

Abstract
Serotonin 1A receptor (5-HT1AR) agonists reduce both L-DOPA- and D1 receptor (D1R) agonist-mediated dyskinesia, but their anti-dyskinetic mechanism of action is not fully understood. Given that 5-HT1AR stimulation reduces glutamatergic neurotransmission in the dopamine-depleted striatum, 5-HT1AR agonists may diminish dyskinesia in part through modulation of pro-dyskinetic striatal glutamate levels. To test this, rats with unilateral medial forebrain bundle dopamine or sham lesions were primed with L-DOPA (12 mg/kg + benserazide, 15 mg/kg, sc) or the D1R agonist SKF81297 (0.8 mg/kg, sc) until abnormal involuntary movements (AIMs) stabilized. On subsequent test days, rats were treated with vehicle or the 5-HT1AR agonist ±8-OH-DPAT (1.0 mg/kg, sc), followed by L-DOPA or SKF81297, or intrastriatal ±8-OH-DPAT (7.5 or 15 mM), followed by L-DOPA. In some cases, the 5-HT1AR antagonist WAY100635 was employed to determine receptor-specific effects. In vivo microdialysis was used to collect striatal samples for analysis of extracellular glutamate levels during AIMs assessment. Systemic and striatal ±8-OH-DPAT attenuated L-DOPA-induced dyskinesia and striatal glutamate efflux while WAY100635 reversed ±8-OH-DPAT’s effects. Interestingly, systemic ±8-OH-DPAT diminished D1R-mediated AIMs without affecting glutamate. These findings indicate a novel anti-dyskinetic mechanism of action for 5-HT1AR agonists with implications for the improved treatment of Parkinson’s disease.
Keywords: 5-HT1A receptor, D1 receptor, Glutamate, Dyskinesia, L-DOPA, Parkinson’s disease
Introduction
Chronic dopamine (DA) replacement therapy with L-3,4-dihydroxyphenylalanine (L-DOPA) for Parkinson’s disease (PD) patients often results in abnormal and excessive movements known as L-DOPA-induced dyskinesia (LID; Jankovic, 2005). Although the mechanisms of LID are not fully understood, it is believed that following DA depletion, serotonergic neurons of the raphe nuclei convert exogenously administered L-DOPA to DA and release it into the striatum in a pulsatile, unregulated manner (Carta et al., 2007; Eskow et al., 2009; Lindgren et al., 2010). This aberrant release of DA is believed to stimulate supersensitive DA D1 (D1R) and D2 (D2R) receptors located in the DA-depleted striatum (Pavese et al., 2006; Cenci, 2007). While both receptor subtypes appear to be involved in LID, it is likely that striatal D1R have a more prominent role (Westin et al., 2007). For instance, striatal D1R expression and signaling have been shown to be significantly enhanced in dyskinetic animals and humans (Cenci et al., 1998; Gerfen et al., 2002; Aubert et al., 2005; Guigoni et al., 2007), and D1R agonists induce dyskinesia in both experimental and clinical models of PD (Rascol et al., 2001, 2006; Delfino et al., 2007; Dupre et al., 2007, 2008a).
It is well known that serotonin (5-HT) 1A receptor (5-HT1AR) agonists diminish LID (Dekundy et al., 2007; Eskow et al., 2007, 2009) and these results have been mostly attributed to stimulation of raphe 5-HT1AR that temper striatal DA release. Interestingly, there is also evidence that stimulation of 5-HT1AR located directly within the striatum attenuates both L-DOPA- (Bishop et al., 2009) and D1R-mediated dyskinesia (Dupre et al., 2008a) and improves movement in DA-depleted rats (Mignon & Wolf, 2002; Matsubara et al., 2006; Dupre et al., 2008a). The mechanism(s) surrounding these striatally-mediated 5-HT1AR effects are not yet known. One possibility is that activation of these receptors, located presynaptically on corticostriatal glutamate neurons, attenuate the release of glutamate into the striatum (Antonelli et al., 2005; Mignon & Wolf, 2005). Indeed, upon DA depletion and subsequent L-DOPA or D1R agonist treatment, augmentation of striatal glutamate levels (Jonkers et al., 2002; Robelet et al., 2004) and increased expression of striatal glutamate receptors have been postulated to result in dyskinetic behaviors (Calon et al., 2002; Ouattara et al., 2010). In support of this, a number of ionotropic and metabotropic glutamate receptor antagonists have been shown to reduce LID and improve D1R-mediated locomotor activity (Goodwin et al., 1992; Ferré et al., 1994; Bibbiani et al., 2005; Rylander et al., 2010; Kobylecki et al., 2010). Thus, whether striatal 5-HT1AR stimulation attenuates LID and D1R agonist-induced dyskinesia through modulation of local glutamate release remains an important mechanistic and translational question.
The aim of the current study was to investigate the effects of systemic and local 5-HT1AR stimulation on extracellular striatal glutamate levels in hemiparkinsonian rats rendered dyskinetic by either L-DOPA or the D1R agonist SKF81297. Using in vivo microdialysis, the full 5-HT1AR agonist (±)-8-Hydroxy-2-(dipropylamino)tetralin hydrobromide (±8-OH-DPAT) was administered systemically or striatally perfused prior to L-DOPA (12 mg/kg, sc + benserazide, 15 mg/kg, sc) or administered systemically prior to R(+)-SKF-81297 hydrobromide (SKF81297; 0.8 mg/kg, sc) in unilaterally DA-depleted and sham-lesioned rats. Striatal sample fractions were collected for analysis of glutamate and dyskinesia was measured using the abnormal involuntary movements (AIMs) scale (Lundblad et al., 2002). The present results indicate that the anti-dyskinetic effect of 5-HT1AR stimulation coincides with a reduction in extracellular striatal glutamate levels with L-DOPA, but not D1R agonist, treatment. These findings implicate a novel glutamatergic mechanism by which 5-HT1AR agonists work to reduce LID with implications for the treatment of PD.
Materials and methods
Animals
Adult male Sprague-Dawley rats (N=61) were used (225–250 g upon arrival; Taconic Farms, Hudson, NY, USA). Rats were kept in plastic cages (22 cm high, 45 cm deep and 23 cm wide) and given free access to food (Rodent Diet 5001; Lab Diet, Brentwood, MO, USA) and water. The colony room was kept on a 12 hr light/dark cycle (light on at 0700 hr) and maintained at 22–23°C. The guidelines of the Institutional Animal Care and Use Committee of Binghamton University and the “Guide for the Care and Use of Laboratory Animals” (Institute of Laboratory Animal Resources, National Academic Press 1996; NIH publication number 85-23, revised 1996) were maintained throughout the study.
Medial forebrain bundle 6-hydroxydopamine lesion and microdialysis guide cannulae implantation surgeries
One week after arrival, rats in Experiments 1 and 2 received unilateral DA (n=24) or sham (n=18) lesions of the left medial forebrain bundle (MFB). All rats in Experiment 3 received DA lesions of the left MFB (n=20). Each rat was administered desipramine HCl (25 mg/kg, ip; Sigma, St. Louis, MO, USA) 30 minutes prior to surgery in order to protect norepinephrine neurons. Rats were anesthetized with inhalant isoflurane (2–3%; Sigma) in oxygen (2.5 L/min) and placed in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA, USA). The following coordinates relative to bregma were used for the site of injection: AP, −1.8 mm; ML, +2.0 mm; DV, −8.6 mm, with the incisor bar positioned at 5.0 mm below interaural line (Paxinos & Watson, 1998). After drilling a small hole in the skull above the site of injection, a 10 µl Hamilton syringe attached to a 26 gauge needle was lowered into the target. At that point, 4 µl of vehicle (0.9% sodium chloride (NaCl) + 0.1% ascorbic acid) or 6-hydroxydopamine hydrobromide (6-OHDA; 3 µg/µl; Sigma) was injected at a rate of 2 µl/min for rats in Experiments 1 and 2, whereas all rats in Experiment 3 received 6-OHDA. The needle was withdrawn 5 min later. During the same surgery, rats were fitted unilaterally with plastic microdialysis guide cannulae (CMA 12 Elite; Stockholm, Sweden) targeting the dorsal striatum ipsilateral to the lesioned side (AP, +1.2 mm; ML, +2.5 mm; DV, −3.7 mm; relative to bregma; Paxinos & Watson, 1998). Cannulae were positioned and affixed to the skull with screws and liquid and powder dental acrylic (Lang Dental, Wheeling, IL). At the completion of surgery, animals were single housed, placed in clean cages and allowed to recover with ad lib food and water. Five min pre-surgery and 1 hr and 1 day post-surgery, rats received an injection of Buprenex (buprenorphine HCl; 0.03 mg/kg, ip; Reckitt Benckiser Pharmaceuticals Inc., Richmond, VA) as analgesic treatment. Soft chow was also provided and rats were monitored and handled twice per week for 3 weeks post-surgery in order to ensure full recovery and acclimation to experimenters.
Pharmacological treatments and in vivo microdialysis procedure
Experiment 1: Effects of systemic 5-HT1AR stimulation on extracellular striatal glutamate levels in L-DOPA-induced dyskinesia
Three weeks after 6-OHDA (n=14) or sham (n=11) lesions of the MFB and unilateral striatal microdialysis cannulations, rats in the first experiment received injections of L-3,4-dihydroxyphenylalanine methyl ester hydrochloride (L-DOPA; 12 mg/kg, sc; Sigma) + DL-serine 2-(2,3,4-trihydroxybenzyl) hydrazide hydrochloride (benserazide; 15 mg/kg, sc; Sigma) once daily for 7 days. The dose of L-DOPA and the length of priming have been extensively utilized in our lab to produce prominent and stable AIMs expression (Eskow et al., 2007; Dupre et al., 2008b; Bishop et al., 2009). L-DOPA and benserazide were dissolved in vehicle (0.9% NaCl + 0.1% ascorbic acid). On the final day of priming, AIMs (see description below) were observed every 20 min for 3 h immediately after L-DOPA injections. 6-OHDA-lesioned rats displaying an AIMs score of ≥25 by the 7th day of L-DOPA priming were retained for further study (n=12).
Microdialysis testing commenced 2 days after the last day of L-DOPA priming. On test day, striatal probes (CMA 12 Elite; membrane length = 3 mm; 20000 Dalton; Stockholm, Sweden) were inserted into rats’ guide cannulae and locked into place so that the dialysis membrane extended −3.7 to −6.7 ventral to bregma within the striatum. After 60 min of probe stabilization (2.0 µl/min of artificial cerebral spinal fluid (aCSF) (in mM: 128 NaCl, 2.5 KCl, 1.3 CaCl2, 2.1 MgCl2, 0.9 NaH2PO4, 2.0 Na2HPO4, and 1.0 glucose, brought to a pH of 7.4), striatal dialysate samples were collected every 20 min for 40 min to determine baseline levels of glutamate. At this point, rats received a systemic treatment injection of vehicle (0.9% NaCl + 0.1% ascorbic acid, sc) and sample fractions were collected every 20 min for 2 h to determine any changes in extracellular glutamate levels due to systemic injection. Following this, using a counter-balanced design, rats received systemic treatment of: Vehicle (0.9% NaCl), the full 5-HT1AR agonist ±8-OH-DPAT (1.0 mg/kg, sc; Sigma), or combined ±8-OH-DPAT (1.0 mg/kg, sc) + the 5-HT1AR antagonist N-[2-[4-(2-Methoxyphenyl)-1-piperazinyl]ethyl]-N-2-pyridinylcyclohexanecarboxamide maleate salt (WAY100635; 0.5 mg/kg, sc; Sigma), immediately followed by L-DOPA (12 mg/kg, + benserazide, 15 mg/kg, sc). Sample fractions were collected every 20 min for 3 h and AIMs were concurrently observed during this time. Each rat underwent this microdialysis procedure for 2 consecutive days. No differences in glutamate nor AIMs were found in animals treated with Vehicle + L-DOPA on microdialysis test day 1 versus test day 2 (data not shown). In addition, drug clearance of ±8-OH-DPAT by day 2 of testing was ensured due to its relatively short half-life of approximately 45 min (Yu & Lewander, 1997). Finally, a post-test with L-DOPA alone was performed at the end of the study to ensure stable AIMs throughout testing (Fig 1).
Figure 1. L-DOPA- and D1R-mediated priming and post-test AIMs.
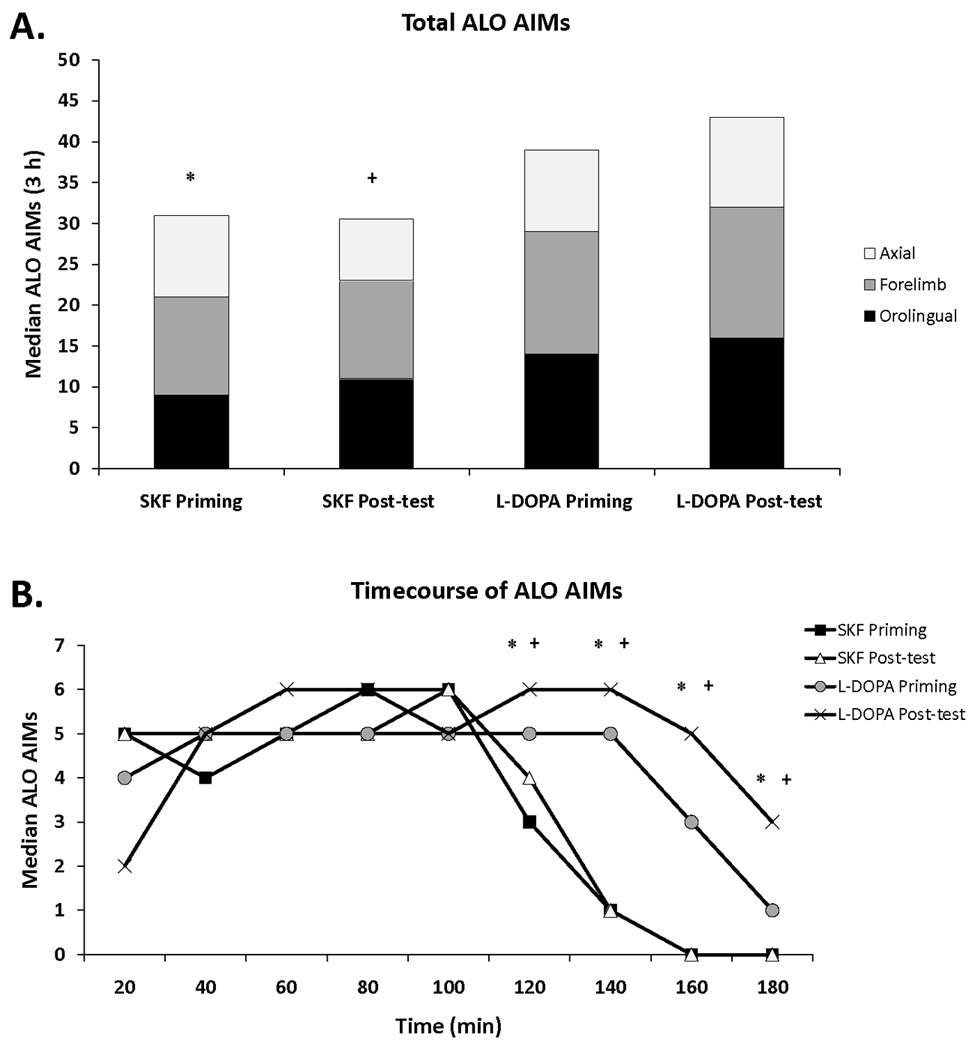
(A) Rats that received unilateral 6-OHDA lesions of the MFB were primed 3 weeks later with L-DOPA (12 mg/kg + benserazide, 15 mg/kg, sc; Experiments 1 and 3; n = 30) or the D1R agonist SKF81297 (SKF; 0.8 mg/kg,sc; Experiment 2; n = 8) and exhibited summed axial, forelimb, and orolingual (ALO) AIMs ≥ 25 by the last day of priming (3 h total). Following microdialysis test days, rats were given a post-test of L-DOPA or SKF alone in order to ensure AIMs stability over the course of testing. The distribution of axial (white), forelimb (gray), and orolingual (black) AIMs are represented. (B) Timecourse of AIMs for the last day of priming and the post-test of 6-OHDA-lesioned animals treated with L-DOPA or SKF. Differences in AIMs (expressed as medians) were analyzed with non-parametric Kruskal-Wallis tests and Mann-Whitney post hoc comparisons.
* p < 0.05 for SKF priming vs L-DOPA priming
+ p < 0.05 for SKF post-test vs L-DOPA post-test
Experiment 2: Effects of systemic 5-HT1AR stimulation on extracellular striatal glutamate levels in D1R agonist-mediated dyskinesia
Three weeks after 6-OHDA (n=10) or sham (n=7) lesions of the MFB and unilateral striatal microdialysis cannulations, rats in the second experiment received injections of the D1R agonist SKF81297 (0.8 mg/kg, sc; Sigma), dissolved in 20% dimethyl sulfoxide (DMSO) in 0.9% NaCl, on 3 separate occasions 2–3 days apart in order to sensitize D1R (Pollack & Yates, 1999; Dupre et al., 2007). The dose of SKF81297 and priming regimen have been used in our lab to produce stable AIMs expression that is similar to the AIMs induced by our current dose of L-DOPA (Dupre et al., 2007; Dupre et al., 2008a). AIMs were observed every 20 min for 3 h immediately after injections. 6-OHDA-lesioned rats displaying an AIMs score of ≥25 by the 3rd day of D1R priming were retained for further study (n=9). Microdialysis testing commenced 2 days after the last day of SKF81297 priming.
Rats in Experiment 2 followed a similar microdialysis procedure as those in Experiment 1. Following baseline sampling, rats received a systemic treatment injection of vehicle (20% DMSO in 0.9% NaCl, sc) and sample fractions were collected every 20 min for 2 h. At this point, using a counter-balanced, within-subjects design, rats received systemic treatment of vehicle (0.9% NaCl) or ±8-OH-DPAT (1.0 mg/kg, sc) immediately followed by SKF81297 (0.8 mg/kg, sc). Sample fractions were collected every 20 min for 3 h and AIMs were concurrently observed during this time. Each rat underwent this microdialysis procedure for 2 consecutive days and no differences in glutamate nor AIMs were found in animals treated with Vehicle + SKF81297 on microdialysis test day 1 versus test day 2 (data not shown). A post-test with SKF81297 alone was performed at the end of the study to ensure that there were stable AIMs throughout testing (Fig 1).
Experiment 3: Effects of intrastriatal 5-HT1AR stimulation on local extracellular glutamate levels in L-DOPA-induced dyskinesia
Three weeks after 6-OHDA lesions of the MFB and unilateral striatal microdialysis cannulations, rats in the third experiment received injections of L-DOPA (12 mg/kg, benserazide, 15 mg/kg, sc) once daily for 7 days. On the final day of priming, AIMs were observed every 20 min for 3 h immediately after L-DOPA injections. Rats displaying an AIMs score of ≥25 by the 7th day of L-DOPA priming were retained for further study (n=18).
Microdialysis testing commenced 2 days after the last day of L-DOPA priming and followed a similar procedure as that described in Experiment 1. After baseline sampling, rats received a systemic treatment injection of vehicle (0.9% NaCl + 0.1% ascorbic acid, sc) and sample fractions were collected every 20 min for 2 h. Following this, using a counter-balanced design, rats received intrastriatal infusion of: Vehicle (aCSF), the full 5-HT1AR agonist ±8-OH-DPAT (7.5 or 15 mM), or combined ±8-OH-DPAT (15 mM) + WAY100635 (4.6 mM), followed 10 min later (when drug reached brain) by systemic treatment injections of L-DOPA (12 mg/kg, + benserazide, 15 mg/kg, sc). Sample fractions were collected every 20 min for 3 h and AIMs were concurrently observed during this time. Each rat underwent this microdialysis procedure no more than 2 times and a post-test with L-DOPA alone was performed at the end of the study to ensure stable AIMs throughout testing (Fig 1).
Abnormal Involuntary Movements
Rats were monitored for AIMs using a procedure similarly described in Dupre et al. (2008a; 2008b). The AIMs model of dyskinesia utilizes distinct behavioral measures and demonstrates face validity with known anti-dyskinetic compounds (Lundblad et al., 2002; Dekundy et al., 2007). AIMs can also be maintained over repeated testing by separating experimental days after initial priming (Bishop et al., 2006; Taylor et al., 2006). On the last day of priming (0900–1700 h), rats were individually placed in clear plastic cylinders (20 × 25 cm) immediately following L-DOPA or SKF81297 injection. On microdialysis test days (0800–1800 h), animals received treatment injections while in clear plastic round bottom microdialysis bowls (CMA Microdialysis Inc.). Following injections, a trained observer assessed each rat for exhibition of axial, limb, and orolingual AIMs. Dystonic posturing of the neck and torso, involving positioning of the neck and torso in a twisted manner directed toward the side of the body contralateral to the lesion, were referred to as ‘axial’ AIMs. ‘Limb’ AIMs were defined as rapid, purposeless movements of the forelimb located on the side of the body contralateral to the lesion. ‘Orolingual’ AIMs were composed of repetitive openings and closings of the jaw and tongue protrusions. The movements were considered abnormal as they occurred at times when the rats were not chewing or gnawing on food or other objects. Every 20th min for 3 h, rats were observed for 1 min for AIMs. During this time, a severity score of 0–4 was assigned for each AIMs category: 0, not present; 1, present for <50% of the observation period (i.e. 1–29 s); 2, present for >50% or more of the observation period (i.e. 30–59 s); 3, present for the entire observation period (i.e. 60 s) and interrupted by a loud stimulus (a tap on the cylinder), or 4, present for the entire observation period but not interrupted by a loud stimulus. Thus, the theoretical maximum score for each type of AIM was 36 (4 × 9 time periods), although observed scores were never this severe. The scores of the 3 AIMs subcategories (axial, limb, and orolingual) were summed (with a theoretical maximum potential score of 12 per time period (4 × 3 AIMs subcategories) and an overall maximum score of 108 for the entire 3 h).
Histology and neurochemical analyses
Tissue dissection and cryostat sectioning
When experiments were completed, a subset of sham-lesioned (n=7) and 6-OHDA-lesioned (n=7) rats were sacrificed by decapitation and brains were immediately removed. Anterior and central striata from these rats were visually examined for verification of striatal placements. To determine the level of DA depletion, posterior striata were freshly dissected, frozen at −80°C and subjected to monoamine analysis using high performance liquid chromatography with electrochemical detection (HPLC-ED; see below). Striata from all other rats were examined for verification of striatal placements using a cryostat. During dissection, anterior and central striata were removed and rapidly frozen in methylbutane (−30°C) and stored at −20°C. Cresyl violet (FD Neurotechnologies, Baltimore, MD) staining was used to determine injection sites and extent of gliosis from cryostat-generated 20 mm coronal sections containing injection sites that were post-fixed with 4% paraformaldehyde (PFA; Fisher Scientific, Hanover Park, IL). All rats that completed these studies were found to have probe placements within the dorsocentral or dorsolateral aspects of the striatum (data not shown).
High-performance liquid chromatography for DOPAC and DA tissue analyses
Reverse-phase HPLC-ED was performed on striatal tissue, obtained from a subset of rats, according to protocol of Kilpatrick et al. (1986), a method for semiautomated catecholamine and indoleamine analysis with coulometric detection. The system included an ESA autoinjector (Model 542; Chelmsford, MA, USA), an ESA solvent delivery system (582), an external pulse dampener (ESA), an ESA column and a MD-150X3.2 (150 × 3.2 mm, 3 µm packing) column (ESA). Samples were homogenized in ice-cold perchloric acid (0.1 M) with 1% ethanol and 0.02% ethylenediaminetetraacetic acid (EDTA). The homogenates were spun for 30 min at 14,000 g with the temperature maintained at 4°C. Aliquots of supernatant were then analyzed for abundance of 3,4-dihydroxyphenylacetic acid (DOPAC) and DA. Samples were separated using a mobile phase composed of sodium phosphate (monobasic, anhydrous), 100 mM; EDTA, 0.05 mM; octane sulphonic acid, 1.4 mM; and acetonitrile, 9% adjusted to pH 3.0 with o-phosphoric acid. A coulometric detector configured with three electrodes (Coulochem III; ESA) measured the content of DOPAC and DA. An ESA model 5020 guard cell (+300 mV) was positioned prior to the autoinjector. The analytical cell (ESA model 5011A; first electrode at −100 mV, second electrode at +250 mV) was located immediately after the column. The second analytical electrode emitted signals that were recorded and analyzed by EZChrom Elite software via Scientific Software Inc. module (SS420x). The final oxidation current values were compared to known standards 10−6–10−9, adjusted to striatal tissue weights, and expressed as nanograms (ng) of DOPAC or DA per milligram (mg) tissue (mean ± SEM).
High-performance liquid chromatography for glutamate dialysate samples
Dialysate samples from all rats were analyzed via HPLC-ED for extracellular glutamate levels. This method, similarly described in Donzanti and Yamamoto (1988) and Rowley et al. (1995), employed electrochemical detection of amino acid derivatives formed by a chemical reaction with o-phthaldialdehyde (OPA) and β-mercaptoethanol (β-ME). The system included an ESA autoinjector (Model 542; Chelmsford, MA, USA), an external pulse dampener (ESA), and an XTerra MSC18 (3.0 × 50 mm, 2.5 µm) column. The OPA/β-ME derivatizing reagent (15 µl) reacted with striatal dialysate samples (30 µl) and 20 µl of the derivatized sample was analyzed for abundance of glutamate. The mobile phase consisted of 100 mM sodium phosphate (dibasic, anhydrous) with 20% by volume methanol and 3.5% by volume acetonitrile, filtered through 0.2 µm nylon membrane filter (Nylaflo; Pall Life Sciences) and adjusted to pH 6.75 with o-phosphoric acid and/or sodium hydroxide. An ESA model 5020 guard cell (+700 mV) was positioned prior to the autoinjector. An analytical cell (ESA model 5014B; first electrode +150 mV, second electrode +400 mV) located immediately after the column was controlled by a coulometric detector configured with three electrodes (Coulochem III; ESA). The areas of oxidation current peaks generated at the second electrode by eluting amino acid derivatives was analyzed by EZChrom Elite software via a Scientific Software Inc. module (SS420x). The oxidation current values were converted to masses (ng of amino acid) using standard curves (2e−6-1e−8 M). The determination of probe recovery (±30%) was performed in vitro by perfusing standard solutions (2e−6-1e−8 M).
Data analyses
DOPAC and DA levels (expressed as mean ng/mg ± SEM) in the striatum were analyzed with one-between (lesion type) and one-within factor repeated-measures (hemisphere) ANOVAs. Basal levels (expressed as means ± SEM) for striatal glutamate concentrations (M) were analyzed with a between-subjects 2×2 two-way ANOVA (lesion type × priming condition). Striatal glutamate (% baseline) was analyzed with a one-between (treatment group) and one-within factor repeated-measures (sample time [20 min samples over 340 min total = 17 sample times]) ANOVA. Treatment effects (expressed as means ± SEM) for striatal glutamate concentrations (M) were analyzed with between-subjects one-way ANOVAs. AIMs (expressed as medians) were analyzed by employing non-parametric Kruskal-Wallis tests. Significant differences between groups were determined by Mann-Whitney post hoc comparisons (Pett, 1997) for AIMs and planned comparison tests for striatal DOPAC, DA, and glutamate (% baseline and M). Analyses were performed with the use of Statistica software ’98 (Statsoft Inc., Tulsa, OK, USA) and alpha was set at p<0.05.
Results
Striatal DOPAC and DA levels
Subsets of 6-OHDA- and sham-lesioned animals were examined by HPLC-ED for determination of average DOPAC and DA content (see Table 1). Mixed design two-way ANOVAs [2 lesion types (between) × 2 hemispheres (within)] of striatal DOPAC and DA revealed main effects of lesion (DOPAC: F1,12=11.57; DA: F1,12=9.52; p<0.01), hemisphere (DOPAC: F1,12=24.53; DA: F1,12=31.57; p<0.01), and lesion × hemisphere interactions (DOPAC: F1,12=10.78; DA: F1,12=18.08; p<0.01). Planned comparison analyses showed that unilateral 6-OHDA injection into the MFB produced significant reductions in lesioned striatal DOPAC and DA compared to the intact striatum, 98.3% and 99.7%, respectively. Unilateral 6-OHDA injection into the MFB also produced significant reductions in lesioned striatal DOPAC and DA compared to the intact and lesioned striata of sham-lesioned animals (all p<0.05). Injections of vehicle into the MFB did not produce significant reductions in sham-lesioned striatal DOPAC or DA compared to the intact striatum.
Table 1.
Effects of unilateral medial forebrain bundle (MFB) 6-OHDA or sham lesion on concentrations of striatal DOPAC and DA.
| Intact vs. lesioned | DOPAC (ng/mg) | DA (ng/mg) |
|---|---|---|
| Subset 6-OHDA-lesioned (n=7) | ||
| Intact (right) | 3.42 (± 0.16) | 11.34 (± 1.44) |
| Lesioned (left) | 0.06 (± 0.01)*+ | 0.03 (± 0.01)*+ |
| (Lesioned/intact %) | 1.70% | 0.30% |
| Subset sham-lesioned (n=7) | ||
| Intact (right) | 4.09 (± 0.74) | 10.30 (± 1.49) |
| Lesioned (left) | 3.41 (± 0.79) | 8.74 (± 1.53) |
| (Lesioned/intact %) | 88.45% | 92.22% |
Values are nanogram DOPAC or DA per milligram protein and percentage of intact striata. Differences between group means were determined by two-way ANOVAs and planned comparison analyses.
p<0.05 compared to intact sides;
p<0.05 compared to lesioned side (sham)
L-DOPA- and D1R-mediated priming and post-test AIMs
Animals that received 6-OHDA-lesions were primed with L-DOPA (Experiments 1 and 3) or SKF81297 (Experiment 2) and developed stable but quantitatively differential dyskinesia (H3 = 30.16; p<0.05). Post hoc analyses revealed that within treatment, L-DOPA- and SKF81297-induced AIMs on the last day of priming did not differ from their respective post-test summed AIMs (180 min total). In contrast, L-DOPA-induced priming and post-test summed AIMs were significantly higher than those of SKF81297 (both p<0.05; Fig 1A). The difference in overall summed AIMs was explained in the timecourse of AIMs as shown in Figure 1B. While L-DOPA provoked dyskinesia for the entire 180 min, SKF81297-mediated AIMs diminished rapidly following 120 min and ceased by 160 min. Importantly, there were no differences in axial, forelimb, nor orolingual AIMs between L-DOPA and SKF81297 priming. Therefore, L-DOPA and SKF81297 priming produced similar severities of axial, forelimb, and orolingual AIMs, but likely due to differences in pharmacokinetics, L-DOPA’s effects lasted longer.
Basal extracellular striatal glutamate levels following priming
Basal concentrations of striatal glutamate of 6-OHDA- and sham-lesioned rats in Experiments 1 (L-DOPA-primed) and 2 (SKF81297-primed) are shown in Figure 2. A between-subjects two-way ANOVA revealed a main effect of lesion (F1,119=21.53, p<0.001), but no main effect of priming treatment and no lesion × priming treatment interaction. Thus, unilateral 6-OHDA injections into the MFB produced significant reductions in basal striatal glutamate concentrations, regardless of priming treatment.
Figure 2. Effects of 6-OHDA-lesion, L-DOPA priming, and D1R agonist priming on basal extracellular striatal glutamate levels.
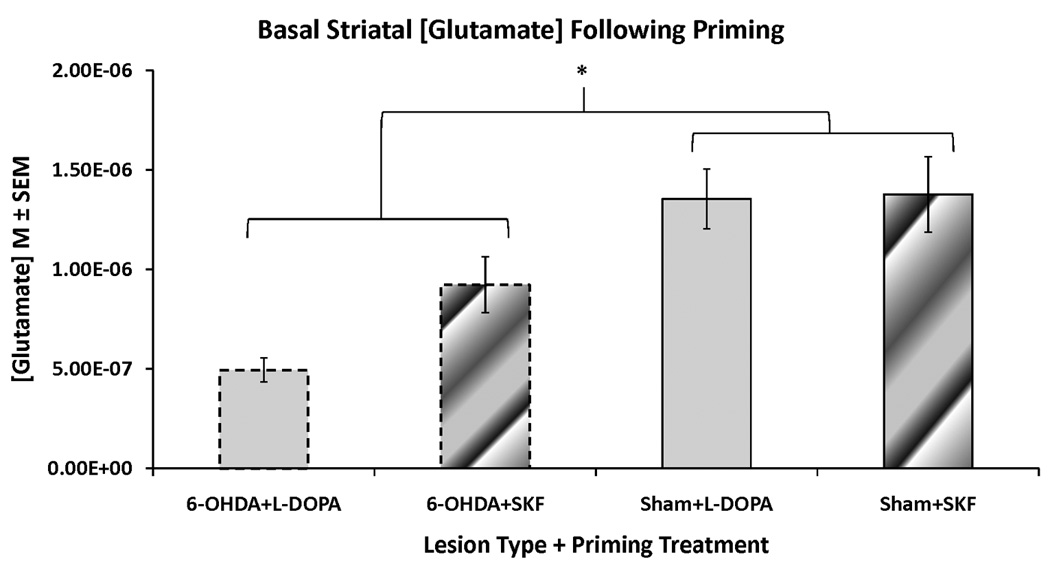
Rats in the first and second experiments received unilateral 6-OHDA or sham lesions of the MFB and 3 weeks later were primed with L-DOPA (12 mg/kg + benserazide, 15 mg/kg, sc) or the D1R agonist SKF81297 (SKF; 0.8 mg/kg,sc). Following priming, rats underwent a microdialysis procedure including 40 min of baseline sampling prior to any vehicle or drug injection. Striatal glutamate concentrations (M) from these baseline samples were averaged and analyzed with a two-way ANOVA (lesion group × priming treatment). The groups include: 6-OHDA+L-DOPA (n=12), 6-OHDA+SKF (n=9), Sham+L-DOPA (n=11), and Sham+SKF (n=7).
* p < 0.05 for 6-OHDA-lesioned groups vs sham-lesioned groups
Experiment 1: Systemic 5-HT1AR stimulation diminishes L-DOPA-induced striatal glutamate efflux while concurrently reducing AIMs in 6-OHDA-lesioned rats
The effects of systemic 5-HT1AR stimulation on striatal glutamate in DA-depleted, chronically L-DOPA-treated rats was examined by injecting vehicle or ±8-OH-DPAT (1.0 mg/kg, sc) immediately prior to L-DOPA treatment. A mixed design two-way ANOVA [3 treatment groups (between) × 17 time points (within)] revealed a main effect of time (F16,272=10.93, p<0.01) and a treatment × time interaction (F32,272=3.19, p<0.01; Fig 3A). Planned comparisons revealed that pretreatment with ±8-OH-DPAT significantly reduced striatal glutamate compared to L-DOPA alone at time points 120–260 (all p<0.05). Co-administration of the full 5-HT1AR antagonist WAY100635 with ±8-OH-DPAT reduced ±8-OH-DPAT’s effects at the 160th and 180th min (both p<0.05). In sham-lesioned rats, a main effect of time for extracellular striatal glutamate levels was found (F16,256=50.78, p<0.001), but there was no main effect of treatment nor a treatment × time interaction (Fig 3D). Since extracellular striatal glutamate levels lessened over sampling time in both lesion groups, main effects of time did not interfere with the treatment effects observed.
Figure 3. Effects of 5-HT1AR stimulation on L-DOPA-related striatal glutamate and AIMs in 6-OHDA- and sham-lesioned rats.
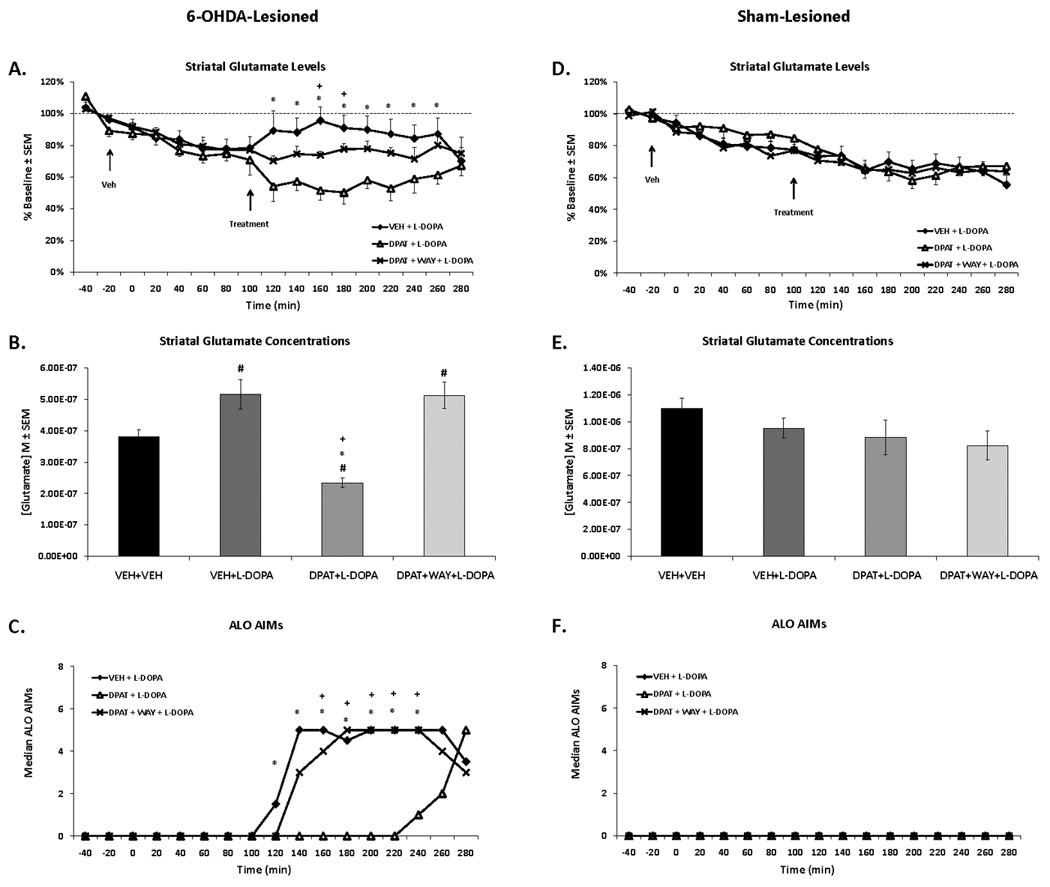
Rats in the first experiment received unilateral 6-OHDA or sham lesions of the MFB and 3 weeks later were primed with L-DOPA (12 mg/kg + benserazide, 15 mg/kg, sc) once daily for 7 d. Dyskinetic (n=12) and sham-lesioned rats (n=11) underwent a microdialysis procedure including: 40 min baseline, 120 min vehicle treatment, 120 min drug treatment, and 60 min post-drug treatment sampling (dialysate collected every 20 min). Drug treatment included: Vehicle (VEH; 0.9% NaCl), the full 5-HT1AR agonist ±8-OH-DPAT (DPAT; 1.0 mg/kg, sc), or combined DPAT + the 5-HT1AR antagonist WAY100635 (WAY; 0.5 mg/kg, sc), immediately followed by L-DOPA (12 mg/kg, + benserazide, 15 mg/kg, sc). AIMs were observed during this time. Lines depict the means ± SEM of striatal glutamate (percent baseline) (A) and AIMs (C) for VEH+L-DOPA (n=8), DPAT+L-DOPA (n=7), and DPAT+WAY+L-DOPA (n=5) in 6-OHDA-lesioned rats. Results for sham-lesioned rats receiving VEH+L-DOPA (n=7), DPAT+L-DOPA (n=6), and DPAT+WAY+L-DOPA (n=6) are expressed in (D) and (F). Bars depict the means ± SEM concentrations of striatal glutamate (M) during the 120 min of vehicle (VEH+VEH) and 120 min of drug treatments in 6-OHDA- (B) and sham-lesioned (E) rats. Effects over time were determined by two-way mixed design ANOVAs for striatal glutamate (% baseline). Striatal glutamate concentrations (M) were analyzed with one-way ANOVAs. Treatment effects for AIMs (expressed as medians) were analyzed by employing non-parametric Kruskal-Wallis tests. Significant differences between treatments were determined by Mann-Whitney post hoc comparisons for AIMs and planned comparison tests for striatal glutamate (% baseline and M).
* p < 0.05 for VEH + L-DOPA vs DPAT + L-DOPA
+ p < 0.05 for DPAT + L-DOPA vs DPAT + WAY + L-DOPA
# p < 0.05 vs VEH + VEH (bar graph)
The average concentrations of striatal glutamate during 120 min of vehicle (time points 0–100) and 120 min of treatment (time points 120–220) are portrayed in Figure 3B and 3E. A one-way ANOVA showed a main effect of treatment in 6-OHDA-lesioned rats (F3,205=12.11, p<0.01; Fig 4B) but not sham-lesioned rats (Fig 4E). Post hoc analyses showed that L-DOPA significantly increased striatal glutamate concentrations compared to vehicle treatment, whereas pre-treatment with ±8-OH-DPAT significantly lowered glutamate concentrations compared to all treatments in 6-OHDA-lesioned rats (all p<0.05). Importantly, WAY100635 reversed ±8-OH-DPAT’s suppressant effects while maintaining L-DOPA’s enhancement of striatal glutamate when compared to vehicle treatment (p<0.05).
Figure 4. Effects of 5-HT1AR stimulation on D1R agonist-related striatal glutamate and AIMs in 6-OHDA- and sham-lesioned rats.
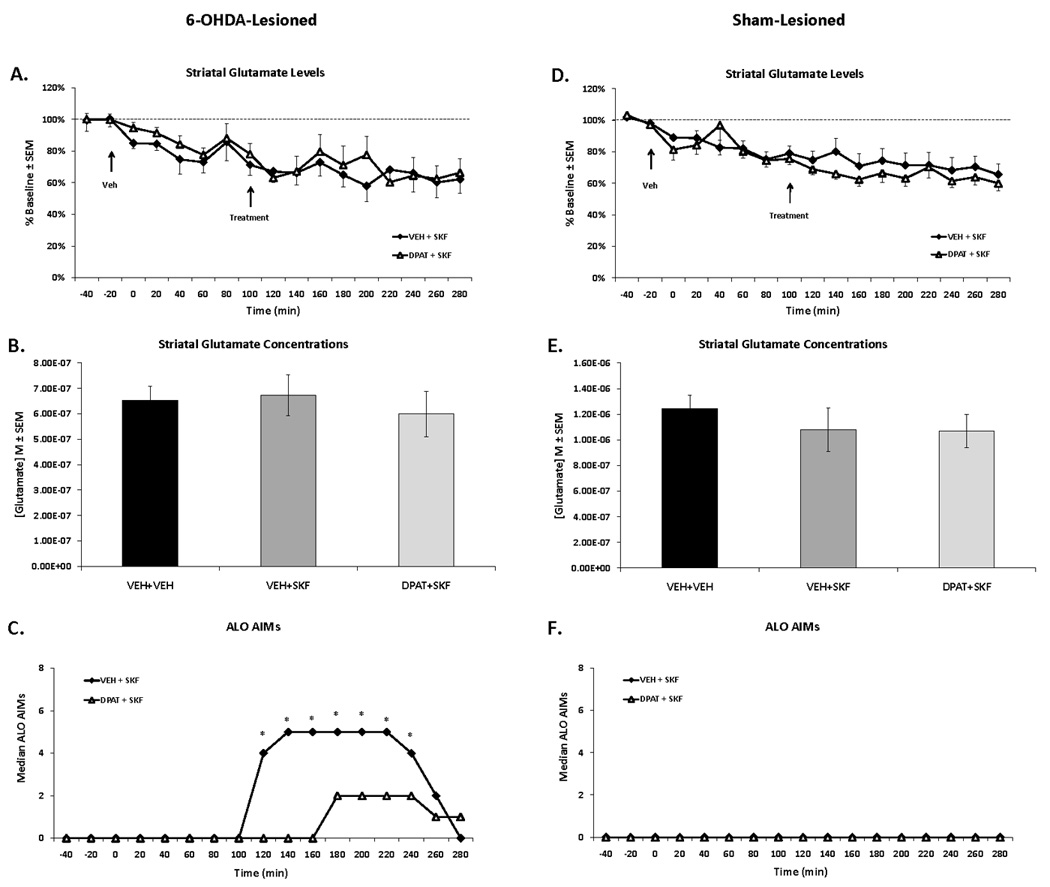
Rats in the second experiment received unilateral 6-OHDA or sham lesions of the MFB and 3 weeks later were primed with the D1R agonist SKF81297 (SKF; 0.8 mg/kg, sc) every 2–3 d for a total of 3 d. Dyskinetic (n=9) and sham-lesioned rats (n=7) underwent a microdialysis procedure including: 40 min baseline, 120 min vehicle treatment, 120 min drug treatment, and 60 min post-drug treatment sampling (dialysate collected every 20 min). Drug treatment included: Vehicle (VEH; 0.9% NaCl) or the full 5-HT1AR agonist ±8-OH-DPAT (DPAT; 1.0 mg/kg, sc), immediately followed by SKF (0.8 mg/kg, sc). AIMs were observed during this time. Lines depict the means ± SEM of striatal glutamate (A) and AIMs (C) for VEH+SKF (n=9) and DPAT+SKF (n=9) in 6-OHDA-lesioned rats. Results for sham-lesioned rats receiving VEH+SKF (n=6) and DPAT+SKF (n=7) are expressed in (D) and (F). Bars depict the means ± SEM concentrations of striatal glutamate (M) during the 120 min of vehicle (VEH+VEH) and 120 min of drug treatments in 6-OHDA- (B) and sham-lesioned (E) rats. Effects over time were determined by two-way mixed design ANOVAs for striatal glutamate (% baseline). Striatal glutamate concentrations (M) were analyzed with oneway ANOVAs. Treatment effects for AIMs (expressed as medians) were analyzed by employing non-parametric Kruskal-Wallis tests. Significant differences between treatments were determined by Mann-Whitney post hoc comparisons for AIMs and planned comparison tests for striatal glutamate (% baseline and M).
* p < 0.05 for VEH+SKF vs DPAT+SKF
While there was no expression of AIMs with any treatment in sham-lesioned rats (Fig 3F), main effects for AIMs in 6-OHDA-lesioned rats were observed at the 120–240th min (all H2>9.0; all p<0.05). As shown in Figure 3C, post hoc analyses revealed that ±8-OH-DPAT significantly attenuated L-DOPA-induced AIMs at all time points compared to L-DOPA alone (all p<0.05). WAY100635 successfully reversed the anti-dyskinetic effects of ±8-OH-DPAT on AIMs from the 160th-240th min (all p<0.05).
Experiment 2: Systemic 5-HT1AR stimulation does not alter extracellular striatal glutamate levels but diminishes D1R agonist-induced AIMs in 6-OHDA-lesioned rats
To explore the effects of systemic 5-HT1AR stimulation on striatal glutamate in D1R agonist-treated rats, vehicle or ±8-OH-DPAT (1.0 mg/kg, sc) was given immediately prior to SKF81297 in 6-OHDA- and sham-lesioned rats. Under these conditions, main effects of time were found in 6-OHDA- (F1,16=7.50, p<0.001; Fig 4A) and sham-lesioned (F1,16=13.34, p<0.001; Fig 4D) rats, but there were no main effects of treatment and no treatment × time interactions for either lesion type. Thus, similar to Experiment 1, extracellular striatal glutamate levels fell from baseline over time in both lesion groups. Next, when comparing the average striatal glutamate concentrations during the 120 min of vehicle (time points 0–100) and treatment sampling (time points 120–220), there were no significant main effects of treatment for 6-OHDA- (Fig 4B) or sham-lesioned (Fig 4E) rats. Therefore, neither SKF81297 nor pretreatment with ±8-OH-DPAT impacted extracellular striatal glutamate levels in 6-OHDA- and sham-lesioned animals that had been primed with SKF81297.
Although systemic 5-HT1AR stimulation did not modify striatal glutamate in DA-depleted, SKF81297-treated rats, ±8-OH-DPAT robustly reduced SKF81297-induced AIMs from the 120–240th min (all H1>5.00; all p<0.05; Fig 5C). In sham-lesioned rats, there was no expression of AIMs (Fig 4F).
Figure 5. Effects of striatal 5-HT1AR stimulation on local glutamate and AIMs in 6-OHDA, L-DOPA-treated rats.
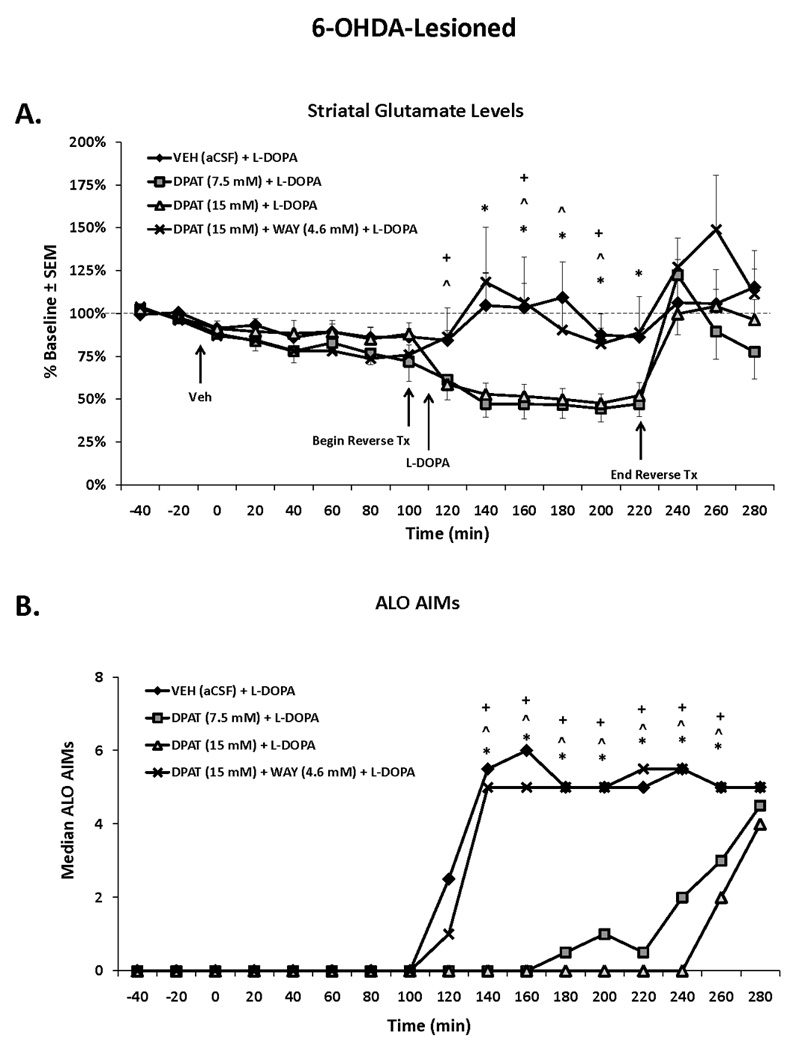
Rats in the third experiment received unilateral 6-OHDA of the MFB and 3 weeks later were primed with L-DOPA (12 mg/kg + benserazide, 15 mg/kg, sc) once daily for 7 d. Dyskinetic rats (n=18) underwent a microdialysis procedure including: 40 min baseline, 120 min vehicle treatment, 120 min drug treatment, and 60 min post-drug treatment sampling (dialysate collected every 20 min and expressed as percent baseline). Intrastriatal drug infusion included: Vehicle (VEH; aCSF), the full 5-HT1AR agonist ±8-OH-DPAT (DPAT; 7.5 or 15 mM), or combined DPAT (15 mM) + WAY100635 (WAY; 4.6 mM), followed 10 min later by systemic treatment injections of L-DOPA (12 mg/kg, + benserazide, 15 mg/kg, sc). AIMs were observed during this time. Lines depict the means ± SEM of striatal glutamate (A) and AIMs (B) for VEH (n=6), DPAT (7.5 mM; n=6), DPAT (15 mM; n=7), and DPAT (15 mM) + WAY (4.6 mM; n=6) groups. Effects over time were determined by a two-way mixed design ANOVA for striatal glutamate. Treatment effects for AIMs (expressed as medians) were analyzed by employing non-parametric Kruskal-Wallis tests. Significant differences between treatments were determined by Mann-Whitney post hoc comparisons for AIMs and planned comparison tests for striatal glutamate.
* p < 0.05 for VEH vs DPAT (7.5 mM)
^ p < 0.05 for VEH vs DPAT (15 mM)
+ p < 0.05 for DPAT (15 mm) vs DPAT (15 mM) + WAY (4.6 mM)
Experiment 3: Striatal 5-HT1AR stimulation reduces local extracellular glutamate levels while concurrently attenuating L-DOPA-induced AIMs in 6-OHDA-lesioned rats
Finally, the effects of striatal 5-HT1AR stimulation on local extracellular glutamate levels in DA-depleted, L-DOPA-treated rats were examined. Using in vivo microdialysis, aCSF or ±8-OH-DPAT (7.5 or 15 mM) was locally administered into the striatum for 2 h. A two-way ANOVA revealed a main effect of time (F16,336=7.08, p<0.001) and a treatment × time interaction (F48,336=2.10, p<0.001) but no main effect of treatment (F3,21=3.39, p=0.067) for extracellular striatal glutamate levels (% baseline) in 6-OHDA-lesioned rats (Fig 5A). Planned comparisons revealed that, similar to systemic ±8-OH-DPAT treatment, striatal infusion of the lower concentration of ±8-OH-DPAT (7.5 mM) significantly reduced local extracellular glutamate levels compared to L-DOPA alone at time points 140–220 (all p<0.05). The higher concentration of ±8-OH-DPAT (15 mM) also diminished striatal glutamate compared to L-DOPA alone at the 120th, 160th, 180th, and 200th min (all p<0.05). WAY100635 reversed ±8-OH-DPAT (15 mM)’s effects at the 120th, 160th, and 200th min (all p<0.05).
For AIMs, main effects were observed from the 140–260th min (all H3>14.00; all p<0.01). As displayed in Figure 5B, post hoc analyses showed that both concentrations of ±8-OH-DPAT significantly attenuated L-DOPA-induced AIMs at all time points (all p<0.05) compared to L-DOPA alone. Importantly, WAY100635 reversed ±8-OH-DPAT’s effects on AIMs at all time points (all p<0.05).
Discussion
The anti-dyskinetic effects of 5-HT1AR stimulation have been well-characterized in both preclinical (Tomiyama et al., 2005; Carta et al. 2007; Dekundy et al., 2007; Dupre et al., 2007, 2008a, 2008b; Eskow et al., 2007; Bishop et al., 2009) and clinical investigations (Bara-Jiminez et al. 2005; Goetz et al. 2007). Unfortunately, how these compounds exert their therapeutic effects is not fully understood, which has limited their use for PD patients. The current study showcases a potential mechanism by which 5-HT1AR stimulation reduces LID by demonstrating that both systemic and intrastriatal pre-treatment with the full 5-HT1AR agonist ±8-OH-DPAT contemporaneously diminishes striatal glutamate efflux and AIMs in L-DOPA-treated, but not D1R agonist-treated, hemiparkinsonian rats.
The loss of DA within the brain has profound, but inconsistent, effects on the corticostriatal glutamate system (Chase & Oh, 2000), such that increases (Lindefors & Ungerstedt, 1990; Meshul et al., 1999; Jonkers et al., 2002; Robinson et al., 2003), reductions (Reid et al., 1990; Meshul et al., 1999; Robinson et al., 2003), and no changes in striatal glutamate levels (Bianchi et al., 2003; Corsi et al., 2003; Robelet et al., 2004) have been reported. In the current study we found that unilateral 6-OHDA-lesions of the MFB, irrespective of the priming drug (L-DOPA or SKF81297), led to a reduction of basal levels of striatal glutamate when compared to sham-lesioned, primed rats (Fig 3). This is similar to recent work by Morgese et al. (2009), who found in unilaterally DA-depleted, chronically L-DOPA-treated rats, basal levels of glutamate within the ipsilateral striatum were diminished compared to the contralateral striatum. Interestingly, Meshul and colleagues (1999) found that there is a time-dependent change in striatal glutamate in non-primed, DA-depleted rats, where an increase is found at 1 month post 6-OHDA-lesion but at 3 months post-lesion, a decrease is found. The early rise in striatal glutamate may be a primary response to the insult of the lesion, while the decrease that we and others report may represent a compensatory response to the initial increase.
In addition to the effects of DA denervation, chronic DA replacement therapy with L-DOPA greatly impacts the corticostriatal pathway (Ouattara et al., 2010). For example, L-DOPA administration (25 and 100 mg/kg) in hemiparkinsonian rats has been shown to enhance striatal glutamate levels (Jonkers et al., 2002; Robelet et al., 2004). Here we show a similar striatal glutamate-enhancing effect (Fig 3A and 3B) with a more clinically-relevant dose of L-DOPA (12 mg/kg) that is paralleled with the induction of dyskinesia (Fig 3C). Although glutamate itself is not sufficient to induce dyskinesia when perfused within the DA-denervated striatum (Buck et al., 2010), it may serve to modulate dyskinetic behaviors as suggested by studies reporting that direct intrastriatal administrations of glutamate antagonists reduce LID (Marin et al., 1996; Nash & Brotchie, 2000).
Similar to L-DOPA, D1R agonists induce dyskinesia (Rascol et al., 2001, 2006; Delfino et al., 2007; Dupre et al., 2007, 2008a) and increase striatal glutamate receptor binding in parkinsonian monkeys (Calon et al., 2002); however, their effects on striatal glutamate release in the DA-depleted brain have not been directly investigated. The present study is the first to report that stimulation with the D1R agonist SKF81297, while inducing dyskinesia (Fig 4C), does not increase extracellular striatal glutamate levels compared to vehicle treatment in DA-depleted rats (Fig 4A and 4B). Thus, our results indicate enhanced extracellular striatal glutamate levels are not directly related to the expression of D1R agonist-mediated dyskinesia. It is important to note that while SKF81297’s timecourse of dyskinesia is shorter (likely attributable to its pharmacokinetic profile), it induced similar severities of axial, forelimb, and orolingual AIMs compared to L-DOPA (Fig 1A; Fig 3C; Fig 4C). Therefore, the differential effects on glutamate between D1R agonism and L-DOPA cannot be explained by differences in AIMs expression. In support of our effects, glutamate antagonism failed to reduce D1R-induced locomotor activity and striatal preprotachykinin mRNA expression in parkinsonian rat pups (Campbell et al., 2006). Taken as a whole, it is possible that stimulation of both DA receptor subtypes is necessary to augment striatal glutamate, as seen in vitro where stimulation of both D1R and D2R were required to enhance glutamate-induced firing of striatal neurons (Hu & White, 1997), in vivo where separate administration of D1R and D2R agonists did not affect striatal glutamate in intact animals (Yamamoto & Davy, 1992), and in vivo with apomorphine in intact rats (Porras & Mora, 1995) and L-DOPA in DA-depleted rats (current study; Robelet et al., 2004). Future work should investigate the effects of D2R agonist treatment on extracellular glutamate levels within the DA-depleted striatum, both alone and in conjunction with D1R agonism, to validate this hypothesis.
Much research has shown 5-HT1AR agonists reduce dyskinesia and most researchers have attributed this to a raphe-mediated mechanism that involves dampening the release of DA delivered to the striatum (Dekundy et al., 2007; Carta et al., 2007; Eskow et al., 2009; Lindgren et al., 2010). However, we have previously shown that local stimulation of striatal 5-HT1AR via microinjections attenuates LID and D1R agonist-induced dyskinesia while improving L-DOPA efficacy and motor performance in hemiparkinsonian rats (Dupre et al., 2008a; Bishop et al., 2009). In the current experiments, we report for the first time that systemic and intrastriatal 5-HT1AR stimulation using ±8-OH-DPAT diminishes L-DOPA-induced striatal glutamate efflux (Figs 3A, 3B, 5A) while concurrently reducing LID (Figs 3C and 5B). The intrastriatal effects lasted the entire time ±8-OH-DPAT was being infused, while the systemic effects lasted for the first 120 min following injection, likely reflecting ±8-OH-DPAT’s short half-life (Yu & Lewander, 1997). Importantly, these results were reversed with the 5-HT1AR antagonist WAY100635 (Fig 3A–C; 5A–B), indicating a specific 5-HT1AR effect. In sham-lesioned rats, pretreatment with ±8-OH-DPAT did not alter striatal glutamate (Fig 3D and 3E), indicating ±8-OH-DPAT’s effects on L-DOPA-related striatal glutamate were dependent on DA-depletion. In support of our findings, striatal 5-HT1AR binding was increased in parkinsonian primates chronically treated with L-DOPA (Huot et al., 2010). This suggests a gain of function for 5-HT1AR in the striatum of DA-depleted, L-DOPA-treated animals and may explain ±8-OHDPAT’s effects in 6-OHDA- and not sham-lesioned animals.
In contrast to L-DOPA-treated, DA-depleted rats, treatment with ±8-OH-DPAT prior to SKF81297, while diminishing dyskinesia (Fig 4C), did not reduce striatal glutamate in D1R agonist-primed, 6-OHDA-lesioned rats (Fig 4A and 4B). Collectively, our results suggest two pathways whereby striatal 5-HT1AR stimulation may diminish dyskinesia. First, 5-HT1AR agonists may act pre-synaptically on glutamatergic corticostriatal neurons to reduce L-DOPA-mediated glutamate efflux. Next, while this mechanism may also relate to LID, D1R agonist-mediated dyskinesia may be modulated via a post-synaptic mechanism(s) that modifies downstream effects. Indeed, 5-HT1AR stimulation may alter D1R-mediated signaling pathways within the striatum, such as those involved in ΔFosB (Cenci et al., 1998; Cao et al., 2010), zif-268 (Carta et al., 2010), and the phosphorylation of extracellular regulated kinase (ERK) 1/2 (Gerfen et al., 2002, 2008; Westin et al., 2007), and subsequent GABA release to output structures, such as the substantia nigra pars reticulata (Weick & Walters, 1986; Radnikow & Misgeld, 1998; Yamamoto et al., 2006; Rangel-Barajas et al., 2008), that are likely involved in dyskinesia caused by D1R agonists. For instance, 5-HT1AR stimulation has been shown to reduce L-DOPA-induced overactive striatal preprodynorphin mRNA that is associated with the D1R pathway (Tomiyama et al., 2005; Bishop et al., 2009). Further studies are underway to test these hypotheses surrounding striatal 5-HT1AR and D1R-mediated dyskinesia.
While 5-HT1AR agonists diminish dyskinesia, these compounds have been shown to produce adverse side effects (Iravani et al., 2006), including flat body posture indicative of 5-HT syndrome (Goodwin et al., 1987; Carey et al., 2004). We used a systemic dose of ±8-OH-DPAT (1.0 mg/kg, sc) that produced transient 5-HT syndrome in the majority of rats treated with L-DOPA; however, no 5-HT syndrome was observed in rats treated in combination with SKF81297. In previous studies using unilaterally DA-depleted rats, this dose has not reduced L-DOPA-efficacy on the forepaw adjusting steps test (Bishop et al., 2009) and has actually increased stepping on this test when given alone or in combination with SKF81297 (Dupre et al., 2008a). The concentrations of ±8-OH-DPAT (7.5 mM and 15 mM) for intrastriatal infusion were determined based on intrastriatal microinjection doses (5 and 10 µg) previously used to produce anti-dyskinetic effects (Dupre et al., 2008a; Bishop et al., 2009). The lower dose of ±8-OH-DPAT variably produced 5-HT syndrome, while the higher dose consistently resulted in symptoms; however, both doses attenuated L-DOPA-induced striatal glutamate efflux and LID to comparable degrees (Fig 5A and B). Thus, while 5-HT syndrome did not appear to be indicative of the reduction in striatal glutamate, it remains an important factor to consider when using 5-HT1AR agonists.
Conclusions
Several important conclusions can be drawn from the current set of experiments. First, L-DOPA treatment produced a moderate augmentation of glutamate in the DA-depleted striatum, whereas SKF81297 did not. These results suggest that enhancement of extracellular striatal glutamate may be important for the expression of LID and not necessarily D1R-mediated dyskinesia. Moreover, stimulation of striatal 5-HT1AR reduces local L-DOPA-induced glutamate efflux while concomitantly diminishing LID. These findings, while providing insight into the striatal workings surrounding dyskinesia in PD, reveal a novel mechanism of 5-HT1AR agonists that may be useful for the treatment of LID and PD.
Acknowledgements
This work was supported by funds from NIH F31NS066684 (K.D.), NIH NS054272 (L.S.), Department of Veterans Affairs MERIT award (WAW) (W.W.), the Center for Development and Behavioral Neuroscience at Binghamton University (C.B.), and NIH NS059600 (C.B.).
Abbreviations
- aCSF
artificial cerebral spinal fluid
- AIMs
Abnormal Involuntary Movements
- Benserazide
DL-Serine 2-(2,3,4-trihydroxybenzyl) hydrazide hydrochloride
- Cc
corpus callosum
- Cpu
caudate putamen
- D1R
D1 receptor
- D2R
D2 receptor
- DA
Dopamine
- DMSO
Dimethyl Sulfoxide
- DOPAC
3,4-dihydroxyphenylacetic acid
- DPAT
±8-OH-DPAT
- EDTA
ethylenediaminetetraacetic acid
- HPLC-ED
high performance liquid chromatography coupled to electrochemical detection
- 5-HT
Serotonin
- 5-HT1AR
Serotonin 1A receptor
- L-DOPA
L-3,4-dihydroxyphenylalanine methyl ester
- LID
L-DOPA-induced dyskinesia
- LV
lateral ventricle
- β-ME
β-mercaptoethanol
- MFB
medial forebrain bundle
- mg
milligram
- NaCl
sodium chloride
- ng
nanogram
- OPA
o-phthaldialdehyde
- ±8-OH-DPAT
(±)-8-Hydroxy-2-(dipropylamino)tetralin hydrobromide
- 6-OHDA
6-hydroxydopamine hydrobromide
- PFA
paraformaldehyde
- PD
Parkinson’s disease
- SKF
- SKF81297
R(+)-SKF-81297 hydrobromide
- VEH
vehicle
- WAY
WAY100635
- WAY100635
N-[2-[4-(2-Methoxyphenyl)-1-piperazinyl]ethyl]-N-2-pyridinylcyclohexanecarboxamide maleate salt
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Antonelli T, Fuxe K, Tomasini MC, Bartoszyk GD, Seyfried CA, Tanganelli S, Ferraro L. Effects of sarizotan on the corticostriatal glutamate pathways. Synapse. 2005;58:193–199. doi: 10.1002/syn.20195. [DOI] [PubMed] [Google Scholar]
- Aubert I, Guigoni C, Hakansson K, et al. Increased D1 dopamine receptor signaling in levodopa-induced dyskinesia. Ann. Neurol. 2005;57:17–26. doi: 10.1002/ana.20296. [DOI] [PubMed] [Google Scholar]
- Bara-Jimenez W, Bibbiani F, Morris MJ, Dimitrova T, Sherzai A, Mouradian MM, Chase TN. Effects of serotonin 5-HT1A agonist in advanced Parkinson’s disease. Mov. Disord. 2005;20:932–936. doi: 10.1002/mds.20370. [DOI] [PubMed] [Google Scholar]
- Bianchi L, Galeffi F, Bolam JP, Della Corte L. The effect of 6-hydroxydopamine lesions on the release of amino acids in the direct and indirect pathways of the basal ganglia: A dual microdialysis probe analysis. Eur. J. Neurosci. 2003;18:856–868. doi: 10.1046/j.1460-9568.2003.02795.x. [DOI] [PubMed] [Google Scholar]
- Bibbiani F, Oh JD, Kielaite A, Collins MA, Smith C, Chase TN. Combined blockade of AMPA and NMDA glutamate receptors reduces levodopa-induced motor complications in animal models of PD. Exp. Neurology. 2005;196:422–429. doi: 10.1016/j.expneurol.2005.08.017. [DOI] [PubMed] [Google Scholar]
- Bishop C, Taylor JL, Kuhn DM, Eskow KL, Park JY, Walker PD. MDMA and fenfluramine reduce L-DOPA-induced dyskinesia via indirect 5-HT1A receptor stimulation. Eur. J. Neurosci. 2006;23:2669–2676. doi: 10.1111/j.1460-9568.2006.04790.x. [DOI] [PubMed] [Google Scholar]
- Bishop C, Krolewski DM, Eskow KL, Barnum CJ, Dupre KB, Deak T, Walker PD. Contribution of the striatum to the effects of 5-HT1A receptor stimulation in L-DOPA-treated hemiparkinsonian rats. J. Neurosci. Res. 2009;87:1645–1658. doi: 10.1002/jnr.21978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buck K, Voehringer P, Ferger B. Site-specific action of L-3,4-dihydroxyphenylalanine in the striatum but not globus pallidus and substantia nigra pars reticulata evokes dyskinetic movements in chronic L-3,4-dihydroxyphenylalanine-treated 6-hydroxydopamine-lesioned rats. Neuroscience. 2010;166:355–358. doi: 10.1016/j.neuroscience.2009.12.032. [DOI] [PubMed] [Google Scholar]
- Calon F, Morissette M, Ghribi O, Goulet M, Grondin R, Blanchet PJ, Bédard PJ, Di Paolo T. Alteration of glutamate receptors in the striatum of dyskinetic 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated monkeys following dopamine agonist treatment. Prog Neuropsychopharmacol. Biol. Psychiatry. 2002;26:127–138. doi: 10.1016/s0278-5846(01)00237-8. [DOI] [PubMed] [Google Scholar]
- Campbell BM, Kreipke CW, Walker PD. Failure of MK-801 to suppress D1 receptor-mediated induction of locomotor activity and striatal preprotachykinin mRNA expression in the dopamine-depleted rat. Neuroscience. 2006;137:505–517. doi: 10.1016/j.neuroscience.2005.09.024. [DOI] [PubMed] [Google Scholar]
- Cao X, Yasuda T, Uthayathas S, Watts RL, Mouradian MM, Mochizuki H, Papa SM. Striatal overexpression of ΔFosB reproduces chronic levodopa-induced involuntary movements. J. Neurosci. 2010;30:7335–7343. doi: 10.1523/JNEUROSCI.0252-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey RJ, DePalma G, Damianopoulos E, Muller CP, Huston JP. The 5-HT1A receptor and behavioral stimulation in the rat: Effects of 8-OHDPAT on spontaneous and cocaine-induced behavior. Psychopharmacology. 2004;177:46–54. doi: 10.1007/s00213-004-1917-4. [DOI] [PubMed] [Google Scholar]
- Carta M, Carlsson T, Kirik D, Bjorklund A. Dopamine released from 5-HT terminals is the cause of L-DOPA-induced dyskinesia in parkinsonian rats. Brain. 2007;130:1819–1833. doi: 10.1093/brain/awm082. [DOI] [PubMed] [Google Scholar]
- Carta AR, Frau L, Pinna A, Morelli M. Dyskinetic potential of dopamine agonists is associated with different striatonigral/striatopallidal zif-268 expression. Exp. Neurology. 2010;224:395–402. doi: 10.1016/j.expneurol.2010.04.016. [DOI] [PubMed] [Google Scholar]
- Cenci MA, Lee CS, Bjorklund A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 1998;10:2694–2706. [PubMed] [Google Scholar]
- Cenci MA. Dopamine dysregulation of movement control in L-DOPA-induced dyskinesia. Trends Neurosci. 2007;30:236–243. doi: 10.1016/j.tins.2007.03.005. [DOI] [PubMed] [Google Scholar]
- Chase TN, Oh JD. Striatal dopamine- and glutamate-mediated dysregulation in experimental parkinsonism. Trends Neurosci. 2000;23:S86–S91. doi: 10.1016/s1471-1931(00)00018-5. [DOI] [PubMed] [Google Scholar]
- Corsi C, Pinna A, Gianfriddo M, Melani A, Morelli M, Pedata F. Adenosine A2A receptor antagonism increases striatal glutamate outflow in dopamine-denervated rats. Eur. J. Pharmacol. 2003;464:33–38. doi: 10.1016/s0014-2999(03)01352-9. [DOI] [PubMed] [Google Scholar]
- Dekundy A, Lundblad M, Danysz W, Cenci MA. Modulation of l-DOPA-induced abnormal involuntary movements by clinically tested compounds: Further validation of the rat dyskinesia model. Beh. Brain Res. 2007;179:76–89. doi: 10.1016/j.bbr.2007.01.013. [DOI] [PubMed] [Google Scholar]
- Delfino M, Kalisch R, Czisch M, et al. Mapping the effects of three dopamine agonists with different dyskinetogenic potential and receptor selectivity using pharmacological functional magnetic resonance imaging. Neuropsychopharmacology. 2007;32:1911–1921. doi: 10.1038/sj.npp.1301329. [DOI] [PubMed] [Google Scholar]
- Donzanti BA, Yamamoto BK. An improved and rapid HPLC-EC method for the isocratic separation of amino acid neurotransmitters from brain tissue and microdialysis perfusates. Life Sci. 1988;43:913–922. doi: 10.1016/0024-3205(88)90267-6. [DOI] [PubMed] [Google Scholar]
- Dupre KB, Eskow KL, Negron G, Bishop C. The differential effects of 5-HT1A receptor stimulation on dopamine receptor-mediated abnormal involuntary movements and rotations in the primed hemiparkinsonian rat. Brain Res. 2007;1158:135–143. doi: 10.1016/j.brainres.2007.05.005. [DOI] [PubMed] [Google Scholar]
- Dupre KB, Eskow KL, Barnum CJ, Bishop C. Striatal 5-HT1A receptor stimulation reduced D1 receptor-induced dyskinesia and improves movement in the hemiparkinsonian rat. Neuropharmacology. 2008a;55:1321–1328. doi: 10.1016/j.neuropharm.2008.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupre KB, Eskow KL, Steiniger A, Klioueva A, Negron G, Lormand L, Park J, Bishop C. Effects of coincident 5-HT1A receptor stimulation and NMDA receptor antagonism on L-DOPA-induced dyskinesia and rotational behaviors in the hemi-parkinsonian rat. Psychopharmacology (Berl) 2008b;199:99–108. doi: 10.1007/s00213-008-1135-6. [DOI] [PubMed] [Google Scholar]
- Eskow KL, Gupta V, Alam S, Park JY, Bishop C. The partial 5-HT1A agonist buspirone reduces the expression and development of l-DOPA-induced dyskinesia in rats and improves l-DOPA efficacy. Pharm. Biochem. Behav. 2007;87:306–314. doi: 10.1016/j.pbb.2007.05.002. [DOI] [PubMed] [Google Scholar]
- Eskow KL, Dupre KB, Barnum CJ, Dickinson SO, Park JY, Bishop C. The role of the dorsal raphe nucleus in the development, expression, and treatment of LID in hemiparkinsonian rats. Synapse. 2009;63:610–620. doi: 10.1002/syn.20630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferré S, Giménez-Llort L, Artigas F, Martínez E. Motor activation in short- and long-term reserpinized mice: role of N-methyl-D-aspartate, dopamine D1 and dopamine D2 receptors. Eur. J. Pharmacol. 1994;255:203–213. doi: 10.1016/0014-2999(94)90099-x. [DOI] [PubMed] [Google Scholar]
- Gerfen CR, Miyachi S, Paletzki R, Brown P. D1 dopamine receptor supersensitivity in the dopamine-depleted striatum results from a switch in the regulation of ERK1/2/MAP kinase. J. Neurosci. 2002;22:5042–5054. doi: 10.1523/JNEUROSCI.22-12-05042.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerfen CR, Paletzki R, Worley P. Differences between dorsal and ventral striatum in drd1a dopamine receptor coupling of dopamine- and cAMP-regulated phosphoprotein-32 to activation of extracellular signal-regulated kinase. J. Neurosci. 2008;28:7113–7120. doi: 10.1523/JNEUROSCI.3952-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goetz CG, Damier P, Hicking C, Laska E, Muller T, Olanow CW, Rascol O, Russ H. Sarizotan as a treatment for dyskinesias in Parkinson’s disease: A double-blind placebo-controlled trial. Mov. Disord. 2007;22:179–186. doi: 10.1002/mds.21226. [DOI] [PubMed] [Google Scholar]
- Goodwin GM, De Souza RJ, Green AR, Heal DJ. The pharmacology of the behavioral and hypothermic responses of rats to 8-hydroxy-2-(di-n-propylamino) tetralin (8-OH-DPAT) Psychopharmacology. 1987;91:506–511. doi: 10.1007/BF00216019. [DOI] [PubMed] [Google Scholar]
- Goodwin P, Starr BS, Starr MS. Motor responses to dopamine D1 and D2 agonists in the reserpine-treated mouse are affected differentially by the NMDA receptor antagonist MK 801. J. Neural Transm. Park. Dis. Dement. Sect. 1992;4:15–26. doi: 10.1007/BF02257618. [DOI] [PubMed] [Google Scholar]
- Guigoni C, Doudnikoff E, Li Q, Bloch B, Bezard E. Altered D1 dopamine receptor trafficking in parkinsonian and dyskinetic non-human primates. Neurobiol. Dis. 2007;26:452–463. doi: 10.1016/j.nbd.2007.02.001. [DOI] [PubMed] [Google Scholar]
- Hu XT, White FJ. Dopamine enhances glutamate-induced excitation of rat striatal neurons by cooperative activation of D1 and D2 class receptors. Neurosci. Lett. 1997;224:61–65. doi: 10.1016/s0304-3940(97)13443-7. [DOI] [PubMed] [Google Scholar]
- Huot P, Johnston TH, Koprich JB, Winkelmolen L, Fox SH, Brotchie JM. Regulation of cortical and striatal 5-HT(1A) receptors in the MPTP-lesioned macaque. Neurobiol. Aging, ahead of print. 2010 doi: 10.1016/j.neurobiolaging.2010.09.011. [DOI] [PubMed] [Google Scholar]
- Iravani MM, Tayarani-Binazir K, Chu WB, Jackson MJ, Jenner P. In 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated primates, the selective 5-hydroxytryptamine 1a agonist(R)-(+)-8-OHDPAT inhibits levodopa-induced dyskinesia but only with increased motor disability. J. Pharmacol. Exp. Ther. 2006;319:1225–1234. doi: 10.1124/jpet.106.110429. [DOI] [PubMed] [Google Scholar]
- Jankovic J. Motor fluctuations and dyskinesias in Parkinson's disease: clinical manifestations. Mov. Disord. 2005;20:S11–S16. doi: 10.1002/mds.20458. [DOI] [PubMed] [Google Scholar]
- Jonkers N, Sarre S, Ebinger G, Michotte Y. MK801 suppresses the L-DOPA-induced increase of glutamate in striatum of hemi-Parkinson rats. Brain Res. 2002;926:149–155. doi: 10.1016/s0006-8993(01)03147-x. [DOI] [PubMed] [Google Scholar]
- Kilpatrick IC, Jones MW, Phillipson OT. A semiautomated analysis method for catecholamines, indoleamines, and some prominent metabolites in microdissected regions of the nervous system: An isocratic HPLC technique employing coulometric detection and minimal sample preparation. J. Neurochem. 1986;46:1865–1876. doi: 10.1111/j.1471-4159.1986.tb08506.x. [DOI] [PubMed] [Google Scholar]
- Kobylecki C, Cenci MA, Crossman AR, Ravenscroft P. Calcium-permeable AMPA receptors are involved in the induction and expression of l-DOPA-induced dyskinesia in Parkinson's disease. J. Neurochem. 2010;114:499–511. doi: 10.1111/j.1471-4159.2010.06776.x. [DOI] [PubMed] [Google Scholar]
- Lindefors N, Ungerstedt U. Bilateral regulation of glutamate tissue and extracellular levels in caudate-putamen by midbrain dopamine neurons. Neurosci. Lett. 1990;115:248–252. doi: 10.1016/0304-3940(90)90463-j. [DOI] [PubMed] [Google Scholar]
- Lindgren HS, Andersson DR, Lagerkvist S, Nissbrandt H, Cenci MA. L-DOPA-induced dopamine efflux in the striatum and the substantia nigra in a rat model of Parkinson's disease: temporal and quantitative relationship to the expression of dyskinesia. J. Neurochem. 2010;112:1465–1476. doi: 10.1111/j.1471-4159.2009.06556.x. [DOI] [PubMed] [Google Scholar]
- Lundblad M, Anderson M, Winkler C, Kirik D, Wierup N, Cenci MA. Pharmacological validation of behavioural measures of akinesia and dyskinesia in a rat model of Parkinson’s disease. Eur. J. Neurosci. 2002;15:120–132. doi: 10.1046/j.0953-816x.2001.01843.x. [DOI] [PubMed] [Google Scholar]
- Marin C, Papa S, Engber TM, Bonastre M, Tolosa E, Chase TN. MK-801 prevents levodopa-induced motor response alterations in parkinsonian rats. Brain Res. 1996;736:202–205. doi: 10.1016/0006-8993(96)00693-2. [DOI] [PubMed] [Google Scholar]
- Matsubara K, Shimizu K, Suno M, et al. Tandospirone, a 5-HT1A agonist, ameliorates movement disorder via non-dopaminergic systems in rats with unilateral 6-hydroxydopamine-generated lesions. Brain Res. 2006;1112:126–133. doi: 10.1016/j.brainres.2006.07.003. [DOI] [PubMed] [Google Scholar]
- Meshul CK, Emre N, Nakamura CM, Allen C, Donohue MK, Buckman JF. Time-dependent changes in striatal glutamate synapses following a 6-hydroxydopamine lesion. Neuroscience. 1999;88:1–16. doi: 10.1016/s0306-4522(98)00189-4. [DOI] [PubMed] [Google Scholar]
- Mignon L, Wolf WA. Postsynaptic 5-HT(1A) receptors mediate an increase in locomotor activity in the monoamine-depleted rat. Psychopharmacology (Berl) 2002;163:85–94. doi: 10.1007/s00213-002-1121-3. [DOI] [PubMed] [Google Scholar]
- Mignon LJ, Wolf WA. 8-Hydroxy-2-(di-n-propylamino)tetralin reduces striatal glutamate in an animal model of Parkinson’s disease. Neuroreport. 2005;16:699–703. doi: 10.1097/00001756-200505120-00009. [DOI] [PubMed] [Google Scholar]
- Morgese MG, Cassano T, Gaetani S, et al. Neurochemical changes in the striatum of dyskinetic rats after administration of the cannabinoid agonist WIN55,212-2. Neurochem. Int. 2009;54:56–64. doi: 10.1016/j.neuint.2008.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nash JE, Brotchie JM. A common signaling pathway for striatal NMDA and adenosine A2a receptors: Implications for the treatment of Parkinson’s disease. J. Neurosci. 2000;20:7782–7789. doi: 10.1523/JNEUROSCI.20-20-07782.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ouattara B, Hoyer D, Grégoire L, Morissette M, Gasparini F, Gomez-Mancilla B, Di Paolo T. Changes of AMPA receptors in MPTP monkeys with levodopa-induced dyskinesias. Neuroscience. 2010;167:1160–1167. doi: 10.1016/j.neuroscience.2010.03.022. [DOI] [PubMed] [Google Scholar]
- Pavese N, Evans AH, Tai YF, et al. Clinical correlates of levodopa-induced dopamine release in Parkinson disease. Neurology. 2006;67:1612–1617. doi: 10.1212/01.wnl.0000242888.30755.5d. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson W. The Rat Brain in Stereotaxic Coordinates. fourth ed. San Diego: Academic Press; 1998. [Google Scholar]
- Pett MA. Nonparametric statistics for health care research: Statistics for small samples and unusual distributions. Thousand Oaks, California: SAGE Publications, Inc.; 1997. [Google Scholar]
- Pollack AE, Yates TM. Prior dopamine D1 receptor stimulation is required to prime D2-mediated striatal fos expression in 6-hydroxydopamine-lesioned rats. Neuroscience. 1999;94:505–514. doi: 10.1016/s0306-4522(99)00338-3. [DOI] [PubMed] [Google Scholar]
- Porras A, Mora F. Dopamine-glutamate-GABA interactions and ageing: Studies in the striatum of the conscious rat. Eur. J. Neurosci. 1995;7:2183–2188. doi: 10.1111/j.1460-9568.1995.tb00640.x. [DOI] [PubMed] [Google Scholar]
- Rangel-Barajas C, Silva I, Garcıa-Ramırez M, Sanchez-Lemus E, Floran L, Aceves J, Erlij D, Floran B. 6-OHDA-induced hemiparkinsonism and chronic L-DOPA treatment increase dopamine D1-stimulated [3H]-GABA release and [3H]-cAMP production in substantia nigra pars reticulata of the rat. Neuropharmacology. 2008;55:704–711. doi: 10.1016/j.neuropharm.2008.06.002. [DOI] [PubMed] [Google Scholar]
- Radnikow G, Misgeld U. Dopamine D1 receptors facilitate GABAA synaptic currents in the rat substantia nigra pars reticulata. J. Neurosci. 1998;18:2009–2016. doi: 10.1523/JNEUROSCI.18-06-02009.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rascol O, Nutt JG, Blin O, et al. Induction by dopamine D1 receptor agonist ABT-431 of dyskinesia similar to levodopa in patients with Parkinson disease. Arch. Neurol. 2001;58:249–254. doi: 10.1001/archneur.58.2.249. [DOI] [PubMed] [Google Scholar]
- Rascol O, Brooks DJ, Korczyn AD, De Deyn PP, Clarke CE, Lang AE, Abdalla M. Development of dyskinesias in a 5-year trial of ropinirole and L-dopa. Mov. Disord. 2006;21:1844–1850. doi: 10.1002/mds.20988. [DOI] [PubMed] [Google Scholar]
- Reid MS, Herrera-Marschitz M, Kehr J, Ungerstedt U. Striatal dopamine and glutamate release: Effects of intranigral injections of substance P. Acta Physiol. Scand. 1990;140:527–537. doi: 10.1111/j.1748-1716.1990.tb09030.x. [DOI] [PubMed] [Google Scholar]
- Robelet S, Melon C, Guillet B, Salin P, Kerkerian-LeGoff L. Chronic l-DOPA treatment increases extracellular glutamate levels and GLT1 expression in the basal ganglia in a rat model of Parkinson’s disease. Eur. J. Neurosci. 2004;20:1255–1266. doi: 10.1111/j.1460-9568.2004.03591.x. [DOI] [PubMed] [Google Scholar]
- Robinson S, Freeman P, Moore C, Touchon JC, Krentz L, Meshul CK. Acute and subchronic MPTP administration differentially affects striatal glutamate synaptic function. Exp. Neurology. 2003;180:74–87. doi: 10.1016/s0014-4886(02)00050-x. [DOI] [PubMed] [Google Scholar]
- Rowley HL, Martin KF, Marsden CA. Determination of in vivo amino acid neurotransmitters by high-performance liquid chromatography with o-phthalaldehyde-sulphite derivatisation. J. Neurosci. Meth. 1995;57:93–99. doi: 10.1016/0165-0270(94)00132-z. [DOI] [PubMed] [Google Scholar]
- Rylander D, Iderberg H, Li Q, et al. A mGluR5 antagonist under clinical development improves L-DOPA-induced dyskinesia in parkinsonian rats and monkeys. Neurobiol. Dis. 2010;39:352–361. doi: 10.1016/j.nbd.2010.05.001. [DOI] [PubMed] [Google Scholar]
- Taylor JL, Bishop C, Ullrich T, Rice KC, Walker PD. Serotonin 2A receptor antagonist treatment reduces dopamine D1 receptor-mediated rotational behavior but not L-DOPA-induced abnormal involuntary movements in the unilateral dopamine-depleted rat. Neuropharmacology. 2006;50:761–768. doi: 10.1016/j.neuropharm.2005.12.004. [DOI] [PubMed] [Google Scholar]
- Tomiyama M, Kimura T, Maeda T, Kannari K, Matsunaga M, Baba M. A serotonin 5-HT1A receptor agonist prevents behavioral sensitization to L-DOPA in a rodent model of Parkinson’s disease. Neurosci. Res. 2005;52:185–194. doi: 10.1016/j.neures.2005.03.004. [DOI] [PubMed] [Google Scholar]
- Weick BG, Walters JR. Effects of D1 and D2 dopamine receptor stimulation on the activity of substantia nigra pars reticulata neurons in 6-hydroxydopamine lesioned rats: D1/D2 coactivation induces potentiated responses. Brain Res. 1986;405:243–246. doi: 10.1016/0006-8993(87)90293-9. [DOI] [PubMed] [Google Scholar]
- Westin JE, Vercammen L, Strome EM, Konradi C, Cenci MA. Spatiotemporal pattern of striatal ERK1/2 phosphorylation in a rat model of l-DOPA-induced dyskinesia and the role of dopamine D1 receptors. Biol. Psychiatry. 2007;62:800–810. doi: 10.1016/j.biopsych.2006.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto BK, Davy S. Dopaminergic modulation of glutamate release in striatum as measured by microdialysis. J. Neurochem. 1992;58:1736–1742. doi: 10.1111/j.1471-4159.1992.tb10048.x. [DOI] [PubMed] [Google Scholar]
- Yamamoto N, Pierce RC, Soghomonian JJ. Subchronic administration of L-DOPA to adult rats with a unilateral 6-hydroxydopamine lesion of dopamine neurons results in a sensitization of enhanced GABA release in the substantia nigra, pars reticulata. Brain Res. 2006;1123:196–200. doi: 10.1016/j.brainres.2006.09.027. [DOI] [PubMed] [Google Scholar]
- Yu H, Lewander T. Pharmacokinetic and pharmacodynamic studies of (R)-8-hydroxy-2-(di-n-propylamino)tetralin in the rat. Eur. Neuropsychopharmacol. 1997;7:165–172. doi: 10.1016/s0924-977x(96)00395-1. [DOI] [PubMed] [Google Scholar]