slowed growth and defects in clathrin-dependent protein sorting in the endocytic or biosynthetic pathways. These results are extended by our data showing that coimmunoprecipitation of Vps10p and Kex2p (our unpublished results) with clathrin is not decreased in the absence of Apl2p. Thus, an alternative mechanism in which distinct clathrin-binding proteins or clathrin itself can substitute for AP function is possible. Indeed, accessory proteins provide alternative strategies for cargo selection (for review, see Jarousse and Kelly, 2000).
The efficient incorporation of Vps10CtΔp-HA in CCVs rules out the possibility that the cytoplasmic tail of Vps10p has a prominent role in clathrin coat formation and for its recruitment in CCVs. Other domains of Vps10p may interact indirectly with clathrin-associated proteins. However the principle sorting step determining the fate of TGN proteins may be in their selective retrieval from the endosome. Kex2p, which is transported efficiently from the TGN in CCVs, must be actively recycled from the endosome to maintain its steady-state location in the Golgi (Redding et al. 1996; Brickner and Fuller, 1997). The anterograde limb of this cycling pathway may be less cargo-selective than the retrieval event. For example, overexpression of Golgi membrane proteins does not saturate transport from the TGN to the endosome, whereas the retrieval pathway that is signal-directed is saturated (Roberts et al., 1992; Wilcox et al., 1992). In addition, Vam3p and ALP, two proteins that require a specific sorting signal to transit through the AP-3–dependent pathway to the vacuole, are rerouted through the endosome in AP-3–deficient strains (Cowles et al. 1997a; Stepp et al., 1997; Darsow et al., 1998). Thus, clathrin may provide a default pathway for the transport of certain mutant or heterologous proteins to the vacuole in S. cerevisiae.
How membrane proteins are recruited into vesicles without reference to a cytoplasmic signal remains to be elucidated. Perhaps the membrane anchor domains and a special phospholipid environment are essential to form CCVs from the yeast trans-Golgi organelle. Recently, Chen et al. (1999) have reported that the integral membrane P-type ATPase (DRS2), a potential aminophospholipid translocase, is required for clathrin function at the late Golgi. Drsp2 may create a special bilayer environment into which protein substrates for clathrin vesicles may partition.
ACKNOWLEDGMENTS
We thank Tom H. Stevens for generously providing polyclonal Vps10p antibodies, and Edina Harsay for providing unpublished strains and for expert advice. We thank members of Schekman lab for helpful discussion. O.D. was supported by a fellowship from the Swiss National Science Foundation. The work was supported by grants from the National Institutes of Health to R.S. and G.P., and by the Howard Hughes Medical Institute to R.S.
Abbreviations used:
- BSA
bovine serum albumin
- CCV
clathrin-coated vesicle
- CPY
carboxylpeptidase Y
- DTT
dithiothreitol
- MES
2-(N-morpholino)ethanesulfonic acid
- Tris
tris(hydroxymethyl)aminomethane
REFERENCES
- Abeijon C, Orlean P, Robbins PW, Hirschberg CB. Topography of glycosylation in yeast: characterization of GDPmannose transport and lumenal guanosine diphosphatase activities in Golgi-like vesicles. Proc Natl Acad Sci USA. 1989;86:6935–6939. doi: 10.1073/pnas.86.18.6935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausubel, F.M., Brent, R., Kingston, R.E., More, D.D., Seidman, J.G., Smith, J.A., and Struhl, K. (1987–1995). Current Protocols in Molecular Biology, New York: John Wiley & Sons.
- Bankaitis VA, Johnson LM, Emr SD. Isolation of yeast mutants defective in protein targeting to the vacuole. Proc Natl Acad Sci USA. 1986;83:9075–9079. doi: 10.1073/pnas.83.23.9075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becherer KA, Rieder SE, Emr SD, Jones EW. Novel syntaxin homologue, Pep12p, required for the sorting of lumenal hydrolases to the lysosome-like vacuole in yeast. Mol Biol Cell. 1996;7:579–594. doi: 10.1091/mbc.7.4.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bensen ES, Costaguta G, Payne GS. Synthetic genetic interactions with temperature-sensitive clathrin in Saccharomyces cerevisiae. Roles for synaptojanin-like Inp53p and dynamin-related Vps1p in clathrin-dependent protein sorting at the trans-Golgi network. Genetics. 2000;154:83–97. doi: 10.1093/genetics/154.1.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonifacino JS, Dell'Angelica EC. Molecular bases for the recognition of tyrosine-based sorting signals. J Cell Biol. 1999;145:923–926. doi: 10.1083/jcb.145.5.923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brickner JH, Fuller RS. SOI1 encodes a novel, conserved protein that promotes TGN-endosomal cycling of Kex2p and other membrane proteins by modulating the function of two TGN localization signals. J Cell Biol. 1997;139:23–36. doi: 10.1083/jcb.139.1.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryant NJ, Piper RC, Gerrard SR, Stevens TH. Traffic into the prevacuolar/endosomal compartment of Saccharomyces cerevisiae: a VPS45-dependent intracellular route and a VPS45-independent, endocytic route. Eur J Cell Biol. 1998;76:43–52. doi: 10.1016/S0171-9335(98)80016-2. [DOI] [PubMed] [Google Scholar]
- Bryant NJ, Stevens TH. Two separate signals act independently to localize a yeast late Golgi membrane protein through a combination of retrieval and retention. J Cell Biol. 1997;136:287–297. doi: 10.1083/jcb.136.2.287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burd CG, Peterson M, Cowles CR, Emr SD. A novel Sec18p/NSF-dependent complex required for Golgi-to-endosome transport in yeast. Mol Biol Cell. 1997;8:1089–1104. doi: 10.1091/mbc.8.6.1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cereghino JL, Marcusson EG, Emr SD. The cytoplasmic tail domain of the vacuolar protein sorting receptor Vps10p and a subset of VPS gene products regulate receptor stability, function, and localization. Mol Biol Cell. 1995;6:1089–1102. doi: 10.1091/mbc.6.9.1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen CY, Ingram MF, Rosal PH, Graham TR. Role for Drs2p, a P-type ATPase and potential aminophospholipid translocase, in yeast late Golgi function. J Cell Biol. 1999;147:1223–1236. doi: 10.1083/jcb.147.6.1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu DS, Pishvaee B, Payne GS. The light chain subunit is required for clathrin function in Saccharomyces cerevisiae. J Biol Chem. 1996;271:33123–33130. doi: 10.1074/jbc.271.51.33123. [DOI] [PubMed] [Google Scholar]
- Chuang JS, Schekman RW. Differential trafficking and timed localization of two chitin synthase proteins, Chs2p and Chs3p [published erratum appears in J. Cell Biol. (1996) Dec, 135(6 Pt 2):1925] J Cell Biol. 1996;135:597–610. doi: 10.1083/jcb.135.3.597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conibear E, Stevens TH. Multiple sorting pathways between the late Golgi and the vacuole in yeast. Biochim Biophys Acta. 1998;1404:211–230. doi: 10.1016/s0167-4889(98)00058-5. [DOI] [PubMed] [Google Scholar]
- Cooper AA, Stevens TH. Vps10p cycles between the late-Golgi and prevacuolar compartments in its function as the sorting receptor for multiple yeast vacuolar hydrolases. J Cell Biol. 1996;133:529–541. doi: 10.1083/jcb.133.3.529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowles R, Odorizzi G, Payne G, Emr S. The AP-3 adaptor complex is essential for cargo-selective transport to the yeast vacuole. Cell. 1997b;91:109–118. doi: 10.1016/s0092-8674(01)80013-1. [DOI] [PubMed] [Google Scholar]
- Cowles C, Snyder W, Burd C, Emr S. Novel Golgi to vacuole delivery pathway in yeast: identification of a sorting determinant and required transport component. EMBO J. 1997a;16:2769–2782. doi: 10.1093/emboj/16.10.2769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darsow T, Burd CG, Emr SD. Acidic di-leucine motif essential for AP-3-dependent sorting and restriction of the functional specificity of the Vam3p vacuolar t-SNARE. J Cell Biol. 1998;142:913–922. doi: 10.1083/jcb.142.4.913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldheim D, Yoshimura K, Admon A, Schekman R. Structural and functional characterization of Sec66p, a new subunit of the polypeptide translocation apparatus in the yeast endoplasmic reticulum. Mol Biol Cell. 1993;4:931–939. doi: 10.1091/mbc.4.9.931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerrard SR, Bryant NJ, Stevens TH. VPS21 controls entry of endocytosed and biosynthetic proteins into the yeast prevacuolar compartment. Mol Biol Cell. 2000;11:613–626. doi: 10.1091/mbc.11.2.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glickman JN, Conibear E, Pearse BM. Specificity of binding of clathrin adaptors to signals on the mannose-6-phosphate/insulin-like growth factor II receptor. EMBO J. 1989;8:1041–1047. doi: 10.1002/j.1460-2075.1989.tb03471.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein A, Lampen JO. Beta-D-fructofuranoside fructohydrolase from yeast. Methods Enzymol. 1975;42:504–511. doi: 10.1016/0076-6879(75)42159-0. [DOI] [PubMed] [Google Scholar]
- Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol. 1995;177:4121–4130. doi: 10.1128/jb.177.14.4121-4130.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilker R, Spiess M, Crottet P. Recognition of sorting signals by clathrin adaptors. Bioessays. 1999;21:558–567. doi: 10.1002/(SICI)1521-1878(199907)21:7<558::AID-BIES4>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- Herman PK, Emr SD. Characterization of VPS34, a gene required for vacuolar protein sorting and vacuole segregation in Saccharomyces cerevisiae. Mol Cell Biol. 1990;10:6742–6754. doi: 10.1128/mcb.10.12.6742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hille-Rehfeld A. Mannose 6-phosphate receptors in sorting and transport of lysosomal enzymes. Biochim Biophys Acta. 1995;1241:177–194. doi: 10.1016/0304-4157(95)00004-b. [DOI] [PubMed] [Google Scholar]
- Honing S, Sosa M, Hille-Rehfeld A, von Figura K. The 46-kDa mannose 6-phosphate receptor contains multiple binding sites for clathrin adaptors. J Biol Chem. 1997;272:19884–19890. doi: 10.1074/jbc.272.32.19884. [DOI] [PubMed] [Google Scholar]
- Huang KM, D'Hondt K, Riezman H, Lemmon SK. Clathrin functions in the absence of heterotetrameric adaptors and AP180-related proteins in yeast. EMBO J. 1999;18:3897–3908. doi: 10.1093/emboj/18.14.3897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarousse N, Kelly RB. Selective inhibition of adaptor complex-mediated vesiculation. Traffick. 2000;1:378–384. doi: 10.1034/j.1600-0854.2000.010502.x. [DOI] [PubMed] [Google Scholar]
- Johnson KF, Kornfeld S. The cytoplasmic tail of the mannose 6-phosphate/insulin-like growth factor-II receptor has two signals for lysosomal enzyme sorting in the Golgi. J Cell Biol. 1992;119:249–257. doi: 10.1083/jcb.119.2.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirchhausen T, Bonifacino JS, Riezman H. Linking cargo to vesicle formation: receptor tail interactions with coat proteins. Curr Opin Cell Biol. 1997;9:488–495. doi: 10.1016/s0955-0674(97)80024-5. [DOI] [PubMed] [Google Scholar]
- Klionsky DJ, Emr SD. A new class of lysosomal/vacuolar protein sorting signals. J Biol Chem. 1990;265:5349–5352. [PubMed] [Google Scholar]
- Klumperman J, Kuliawat R, Griffith JM, Geuze HJ, Arvan P. Mannose 6-phosphate receptors are sorted from immature secretory granules via adaptor protein AP-1, clathrin, and syntaxin 6-positive vesicles. J Cell Biol. 1998;141:359–371. doi: 10.1083/jcb.141.2.359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Le Borgne R, Hoflack B. Mannose 6-phosphate receptors regulate the formation of clathrin-coated vesicles in the TGN. J Cell Biol. 1997;137:335–345. doi: 10.1083/jcb.137.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemmon S, Lemmon VP, Jones EW. Characterization of yeast clathrin and anticlathrin heavy-chain monoclonal antibodies. J Cell Biochem. 1988;36:329–340. doi: 10.1002/jcb.240360403. [DOI] [PubMed] [Google Scholar]
- Marcusson EG, Horazdovsky BF, Cereghino JL, Gharakhanian E, Emr SD. The sorting receptor for yeast vacuolar carboxypeptidase Y is encoded by the VPS10 gene. Cell. 1994;77:579–586. doi: 10.1016/0092-8674(94)90219-4. [DOI] [PubMed] [Google Scholar]
- Marks MS, Ohno H, Kirchhausen T, Bonifacino JS. Protein sorting by tyrosine-based signals: adapting to the Ys and wherefores. Trends Cell Biol. 1997;7:292–297. doi: 10.1016/S0962-8924(96)10057-X. [DOI] [PubMed] [Google Scholar]
- Nothwehr SF, Conibear E, Stevens TH. Golgi and vacuolar membrane proteins reach the vacuole in vps1 mutant yeast cells via the plasma membrane. J Cell Biol. 1995;129:35–46. doi: 10.1083/jcb.129.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owen DJ, Evans PR. A structural explanation for the recognition of tyrosine-based endocytotic signals. Science. 1998;282:1327–1332. doi: 10.1126/science.282.5392.1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Payne GS, Schekman R. Clathrin: a role in the intracellular retention of a Golgi membrane protein. Science. 1989;245:1358–1365. doi: 10.1126/science.2675311. [DOI] [PubMed] [Google Scholar]
- Phan HL, Finlay JA, Chu DS, Tan PK, Kirchhausen T, Payne GS. The Saccharomyces cerevisiae APS1 gene encodes a homolog of the small subunit of the mammalian clathrin AP-1 complex: evidence for functional interaction with clathrin at the Golgi complex. EMBO J. 1994;13:1706–1717. doi: 10.1002/j.1460-2075.1994.tb06435.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piper R, Bryant N, Stevens T. The membrane protein alkaline phosphatase is delivered to the vacuole by a route that is distinct from the VPS-dependent pathway. J Cell Biol. 1997;138:531–546. doi: 10.1083/jcb.138.3.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piper RC, Cooper AA, Yang H, Stevens TH. VPS27 controls vacuolar and endocytic traffic through a prevacuolar compartment in Saccharomyces cerevisiae. J Cell Biol. 1995;131:603–617. doi: 10.1083/jcb.131.3.603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pishvaee B, Munn A, Payne GS. A novel structural model for regulation of clathrin function. EMBO J. 1997;16:2227–2239. doi: 10.1093/emboj/16.9.2227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rad MR, Phan HL, Kirchrath L, Tan PK, Kirchhausen T, Hollenberg CP, Payne GS. Saccharomyces cerevisiae Apl2p, a homologue of the mammalian clathrin AP beta subunit, plays a role in clathrin-dependent Golgi functions. J Cell Sci. 1995;108:1605–1615. doi: 10.1242/jcs.108.4.1605. [DOI] [PubMed] [Google Scholar]
- Redding K, Seeger M, Payne GS, Fuller RS. The effects of clathrin inactivation on localization of Kex2 protease are independent of the TGN localization signal in the cytosolic tail of Kex2p. Mol Biol Cell. 1996;7:1667–1677. doi: 10.1091/mbc.7.11.1667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts CJ, Nothwehr SF, Stevens TH. Membrane protein sorting in the yeast secretory pathway: evidence that the vacuole may be the default compartment. J Cell Biol. 1992;119:69–83. doi: 10.1083/jcb.119.1.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson JS, Klionsky DJ, Banta LM, Emr SD. Protein sorting in Saccharomyces cerevisiae: isolation of mutants defective in the delivery and processing of multiple vacuolar hydrolases. Mol Cell Biol. 1988;8:4936–4948. doi: 10.1128/mcb.8.11.4936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothman JH, Howald I, Stevens TH. Characterization of genes required for protein sorting and vacuolar function in the yeast Saccharomyces cerevisiae. EMBO J. 1989;8:2057–2065. doi: 10.1002/j.1460-2075.1989.tb03614.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothman JH, Stevens TH. Protein sorting in yeast: mutants defective in vacuole biogenesis mislocalize vacuolar proteins into the late secretory pathway. Cell. 1986;47:1041–1051. doi: 10.1016/0092-8674(86)90819-6. [DOI] [PubMed] [Google Scholar]
- Schmid SL. Clathrin-coated vesicle formation and protein sorting: an integrated process. Annu Rev Biochem. 1997;66:511–548. doi: 10.1146/annurev.biochem.66.1.511. [DOI] [PubMed] [Google Scholar]
- Schu PV, Takegawa K, Fry MJ, Stack JH, Waterfield MD, Emr SD. Phosphatidylinositol 3-kinase encoded by yeast VPS34 gene essential for protein sorting. Science. 1993;260:88–91. doi: 10.1126/science.8385367. [DOI] [PubMed] [Google Scholar]
- Seeger M, Payne GS. A role for clathrin in the sorting of vacuolar proteins in the Golgi complex of yeast. EMBO J. 1992a;11:2811–2818. doi: 10.1002/j.1460-2075.1992.tb05348.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeger M, Payne GS. Selective and immediate effects of clathrin heavy chain mutations on Golgi membrane protein retention in Saccharomyces cerevisiae. J Cell Biol. 1992b;118:531–540. doi: 10.1083/jcb.118.3.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sherman F. Getting started with yeast. Methods Enzymol. 1991;194:2–21. doi: 10.1016/0076-6879(91)94004-v. [DOI] [PubMed] [Google Scholar]
- Silveira LA, Wong DH, Masiarz FR, Schekman R. Yeast clathrin has a distinctive light chain that is important for cell growth. J Cell Biol. 1990;111:1437–1449. doi: 10.1083/jcb.111.4.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stack JH, DeWald DB, Takegawa K, Emr SD. Vesicle-mediated protein transport: regulatory interactions between the Vps15 protein kinase and the Vps34 PtdIns 3-kinase essential for protein sorting to the vacuole in yeast. J Cell Biol. 1995;129:321–334. doi: 10.1083/jcb.129.2.321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stack JH, Emr SD. Vps34p required for yeast vacuolar protein sorting is a multiple specificity kinase that exhibits both protein kinase and phosphatidylinositol-specific PI 3-kinase activities. J Biol Chem. 1994;269:31552–31562. [PubMed] [Google Scholar]
- Stepp JD, Huang K, Lemmon SK. The yeast adaptor protein complex, AP-3, is essential for the efficient delivery of alkaline phosphatase by the alternate pathway to the vacuole. J Cell Biol. 1997;139:1761–1774. doi: 10.1083/jcb.139.7.1761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepp J, Pellicena-Palle A, Hamilton S, Kirchhausen T, Lemmon S. A late Golgi sorting function for Saccharomyces cerevisiae Apm1p, but not for Apm2p, a second yeast clathrin AP medium chain-related protein. Mol Biol Cell. 1995;6:41–58. doi: 10.1091/mbc.6.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vowels JJ, Payne GS. A dileucine-like sorting signal directs transport into an AP-3-dependent, clathrin-independent pathway to the yeast vacuole. EMBO J. 1998;17:2482–2493. doi: 10.1093/emboj/17.9.2482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilcox CA, Redding K, Wright R, Fuller RS. Mutation of a tyrosine localization signal in the cytosolic tail of yeast Kex2 protease disrupts Golgi retention and results in default transport to the vacuole. Mol Biol Cell. 1992;3:1353–1371. doi: 10.1091/mbc.3.12.1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanagisawa K, Resnick D, Abeijon C, Robbins PW, Hirschberg CB. A guanosine diphosphatase enriched in Golgi vesicles of Saccharomyces cerevisiae. Purification and characterization. J Biol Chem. 1990;265:19351–19355. [PubMed] [Google Scholar]
- Yeung BG, Phan HL, Payne GS. Adaptor complex-independent clathrin function in yeast. Mol Biol Cell. 1999;10:3643–3659. doi: 10.1091/mbc.10.11.3643. [DOI] [PMC free article] [PubMed] [Google Scholar]